Effects of beech and ash on small-scale variation of soil acidity and
nutrient stocks in a mixed deciduous forest
Frédéric M. Holzwarth1,2*, Max Daenner1,andHeiner Flessa3,4
1Department for Ecoinformatics, Biometrics, and Forest Growth, Büsgen Institute, Georg-August-Universität Göttingen, Büsgenweg 4,
37077 Göttingen, Germany
2present address: Department for Systematic Botany and Functional Biodiversity Research, Institute of Biology, Universität Leipzig,
Johannisallee 21, 04103 Leipzig, Germany
3Soil Science of Temperate and Boreal Ecosystems, Büsgen Institute, Georg-August-Universität Göttingen, Büsgenweg 2, 37077 Göttingen,
Germany
4present address: Institute of Agricultural Climate Research, Johann Heinrich von Thünen-Institut, Federal Research Institute for Rural Areas,
Forestry and Fisheries, Bundesallee 50, 38116 Braunschweig, Germany
Abstract
Trees interact in a complex manner with soils: they recycle and redistribute nutrientsviamany ecolo-gical pathways. Nutrient distributionvialeaf litter is assumed to be of major importance. Beech is commonly known to have lower nutrient concentrations in its litter than other hardwood tree species occurring in Central Europe. We examined the influences of distribution of beech (Fagus sylvaticaL.), ash (Fraxinus excelsiorL.), lime (Tilia cordataMill. andT. platyphyllosScop.), maple (Acer spp.L.), and clay content on small-scale variability of pH and exchangeable Ca and Mg stocks in the mineral soil and of organic-C stocks in the forest floor in a near-natural, mature mixed deciduous forest in Central Germany. The soil is a Luvisol developed in loess over limestone. We found a positive effect of the proportion of beech on the organic-C stocks in the forest floor and a negative effect on soil pH and exchangeable Ca and Mg in the upper mineral soil (0 to 10 cm). The proportion of ash had a similar effect in the opposite direction, the other species did not show any such effect. The ecological impact of beech and ash on soil properties at a sample point was explained best by their respective proportion within a radius of 9 to 11 m. The propor-tion of the species based on tree volume within this radius was the best proxy to explain species effects. The clay content had a significant positive influence on soil pH and exchangeable Ca and Mg with similar effect sizes.
Our results indicate that beech, in comparison to other co-occurring deciduous tree species, mainly ash, increased acidification at our site. This effect occurred on a small spatial scale and was probably driven by species-related differences in nutrient cyclingvialeaf litter. The distribu-tion of beech and ash resulted not only in aboveground diversity of stand structures but also induced a distinct belowground diversity of the soil habitat.
Key words:tree–soil interaction / species-specific effect / leaf litter / deciduous-tree species / forest floor / exchangeable cations
Accepted March 29, 2011
1 Introduction
Tree species differ in their influence on the acidification and the redistribution of nutrients in soils (see reviews byBinkley
andGiardina, 1998;Augustoet al., 2002). Many studies have analyzed the differences between conifer and broadleaved species (e.g., Sanborn, 2001; Rothe et al., 2002; Binkley, 2003; Rothe et al., 2003). A smaller number have investi-gated broadleaf species (e.g., Nordén, 1994a, b;Finziet al., 1998a, b; Neyrinck et al., 2000;Hagen-Thorn et al., 2004;
Oostraet al., 2006), and only very few studies have studied species effects along a gradient of species mixture (Klemmedson, 1991; Rothe, 1997; Sanborn, 2001; Rothe
et al., 2002).
One approach for treating small-scale heterogeneity and establishing a gradient of species admixtures is garden
experiments (Challinor, 1968; Binkley and Valentine, 1991;
Scherer-Lorenzen et al., 2007). However, studying estab-lished forests may also reveal and describe mechanisms of tree–soil interactions in detail. Although near-natural forests might contain greater noise in the data, they are more repre-sentative of natural ecosystems and also cover long-term effects (BinkleyandMenyailo, 2005;Leuschneret al., 2009).
Although a multitude of pathways on how trees interact with soils has been described, litterfall is commonly thought to be of major importance (Ovington, 1953;Challinor, 1968;Finzi
et al., 1998a, b;WashburnandArthur, 2003). Other factors, which were found to contribute to differences in deciduous tree species, are, e.g., canopy interception (throughfall), canopy–precipitation interaction, stem flow, rooting patterns,
root litter (Falkengren-Grerup, 1989;Nordén, 1991;Dijkstra
and Smits, 2002; Dijkstra, 2003; Meinen, 2009). Horizontal ranges of litter fall have been studied (Ferrari and Sugita, 1996;Staelens et al., 2003, 2004), but horizontal ranges of tree effects on soils properties have been investigated less (Zinke, 1962;Rothe, 1997;Rotheet al., 2002;Hojjatiet al., 2009).
Gucklandet al. (2009) analyzed mixed deciduous forests on a plot basis indicating differences in species-specific tree–soil interactions, as well as clay content in the soil being an impor-tant covariable. The results suggested that beech affected the soil nutrient status in a different manner than ash, lime, maple, hornbeam, and elm. However, it was not possible to quantify and separate effects of tree species and variations of soil clay content.Nordén(1994c) andJacobet al. (2009) con-firmed that beech litter contains less Ca and Mg than litter of several other deciduous European tree species and ash has very low C : N ratios compared to other species especially beech, which has rather high ratios.
We used some of the study sites described byGucklandet al. (2009) for a more detailed analysis of the species effect on small-scale variation of soil properties in these mixed decid-uous stands. On the basis that beech furnishes more recalci-trant litter, which also contains less basic cations, and that lit-terfall shows spatial patterns, we hypothesize that higher pro-portions of beech result in (1) higher stocks of organic C in the forest floor, lead to (2) a stronger acidification and to smaller stocks of exchangeable base cations in the upper mineral soil, and (3) to spatial patterns of the studied soil parameters. The objectives of this study were (1) to deter-mine the effects of tree species, especially beech, on the stocks of organic C in the forest floor and the acidification of the mineral soil (pH value, stocks of exchangeable Ca and Mg cations), (2) to analyze the horizontal and vertical (soil depth) extension of species effects, and (3) to separate effects of tree species from effects related to small-scale var-iation of soil clay content.
2 Material and methods
2.1 Study site
The study was conducted in mature deciduous forest stands (trees aged 1 to≈250 y), which are located in the Hainich National Park (51°06′ N, 10°31′ E), Thuringia, Germany, at an elevation of≈350 m asl (mean annual temperature 7.5°C; mean annual precipitation 670 mm). The soil type at the study plots is a Luvisol (FAO, 2006) developed from loess, which is underlain by limestone. The loess cover is generally free of carbonates, and its thickness varies between 60 and 120 cm. However, the tree roots still extend into the calcareous sub-soil.
The stands had been managed as coppice-with-standards (timber trees above coppiced woodland, including all the recent tree species) until≈1900, after which they were gra-dually transformed into a beech-selection forest (no coppice). Management and harvesting ceased by the year 1965 (
But-ler-Manning, 2007). In December 1997, it became the core zone of the Hainich National Park.
Four study plots with an extension of roughly 54 m × 54 m (inclination
<
5%) were established in the NE part of the national park. Tree species on these plots were beech (Fagus sylvaticaL.), ash (Fraxinus excelsiorL.), lime (Tilia cordataMill. andT. platyphyllosScop.), hornbeam (Carpinus betulus
L.), maple (Acer pseudoplatanusL.,A. platanoidesL., andA. campestreL.), and elm (Ulmus glabraHuds.). All plots con-tained a fair mixture of beech and the other species.
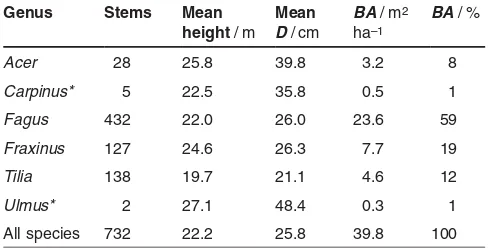
On each plot, all trees with a diameter at breast height (D) of at least 7 cm were recorded in the spring of 2005. Measured tree parameters were: coordinates of the trunk base, species, D, height, and crown projection area (C) (Tab. 1).
2.2 Sampling and laboratory analysis
For soil sampling within plots, a grid of 12 m × 12 m (aligned northward) was established within a radius of 20 m around the plot center (12 sample points per plot). In addition, a soil-profile pit was dug adjacent to each plot.
In the winter of 2004/05, soil cores (∅ 6.4 cm) were taken from the upper 30 cm of the mineral soil. They were divided into three parts representing the soil depths of 0–10 cm, 10–20 cm, and 20–30 cm. Samples were dried at 40°C and passed through a 2 mm sieve. Thickness of the loess cover was determined using a soil auger. Samples of the forest floor were collected at each sample point (300 cm2surface).
Soil pH was measured in a suspension with 1M KCl (5 g of soil, 15 mL KCl solution). Organic C (Corg) in the forest-floor samples was determined by an automated analyzer (Heraeus Elementar Vario EL, Hanau, Germany). Effective cation-exchange capacity (CEC) was quantified by leaching soil samples with 100 mL of 1M NH4Cl for 4 h. Cations in the extract were quantified by AAS. We determined the soil tex-ture using the sieving and pipette method (Schlichtinget al., 1995). Soil bulk density (after drying at 105°C) was calculated
2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.plant-soil.com
Table 1:Inventory data of all trees (diameter at breast height,D> 7 cm) on the four studied plots (total area 12 395 m2), shown for each
genus and in total: number of stems, mean height, mean diameter at breast height (D), basal area per hectare (BA), and proportion of basal area. Species marked with an asterisk occur only on one of the plots, respectively. All other species occur on all plots.
Genus Stems Mean
Carpinus* 5 22.5 35.8 0.5 1
Fagus 432 22.0 26.0 23.6 59
Fraxinus 127 24.6 26.3 7.7 19
Tilia 138 19.7 21.1 4.6 12
Ulmus* 2 27.1 48.4 0.3 1
gravimetrically from undisturbed soil cores taken from soil-profile pits adjacent to each plot (125 cm3, n = 3 each). Stocks of exchangeable cations in the mineral soil were cal-culated with the analytical value (mmolcg–1), the bulk density (g cm–3), and the thickness of the relevant layer (10 cm each). The properties of the stands and soils were described in detail byGucklandet al. (2009). Summary statistics of the soil parameters used in this study are given in Tab. 2.
2.3 Analysis of species effects
The soil-sample grid comprised 48 sample points (12 points at each of the four sites), 3 samples were missing, one soil sample (10 to 20 cm) missed only at the time of clay-content measurement, but we could impute the clay content using other measurements. We checked influence of stem flow, which would add to any spatial signal, by plotting the soil parameters against the distance to the nearest trunk, but did not find any trend and, thus, used samples from all 45 sample points in the analysis without further considering stem flow as an influential factor. We also checked the influence of abso-lute stand density in order not to confound species effects with mere density effects.
As dependent variables, we used the following soil para-meters: in the forest floor the stock of Corg(Mg ha–1) and in the upper mineral soil in both depth levels: the stocks of exchangeable cations of Ca and Mg (kmolc ha–1), and pH value (KCl).
Possible independent variables in this study were: the spe-cies proportion at a given point (Eq. 1), the clay content in the upper mineral soil (soil depths: 0–10 cm, 10–20 cm) and the absolute stand density (m2ha–1) in different radii around the sample point.
The potential influence of trees on soil properties at a given point in a mixed stand can be estimated from several indica-tors, among these: species identity, tree distance, tree dimen-sion, or other measurable proxies for the potential impact.
Rotheet al. (2002) proposed estimation of the influence of a tree species on soil properties of a given point from the
per-centage of the sum of the area of its crowns related to the sum of the crowns of all trees within a 10 m radius. We used a similar approach and additionally compared the suitability of four different attributes (projected crown area [C], diameter at breast height [D], basal area [BA], and tree volume [V]) for all major tree species (beech, ash, lime, and maple) as proxy. We calculated the proportion of a given species at a given sample point as the percentage of the sum of a species’ attri-butes related to the sum of all trees attriattri-butes (Eq. 1):
proportion of a species =
X
m i;r
j
Aij
X
n r
i
X
m i;r
j
Aij
; (1)
whereiis the tree species,Ais one of a set of measured attri-butes of each individual tree, andr the radius of the circular surrounding of the sample point ranging from 4.5 to 24 m with intervals of 0.5 m. Furthermore,m(i, r) is the number of trees of the speciesiwith the treesj= 1 tom(i, r) in a circle of the radiusr, n(r) the number of species in the relevant circle with the speciesi= 1 ton(r), andAijthe value of the attribute of treejof the speciesi. The attributeAof a tree (see above) shall represent its ecological footprint with regard to its influ-ence on the soil. For greater radii, parts of the circular sur-roundings would lie outside of the plot, where no tree data were available. However, as will be shown later, greater radii are not relevant for the analysis and, thus, a border correction was not necessary.
The basal area (BA) was calculated asa×D2, while the vol-ume (V) was approximated by V=a ×D2.6, witha being a species-specific parameter. The volume of a tree can be approximated by V = BA × height × form factor, and the height is typically estimated with height= a × Db. For the sake of simplicity, we assumed the form factor and the allo-metric constants to be equal among species, which we deem legitimate for our purpose and which is underpinned by height regressions done with the present data set, where heights were available for all trees (data not shown). While the factors would drop out in Eq. 1, the estimated exponent with all stu-died species pooled together was 2.6.
Table 2:Statistics (sample mean and standard deviation [SD] as well as minimum and maximum) of soil parameters measured in the analyzed stands at two soil depths (0–10 cm, 10–20 cm).
Parameter Mean (SD) Min. Max.
Corgin forest floor / Mg ha–1 3.2 (1.2) 0.7 5.9
Clay content / % 0–10 cm 24.7 (7) 14.4 41.6
10–20 cm 24.5 (7) 14.0 41.6
pH (KCl) 0–10 cm 4.4 (0.8) 3.5 6.3
10–20 cm 4.7 (0.8) 3.4 6.4
Ex.aCa / kmol
cha–1 0–10 cm 140 (86) 8.6 450
10–20 cm 150 (76) 3.2 330
Ex. Mg / kmolcha–1 0–10 cm 10 (4.7) 2.4 26
10–20 cm 8.9 (4.2) 1.5 21
2.4 Statistical analysis
Visualization of the data suggested a linear relationship be-tween the species proportions, the stand densities, as well as the clay content on the one side and the studied soil para-meters Corg, pH value, exchangeable Ca and Mg on the other side. For all regressions, we used linear mixed-effect models to account for the nested sampling design. Models were fit by maximizing the likelihood (ML). Effects in the models were chosen by comparing the models BIC (Bayesian information criterion). We refer to the null model as the model with only the random effects. Relating the model likelihood with the likelihood of the null model, served us as goodness of fit (Maddala pseudo-R2).
For Corgin the forest floor, possible fixed effects were: clay content (in the upper layer) and species proportion, and ran-dom effect: plot identity. For the other soil parameters (meas-urements in two depth layers), interaction of the species pro-portion with the depth layer was also possible and as random effects: plot identity and sample-point identity nested within plot identity. As the species proportion and the clay content did not correlate, we used them as independent variables, where applicable. To underline this, we also reported covar-iance of parameter estimates for these two variables in the models. To check model assumptions, we analyzed the resi-duals with Tukey-Anscombe and QQ plots.
The species proportions were of course highly correlated, so it is impossible to include all different proportions in a single model. Moreover, we assume only a single gradient of spe-cies abundance to be responsible for the majority of variation in the soil parameters caused by species identity. We did mul-tivariate analysis (PCA) with the species proportions to iden-tify the main axes of variation. The main axes as well as the four species proportions from all available calculations (depending on the species, the radius of the circular sur-rounding, and the attribute to represent the trees) were used as explanatory variables. We compared the pseudo-R2of the models between all possible proportions and the dependent soil parameter to find the proportion that best explains a given soil parameter. A similar approach (with the Pearson correla-tion coefficient) was described byRothe (1997) andRothe
et al. (2002).
We checked, whether distance signals in the models would be artifacts of the rather dense and regular sample grid. Therefore, we also calculated the models with subsets of the points: in each plot we selected six points, which had the furthest distance from each other (minimum distance not 12 m but 17 m). With two possible subsets for each plot and four plots there were 16 permutations, which we all calculated independently and finally checked, whether, on average, any previously found distance signal would change.
We used “R: A language and environment for statistical com-puting” (R Development Core Team, 2010) for all computa-tions, including the package “lme4: Linear mixed-effects mod-els using S4 classes” (Bateset al., 2011).
3 Results
3.1 Influence of covariables “clay content” and “stand density”
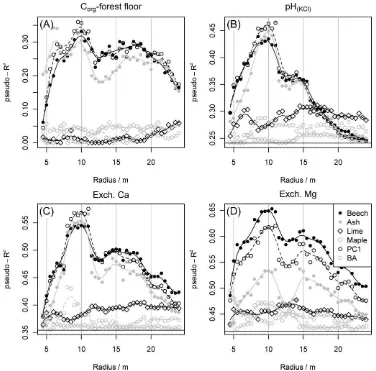
The clay content in the upper 10 cm of the mineral soil as an explaining factor accounted for a substantial degree of var-iance in stocks of exchangeable Ca and Mg (pseudo-R2 = 0.38 and 0.36). In contrast, its influence on the pH value of the mineral soil was quite low (pseudo-R2 = 0.15) and non-existent on Corgin the forest floor (Tab. 3). The clay content in the underlying layer (10–20 cm) explained the variation of the soil parameters (in the corresponding layer) even better with pseudo-R2for pH, Ca, and Mg being 0.26, 0.58, and 0.63, re-spectively. Hence, clay content was an important predictor for stocks of exchangeable Ca and Mg as well as the pH value in the mineral soil and was thus used as covariable in further models of these three parameters. Absolute stand densities (m2ha–1) in increasing radii around the sample points did not show any explanatory effect nor any trend with the radius (analogous: also no correlation with litter mass in litter traps [15 traps per plot], data not shown) and thus allowed the use of species proportions instead of absolute values (Fig. 2).
3.2 Influence of species proportions and their horizontal ranges
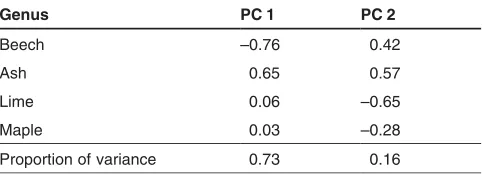
Figure 1 and Tab. 4 give the results of the PCA of the species proportions as an example for one radius (10 m) and one attribute (V). It reveals a rather strict dichotomy of beechvs.
ash on the first axis, covering 73% of the variation. The sec-ond axis spans beech and ashvs.lime and maple (16%), and
2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.plant-soil.com
Table 3:Goodness of fit (pseudo-R2) of mixed models with clay
con-tent in respective depth layer as fixed effect.
Soil parameter
Table 4:Statistics of the first two axes of a PCA on species propor-tions calculated with tree volume (V) as attribute in a radius of 10 m. Loadings of species proportions on respective axis and their propor-tion of explained variance.
the third axis (not shown in Fig. 1) differentiates between lime and maple (10%).
We show all pseudo-R2of models using the four species pro-portions and the first axis of the PCA in all different radii as explanatory variable (Fig. 2). We only present results of mod-els using species proportions based on the attribute tree vol-ume (V), because projected crown area (C), diameter at breast height (D), and basal area (BA) proved to convey less explanatory power (withBAbeing significantly better thanC
and D and only slightly worse than V), and we shall not regard the latter anymore.
Models using species proportions of beech and ash as well as the first PCA axis exhibited a significant explanatory power in radii between 9 and 11 m for all modeled parameters, except for Mg, where the proportion of ash did not perform well. We observed a distinct change of the goodness of fit with changing radius (steep increase of the model pseudo-R2 with increasing radius and then smooth decline), which clearly indicates a nonrandom correlation. Otherwise, a more erratic development of the pseudo-R2with increasing radius would have been observed. This trend was strongly pro-nounced for the models of pH, Ca, and Mg. Proportions of lime and maple as well as the second and third axis of the PCA explained far less variation of the soil parameters and had no consistent trend. The spatial aspect of the sampling grid had no influence on the estimated ranges. On average, the models with 16 different subsets of points showed the Figure 1:Bi-Plot of the first two axes of a PCA on species proportions
calculated with tree volume (V) as attribute in a radius of 10 m. Points represent the observations in the reduced space, vectors represent variable loadings on the two axes.
Figure 2: Goodness of fit (pseudo-R2) of mixed-effect
models regressing Corg in the forest floor (A) as well as pH values (B), exchangeable Ca (C), and Mg (D) stocks in two layers of the mineral soil on species proportions (beech, ash, lime, and maple) calculated on the basis of estimated tree volume (V) as well as on the first PCA axis (PC1, Fig. 1) and on stand densities (m2ha–1) (basal
same distance trend. This corroborates our finding of a spa-tial signal in the species proportions and precludes that it is a mere sampling artifact.
We selected the species proportions and the radii, that entailed high pseudo-R2 (beech for all soil parameters, ash for Corg, pH, and Ca; in radii of 9 to 11 m) for candidate mod-els, on which we had a closer look. The proportions were averaged for the radii of 9 to 11 m, since the respective mod-els had very similar pseudo-R2 for these radii. We did not choose to use both proportions in a model, because this would lead to high intercorrelations and thus a great uncer-tainty in parameter estimations. We also did not choose to use the first PCA axis, although it sometimes performed bet-ter than the mere species proportions. First, because the PCA axis contained information from four species proportions and thus, naturally has the potential to perform better than a single proportion. And second, because the difference in goodness of fit was not large enough to prefer it over simpler and more intuitive predictors.
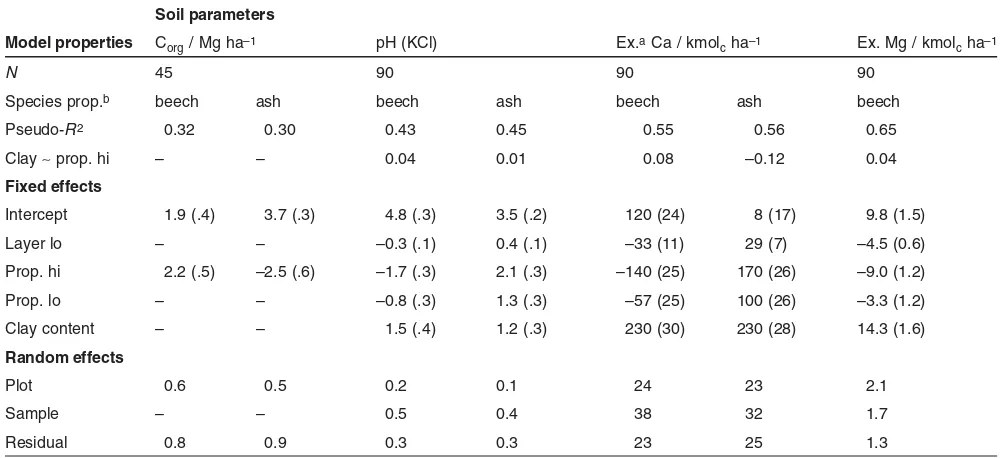
The candidate models are summarized in Tab. 5. The fixed effect parameter “clay content” was normalized to an interval of [0, 1], to better comprehend and compare the effect sizes. In the models, where both fixed effects were included (pH, Ca, and Mg), the parameter estimates for the species propor-tion and the clay content were not or only very weakly corre-lated. No major deviations from the model assumptions (homoscedasticity, normally distributed residuals) were found in any of the models. There was a slight trend of higher var-iance of the residuals with increasing estimates for Ca, but
we did not deem it strong enough and thus necessary to counter it with,e.g.,a transformation of the dependent vari-able.
The stocks of organic C in the forest floor (Corg) (Mg ha–1) could be explained by the proportion of beech (pseudo-R2 0.32) equally well as by the proportion of ash (0.30), and no influence of the clay content was found (Tab. 5). A change from 0% to 100% beech proportion resulted in an estimated increase of the Corgstocks in the forest floor of more than 100% (1.9 to 4.1 Mg ha–1). Alternatively, a decrease from 100% ash proportion to 0% predicted a similar increase (1.3 to 3.7 Mg ha–1). The uncertainties of the parameter estimates were considerable (0.5 Mg ha–1) (Tab. 5).
The variation of the pH value in the upper 20 cm of the mineral soil could be explained to a certain degree by a model comprising the proportion of beech (pseudo-R20.43) or ash (0.45) and the clay content (Tab. 5). Species-propor-tion effects were strong in the upper layer (beech: –1.7, ash: 2.1 pH units) and noticeably weaker in the lower layer (beech: –0.8, ash: 1.3 pH units). The effect of the clay content was estimated to be of similar magnitude (1.5 and 1.2 pH units, respectively).
Variation of exchangeable Ca (kmolcha–1) (0 to 20 cm) was explained to an even greater extent with a pseudo-R2of 0.55 (beech) or 0.56 (ash) and clay content. Of greatest impor-tance was clay content (230 kmolcha–1in both models), fol-lowed by the species proportion with an effect of –140 (beech) or 170 (ash) in the upper 10 cm of the mineral soil
2011 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.plant-soil.com
Table 5:Candidate models. Model properties: sample size (N), attribute and radius of the proportion of beech used, goodness of fit in relation to null model (Maddala pseudo-R2), and correlation of parameter estimates for clay content and species proportion (Clay∼prop. hi; in the upper layer [hi]; values for the lower layer [lo] are very similar). Fixed effects: parameter estimates (±SE) for lower depth layer (10–20 cm), clay content (normalized), and species proportion. Parameters for species proportion differ in the two depth layers (upper = 0–10 cm, lower = 10–20 cm). The parameter values also resemble the respective effect size for the variation from minimum to maximum values of each variable. Random effects: variance attributed to random effects and residual variance, given as standard deviations.
Soil parameters
Model properties Corg/ Mg ha–1 pH (KCl) Ex.aCa / kmol
cha–1 Ex. Mg / kmolcha–1
N 45 90 90 90
Species prop.b beech ash beech ash beech ash beech
Pseudo-R2 0.32 0.30 0.43 0.45 0.55 0.56 0.65
Clay∼prop. hi – – 0.04 0.01 0.08 –0.12 0.04
Fixed effects
Intercept 1.9 (.4) 3.7 (.3) 4.8 (.3) 3.5 (.2) 120 (24) 8 (17) 9.8 (1.5)
Layer lo – – –0.3 (.1) 0.4 (.1) –33 (11) 29 (7) –4.5 (0.6)
Prop. hi 2.2 (.5) –2.5 (.6) –1.7 (.3) 2.1 (.3) –140 (25) 170 (26) –9.0 (1.2)
Prop. lo – – –0.8 (.3) 1.3 (.3) –57 (25) 100 (26) –3.3 (1.2)
Clay content – – 1.5 (.4) 1.2 (.3) 230 (30) 230 (28) 14.3 (1.6)
Random effects
Plot 0.6 0.5 0.2 0.1 24 23 2.1
Sample – – 0.5 0.4 38 32 1.7
Residual 0.8 0.9 0.3 0.3 23 25 1.3
and mere –57 (beech) or 100 (ash) one layer deeper (Tab. 5). The species effect weakened considerably with increasing soil depth.
Stocks of exchangeable Mg (kmolcha–1) in the upper mineral soil could be modeled by clay content and proportion of beech with a high pseudo-R2 of 0.65 (Tab. 5). Again, clay content had the strongest influence, leading to an estimated shift of up to 14.3 kmolcha–1. Changes of –9.0 in the upper layer and just –3.3 in the lower layer were attributed to an increase of beech proportion from 0% to 100%.
4 Discussion
As we hypothesized at the outset, the occurrence of beech in the investigated mixed deciduous forest resulted in higher C stocks in the forest floor and lower stocks of exchangeable Ca and Mg as well as lower pH values in the upper mineral soil. However, the occurrence of ash was negatively corre-lated with the occurrence of beech and the proportion of ash had a similar explanatory power with opposite directions for the first three parameters. We could hardly separate the effect of these two species by our methods alone. We argue that in the studied stands both species act antagonistically and the relative importance of these two species varied with the soil parameter. While for stocks of exchangeable Mg, beech stood against all other species and ash did not show a remarkably different behavior than lime and maple, for stocks of Corg, exchangeable Ca, and the pH value, beech and ash formed a dichotomy, where lime and maple stood in between.
The effect of species identity interfered with the influence of clay content, which was of opposite direction (except for the forest-floor C stock, which was not influenced by clay con-tent). Our results support the conclusion byGucklandet al. (2009) that soil acidification and soil nutrient distribution in these stands are affected by the abundance of beech and add to it that ash stands out of the other deciduous species and spans the other end of the gradient. Their conclusion was constrained by the interfering effects of species abun-dance and soil clay content, which could not be separated, as well as the coarse resolution of the species distribution on just the whole plot size. Our methods allowed a clear quanti-tative separation of these effects and a small-scale resolution of the species distribution. Our results showed that in the analyzed stands the proportions of beech and ash had a sim-ilar effect magnitude on soil pH and exchangeable Ca and Mg in the upper 10 cm of the mineral soil as the soil clay con-tent.
The influence of tree species diminished with soil depth. Sim-ilar results were described for beech and spruce (Rotheet al., 2002), and some genera also used in this study (Acer sp., Fagus sp., Fraxinus sp. among others,Finzi et al., 1998a). The weakening of the effect with soil depth indicates that the analyzed soil properties were probably influenced by an aboveground process, most probably leaf litterfall. Analyses of leaf litter from all occurring species clearly showed a differ-ence in cation content between beech, on the one hand, and the remaining species on the other (Nordén, 1994c).Jacob
et al. (2009) analyzed the leaf-litter composition of the
investi-gated stands: beech litter had lower contents of Ca and Mg than the nonbeech litter (16.8vs.21.5 to 30.2, 1.2vs.2.1 to 3.2 [mg (g dry mass)–1]). This resulted in an increased annual deposition of Ca, Mg, and alkalinityvialeaf litter with decreas-ing abundance of beech (Gucklandet al., 2009).
Our results support the observations of other studies that species-related differences in nutrient recyclingvialitter can have significant implications for the pattern of soil fertility and soil acidity in mixed stands (Nordén, 1994b; Finzi et al., 1998a;RotheandBinkley, 2001;Sayer, 2006).Augustoet al. (2002) summarized effects of tree species in European tem-perate forests on soil fertility. They concluded that beech and oak species were the deciduous species, which exerted the greatest acidifying effect on soils. At our site, the redistribu-tion of Ca and Mg in the soil profilevianutrient uptake and lit-ter deposition had a highly beneficial effect since it led to the translocation of base cations and alkalinity from the alkaline subsoil (limestone) to the surface parent material (loess), which has rather low buffer capacity and tends to form strongly acidic forest soils (Guckland et al., 2009). Differ-ences of cation contents in fine-root litter were not observed (C. Meinen, pers. communication), which excludes fine-root litter as the cause of the observed differences.
The C stock in the forest floor was found to be independent of the underlying clay content but depended on the species pro-portion. The accumulation of litter on the mineral soil is mainly a function of decay rates, not of litter production (Guckland
et al., 2009). Increasing litter accumulation with increasing abundance of beech can be explained by the relatively high recalcitrance of beech litter, which has lower nutrient concen-trations and a higher C : N ratio than litter ofAcerand Fraxi-nusspecies (Melliloet al., 1982;Finzi et al., 1998b).Jacob
et al. (2009) reported that beech leaf litter at our sites had higher C : N ratios than all other occurring species (means: 56vs.28 to 40), while ash marked the lower end. In addition, beech litter may affect the activity of decomposers due to its influence on soil pH and soil-nutrient availability (Reichet al., 2005;Jacobet al., 2009). Our observations on beech effects on the forest floor agree with the conclusions of Neirynck
et al. (2000) that beech belongs to the mullmoder-forming species, whereas maple, lime, and ash are mull-forming trees.
Soil properties at a given point in this mixed stand were influ-enced by stand structure and species composition within a radius of 9 to 11 m. This is in the near range of the radius around litter traps, in which the proportion of beech was cor-related to the proportion of beech litter in the traps (13 to 16 m;
Holzwarth, 2008). Similar radii (11 to 18 m) for litter dispersal of deciduous trees in mature stands were found byStaelens
et al. (2004), and Ferrari and Sugita (1996) modeled that 50% of the leaf litter would fall down in distances of 5 to 13 m from the stems. Nevertheless, litter dispersal of different deciduous tree species depends on the canopy structure, the form of falling leaves and the wind regime (Hirabuki, 1991;
autumnal rainfall and moisture surplus during winter months, decreased wind velocities in a closed forest, due to the understory vegetation (Guckland et al., 2009), and fungal hyphae binding fresh litter to the forest floor (Ferguson, 1985;
LodgeandAsbury, 1988;Hirabuki, 1991).
These factors may thus affect the horizontal extension of tree-species effects on soil chemical properties. The results of studies on litter dispersal (Staelens et al., 2004; Ferrari
andSugita, 1996;Rothe, 1997;Staelenset al., 2003), on the range of species-specific effects on soil properties (Rothe, 1997; Rothe et al., 2002; this study), on litter chemistry (Jacob et al., 2009), as well as on effects of litter on soil chemistry (Ferguson, 1985; Binkley and Valentine, 1991;
Sayer, 2006) show that distribution of tree litter is an impor-tant factor, which causes small-scale variability of chemical soil properties in mixed stands. The negative correlation of the occurrence of beech and ash found on the plots might be the reason for their similar explanatory power in the models, while underneath there is still a dichotomy between beech and the rest of the species lumped together, as indicated by litter chemistry (see above andJacobet al., 2009). More and larger plots with different species admixtures could have facilitated a clear separation of species effects.
We also stress the need to consider confounding soil factors (e.g.,texture) in this research area. In our study, the species effect was nearly as strong as or slightly stronger than the effect of clay content, thus species identities can be as impor-tant as the parent material for soil development. This under-lines the responsibility that forest management has not only for a sustainable yield of wood and other ecosystem services, but also for sustainable soil fertility. Our results also indicate that small-scale variability of chemical soil properties is not only driven by species mixture and identity, but also by the spatial pattern of individual trees. On the one hand, this can be applied in forestry planting to help regulating soil chem-istry, but on the other hand, it needs to be carefully consid-ered within biodiversity studies.
Even though we were able to explain a considerable part of the small-scale variability of C accumulation in the forest floor and soil acidity in the upper mineral soil, our models left sub-stantial parts of the variation in these properties unexplained. This variation might be explained by other covariables,e.g.,
multidimensional representation of species proportions, intraspecific variation of litter quality, interspecific or interindi-vidual variation of horizontal extension of litterfall (Staelens
et al., 2003), management history (Hüttland Schaaf, 1995;
Johnsonand Curtis, 2001), or rooting patterns (Rothe and
Binkley, 2001). Yet, it is also common to encounter small-scale heterogeneity of random nature in near-natural ecosys-tems (Jackson and Caldwell, 1993; Ettema and Wardle, 2002) and a number of unknown or uncontrolled sources of influence (Ehrenfeldet al., 2005). Unfortunately, no garden experiments of similar extent, age, and species composition are available thus far (Leuschneret al., 2009), which calls for continued research in near-natural ecosystems (Ehrenfeld
et al., 2005;Leuschneret al., 2009).
The published results on litter quality in the studied stands (Jacobet al., 2009) and the vertical and horizontal ranges of the effects of the proportion of beech found in this study indi-cate that the main cause of species-specific effects on soil parameters mentioned here are differences in leaf-litter chemistry. In addition, our results show that the distribution of beech and ash resulted not only in aboveground diversity of stand structures but also caused a distinct small-scale below-ground diversity of the soil habitat.
Acknowledgments
Funding by the Deutsche Forschungsgemeinschaft (DFG; Graduiertenkolleg 1086)is gratefully acknowledged. We want to thankAnja GucklandandMascha Jacobfor sharing their data with us. The authors are grateful to an anonymous referee for many useful suggestions.
References
Augusto, L., Ranger, J., Binkley, D., Rothe, A. (2002): Impact of several common tree species of European temperate forests on soil fertility.Ann. Forest Sci.59, 233–253.
Bates, D., Mächler, M., Bolker B.(2011): lme4: Linear mixed-effects models using S4 classes. Available at: http://CRAN.R-project.org/ package=lme4
Binkley, D.(2003): Seven decades of stand development in mixed and pure stands of conifers and nitrogen-fixing red alder. Can. J. Forest Res.33, 2274–2279.
Binkley, D., Giardina, C. (1998): Why do tree species affect soils? The warp and woof of tree-soil interactions.Biogeochemistry42, 89–106.
Binkley, D., Menyailo, O. (2005): Gaining Insights on the Effects of Tree Species on Soils, in Binkley, D., Menyailo, O. (eds.): Tree Species Effects on Soils: Implications for Global Change. Springer Verlag, Dordrecht, The Netherlands, pp. 1–16.
Binkley, D., Valentine, D.(1991): Fifty-year biogeochemical effects of green ash, white pine, and Norway spruce in a replicated experiment.Forest Ecol. Manage.40, 13–25.
Butler-Manning, D.(2007): Stand structure, gap dynamics and regen-eration of a semi-natural mixed beech forest on limestone in central Europe – a case study. Schriftenreihe Freiburger Forstliche Forschung 38, University Freiburg, Freiburg im Breisgau, Germany.
Challinor, D.(1968): Alteration of surface soil characteristics by four tree species.Ecology49, 286–290.
Dijkstra, F. A.(2003): Calcium mineralization in the forest floor and surface soil beneath different tree species in the northeastern US. Forest Ecol. Manage.175, 185–194.
Dijkstra, F. A., Smits, M. M.(2002): Tree species effects on calcium cycling: The role of calcium uptake in deep soils.Ecosystems5, 385–398.
Ehrenfeld, J. G., Ravit, B., Elgersma, K. (2005): Feedback in the plant-soil system.Annu. Rev. Environ. Resour.30, 75–115.
Ettema, C. H., Wardle, D. A. (2002): Spatial soil ecology. Trends Ecol. Evol.17, 177–183.
Falkengren-Grerup, U. (1989): Effect of stemflow on beech forest soils and vegetation in southern Sweden.J. Appl. Ecol.(UK) 26, 341–352.
FAO(2006): World Reference Base for Soil Resources. Roma, Italy.
Ferguson, D. K.(1985): The origin of leaf-assemblages – new light on an old problem. Rogier Vanhoorne – An appreciation. Rev. Palaeobotany Palynology46, 117–188.
Ferrari, J. B., Sugita, S.(1996): A spatially explicit model of leaf litter fall in hemlock-hardwood forests. Can. J. Forest Res. 26, 1905–1913.
Finzi, A. C., Canham, C. D., van Breemen, N.(1998a): Canopy tree-soil interactions within temperate forests: Species effects on pH and cations.Ecol. Applic.8, 447–454.
Finzi, A. C., van Breemen, N., Canham, C. D.(1998b): Canopy tree-soil interactions within temperate forests: Species effects on tree-soil carbon and nitrogen.Ecol. Appl.8, 440–446.
Guckland, A., Jacob, M., Flessa, H., Thomas, F. M., Leuschner, C. (2009): Acidity, nutrient stocks, and organic-matter content in soils of a temperate deciduous forest with different abundance of European beech (Fagus sylvaticaL.).J. Plant Nutr. Soil Sci.172, 500–511.
Hagen-Thorn, A., Callesen, I., Armolaitis, K., Nihlgård, B.(2004): The impact of six European tree species on the chemistry of mineral topsoil in forest plantations on former agricultural land. Forest Ecol. Manage.195, 373–384.
Hirabuki, Y.(1991): Heterogeneous dispersal of tree litterfall corre-sponding with patchy canopy structure in a temperate mixed forest.Plant Ecol.94, 69–79.
Hojjati, S., Hagen-Thorn, A., Lamersdorf, N.(2009): Canopy compo-sition as a measure to identify patterns of nutrient input in a mixed European beech and Norway spruce forest in central Europe.Eur. J. Forest Res.128, 13–25.
Holzwarth, F. M.(2008): Variability of the nutrient regime of soils in the National Park Hainich: Influence of the proportion of beech and the soil texture. Master thesis, Georg-August-Universität, Göttingen, Germany, available at: http://www.uni-goettingen.de/de/ document/download/7a8555f62b56a0c3deb97c8bc176d500.pdf/ Masterarbeit_Frederic%20Holzwarth.pdf
Hüttl, R. F., Schaaf, W. (1995): Nutrient supply of forest soils in relation to management and site history. Plant Soil 168–169, 31–41.
Jackson, R. B., Caldwell, M. M.(1993): The scale of nutrient hetero-geneity around individual plants and its quantification with geosta-tistics.Ecology74, 612–614.
Jacob, M., Weland, N., Platner, C., Schaefer, M., Leuschner, C., Thomas, F. M. (2009): Nutrient release from decomposing leaf litter of temperate deciduous forest trees along a gradient of increasing tree species diversity. Soil Biol. Biochem. 41, 2122–2130.
Johnson, D. W., Curtis, P. S.(2001): Effects of forest management on soil C and N storage: meta analysis.Forest Ecol. Manage.140, 227–238.
Klemmedson, J. O.(1991): Oak influence on nutrient availability in pine forests of Central Arizona.Soil Sci. Soc. Am. J.55, 248
Leuschner, C., Jungkunst, H. F., Fleck, S.(2009): Functional role of forest diversity: Pros and cons of synthetic stands and across-site comparisons in established forests.Basic Appl. Ecol.10, 1–9.
Lodge, D., Asbury, C. (1988). Basidiomycetes reduce export of organic matter from forest slopes.Mycologia80, 888–890.
Meinen, C., Hertel, D., Leuschner, C. (2009): Biomass and
morphology of fine roots in temperate broad-leaved forests differing in tree species diversity: is there evidence of below-ground overyielding?Oecologia161, 99–111.
Melillo, J. M., Aber, J. D., Muratore, J. F.(1982): Nitrogen and lignin control of hardwood leaf litter decomposition dynamics.Ecology 63, 621–626.
Neirynck, J., Mirtcheva, S., Sioen, G., Lust, N.(2000): Impact ofTilia platyphyllosScop.,Fraxinus excelsiorL.,Acer pseudoplatanusL., Quercus roburL. andFagus sylvaticaL. on earthworm biomass and physico-chemical properties of a loamy topsoil.Forest Ecol. Manage.133, 275–286.
Nordén, U.(1991): Acid deposition and throughfall fluxes of elements as related to tree species in deciduous forests of South Sweden. Water, Air Soil Poll.60, 209–230.
Nordén, U.(1994a): Influence of broad-leaved tree species on pH and organic matter content of forest topsoils in Scania, south Sweden.Scand. J. Forest Res.9, 1–8.
Nordén, U.(1994b): Influence of tree species on acidification and mineral pools in deciduous forest soils of South Sweden.Water, Air Soil Poll.76, 363–381.
Nordén, U. (1994c): Leaf litterfall concentrations and fluxes of elements in deciduous tree species. Scand. J. Forest Res. 9, 9–16.
Oostra, S., Majdi, H., Olsson, M.(2006): Impact of tree species on soil carbon stocks and soil acidity in southern Sweden. Scand. J. Forest Res.21, 364–371.
Ovington, J. D. (1953): Studies of the development of woodland conditions under different trees: soils pH.J. Ecol.41, 13–34.
R Development Core Team(2010): R: A Language and Environment for Statistical Computing. Wien, Austria, available at: http://www.R-project.org.
Reich, P. B., Oleksyn, J., Modrzynski, J., Mrozinski, P., Hobbie, S. E., Eissenstat, D. M., Chorover, J., Chadwick, O. A., Hale, C. M., Tjoelker, M. G.(2005): Linking litter calcium, earthworms and soil properties: a common garden test with 14 tree species.Ecol. Lett. 8, 811–818.
Rothe, A. (1997): Einfluss des Baumartenanteils auf
Durch-wurzelung, Wasserhaushalt, Stoffhaushalt und Zuwachsleistung eines Fichten-Buchen-Mischbestandes am Standort Höglwald. Forstliche Forschungsberichte München 163, Technical University München.
Rothe, A., Binkley, D.(2001): Nutritional interactions in mixed species forests: a synthesis.Can. J. Forest Res.31, 1855–1870.
Rothe, A., Kreutzer, K., Küchenhoff, H. (2002): Influence of tree species composition on soil and soil solution properties in two mixed spruce-beech stands with contrasting history in Southern Germany.Plant Soil240, 47–56.
Rothe, A., Ewald, J., Hibbs, D. E.(2003): Do admixed broadleaves improve foliar nutrient status of conifer tree crops? Forest Ecol. Manage.172, 327–338.
Sanborn, P. (2001): Influence of broadleaf trees on soil chemical properties: A retrospective study in the Sub-Boreal Spruce Zone, British Columbia, Canada.Plant Soil236, 75–82.
Sayer, E. J.(2006): Using experimental manipulation to assess the roles of leaf litter in the functioning of forest ecosystems.Biol. Rev. 81, 1–31.
Scherer-Lorenzen, M., Schulze, E.-D., Don, A., Schumacher, J., Weller, E. (2007): Exploring the functional significance of forest diversity: A new long-term experiment with temperate tree species (BIOTREE).Persp. Plant Ecol., Evol. Syst.9, 53–70.
Schlichting, E., Blume, H.-P., Stahr, K. (1995): Bodenkundliches Praktikum. 2nd edn., Blackwell Wissenschafts-Verlag, Wien, Austria.
Staelens, J., Nachtergale, L., Luyssaert, S. (2004): Predicting the spatial distribution of leaf litterfall in a mixed deciduous forest. Forest Sci.50, 836–847.
Washburn, C. S., Arthur, M. A. (2003): Spatial variability in soil nutrient availability in an oak-pine forest: potential effects of tree species.Can. J. Forest Res.33, 2321–2330.
Welbourn, M. L., Stone, E. L., Lassoie, J. P.(1981): Distribution of net litter inputs with respect to slope position and wind Direction. Forest Sci.27, 651–659.
Wilke, B., Bogenrieder, A., Wilmanns, O.(1993): Differenzierte Streu-verteilung im Walde, ihre Ursachen und Folgen.Phytocoenologia 23, 129–155.
Zinke, P. J.(1962): The pattern of influence of individual forest trees on soil properties.Ecology43, 130–133.
![Table 2: Statistics (sample mean and standard deviation [SD] as well as minimum and maximum) of soil parameters measured in the analyzedstands at two soil depths (0–10cm, 10–20cm).](https://thumb-ap.123doks.com/thumbv2/123dok/2116865.1609437/3.595.56.553.566.710/statistics-standard-deviation-minimum-maximum-parameters-measured-analyzedstands.webp)