PRODUKTIVITAS PRIMER FITOPLANKTON DAN
KETERKAITANNYA DENGAN UNSUR HARA DAN CAHAYA
DI PERAIRAN TELUK BANTEN
OLEH :
A L I A N T O
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ABSTRAK
ALIANTO. Produktivitas Primer Fitoplankton dan Keterkaitannya Dengan Unsur
Hara dan Cahaya Di Perairan Teluk Banten. Di bawah Bimbingan Enan M. Adiwilaga
sebagai ketua komisi, dan Ario Damar sebagai anggota komisi.
Penelitian ini bertujuan untuk mengetahui hubungan antara produktivitas
primer fitoplankton dengan intensitas cahaya dan unsur hara di perairan Teluk Banten.
Metode pengukuran produktivitas primer fitoplankton dengan menggunakan
metode oksigen botol terang botol gelap dengan waktu inkubasi selama 5 jam.
Hasil penelitian didapatkan 36 genera fitoplankton yang terdiri dari 29 genera
Bacillariophyceae, 6 genera Dinophyceae, dan 1 genera Cyanophyceae. Kisaran
rata-rata konsentrasi unsur hara DIN (ammonia-nitrogen, nitrat-nitrogen,
nitrit-nitrogen), DIP (ortofosfat) dan silikat yang didapatkan selama pengamatan
berturut-turut sebesar 0.467–0.610 mg at N/4 I, 0.006–0.043 mg at P/4 I di stasiun A
dan 0.430–0.746 mg at N/4 I, 0.006–0.032 mg at P/4 I di stasiun B, serta 1.019–6.294
mg at Si/4 I di stasiun A dan 0.581–3.378 mg at Si/4 I di stasiun B. Nilai kisaran
rata-rata produktivitas primer fitoplankton menurut kedalaman inkubasi di stasiun A
sebesar 14.15–29.59 mg C/m3/5 jam (33.96–71.01 mg C/m3
/hari), dan di stasiun B
sebesar 13.56–25.68 mg C/m3/5 jam (32.54–61.63 mg C/m3
/hari) dengan intensitas
cahaya optimum berturut-turut sebesar 32.6% dan 48.2% menurut kedalaman
inkubasi.
Hasil analisis menunjukkan terdapat hubungan yang sangat erat antara cahaya
dengan peroduktivitas primer fitoplankton, dengan koefisien determinasi (R2) yang
diperoleh sebesar 82% di stasiun A dan 64% di stasiun B. Hasil analisis menunjukkan
pula terdapat hubungan yang kurang erat antara unsur hara DIN dan DIP serta silikat
dengan produktivitas primer fitoplankton, dengan kisaran nilai koefisien determinasi
SURAT PERNYATAAN
Dengan ini saya menyatakan bahwa tesis yang berjudul :
PRODUKTIVITAS PRIMER FITOPLANKTON DAN KETERKAITANNYA DENGAN UNSUR HARA DAN CAHAYA DI PERAIRAN TELUK BANTEN
adalah benar merupakan hasil karya saya sendiri dan belum pernah dipublikasikan.
Semua sumber data dan informasi yang digunakan telah dinyatakan secara jelas dan
dapat diperiksa kebenarannya.
Bogor, Januari 2006
PRODUKTIVITAS PRIMER FITOPLANKTON DAN
KETERKAITANNYA DENGAN UNSUR HARA DAN CAHAYA
DI PERAIRAN TELUK BANTEN
A L I A N T O
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Ilmu Perairan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Produktivitas Primer Fitoplankton dan Keterkaitannya Dengan Unsur Hara dan Cahaya Di Perairan Teluk Banten
N a m a : A l i a n t o
NRP : C151030061
Disetujui
Komisi Pembimbing
Dr. Ir. Enan M. Adiwilaga Dr. Ir. Ario Damar, M.Si Ketua Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana Ilmu Perairan
Dr. Chairul Muluk, M.Sc Prof. Dr. Ir. Syafrida Manuwoto, M.Sc
RIWAYAT HIDUP
Penulis dilahirkan di Wanci Kecamatan Wangi-Wangi Kabupaten Buton
(sekarang Kabupaten Wakatobi) Propinsi Sulawesi Tenggara pada tanggal 05 Maret
1970 dari ayah La Hamuna dan ibu Hj. Wa Nuru. Penulis merupakan anak ketiga dari
tujuh bersaudara.
Pada tahun 1989 penulis lulus dari SMA Negeri Wanci dan pada tahun 1990
penulis lulus seleksi masuk perguruan tinggi melalui jalur UMPTN pada Program
Studi dan Jurusan Manajemen Sumberdaya Perairan Fakultas Perikanan Universitas
Pattimura Ambon. Penulis menyelesaikan studi di Universitas Pattimura tahun 1997.
Pada tahun 2001 penulis diterima sebagai staf pengajar pada Program Studi
Diploma Tiga Budidaya Perikanan Fakultas Pertanian Universitas Cenderawasih
sekarang Program Studi Manajemen Sumberdaya Perairan Jurusan Perikanan Fakultas
Peternakan Perikanan dan Ilmu Kelautan Universitas Negeri Papua Manokwari, Irian
Jaya Barat.
Pada tahun 2003 penulis mendapat kesempatan untuk melanjutkan studi
Magister Sains pada Program Studi Ilmu Perairan, Sekolah Pascasarjana Institut
Pertanian Bogor dengan sponsor Beasiswa Program Pascasarjana (BPPS) dari
PRAKATA
Puji syukur penulis panjatkan kehadirat Allah SWT karena atas berkat rahmat
dan karunia-Nya jualah sehingga penulisan karya ilmiah ini dapat diselesaikan. Judul
penelitian yang dilaksanakan sejak bulan Maret 2005–April 2005 ini adalah
“Produktivitas Primer Fitoplankton dan Keterkaitannya Dengan Unsur Hara dan
Cahaya Di Perairan Teluk Banten”.
Pada kesempatan ini penulis mengucapkan banyak terima kasih kepada Bapak
Dr. Ir. Enan M. Adiwilaga dan Dr. Ir. Ario Damar, M.Si, selaku ketua dan anggota
komisi pembimbing, yang telah banyak memberikan saran dan mengarahkan penulis
sejak penulisan proposal, pelaksanaan penelitian sampai dengan penulisan karya
ilmiah ini. Kepada Bapak Yusli Wardiatno yang telah banyak memberi masukan demi
perbaikan tulisan ini sebagai penguji luar komisi. Bapak Dr. Chairul Muluk, M.Sc,
selaku Ketua Program Studi Ilmu Perairan beserta seluruh staf pengajar yang telah
banyak membekali ilmu pengetahu an kepada penulis.
Ucapan terima kasih penulis sampaikan pula kepada Direktur Jenderal
Pendidikan Tinggi DEPDIKNAS atas bantuan beasiswa melalui BPPS. Rektor
Universitas Negeri Papua dan Dekan Fakultas Peternakan Perikanan dan Ilmu
Kelautan atas izin yang diberikan untuk melanjutkan pendidikan pascasarjana.
Ucapan terima kasih disampaikan kepada ayah, ibu serta istri dan anakku tersayang
serta seluruh keluarga atas segala doa yang telah diberikan. Kepada pihak yang telah
banyak membantu pada saat penelitian diucapkan terima kasih yang tak terhingga.
Harapan penulis semoga karya ilmiah ini dapat memberikan manfaat.
Bogor, Januari 2006
DAFTAR ISI
Fosfor Inorganik Terlarut (DIP) ... 9
Silikat ... 10
Intensitas Cahaya Matahari ... 20
Produktivitas Primer ... 20
Analisis Unsur Hara ... 21
Identifikas i Fitoplankton ... 22
Klorofil-a ... 24
Intensitas Cahaya Matahari ... 29
Intensitas Cahaya Matahari Permukaan ... 29
Intensitas Cahaya Matahari di Kolom Air ... 30
Nitrogen Inorganik Terlarut (DIN) ... 33
Ammonia-Nitrogen (NH3-N) ... 34
Nitrit-Nitrogen (NO2-N) ... 36
Fosfor Inorganik Terlarut (DIP) ... 36
Ortofosfat (PO4-P) ... 36
Silikat (Si) ... 37
Struktur Komunitas Fitoplankton ... 38
Komposisi dan Kelimpahan Fitoplankton ... 38
Indeks Biologi Fitoplankton ... 41
Klorofil-a ... 43
Produktifitas Primer Perairan ... 45
Produktivitas Primer Pada Berbagai Kedalaman Inkubasi ... 45
Produktivitas Primer Pada Berbagai Lapisan Kolom Air ... 47
Hubungan Cahaya Dengan Produktivitas Primer ... 48
Hubungan Unsur Hara Dengan Produktivitas Primer ... 50
Hubungan Klorofil-a Dengan Produktivitas Primer ... 54
KESIMPULAN DAN SARAN Kesimpulan ... 55
Saran ... 55
DAFTAR PUSTAKA ... 56
DAFTAR TABEL
Halaman
1 Nilai Produktifitas Primer Dengan Metode Oksigen (O2) di Beberapa
Wilayah Tropik dan Temperate ... 7
2 Parameter Fisika, Kimia dan Biologi Yang Di ukur Serta Metode dan Alat Ukur yang Digunakan ... 19
3 Kondisi Oseanografi Perairan Teluk Banten ... 27
4 Rataan Unsur Hara Menurut Stasiun Pengamatan di Perairan
Teluk Banten ... 33
5 Rataan Konsentrasi Unsur Hara Menurut Kedalaman Inkubasi
di Perairan Teluk Banten ... 34
6 Rataan Jumlah Genera Fitoplankton Menurut Stasiun Pengamatan
di Perairan Teluk Banten ... 39
7 Rataan Kelimpahan Sel Fitoplankton Menurut Stasiun Pengamatan
di Perairan Teluk Banten ... 40
8 Indeks Bio logi Menurut Stasiun Pengamatan di Perairan
Teluk Banten ... 42
9 Indeks Biologi Menurut Kedalaman Inkubasi di Perairan
Teluk Banten ... 42
10 Konsentrasi Klorofil-a Menurut Stasiun Pengamatan
di Perairan Teluk Banten ... 44
11 Konsentrasi Klorofil-a Menurut Kedalaman Inkubasi di Perairan
Teluk Banten ... 44
12 Nilai Produktivitas Primer Bersih Pada Kedalaman Inkubasi di Perairan Teluk Banten ... 46
DAFTAR GAMBAR
Halaman
1 Kerangka Proses Perumusan dan Pendekatan Masalah ... 4
2 Siklus Nitrogen di Zona Eufotik ... 8
3 Penyerapan dari Tiga Sumber Nitrogen (NH3, Urea, NO3) Oleh Dua Ukuran Fitoplankton Yang Berbeda ... 9
4 Grafik Distribusi Vertikal Cahaya dan Fotosintesis di Perairan ... 14
5 Lokasi Penelitian Perairan Teluk Banten ... 18
6 Intensitas Cahaya Permukaan ... 29
7 Intensitas Cahaya di Lapisan Permukaan ... 31
8 Distribusi Cahaya di Kolom Air Perairan Teluk Banten ... 32
9 Rataan Jumlah Genera Fitoplankton Menurut Kedalaman Inkubasi di Perairan Teluk Banten ... 39
10 Rataan Kelimpahan Fitoplankton Menurut Kedalaman Inkubasi di Perairan Teluk Banten ... 41
11 Nilai Produktivitas Primer Bersih Pada Berbagai Kedalaman Inkubasi di Perairan Teluk Banten ... 47
12 Pola Hubungan Intensitas Cahaya Matahari Dengan Produktivitas Primer Bersih di Perairan Teluk Banten ... 49
13 Pola Hubungan Intensitas Cahaya Matahari Dengan Produktivitas Primer Bersih Pada Berbagai Lapisan Kolom Air di Perairan Teluk Banten ... 50
14 Grafik Pola Hubungan Unsur Hara Dengan Produktivitas Primer Bersih di Stasiun A ... 52
15 Grafik Pola Hubungan Unsur Hara Dengan Produktivitas Primer Bersih di Stasiun B ... 53
DAFTAR LAMPIRAN
Halaman
1 Prosedur Pengisian Air Kedalam Botol BOD ... 63
2 Prosedur Pengukuran Oksigen Terlarut Dengan Titrasi Winkler ... 63
3 Prosedur Pengukuran Produktivitas Primer ... 64
4 Tahapan Analisis Unsur Hara ... 64
5 Prosedur Analisis Klorofil-a ... 66
6 Intensitas Cahaya Matahari Permukaan di Perairan Teluk Banten ... 67
7 Intensitas Cahaya Matahari Pada Lapisan Permukaan di Perairan Teluk Banten ... 67
8 Intensitas Cahaya Matahari Pada Berbagai Kedalaman Inkubasi di Perairan Teluk Banten ... 68
9 Konsentrasi Parameter Utama Yang Diukur di Perairan Teluk Banten ... 70
10 Hasil Uji Sidik Ragam (Anova) Produktivitas Primer Bersih Terhadap Stasiun dan Kedalaman di Perairan Teluk Banten ... 72
11 Kelimpahan Fitoplankton (sel/I) Pada Periode I di Perairan Teluk Banten ... 73
12 Kelimpahan Fitoplank ton (sel/I) Pada Periode II di Perairan Teluk Banten ... 74
13 Kelimpahan Fitoplankton (sel/I) Pada Periode III di Perairan Teluk Banten ... 75
14 Nilai Produktivitas Primer Bersih Pada Setiap Lapisan Kolom Air Stasiun A Selama Waktu Inkubasi di Perairan Teluk Banten... 76
PENDAHULUAN
Latar Belakang
Kemampuan potensial suatu perairan untuk menghasilkan sumberdaya alam
hayati ditentukan oleh produktivitas primer fitoplanktonnya. Produktivitas primer
fitoplankton memainkan peran penting dalam proses bahan pada jaring makanan
planktonik, yang dihasilkan sebagai senyawa organik yang menjadi sumber carbon
dan energi terpenting bagi organisme di berbagai lingkungan perairan (Lignell 1992).
Fitoplankton merupakan tumbuhan yang paling luas tersebar dan ditemukan di
seluruh permukaan laut dan pada kedalaman sampai setebal lapisan eufotik.
Fitoplankton men ghasilkan karbon 1010 ton setiap tahun atau kira-kira 50% dari
seluruh karbon yang dihasilkan oleh seluruh tumbuh-tumbuhan (Smayda 1970;
Meadows dan Campbell 1988; Brotowidjoyo et al. 1995) dan diperkirakan 50% produktivitas primer di laut dihasilkan oleh fitoplankton (Rost et al. 2003).
Fitoplankton merupakan salah satu tumbuhan laut yang mengandung klorofil-a
sehingga sangat memungkinkan untuk melakukan fotosintesis. Pertumbuhan dan
perkembangan fitoplankton sangat tergantung pada keberadaan cahaya matahari dan
suplai bahan inorganik terutama nitrogen, fosfor dan silikat di perairan (Tett dan
Edwars 1984; Duarte 1992).
Sumber energi yang utama bagi kehidupan fitoplankton di laut berasal dari
cahaya matahari. Cahaya merupakan salah satu faktor utama yang mengontrol laju
fotosintesis di laut. Di samping itu, proses fotosintesis fitoplankton berjalan dengan
memanfaatkan unsur hara yang ada di lingkungannya. Unsur hara yang dibutuhkan
oleh fitoplankton untuk pertumbuhan dan perkembangannya terutama nitrogen, dan
biogeokimia seperti produksi dan dekomposisi bahan organik biogenik dan laju
penenggelaman bahan partikulat (Smith 1984; Hirose dan Kamiya 2003).
Nitrogen di laut terdiri dari ammonia, nitrat, dan nitrit. Nitrogen sendiri
merupakan unsur kimia yang penting dalam proses pembentukan protoplasma.
Bentuk-bentuk senyawa nitrogen tersebut berperan penting sebagai sumber nitrogen
bagi fitoplankton meskipun peranan masing-masing ion tersebut tidak selalu sama
terhadap berbagai jenis fitoplankton.
Fosfor merupakan unsur esensial penting bagi fitoplankton dan ketersediannya
di perairan selalu lebih rendah dari nitrogen serta keberadaannya di zona eufotik
selalu ditemukan dalam konsentrasi yang rendah (Dawes 1981). Fosfor dibutuhkan
oleh fitoplankton sebagai sumber energi terutama untuk memproduksi fosfolipid dan
ikatan fosfat gula dan molekul seperti adenosin triphosphat (ATP).
Keberadaan unsur hara nitrogen dan fosfor di perairan, mempunyai dua
pengaruh terhadap fitoplankton, yaitu (1) unsur hara dalam jumlah yang cukup sangat
diperlukan oleh fitoplankton dalam proses fotosintesis untuk metabolisme sel
hidupnya. (2) kandungan unsur hara dalam jumlah yang tinggi dapat menyebabkan
gangguan yang berlanjut pada lingkungan perairan seperti penurunan penetrasi cahaya
matahari, dan ledakan populasi alga yang beracun (Rabalais 1999).
Sehubungan dengan uraian tersebut di atas, masih sangat diperlukan berbagai
informasi mengenai perairan Teluk Banten. Maka penelitian-penelitian khususnya
mengenai kondisi lingkungan yang berhubungan dengan cahaya, unsur hara dan
produktivitas primer perairan sangat penting karena informasi mengenai kedua hal
tersebut merupakan suatu ukuran terhadap kualitas dan kemampuan perairan Teluk
Perumusan dan Pendekatan Masalah
Kelimpahan dan potensi tumbuh fitoplankton di perairan Teluk Banten diduga
tidak merata dengan ketersediaan unsur hara yang berada di dalam maupun masukan
unsur hara dari luar perairan. Ketersediaan unsur hara di perairan sangat menentukan
struktur komunitas fitoplankton. Disamping itu, struktur komunitas fitoplankton di
perairan sangat ditentukan pula oleh intensitas cahaya yang ada dalam kolom air.
Intensitas cahaya yang ada di kolom air umumnya tidak serasi dengan kebutuhan
fitoplankton, sehingga pada suatu perairan akan didominasi oleh jenis -jenis
fitoplankton tertentu.
Sumber penyebab tidak serasinya hal tersebut, terutama disebabkan karena
adanya kekeruhan yang disebabkan oleh partikel-partikel tersuspensi yang akan
menjadi faktor pembatas terutama bagi distribusi horisontal dan vertikal fitoplankton,
sehingga di setiap kolom air perkembangan potensi tumbuh fitoplankton akan
berbeda-beda. Hal ini, akan berpengaruh pada nilai kandungan produktivitas primer
fitoplankton yang berbeda-beda pula.
Berdasarkan hal tersebut diatas, maka diperlukan suatu kajian untuk
mengetahui seberapa jauh produktivitas primer fitoplankton dibatasi oleh intensitas
cahaya dan ketersediaan unsur hara di perairan Teluk Banten. Rumusan dan
pendekatan masalah dari penelitian ini secara sederhana disajikan dalam bentuk
Gambar 1. Kerangka proses perumusan dan pendekatan masalah.
Keterangan : Tanda ( ) adalah jalur penelitian utama. Tanda (- - - -) adalah jalur penelitian penunjang.
Tujuan
Penelitian ini bertujuan untuk mengetahui hubungan antara produktivitas
primer fitoplankton dengan intensitas cahaya matahari dan unsur hara di perairan
Teluk Banten.
Hipotesis
Jika intensitas cahaya matahari yang mencapai kolom air optimum serta
ditunjang oleh konsentrasi unsur hara yang tinggi sampai pada batas tertentu yang
masih sesuai dengan kebutuhan fitoplankton maka tingkat produktivitas primer
fitoplankton akan maksimal. Unsur hara
Struktur
Komunitas Produktivitas Primer
Fitoplankton Kelimpahan
TINJAUAN PUSTAKA
Produktivitas Primer
Pada umumnya produktivitas primer dianggap sebagai padanan fotosintesis,
walaupun sejumlah kecil produktivitas primer dapat dihasilkan oleh bakteri
kemosintetik (Nybakken 1988). Terdapat hubungan yang positif antara kelimpahan
fitoplankton dengan produktivitas primer, jika kelimpahan fitoplankton di suatu
perairan tinggi, maka perairan tersebut cenderung mempunyai produktivitas yang
tinggi pula (Raymont 1963).
Odum (1971) mendefinisikan produktivitas primer sebagai derajat
penyimpanan energi matahari dalam bentuk bahan organik, sebagai hasil fotosintesis
dan kemosintesis dari produsen primer. Produktivitas primer diistilahkan sebagai laju
fiksasi karbon (sintesis organik) di dalam perairan dan biasanya diekspresikan sebagai
gram karbon yang diproduksi per satuan waktu (Kennish 1990). Hal yang sama
dikemukakan oleh Levinton (1982) dan Barnabe dan Barnabe (2000), bahwa
produktivitas adalah jumlah yang dihasilkan oleh organisme hidup per satuan waktu
dan sering diestimasi sebagai jumlah karbon yang terdapat di dalam material hidup
dan secara umum dapat dinyatakan sebagai gram karbon yang dihasilkan dalam
satuan meter kuadrat kolom air per hari (g C/m2/hari) atau sebagai gram karbon yang
dihasilkan dalam satu meter kubik per hari (g C/m3/hari).
Produktivitas primer merupakan sumber utama energi bagi proses metabolik
yang terjadi dalam perairan. Pada ekosistem perairan sebagian besar produktivitas
primer dihasilkan oleh fitoplankton (Kennish 1990; Barnabe dan Barnabe 2000).
Aliran energi dalam ekosistem perairan dimulai dengan fiksasi energi oleh
mengakumulasi energi, energi yang diakumulasi oleh fitoplankton inilah yang disebut
produksi atau secara lebih spesifik disebut produksi primer (Sumich 1994).
Produktivitas dibedakan atas dua, yaitu produktivitas primer kotor (Gross
Primary Production) dan produktivitas primer bersih (Net Primary Production).
Produktivitas primer kotor adalah laju produksi primer zat organik secara
keseluruhan, termasuk yang digunakan untuk respirasi, sedangkan produktivitas
primer bersih adalah laju produktivitas primer zat organik setelah dikurangi dengan
yang digunakan untuk respirasi (Nybakken 1988).
Pengukuran produktivitas primer fitoplankton merupakan satu syarat dasar
untuk mempelajari struktur dan fungsi ekosistem perairan. Metode yang digunakan
untuk pengukuran produktivitas primer fitoplankton pertama kali menggunakan
metode O2 yang diperkenalkan oleh Garder dan Gran serta metode 14C oleh Steemann
Nielsen dengan menggunakan tiga tipe metode inkubasi, yaitu inkubasi in situ, simulasi in situ dan metode cahaya (Gocke dan Lenz 2004).
Inkubasi in situ pada umumnya lebih mendekati kondisi alam dan dianggap sebagai metode yang lebih dipercaya (Gocke dan Lenz 2004). Nilai produktivitas
primer fitoplankton dengan menggunakan metode O2 tipe inkubasi in situ telah banyak dilakukan pada beberapa perairan dunia dan memperlihatkan nilai yang
bervariasi (Tabel 1). Produktivitas primer fitoplankton dalam suatu perairan
dipengaruhi oleh faktor fisika, kimiawi dan biologi. Faktor-faktor tersebut meliputi
cahaya, suhu, sirkulasi massa air, unsur hara dan grazing oleh zooplankton
Tabel 1. Nilai produktivitas primer dengan metode oksigen (O2) di beberapa wilayah tropik dan temperate
Catatan : # g C/m3/tahun, ## g C/m2/tahun, ### g C/m2/hari.
Nitrogen Inorganik Terlarut (DIN)
Nitrogen inorganik terlarut di perairan terdiri dari ammonia-nitrogen (NH3-N),
nitrat-nitrogen (NO3-N), dan nitrit -nitrogen (NO2-N). Nitrogen dalam laut di dapatkan
dalam 5 tingkat oksidasi, dan dari kelima tingkatan tersebut yang melimpah dan
Daerah Produkti fitas primer Sumber
Perairan Tropik Brown (1980) in Shannon dan Pilar (1986) Brown (1984) in Shannon dan Pilar (1986) Carter (1982, 1983) in Shannon dan Pilar (1986) Brown (1984) in Shannon dan Pilar (1986)
aktif adalah ion nitrat, berturut-turut menyusul nitrit dan ammonia (Libes 1992;
Valiela 1984).
Sumb er nitrogen di laut terbesar berasal dari udara, sekitar 80% dalam bentuk
nitrogen bebas yang masuk melalui sistem fiksasi biologis dalam kondisi aerobik
(Dawes 1981). Meskipun nitrogen ditemukan berlimpah di lapisan atmosfer akan
tetapi unsur ini tidak dapat dimanfaatkan secara langsung oleh makhluk hidup. Untuk
dapat dimanfaatkan nitrogen dari atmosfir yang masuk ke dalam perairan difiksasi
(diserap) oleh sebagian bakteri atau fitoplankton menjadi senyawa-senyawa tertentu,
seperti NH3, NH4 dan NO3 (Valiela 1984).
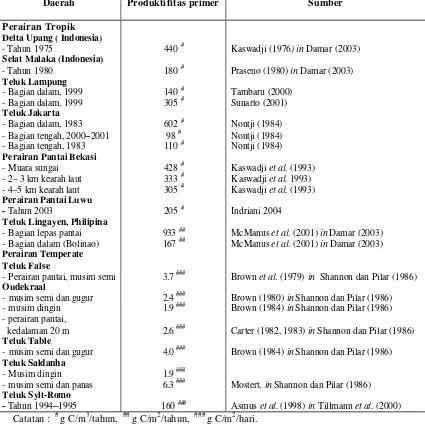
Sumber lain nitrogen di laut berasal dari zona dalam perairan yang disebut
sebagai produksi baru terutama dalam bentuk nitrat-nitrogen (NO3-N) yang masuk ke
zona euphotik yang terangkut melalui vertikal mixing dan upwelling (Tett dan
Edwards 1984; Lalli dan Parsons 1993) (Gambar 2). Fitoplankton lebih banyak
menyerap NH4-N dari pada NO3-N karena lebih banyak dijumpai dalam kondisi
aerobik maupun anaerobik.
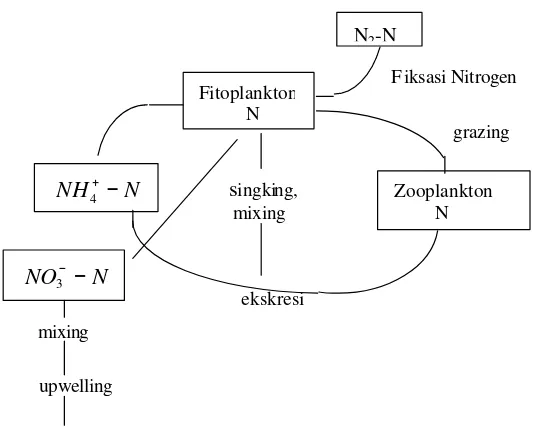
Kebanyakan spesies fitoplankton dapat mengasorbsi ammonium, ammonia,
nitrat, maupun nitrit, tetapi jika ketiganya tersedia, fitoplankton pada umumnya lebih
menyukai ammonium (Raymont 1963; Riley dan Chester 1971; Millero dan Sohn
1991; Libes 1992 ). Laju penyerapan nitrogen lebih cepat dari sel fitoplankton yang
berukuran kecil daripada yang berukuran besar (Eppley et al. 1969; Fricble et al. 1978 in Smith and Kalf 1983; Harrison et al. 2004) (Gambar 3).
Ortofosfat adalah bentuk fosfor yang secara langsung dimanfaatkan oleh
tumbuhan akuatik. Sedangkan polifosfat harus mengalami hidrolisis untuk
membentuk ortofosfat sebelum dimanfaatkan sebagai fosfor. Fosfor merupakan salah
satu unsur penting dalam pertumbuhan dan metabolisme tubuh diatom. Fosfat dapat
menjadi faktor pembatas, baik secara temporal maupun spasial (Raymont 1963).
Keberadaan fosfor di perairan alami biasanya relatif kecil, kadarnya lebih kecil
daripada nitrogen, karena sumber fosfor yang lebih sedikit bila dibandingkan dengan
sumber nitrogen. Sumber fosfor alami yang terdapat di dalam air berasal dari
pelapukan batuan mineral dan hasil dekomposisi organisme yang telah mati, Fitoplankton
< 2 µm
sedangkan sumber antropogenik fosfor berasal dari limbah industri dan domestik,
limbah deterjen, serta limpasan limbah pertanian yang menggunakan pupuk
(Libes 1992).
Soegiarto dan Birowo (1975) menyatakan kandungan fosfat pada lapisan
permukaan lebih rendah dari lapisan di bawahnya, sehingga kandungan fosfat yang
tinggi di lapisan permukaan dapat dipakai sebagai indikasi terjadinya proses penaikan
massa air. Kandungan fosfat yang optimal bagi pertumbuhan fitoplankton berada pada
kisaran 0.27–5.51 ppm (Bruno et al. 1979 in Widjaja et al. 1994).
Fosfat mempengaruhi komposisi fitoplankton, pada perairan yang memiliki
nilai fosfat rendah (0,00–0,02 ppm) akan dijump ai dominansi diatom terhadap
fitoplankton yang lain, dan pada perairan dengan nilai fosfat sedang (0,02–0,05 ppm)
akan banyak dijumpai jenis Chlorophyceae, sedangkan pada perairan dengan nilai
fosfat tinggi (>0,10 ppm) akan didominasi oleh Cyanophyceae (Moyle 1946 in Kaswadji 1976).
Silikat
Silika atau silika dioksida (SiO2) merupakan bagian yang penting dalam
pertumbuhan struktur silikoflagelata seperti diatom, radiolaria dan sponge. Menurut
Millero dan Sohn (1991) silika di laut rata-rata 50% dalam bentuk anorganik dan
sisanya kebanyakan menjadi kalsium karbonat (CaCO3). Silikat di perairan terdapat
dalam bentuk larutan asam silikat, dalam bentuk biota (dinding sel diatom),
silikoflagellata dan radiolaria, baik yang hidup maupun yang mati (Spencer 1975).
Bagi diatom, silikat merupakan nutrien yang sangat penting untuk membangun
dinding selnya dan mengasimilasi sejumlah besar silikat untuk disintesis menjadi
penguraiannya dipengauhi oleh karbondioksida bebas dan asam-asam organik dalam
perairan (Chen 1971).
Spencer (1975) mengemukakan dalam air laut silikon kemungkinan berada
dalam bentuk Si(OH)4. Unsur silikat terdapat sebagai silikat dalam air laut dan
mungkin juga dalam bentuk larutan sejati sebagai ion-ion silikat (Raymont 1963).
Diatom menutupi dirinya dengan kerangka transparan yang merupakan timbunan dari
sebagian besar silikat dalam bentuk senyawa-senyawa SiO2 (Black 1986).
Distribusi silikat di perairan tergantung pada lokasi dan kedalaman perairan.
Distribusi silikat di perairan pantai biasanya lebih tinggi daripada di laut terbuka
karena pengaruh oleh aliran sungai. Di laut terbuka kandungan silikat akan meningkat
bila kedalaman meningkat. Di perairan laut konsentrasi silikat bervariasi dari
0–0.05 mg/L (Dawes 1981), atau 0–200 µM (Millero dan Sohn 1991).
Fitoplankton
Parson et al. (1984) mengatakan bahwa terdapat 13 kelas dari fitoplankton yang terdapat di laut yang terdiri dari Cyanophyceae (alga biru hijau), Rhodophyceae
(alga merah), Bacillarophyceae (Diatom), Cryptophyceae (Cryptomonads),
Dinophyceae (Dinofllagellata), Chrysophyceae (Chrysomonads, Silicoflagellata),
Haptophyceae atau Prymnesiophyceae (Coccolithophorids, Prymnesiomonads),
Raphidiophyceae (Choromonadea), Xanthophyceae (alga kuning hijau),
Eustigmatophyceae, Euglenophyceae (Euglenoids), Prasinophyceae
(Prasinomonads), dan Chlorophyceae (alga hijau). Tetapi hanya 4 kelas saja yaitu
Bacillariophyceae, Cryptophyceae, Dinophyceae, dan Haptophyceae yang memegang
fitoplankton yang mempunyai kelimpahan tertinggi di ekosistem laut adalah dari kelas
diatom (Sze 1993 ).
Nybakken (1988) mengemukakan jenis fitoplankton yang sering dijumpai di
laut dalam jumlah besar adalah Diatom dan Dinoflagellata. Fitoplankton yang
minoritas di laut ialah berbagai jenis alga hijau biru (Cyanophyceae), kokolitofor
(Coccolithophorids), dan silikoflagellata (Dyctyochaceae, Chrysophyceae)
(Nybakken 1988; Romimohtarto dan Juwana 1999).
Nontji (1984) mengatakan bahwa fitoplankton dengan kelimpahan tinggi
umumnya terdapat di perairan sekitar muara sungai, dimana terjadi proses penyuburan
karena masuknya nutrien dari daratan yang dialirkan oleh sungai ke laut. Fitoplankton
umumnya lebih padat di perairan dekat pantai dan makin berkurang pada perairan
yang ke arah laut lepas, selain itu penyebarannya tidak merata melainkan hidup secara
berkelompok (Arinardi et al. 1997).
Goldman dan Horne (1983) dan Wetzel (1983) mengemukakan dominasi suatu
jenis fitoplankton pada suatu perairan dapat diganti oleh jenis lain, disebabkan
berubahnya kondisi fisik kimia perairan. Kondisi lingkungan yang merupakan faktor
penentu keberadaan fitoplankton adalah suhu, salinitas, cahaya matahari, pH,
kekeruhan, konsentrasi nutrien, dan berbagai senyawa lainnya (Nybakken 1988).
Klorofil-a
Klorofil-a dengan rumus kimia C55H72O5N4Mg (Weyl 1970) merupakan salah
satu pigmen fotosintesa yang paling penting bagi tumbuhan yang ada di perairan
khususnya fitoplankton (Parsons et al. 1984; Susilo 1999). Klorofil-a terdapat dalam
mengukur biomass fitoplankton (Strickland dan Parsons 1965) dan dapat digunakan
sebagai petunjuk nilai potensi fotosintetik di perairan (Wiadnyana 1997).
Pada perairan laut fitoplankton memegang peranan terpenting sebagai
produsen primer, karena merupakan komponen utama tumbuhan yang mengandung
klorofil. Pigmen fitoplankton yang sering digunakan dalam mempelajari produktivitas
perairan adalah klorofil-a (Strickland dan Parsons 1965).
Sebaran klorofil-a di dalam kolom perairan sangat tergantung pada konsentrasi
unsur hara. Unsur hara memiliki konsentrasi rendah dan berubah-ubah pada
permukaan laut dan konsentrasi akan meningkat dengan bertambahnya kedalaman
(Millero dan Sohn 1991).
Cahaya
Parsons et al. (1984) mengatakan bahwa aspek dasar dari cahaya yang penting
secara biologi adalah kuantitas dan kualitasnya. Kedua karakter ini berfluktuasi di
laut, bergantung kepada waktu (harian, musiman, dan tahunan), ruang (perbedaan
lokasi di bumi dan kedalaman), kondisi cuaca, penyebaran sudut datang termasuk arah
perubahan maksimum dan tingkat difusi, dan polarisasi. Cahaya merupakan sumber
energi dalam fotosintesis. Proses fotosintesis di dalam perairan hanya dapat
berlangsung jika ada cahaya sampai pada kedalaman tertentu dimana fitoplankton
berada. Pada tahap awal cahaya matahari ditangkap oleh fitoplankton, kemudian
energi ini digunakan untuk aktivitas proses fotosintesis. Tidak semua radiasi
elektromanetik yang jatuh pada tumbuhan berfotosintesis dapat diserap, tetapi hanya
cahaya tampak (visible light) yang memiliki panjang gelombang berkisar antara
400–720 nm yang diabsorpsi dan digunakan untuk melakukan aktivitas fotosintesis
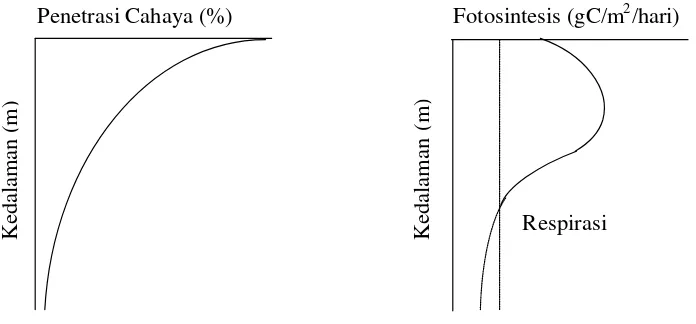
Ruttner (1973) mengatakan bahwa makin dalam penetrasi cahaya ke dalam
perairan menyebabkan semakin besar daerah dimana proses fotosintesis dapat
berlangsung, sehingga kandungan oksigen terlarut masih tinggi pada lapisan air yang
lebih dalam. Penetrasi cahaya matahari dalam air, semakin menurun dengan
bertambahnya kedalaman (Gambar 4).
Penetrasi Cahaya (%) Fotosintesis (gC/m2/hari)
Respirasi
Gambar 4. Grafik distribusi vertikal cahaya dan fotosintesis di perairan (Lalli dan Parsons 1993; Mann dan Lazier 1996).
Umumnya fotosintesis bertambah sejalan dengan meningkatnya intensitas
cahaya sampai pada suatu nilai optimum tertentu (cahaya saturasi). Di atas nilai
optimum, cahaya merupakan penghambat bagi fotosintesis (cahaya inhibisi),
sedangkan dibawahnya merupakan cahaya pembatas (limitasi) sampai pada suatu
kedalaman dimana fotosintesis sama dengan respirasi (Mann 1982; Parsons et al. 1984; Valiela 1984). Kennish (1990 ) mengatakan bahwa intensitas cahaya yang
masuk di perairan sangat dipengaruhi oleh banyaknya padatan tersuspensi, jasad renik
yang melayang, kekeruhan dan warna air. Intensitas cahaya ini semakin melemah saat
penetrasi ke dalam kolom air. Hukum Lambert-Beer (Foog 1975; Parsons et al. 1984) dapat digunakan untuk menghitung besarnya tingkat absorpsi cahaya yang
ditunjukkan oleh besarnya koefisien absorbsi, yaitu :
Iz = Io e -kz
Dimana Iz adalah intensitas cahaya pada suatu kedalaman z, Io adalah intensitas
cahaya pada permukaan air, e adalah bilangan dasar logaritma (2,7), dan k adalah
koefisien absorbsi.
Respon fitoplankton terhadap intensitas cahaya juga sangat dipengaruhi oleh
pigmen yang dikandungnya. Perbedaan pigmen yang dikandung antara jen is
fitoplankton menyebabkan perbedaan intensitas cahaya yang diabsorbsi. Hal ini
berpengaruh terhadap tingkat efisiensi fotosintesis. Spektrum cahaya yang terpenting
dalam mengontrol fotosintesis fitoplankton adalah yang mempunyai panjang
gelombang 400–700 nm, atau yang dikenal dengan photosynthetically active radiation
(PAR) (Kennish 1990 ; Lalli dan Parsons 1993).
Suhu
Suhu merupakan salah satu faktor fisika yang dapat mempengaruhi
fotosintesis dan pertumbuhan fitoplankton. Suhu berpengaruh pada sistem biologi
melalui dua cara. Pertama suhu, berpengaruh terhadap kecepatan reaksi-reaksi secara
enzimatik dalam tubuh organisme. Kedua, suhu berpengaruh terhadap proses respirasi
organisme. Peningkatan suhu pada batas kisaran toleransi akan meningkatkan laju
metabolisme dan aktivitas fotosintesis fitoplankton. Peningkatan suhu sebesar
10oC akan menyebabkan peningkatan laju fotosintesis sebesar dua kali lipat
(Kennish 1990).
Dalam proses fotosintesis di laut, suhu dapat berpengaruh secara langsung dan
tidak langsung. Pengaruh secara langsung yaitu mengontrol reaksi kimia enzimatik
struktur hidrologi perairan, seperti kerapatan air yang akhirnya mempengaruhi laju
penenggelaman fitoplankton (Raymont 1963; Tomascik et al. 1997).
Soegiarto dan Birowo (1975) mengemukakan bahwa keadaan sebaran suhu
secara horisontal di perairan Indonesia memperlihatkan variasi tahunan yang kecil
namun masih memperlihatkan adanya perubahan musiman. Hal tersebut, berhubungan
dengan yang dikemukakan oleh Damar (2003) bahwa suhu perairan tropik
menunjukkan variasi harian yang kecil (siang dan malam), termasuk fluktuasi
musiman. Selanjutnya dinyatakan bahwa kisaran suhu tahunan perairan berkisar dari
29.1–30.0°C.
Suhu dalam lautan bervariasi sesuai dengan kedalaman. Massa air permukaan
di wilayah tropik panas sepanjang tahun, yaitu 20–30oC. Suhu lapisan permukaan di
perairan Indonesia berkisar antara 26–30oC, lapisan termoklin berkisar antara 9–26oC
dan pada lapisan dalam berkisar antara 2–8oC (Soegiarto dan Birowo 1975).
Salinitas
Berdasarkan salinitas dikenal dua jenis fitoplankton, yaitu yang bersifat
stenohaline (dapat hidup pada kisaran salinitas yang sempit) dan euryhaline (dapat
hidup pada kisaran salinitas yang lebar). Salinitas mempengaruhi fitoplankton dalam
hal densitas dan stabilitas dari kolom air. Peningkatan kedalaman akan menurunkan
suhu dan meningkatkan salinitas, hal ini menyebabkan densitas meningkat sehingga
mempengaruhi laju penenggelaman fitoplankton dan akan mempunyai stratifikasi
yang kuat dengan lapisan pegat (discontinuity) yang tajam, akan sukar ditembus oleh
fitoplankton (Raymont 1963).
Kaswadji et al. (1993) mengatakan bahwa variasi salinitas mempengaru hi laju
bertahan pada batas-batas salinitas yang kecil. Sachlan (1982) mengatakan bahwa
salinitas yang cocok bagi fitoplankton adalah lebih dari 20‰. Salinitas seperti ini
memungkinkan fitoplankton dapat bertahan hidup, memperbanyak diri, dan aktif
melakukan fotosintesis (NairdanThampy 1980).
pH
Wardoyo (1982) mengatakan bahwa pH sangat mempengaruhi kehidupan
makhluk hidup termasuk fitoplankton. Selain itu pH merupakan salah satu parameter
yang dapat menentukan produktivitas suatu perairan, dan pH yang ideal untuk
kehidupan fitoplankton dalam perairan adalah 6,5–8,0 (Pescod 1973). pH permukaan
laut sangat stabil biasanya berkisar antara 8.1 dan 8.3 (Reid 1961).
Swingle (1969) mengatakan bahwa pada perairan yang berkondisi asam
dengan pH kurang dari 6, fitoplankton tidak akan hidup dengan baik. Perairan dengan
nilai pH lebih kecil dari 4 merupakan perairan yang sangat asam dan dapat
menyebabkan kematian makhluk hidup, sedangkan pH lebih dari 9,5 merupakan
perairan yang sangat basa dapat pula menyebabkan kematian dan mengurangi
produktifitas (Wardoyo 1982). Air yang bersifat basa dan netral cenderung lebih
produktif dibandingkan dengan air yang bersifat asam (Hickling 1971).
Fitoplankton dapat hidup subur pada pH 7–8 asalkan terdapat cukup mineral di
dalam suatu perairan (Sachlan 1982), sedangkan pH optimal untuk perkembangan
METODE PENELITIAN
Tempat dan Waktu Penelitian
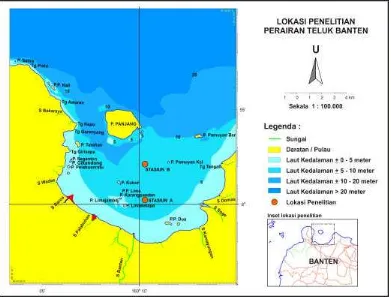
Penelitian ini dilaksanakan di perairan Teluk Banten yang secara geografis
terletak pada 05°55’–06°05’ LS dan 106°05’–106°15’ BT. Tempat pengambilan
contoh air laut terbagi dalam dua stasiun, yakni stasiun A dengan posisi geografis
pada 06°00’20.3” LS dan 106°09’06.2” BT serta stasiun B dengan posisi geografis
pada 05°58’38.3” LS dan 106°09’35.7” BT (Gambar 5). Pengukuran produktivitas
primer serta analisis unsur hara, fitoplankton dan klorofil-a berlangsung pada musim
peralihan I (pertama) dan dilaksanakan dalam 3 periode, dimana periode pertama
berlangsung pada Tanggal 27 Maret 2005, periode kedua berlangsung pada Tanggal
9 April 2005 dan periode ketiga berlangsung pada Tanggal 25 April 2005.
Pengambilan contoh Air Laut
Pengambilan contoh air laut dengan menggunakan Van Dorn kapasitas
5 liter pada kedalaman 0.20 meter, 1 meter, 4 meter dan 5 meter di stasiun A dan
kedalaman 0.20 meter, 2 meter, 5 meter dan 6 meter di stasiun B. Pembagian atas
beberapa kedalaman tersebut dimaksudkan karena distribusi vertikal cahaya akan
semakin berkurang dengan bertambahnya kedalaman serta distribusi vertikal unsur
hara konsentrasinya selalu bervariasi dengan kecenderungan akan semakin besar
dengan bertambahnya kedalaman.
Pengambilan contoh air laut dilakukan satu kali pada masing-masing
kedalaman setiap periode, dimulai pada jam 08.00 bersamaan akan dilakukan proses
inkubasi. Parameter yang diukur meliputi produktivitas primer, unsur hara DIN
(nitrogen inorganik terlarut) yang meliputi NH3-N, NO3-N, NO2-N, DIP (fosfat
inorganik terlarut) yang meliputi ortofosfat, silikat, fitoplankton dan klorofil-a sebagai
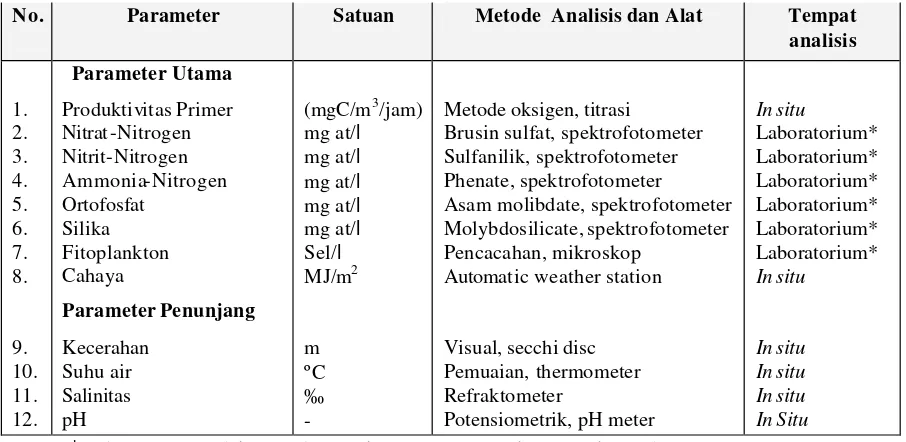
parameter utama serta suhu, salinitas, dan pH sebagai parameter penunjang (Tabel 2).
Tabel 2. Parameter fisika, kimia dan biologi yang diukur serta metode dan alat ukur yang digunakan
Intensitas Cahaya Matahari
Intensitas Cahaya Matahari permukaan tercatat setiap 10 menit dengan
alat Automatic Weather Station (AWS) tipe JY 106 dari Badan Meteorologi dan Geofisika Serang. Distribusi intensitas cahaya matahari pada setiap kedalaman kolom
air ditentukan menurut Hukum Beer -Lambert (Fogg, 1975; Parsons et al. 1984) sebagai berikut :
Dimana : Iz = Intensitas cahaya pada kedalaman z
Io = Intensitas cahaya permukaan
kT = Koefisien peredupan
z = Kedalaman.
Koefisien peredupan dihitung dari pembacaan kedalaman keping secchi disk
(Sd (m)) dengan menggunakan hubungan persamaan empiris (Tillmann et al. 2000), sebagai berikut : k = 0.191 + 1.242/Sd (r2 = 0.853).
Produktivitas Primer
Pengukuran produktivitas primer dilakukan dengan metode oksigen botol
terang-botol gelap. Prinsip kerja metode ini adalah mengukur perubahan kandungan
oksigen dalam botol terang dan botol gelap yang berisi contoh air setelah
diinkubasikan pada perairan yang mendapat sinar matahari (Lampiran 1, 2 dan 3).
Produktivitas primer bersih dengan nilai oksigen terlarut dikonversi kedalam satuan
mgC/m3/jam (Umaly danCuvin 1988) sebagai berikut :
z k o z
T e I
dimana :
NPP = Produktivitas primer bersih (mg C/m3/jam)
O2BT = Oksigen pada botol terang (BT) setelah inkubasi (mg/l)
O2BA = Oksigen pada botol inisial (BI) (mg/l)
PQ = Photosintetic Quotien = 1,2; dengan asumsi bahwa hasil metabolisme sebagian besar didominasi oleh fitoplankton
t = Waktu inkubasi (jam) 1000 = Konversi liter menjadi m3
0,375 = Koefisien konversi oksigen menjadi carbon (=12/32)
Catatan : PQ merupakan perbandingan O2 yang dihasilkan dengan CO2 yang digunakan melalui proses fotosintesis. Nilai PQ berkisar 1.1–1.3 (Ryther 1965inParsons et al. 1984 ; Lalli dan Parsons 1993).
Analisis Unsur Hara
Contoh air laut dimasukkan pada botol sampel polyetilen kapasitas 250 ml
untuk analisis ammonia, nitrat, nitrit, dan silika, serta untuk analisis ortofosfat. Botol
sampel dimasukkan kedalam pendingin sebelum dianalisis.
Sebelum analisis lanjutan di laboratorium terlebih dahulu dilakukan filtrasi
yaitu contoh air laut disaring dengan filter nukleopore (diameter 47 mm dan porositas
0,45 µm) yang dibantu dengan menggunakan pompa vakum melewati suatu glass
microfibre filter (Lampiran 4). NPP =
(
) (
)
( )
t PQx x BA
BT
O
Identifikasi Fitoplankton
Contoh air laut diambil sebanyak 20 liter dan disaring dengan plankton net mesh size 25 µm. Contoh air laut yang telah disaring dimasukkan kedalam botol
sampel 100 ml lalu diawetkan dengan larutan lugol pekat 1 mI/100 ml, untuk
diidentifikasi. Identifikasi jenis fitoplankton dilakukan dengan menggunakan literatur
dari Davis (1955), Smith (1977), Yamaji (1979), dan Tomas (1997).
Kelimpahan fitoplankton dihitung dengan menggunakan metode sensus
(penyapuan) di atas Sedwick Rafter Cell (SRC) (APHA 1998), dengan rumus sebagai
berikut:
Analisis komunitas fitoplankton dengan menggunakan beberapa indeks biologi
seperti indeks keanekaragaman (H’), indeks keseragaman (E), indeks dominansi (D).
Hasil identifikasi dan perhitungan kelimpahan fitoplankton digunakan untuk
menentukan indeks keanekaragaman Shannon-Wienner, yaitu :
dimana : H’ = Indeks keanekaragaman Shannon-Wiener (bits)
Kisaran indeks keanekaragaman Shannon-Wiener dapat dikategorikan sebagai
berikut : (Wilhm dan Doris 1968 inMasson 1981).
H’ < 2.3062 = Keanekaragaman rendah dan kestabilan komunitas rendah
2.3062<H’<6.9078 = Keanekaragaman sedang dan kestabilan komunitas sedang
H’>6.9078 = Keanekaragaman tinggi dan kestabilan komunitas tinggi.
Untuk melihat keseragaman populasi fitoplankton pada setiap pengambilan
sampel dilakukan perhitungan indeks keseragaman (E) atau Equitability, yaitu :
dimana : E = indeks keseragaman
H’ = indeks keanekaragaman H’maks = Ln S
S = jumlah spesies.
Indeks keseragaman berkisar antara 0–1. Semakin kecil nilai E, semakin kecil
pula keseragaman populasi yang berarti penyebaran jumlah individu setiap spesies
tidak sama dan ada kecenderungan terjadi dominansi oleh satu spesies dari jenis yang
ada. Semakin besar nilai E tidak ada yang mendominasi antar jenis yang ada
(Odum 1971).
Untuk melihat adanya dominansi oleh spesies tertentu pada suatu populasi
digunakan indeks dominansi Simpson, yaitu :
Indeks dominansi berkisar 0–1, bila D mendekati 0 berarti dalam struktur
spesies lainnya dan bila D mendekati 1 berarti di dalam struktur komunitas yang
sedang diamati dijumpai spesies yang mendominasi spesies lainnya (Odum 1971).
Hubungan antara H’, E, dan D adalah apabila nilai indeks keanekaragaman
(H’) spesies tinggi berarti nilai keseragaman (E) rendah dan tidak ada spesies yang
mendominasi spesies lainnya (D rendah).
Klorofil-a
Contoh air laut untuk analisis klorofil-a diambil sebanyak 1 liter dan
dimasukkan kedalam botol polietilen kapasitas 1 liter (ditutup rapat dengan plastik
hitam), dan disimpan dalam box ice bersuhu dingin (-4°C) untuk dianalisis di
laboratorium (Lampiran 5). Konsentrasi klorofil-a diukur dengan menggunakan
spekrofometer. Konsentrasi klorofil-a dihitung dengan menggunakan persamaan
menurut APHA (1998), sebagai berikut :
Analisis Data
Data penelitian dikelompokkan dalam dua kelompok, yaitu data penelitian
utama yang terdiri dari data produktivitas primer, unsur hara, fitoplankton, klorofil-a,
dan cahaya serta data penunjang yang terdiri data suhu, salinitas dan pH. Data hasil
analisis akan ditabulasikan dalam tabel berdasarkan lokasi (stasiun) dan kedalaman
inkubasi di kedua stasiun pengamatan.
Analisis data dilakukan secara komputasi dengan menggunakan software
Kgraph, SAS versi 8.0, dan Excel. Sedangkan rancangan yang digunakan dalam
penelitian ini adalah rancangan acak faktorial blok (Mattjik dan Sumertajaya 2000)
pada 2 lokasi dan setiap lokasi terdiri atas 4 titik kedalaman dengan 3 kali ulangan.
Untuk mengetahui perbedaan produktifitas primer antar lokasi dilakukan
analisis ragam. Sedangkan untuk mengetahui pola hubungan antara unsur hara,
klo rofil-a dengan produktifitas primer, pada setiap stasiun dan kedalaman inkubasi di
gunakan analisis regresi sederhana (Mattjik dan Sumertajaya 2000 ), dengan
persamaan sebagai berikut :
Y = a + bX
dimana : Y = Produktivitas primer sebagai peubah tak bebas
X = peubah bebas berupa unsur hara (ammonia-nitrogen, nitrat- nitrogen,
nitrit-nitrogen, ortofosfat, dan silikat)
b = interseps
a = koefesien regresi.
Sedangkan untuk mengetahui pola hubungan cahaya dengan produktivitas
primer dengan menggunakan model Von Platt (Platt et al. 1980; Damar 2003) dengan
Y = a (1 – e-bX) e-cX
Dimana : Y = Produktivitas primer
X = Cahaya
a, b, dan c = Konstanta.
Nilai koefisien determinasi (R2) digunakan untuk mengetahui keeratan dari
peubah X terhadap Y. Kisaran nilai R2 antara 0–1. Jika nilainya lebih besar dari 0,5
atau mendekati 1, maka dapat diartikan bahwa X memiliki peranan terhadap Y.
Besarnya peranan X terhadap Y, ditelaah dengan sidik ragam regresi. Jika Fhitung lebih
besar dari Ftabel berarti peubah X memberikan pengaruh terhadap peubah Y, demikian
pula sebaliknya jika Fhitung lebih kecil dari Ftabel berarti peubah X tidak memberikan
HASIL DAN PEMBAHASAN
Kondisi Oseanografi
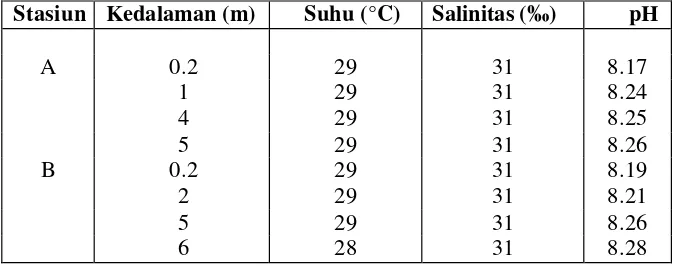
Suhu
Suhu perairan selama penelitian di perairan Teluk Banten relatif sama di
seluruh kedalaman kolom air di stasiun A dan B yang berkisar dari 28–29°C (Tabel
3). Hal ini disebabkan karena kondisi cuaca pada saat pengamatan cenderung sama.
Berhubungan dengan hal tersebut Lalli dan Parsons (1993) menyatakan bahwa
perubahan suhu di perairan tropik yang dangkal lebih kecil dari 2°C. Hasil
penelitian yang didapatkan pada pengamatan ini relatif sama dengan penelitian
sebelumnya, di mana kisaran suhu perairan pada kedalaman 0 meter, 5 meter, dan
10 meter sebesar 28–30°C (Purwanto 1992).
Tabel 3. Kondisi oseanografi perairan Teluk Banten
Stasiun Kedalaman (m) Suhu (°C) Salinitas (‰) pH
A 0.2 29 31 8.17
1 29 31 8.24
4 29 31 8.25
5 29 31 8.26
B 0.2 29 31 8.19
2 29 31 8.21
5 29 31 8.26
6 28 31 8.28
Secara keseluruhan, suhu di perairan Teluk Banten relatif lebih tinggi dari
suhu air laut rata-rata di laut Jawa yang nilainya sebesar 27.93°C dengan kisaran
27.60–28.60°C (Wyrtki 1961). Kisaran nilai suhu yang didapatkan pada penelitian
ini merupakan suhu yang optimum bagi pertumbuhan fitoplankton. Kisaran suhu yang
Salinitas
Salinitas di perairan Teluk Banten relatif sama di stasiun A dan B maupun
kedalaman inkubasi selama pengamatan, nilai salinitas yang teramati adalah
31‰ (Tabel 3). Tidak adanya perbedaan salinitas di kedua stasiun tersebut
kemungkinan disebabkan oleh posisi stasiun pengamatan berada pada bagian tengah
perairan Teluk Banten, yang sama-sama mendapat pengaruh air laut dan air tawar
yang terbawa oleh aliran sungai. Salinitas yang didapatkan pada pengamatan
ini tidak banyak mengalami perubahan bila dibandingkan dengan penelitian
sebelumnya pada perairan yang sama, dimana salinitas pada kedalaman 0 meter,
5 meter, dan 10 meter berkisar dari 31–32‰ (Purwanto 1992).
Dilihat dari fluktuasi salinitas dengan penelitian sebelumnya, maka massa air
di perairan Teluk Banten tergolong kedalam massa air pantai dengan salinitas kurang
dari 32.0‰ (Wyrtki 1961). Kisaran nilai salinitas masih merupakan nilai yang sesuai
untuk pertumbuhan dan perkembangan fitoplankton. Nilai salinitas di atas 20‰
memungkinkan fitoplankton dapat bertahan hidup, memperbanyak diri, dan aktif
melakukan proses fotosintesis (Sachlan 1982).
pH
Nilai pH di stasiun A dan B maupun pada semua kedalaman inkubasi relatif
sama selama pengamatan dengan kisaran 8.17–8.28 (Tabel 3). Kisaran nilai tersebut
tidak jauh berbeda bila dibandingkan dengan penelitian sebelumnya di perairan yang
sama, dimana pada kedalaman 0 meter, 5 meter, dan 10 meter berkisar dari 7.6–8.2
(Purwanto 1992).
Nilai pH yang didapatkan masih dalam batas untuk pertumbuhan fitoplankton.
pH tersebut masih merupakan kisaran pH perairan laut Indonesia yang berkisar dari
6.0–8.5 (Romimohtarto 1991). Perairan laut tropis memiliki kisaran pH dari 7.5–8.4
(Nybakken 1988).
Intensitas Cahaya Matahari
Intensitas Cahaya Matahari Permukaan
Sumber energi di laut berasal dari cahaya matahari. Cahaya yang dipancarkan
matahari terdiri dari 50% sebagai infra merah, 41% sebagai cahaya terang, 9%
sebagai ultraviolet, sinar lamda dan sinar gamma (Anikouchine dan Sternberg 1981).
Penetrasi cahaya matahari pertama mencapai atmosfir kemudian laut. Cahaya yang
terserap atau terpencar di atmosfer sekitar 50% dan mencapai permukaan laut 50%
(Lalli dan Parsons 1993). Besarnya nilai intensitas cahaya matahari yang mencapai
permukaan laut berbeda-beda dari waktu ke waktu. Fenomena tersebut terjadi pula di
perairan Teluk Banten, dimana intensitas cahaya matahari yang mencapai permukaan
laut (di udara) selama pengamatan bervariasi (Gambar 6; Lampiran 6).
0 10 20 30 40 50 60 70 80
06:00 07:00 08:00 09:00 10:00 11:00 12:00 13:00 14:00 15:00 16:00 17:00 18:00
Waktu Pengamatan
Intensitas Cahaya Matahari (MJ/m
2 )
Gambar 6. Intensitas cahaya permukaan.
Bervariasinya intensitas cahaya disebabkan oleh kondisi perawanan pada saat
matahari yang mencapai permukaan laut. Hal ini berhubungan dengan pernyataan
Fritz (1957) in Parsons et al. (1984) bahwa cakupan awan di udara akan mempengaruhi intensitas cahaya matahari yang mencapai permukaan laut.
Intensitas Cahaya Matahari di Kolom Air
Penetrasi cahaya matahari (50% cahaya permukaan) akan mengalami
pengurangan sekitar 10% di lapisan permukaan (Kirk 1994) atau 90% mencapai
lapisan permukaan laut (Iwasaka et al. 2000). Sama halnya dengan intensitas cahaya permukaan, intensitas cahaya di lapisan permukaan bervariasi dari waktu ke waktu
(Gambar 7; Lampiran 7).
Dari Gambar 6 dan 7 terlihat bahwa besarnya intensitas cahaya pada
permukaan laut akan mempengaruhi pula nilai intensitas cahaya di lapisan
permukaan. Variasi ini terutama terjadi menurut waktu, dimana pada pagi hari
intensitasnya rendah dan akan mengalami peningkatan atau memuncak pada siang
hari (tengah hari) dan menurun kembali pada sore hari.
Berdasarkan Gambar 7, terlihat intensitas cahaya matahari selama inkubasi
pada lapisan permukaan cenderung merata dari jam 10:00–12:00 WIB dimana pada
saat itu sudut datang cahaya matahari hampir tegak lurus permukaan laut. Fenomena
tersebut akan menyebabkan lapisan permukaan laut semakin besar pula menerima
intensitas cahaya matahari dan akan mengalami penurunan dengan bertambahnya
kedalaman (Gambar 8). Hal ini berhubungan dengan pernyataan Sumich (1992)
bahwa adanya perbedaan nilai intensitas cahaya matahari yang masuk ke kolom air
dipengaruhi oleh sudut datang matahari, nilai intensitas semakin besar seiring dengan
besarnya sudut datang matahari dan mengalami penurunan dengan semakin kecilnya
0 10 20 30 40 50 60 70
06:00 07:00 08:00 09:00 10:00 11:00 12:00 13:00 14:00 15:00 16:00 17:00 18:00
Waktu pengamatan
Intensitas cahaya matahari (MJ/m
2)
Gambar 7. Intensitas cahaya di lapisan permukaan.
Distribusi cahaya matahari di kolom air akan mengalami peredupan.
Peredupan cahaya matahari tersebut terlihat dari nilai koefisien peredupan yang
diperoleh selama pengamatan di stasiun A sebesar 0.55 per meter dan di stasiun B
sebesar 0.47 per meter. Peredupan ini disebabkan oleh bahan -bahan yang ada dalam
perairan baik berupa bahan organik maupun partikel-partikel tersuspensi termasuk di
dalamnya organisme planktonik dan molekul-molekul air itu sendiri. Hal ini
berhubungan dengan pernyataan Wyatt dan Jackson (1989) bahwa distribusi cahaya di
kolom air tergantung pada kandungan dan kelompok partikel tersuspensi, seperti tipe
partikel termasuk ukuran, bentuk dan struktur internal.
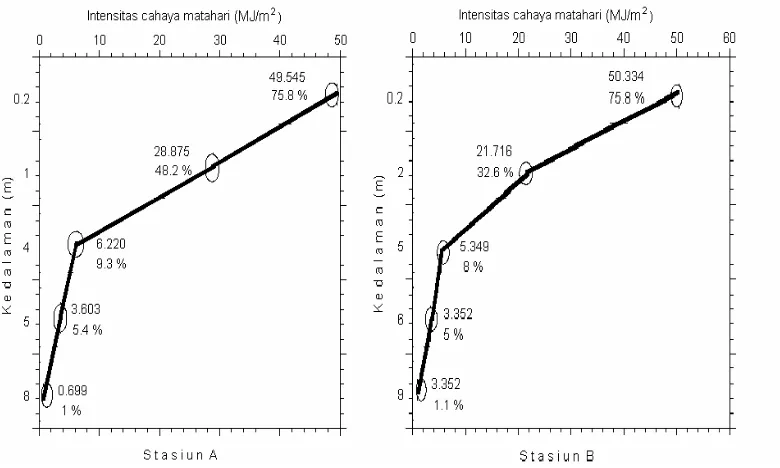
Kedalaman zona eufotik selama pengamatan diukur melalui pembacaan
keping Secchi dengan nilai yang diperoleh sebesar 3.5 meter di stasiun A dan 4.5
meter di stasiun B. Pendugaan nilai dan persentase intensitas cahaya matahari yang
masuk ke kolom air dengan hukum Lambert memperlihatkan pola distribusi cahaya di
perairan semakin dalam cahaya menembus lapisan air maka semakin berkurang nilai
lapisan permukaan pada kedalaman 8 meter di stasiun A dan 9 meter di stasiun B
(Gambar 8; Lampiran 8).
Gambar 8. Distribusi cahaya di kolom air perairan Teluk Banten.
Berdasarkan hal tersebut, dapat dikatakan bahwa kedalaman inkubasi
0.20 meter, 1 meter, 4 meter dan 5 meter di stasiun A serta 0.20 meter, 2 meter, 5
meter dan 6 meter di stasiun B masih termasuk dalam zona eufotik. Dengan demikian
di kedalaman inkubasi tersebut cahaya terdistribusi pada semua kolom air sehingga
akan menunjang terjadinya proses fotosintesis di semua kedalaman inkubasi.
Perbedaan nilai pencahayaan di stasiun A dan B disebabkan oleh berbedanya
nilai koefisien peredupan di kedua stasiun tersebut. Nilai koefisien peredupan di
stasiun A sebesar 0.55 per meter dan di stasiun B sebesar 0.47 per meter. Semakin
besar nilai koefisien peredupan maka nilai intensitas cahaya yang menembus kolom
air akan semakin kecil. Sebaliknya nilai koefisien peredupan yang kecil maka nilai
Nitrogen Inorganik Terlarut (DIN)
Rata-rata total DIN yang diperoleh selama pengamatan sebesar 0.555 mg at
N/l di stasiun A dan 0.550 mg at N/l di stasiun B (Lampiran 9). Nilai ini masih berada
pada kisaran konsentrasi DIN yang umumnya ditemukan di perairan estuari yang
berkisar dari bawah deteksi (0.1 µM) sampai di atas 100 µM atau setara 0.067 mg at
N/l (Nixon dan Pilson 1983). Selama pengamatan terlihat bahwa nitrat merupakan
salah satu unsur yang memberikan kontribusi besar terhadap total DIN di perairan.
Hal ini disebabkan karena nitrat merupakan unsur yang lebih melimpah di lapisan
permukaan perairan (Dawes 1981) dan merupakan bentuk stabil dari nitrogen di laut
(Riley dan Chester 1971).
Kisaran konsentrasi DIN (ammonia, nitrat, dan nitrit) selama pengamatan
cenderung sama dengan variasi yang sangat kecil di kedua stasiun pengamatan
(Tabel 4). Hal ini kemungkinan disebabkan oleh posisi stasiun A dan B yang berada
pada bagian tengah perairan Teluk Banten.
Begitu pula dengan sebaran vertikal DIN cenderung sama dengan variasi yang
sangat kecil pada setiap kedalaman inkubasi di stasiun A dan B (Tabel 5). Adanya
variasi konsentrasi DIN kemungkinan disebabkan oleh posisi kedalaman inkubasi
pada pengamatan ini berada pada zona eufotik, sehingga unsur DIN yang tersedia di Tabel 4. Rataan unsur hara menurut stasiun pengamatan di perairan Teluk Banten
Unsur hara (mg -at/4 l) Stasiun
Periode
ke - NH3-N NO3-N NO2-N PO4-P Si
A I 0.049 0.066 0.002 0.002 1.573
II 0.040 0.106 0.007 0.002 0.255
III 0.076 0.069 0.002 0.011 1.251
B I 0.034 0.082 0.002 0.002 0.752
II 0.039 0.062 0.006 0.002 0.145
perairan dengan cepat dimanfaatkan oleh fitoplankton. Hal ini berhubungan dengan
pernyataan Grasshoff (1976) bahwa konsentrasi unsur hara di zona eufotik cepat
berkurang karena pemanfaatan oleh fitoplankton. Pernyataan tersebut didukung pula
ditemukannya kelimpahan fitoplankton yang tergolong tinggi pada setiap kedalaman
inkubasi di stasiun A dan B (Gambar 9; Lampiran 9).
Tabel 5. Rataan konsentrasi unsur hara menurut kedalaman inkubasi di perairan Teluk Banten
Unsur hara (mg at/3 l) Stasiun
Kedalaman
(m) NH3-N NO3-N NO2-N PO4-P Si
A 0.2 0.045 0.09 0.005 0.006 0.648
1 0.055 0.096 0.005 0.005 0.669
4 0.060 0.094 0.006 0.004 0.529
5 0.047 0.094 0.005 0.004 0.502
B 0.2 0.047 0.053 0.005 0.003 0.372
2 0.051 0.054 0.005 0.003 0.401
5 0.058 0.103 0.005 0.005 0.368
6 0.054 0.096 0.005 0.005 0.373
Ammonia-Nitrogen (NH3-N)
Nilai konsentrasi ammonia di stasiun A berkisar dari 0.040–0.076
mg at NH3-N/4 l, dan stasiun B berkisar dari 0.034–0.079 mg at NH3-N/4 l (Tabel 4;
Lampiran 9). Kisaran nilai konsentrasi ammonia yang didapatkan pada pengamatan
ini lebih rendah bila dibandingkan dengan penelitian sebelumnya yang dilakukan oleh
Purwanto (1992) di perairan yang sama. Kisaran nilai konsentrasi ammonia yang
didapatkan oleh Purwanto (1992) pada kedalaman 0 meter, 5 meter, dan 10 meter
berturut-turut 0.1708–0.244 mg/3 l atau 0.140–0.200 mg at NH3-N/3 l, 0.1859–0.488
mg/3 l atau 0.152–0.401 mg at NH3-N/3 l, 0.0976–0.195 mg/3 l atau 0.080–0.160
Pola distribusi vertikal ammonia di kedua stasiun pengamatan terlihat
cenderung sama dengan variasi yang sangat kecil (Tabel 4). Hal ini berhubungan
dengan pernyataan bahwa distribusi vertikal ammmonia lebih seragam bila dibanding
dengan unsur nitrogen lainnya (Dawes 1981).
Nitrat-Nitrogen (NO3-N)
Konsentrasi nitrat yang didapatkan di stasiun A berkisar dari 0.066–0.106
mg at NO3-N/4 l, dan di stasiun B berkisar dari 0.062–0.105 mg at NO3-N/4 l
(Tabel 4; Lampiran 9), dengan sebaran vertikal di setiap kedalaman inkubasi relatif
sama (Tabel.5; Lampiran 9). Nilai konsentrasi nitrat yang didapatkan pada
pengamatan ini lebih rendah bila dibandingkan dengan yang didapatkan oleh
Purwanto (1992) pada kedalaman 0 meter, 5 meter, dan 10 meter berturut-turut
0.1326–0.353 mg/3 l atau 0.109–0.290 mg at NH3-N/3 l, 0.1768–0.4420
mg/3 l atau 0.145–0.363 mg at NH3-N/3 l, 0.2210–0.3978 mg/3 l atau
0.181–0.327 mg at NH3-N/3 l.
Konsentrasi nitrat yang didapatkan, bila dilihat dari ketersediaannya di
perairan masih berada dalam batas keseimbangan, dimana konsentrasi NO3 d i zona
eufotik sebesar 0.03 µg-at/L atau setara 0.042 mg at NO3/l (Goldman dan Glibert
1983). Bila
konsentrasi nitrat di perairan di bawah 0.7 µM atau setara dengan 0.035 µg at NO3-N/l
maka pembelahan sel fitoplankton akan berhenti (Millero dan Sohn 1992). Sedangkan
untuk pertumbuhan optimal fitoplankton memerlukan kandungan nitrat 0.9–3.5 mg/l
atau 0.7–2.8 mg at NO3-N/l (Mackenthum 1969). Sehubungan dengan hal ini dapat
dikatakan bahwa nitrat yang tersedia di perairan pada saat pengamatan telah
dimanfaatkan oleh fitoplankton untuk pertumbuhannya. Hal ini dipertegas jika
menunjukkan telah terjadi penyerapan nitrat dengan cepat oleh fitoplankton
(Goes et al. 2004).
Nitrit-Nitrogen (NO2-N)
Nitrit ditemukan dalam konsentrasi rendah, karena nitrit merupakan bentuk
senyawa peralihan dari nitrat ke ammonia atau sebaliknya. Kisaran konsentrasi nitrit
di stasiun A berkisar dari 0.002–0.007 mg at NO2-N/3 l, dan di stasiun B berkisar dari
0.002–0.006 mg at NO2-N/3 l (Tabel 4; Lampiran 9). Sedangkan sebaran vertikal di
setiap kedalaman inkubasi terlihat seragam (Tabel 5; Lampiran 9).
Fosfor Inorganik Terlarut (DIP)
Ortofosfat (PO4-P)
Kisaran konsentrasi ortofosfat yang didapatkan selama pengamatan di stasiun
A berkisar dari 0.002–0.011 mg at PO4-P/3 l, dan di stasiun B berkisar dari 0.002–
0.008 mg at PO4-P/3 l (Tabel 4). Bila dibandingkan dengan hasil penelitian yang
dilakukan oleh Purwanto (1992), nilai konsentrasi yang didapatkan pada pengamatan
ini lebih tinggi. Purwanto (1992) mendapatkan konsentrasi fosfat pada lapisan
permukaan, kedalaman 5 meter, dan 10 meter berturut-turut berkisar dari 0.0100–
0.0425 mg/3 l atau 0.00326–0.0138 mg at PO4-P/3 l, 0.0125–0.0450 mg/3 l atau
0.0040–0.0146 mg at PO4-P/3 l, dan 0.0350–0.0450 mg/3 l atau 0.0114–0.0146
mg at PO4-P/3 l.
Millero dan Sohn (1992) menyatakan bahwa pertumbuhan semua jenis
fitoplankton tergantung pada konsentrasi ortofosfat, bila konsentrasi ortofosfat
dibawah 0.3 µM atau setara 0.009 mg at P/l maka perkembangan sel menjadi
terhambat. Konsentrasi yang didapatkan berada dibawah pertumbuhan optimal
ortofosfat yang ada langsung dimanfaatkan oleh fitoplankton. Sedangkan untuk
pertumbuhan optimal fitoplankton konsentrasi ortofosfat yang dibutuhkan berkisar
dari 0.27–5.51 mg/l atau 0.088–1.79 mg-at PO4-P/l (Bruno et al. 1979 in Widjaja et al. 1994).
Rendahnya konsentrasi ortofosfat yang didapatkan pada pengamatan ini
kemungkinan disebabkan (1) ortofosfat yang ada diperairan telah dimanfaatkan oleh
fitoplankton, (2) waktu sampling mempengaruhi nilai konsentrasi ortofosfat, seperti
yang terjadi di Teluk Chesapeake konsentrasi ortofosfat menurun dari 0.4 µM–0.1 µM
atau setara 0.01–0.003 mg at/l pada jam 3 sore dan meningkat dengan cepat setelah
matahari terbenam sampai maksimum pada jam 2 siang (Millero dan Sohn 1992),
(3) kandungan fosfat yang dibutuhkan lebih rendah jika nitrogen yang digunakan
tersedia dalam bentuk ammonia, sedangkan jika nitrogen dalam bentuk nitrat, maka
fosfat yang dibutuhkan lebih tinggi (Chu 1984 in Andarias 1984), atau (4) konsentrasi
ortofosfat makin berkurang ke arah laut, karena sumber utama fosfor di laut adalah
dari aliran sungai (Lewis et al. 1985).
Silikat (Si)
Kisaran konsentrasi silikat yang didapatkan selama pengamatan di stasiun A
berkisar dari 0.255–1.573 mg at Si/4 l, dan di stasiun B berkisar dari 0.145–0.845
mg at Si/4 l (Tabel 4; Lampiran 9). Sedangkan sebaran vertikal di setiap kedalaman
inkubasi cenderung sama den gan variasi yang kecil (Tabel 5; Lampiran 9). Kisaran
silikat yang didapatkan pada pengamatan ini lebih tinggi dari yang didapatkan oleh
Purwanto (1992), dimana pada kedalaman permukaan, 5 meter dan 10 meter berkisar
Silikat yang didapatkan selama pengamatan masih berada pada konsentrasi
yang sesuai untuk pertumbuhan diatom. Diatom tidak dapat berkembang dengan baik
pada konsentrasi silikat yang lebih kecil dari 0.5 mg at Si/l (Effendi 2003). Kisaran
konsentrasi silikat terlarut di laut adalah 1 mg/l, tapi konsentrasi tersebut bervariasi
pada permukaan laut dan perairan laut yang dangkal (Grasshoff 1976).
Dengan melihat kisaran konsentrasi DIN (ammonia, nitrat, dan nitrit), DIP
(ortofosfat) maupun silikat di perairan selama pengamatan, dapat dikatakan bahwa
tingkat variasi unsur-unsur tersebut pada setiap kedalaman inkubasi maupun antar
stasiun tidak terlalu jauh berbeda atau cenderung sama. Perbedaan tersebut selain
disebabkan oleh pemanfaatan fitoplankton di zona eufotik, kemungkinan juga
disebabkan oleh lokasi pengamatan merupakan perairan yang dangkal sehingga
pengadukan massa air sangat kuat, yang menyebabkan kolom air menjadi tercampur
sehingga unsur hara tidak terperangkap pada suatu kedalaman tertentu.
Struktur Komunitas Fitoplankton
Komposisi dan Kelimpahan Fitoplankton
Terdapat tiga kelas fitoplankton yang ditemukan selama pengamatan di
perairan Teluk Banten. Ketiga kelas fitoplankton tersebut terdiri dari
Bacillariophyceae, Dinophyceae, dan Cyanophyceae yang tersebar pada semua
kedalaman inkubasi di kedua stasiun pengamatan dengan jumlah genera yang
bervariasi. Genera fitoplankton dari kelas Bacillariophyceae merupakan yang
dominan pada semua kedalaman inkubasi di stasiun A maupun B dengan 29 genera,
Dinophyceae 6 genera, dan Cyanophyceae 1 genera (Lampiran 11, 12, dan 13).
Bacillariophyceae baik menurut stasiun pengamatan (Tabel 6) maupun menurut
kedalaman inkubasi (Gambar 9).
Melimpahnya genera fitoplankton dari kelas Bacillariophyceae (diatom) dan
Dinophyceae (dinoflagellata) pada setiap kedalaman inkubasi disebabkan karena
fitoplankton dari kedua kelas tersebut merupakan anggota utama fitoplankton yang
terdapat di seluruh bagian perairan laut, baik perairan pantai maupun perairan oseanik
(Arinardi et al. 1997).
Tabel 6. Rataan jumlah genera fitoplankton menurut stasiun pengamatan di perairan Teluk Banten
Kelas fitoplankton Sta siun
Periode
ke - Bacillariophyceae Cyanophyceae Dinophyceae
A I 17 1 4
II 22 1 6
III 21 1 4
B I 19 1 5
II 21 1 2
III 19 1 5
Genera fitoplankton dari kelas Bacillariophyceae yang dominan ditemukan
selama pengamatan terdiri dari Chaetoceros sp, Bacteriastrum sp, Guinardia sp, Rhizosolenia sp, Leptocylindrus sp, Nitzschia sp, Thallasiosira sp, Bidulphia sp, dan Skeletonema sp. Sedangkan dari kelas Dinophyceae adalah Peridinium sp, Ceratium sp, Dinophysis sp, dan Noctiluca sp. Jenis-jenis fitoplankton yang ditemukan dari penelitian sebelumnnya yang predominan diatas 10 % di perairan yang sama terdiri
Gambar 9. Rataan jumlah genera fitoplankton menurut kedalaman inkubasi di perairan Teluk Banten.
Berhubungan dengan hal tersebut Arinardi et al. (1994) menyatakan bahwa
jenis-jenis fitoplankton dari kelas Bacillariophyceae yang umumnya dijumpai
di perairan lepas pantai Indonesia antara lain Chaetoceros sp, Thallasiosira sp, dan Bacteriastrum sp, sedangkan dari kelas Dinophyceae yang umumnya dijumpai di laut adalah Noctiluca sp, Ceratium sp, Peridinium sp, dan Dinophysis sp.
Kelimpahan sel fitoplankton yang didapatkan selama pengamatan bervariasi di
kedua stasiun pengamatan. Secara keseluruhan kelimpahan sel fitoplanton lebih tinggi
di stasiun A dibandingkan dengan stasiun B (Tabel 7; Lampiran 9, 11, 12 dan 13).
Perbedaan tersebut disebabkan oleh posisi stasiun A yang masih mendapat pengaruh
perairan pantai yang mendapat suplai unsur hara dari aliran sungai di
sekitarnya.
Tabel 7. Rataan kelimpahan sel fitoplankton menurut stasiun pengamatan di perairan Teluk Banten
Kelimpahan fitoplankton (sel/l) Stasiun
Periode
ke - Bacillariophyceae Dinophyceae Cyanophyceae