PERTUMBUHAN DAN PERKEMBANGAN SEL
TUMBUHAN
Untuk Memenuhi Tugas Matakuliah Fisiologi Tumbuhan
Yang dibina oleh Ibu Prof. Dra. Herawati Susilo, M. Sc., Ph. D Disusun oleh :
Kelompok 11 Offering B 2016
1. Amanda Valentina Santoso NIM. 160341606043
2. Mawaddatul Khasanah NIM. 160341606058
3. Merinda Oktaviana NIM. 160341606002
Disajikan pada, 15 November 2017
UNIVERSITAS NEGERI MALANG
FAKULTAS MATEMATIKA DAN PENGETAHUAN ALAM JURUSAN BIOLOGI
ii DAFTAR ISI
DAFTAR ISI ………. ii
DAFTAR GAMBAR... iii
ABSTRAK………. iv BAB I PENDAHULUAN 1.1 Latar Belakang………... 1 1.2 Rumusan Masalah……… 1 1.3 Tujuan…………..……….... 2 1.4 Manfaat ………... 2 BAB II BAB III TINJAUAN PUSTAKA 2.1 Pengertian penuaan ... 2.2 Pengertian program kematian sel... 2.3 Pengertian pembelahan sel... 2.4 Pengertian siklus sel... 2.5 Pengertian persepsi sinyal... 2.6 Pengertian transduksi sinyal... 2.7 Pengertian respon sinyal... PEMBAHASAN 3 3 3 3 3 3 4 3.1 Penuaan dan program kematian sel ………..…………... 5
3.2 Mekanisme pembesaran dan pemanjangan sel tumbuhan....………….... 12
3.3 Mekanisme persepsi dan transduksi signal... 22
BAB IV PENUTUP 4.1 Simpulan……….. 28
4.2 Saran………. 28
iii DAFTAR GAMBAR
Gambar 3.1.2. Grafik sigmoid pada penuaan sel tumbuhan... 9 Gambar 3.1.4. Siklus hidup tumbuhan ... 11 Gambar 3.2.3 The phragmoplast dan plat sel dalam sel tanaman
pemisah...
14
Gambar 3.2.2. Fase siklus sel... 15
Gambar 3.2.4. Plasmodesmata... 16
Gambar 3.2.5 Mikrotubulus dan ratapan mikrofil 19
Gamabr 3.2.6. Sebuah model untuk relaksasi stress di dinding sel
tumbuhan... 22 Gambar 3.3.1. Kalsium sebagai secondary
message... 24 Gambar 3.3.2. Transduksi sinyal pada sistem reseptor G-Protein ... 27
iv ABSTRAK
Santoso A. V, Hasanah Mawaddatul, Oktaviana Merinda. 2017. Pertumbuhan dan Perkembangan Sel Tumbuhan.Makalah Fisiologi Tumbuhan, Offering B S1 Pendidikan Biologi Universitas Negeri Malang. Dosen Pembimbing : Prof. Dra. Herawati Susilo, M. Sc., Ph. D . E-mail: [email protected]
Sel adalah unit dasar kehidupan Pertumbuhan dan perkembangan sel-sel tersebut diatur oleh berbagai sinyal internal dan eksternal. Seperti hormon, cahaya, suhu, gravitasi, predasi serangga, penyakit dan hubungan sel dengan sel lainnya. Dalam mengetahui perkembangan pada sel tumbuhan diperlukan pengetahuan mengenai dinding pada sel tumbuhan, pembelahan sel serta sinyal yang memengaruhi peristiwa biologi pada pertumbuhan dan perkembangan tumbuhan. Tujuan pembuatan makalah adalah untuk mengetahui proses penuaan dan program kematian sel tumbuhan, mekanisme pembesaran dan pemanjangan sel tumbuhan dan mekanisme persepsi dan transduksi signal dalam proses perkembangan tumbuhan.
Kata kunci : sel tumbuhan, perkembangan, pertumbuhan
ABSTRACT
Santoso A. V., Mawaddatul, Merinda Oktaviana Treasures. 2017. Growth and development of plant cell. Plant Physiology Paper, Offering Undergraduate Biology Education B State University Of Malang. Supervising Professor: Prof. Dra. Herawati Susilo, M. Sc., Ph. D. E-mail: [email protected]
The cell is the basic unit of the life of the growth and development of these cells is regulated by various internal and external signals. Such as hormones, light,
temperature, gravity, predation of insects, diseases and the relationship of other cells with cells. In knowing the development of the necessary knowledge of the plant cell wall in plant cells, the cell division as well as biological events that affect signals on growth and development of plants. The purpose of the making of the paper is to know the aging process and plant cell death program, the
mechanism of enlargement and lengthening the cells of plants and the mechanism of perception and transduction signal in the process of development of the plants. Keywords: plant cell, development, growth
1 BAB I
PENDAHULUAN 1.1 Latar Belakang
Sel adalah unit dasar kehidupan. Perkembangan pada tanaman adalah serumit seperti kelihatannya. Perkembangan pada tumbuhan tergantung juga pada pertumbuhan dan perkembangan yang dikoordinasi oleh sel, baik sel secara individu maupun kelompok. Pertumbuhan dan perkembangan sel-sel tersebut diatur oleh berbagai sinyal internal dan eksternal. Seperti hormon, cahaya, suhu, gravitasi, predasi serangga, penyakit dan hubungan sel dengan sel lainnya.
Dalam mengetahui perkembangan pada sel tumbuhan diperlukan pengetahuan mengenai dinding pada sel tumbuhan, pembelahan sel serta sinyal yang memengaruhi peristiwa biologi pada pertumbuhan dan perkembangan tumbuhan.
1.2 Rumusan Masalah
Berdasarkan latar belakang di atas, permasalahan dapat dirumuskan sebagai berikut :
1. Bagaimana proses penuaan dan program kematian sel tumbuhan? 2. Bagaimanakah mekanisme pembesaran dan pemanjangan sel tumbuhan? 3. Bagaimana mekanisme persepsi dan transduksi signal dalam proses
2 1.3 Tujuan
Berdasarkan rumusan masalah di atas, tujuan pembuatan makalah adalah untuk mengetahui :
1. Proses penuaan dan program kematian sel tumbuhan 2. Mekanisme pembesaran dan pemanjangan sel tumbuhan
3. Mekanisme persepsi dan transduksi signal dalam proses perkembangan tumbuhan
1.4 Manfaat
Manfaat pembuatan makalah adalah dapat mengetahui: 1. Proses penuaan dan program kematian sel tumbuhan 2. Mekanisme pembesaran dan pemanjangan sel tumbuhan
3. Mekanisme persepsi dan transduksi signal dalam proses perkembangan tumbuhan
3 BAB II
TINJAUAN PUSTAKA 2.1 Pengertian penuaan
Bagian akhir dari proses perkembangan, dari dewasa sampai hilangnya pengorganisasian dan fungsi disebut senesen atau penuaan. Sel-sel yang telah berdifferensiasi pada dasarnya mempunyai masa hidup terbatas, sehingga penuaan akan dialami oleh semua sel pada saat yang berbeda-beda. Selama proses penuaan, pada tingkat sel terjadi penyusutan struktur dan rusaknya membran seluler (Fiter & Hay, 1998).
2.2 Pengertian program kematian sel
Senescence adalah suatu bentuk kematian sel terprogram (programmed cell death/ PCD) ,Secara luas didefinisikan sebagai sebuah proses di mana kematian sel yang dikendalikan oleh organisme sebagai bagian dari proses pertumbuhan dan perkembangan, termasuk di dalamnya mekanisme inisiasi dan eksekusi. PCD memerlukan energi dan biasanya diatur oleh serangkaian gen-gen (Hopkins,2009).
2.3 Pengertian pembelahan sel
Pembelahan sel adalah suatu proses dimana material seluler dibagi kedalam dua sel anak (Ruddon,2007).
2.4 Pengertian siklus sel
Siklus sel merupakan proses perkembangbiakan sel yang memperantarai pertumbuhan dan perkembangan makhluk hidup (Ruddon,2007).
2.5 Pengertian persepsi sinyal
Pada tahapan ini sel target mendeteksi molekul sinyal yang berasal dari luar sel. Sinyal kimiawi terdeteksi ketika molekul sinyal berikatan dengan protein reseptor yang terletak dipermukaan atau didalam sel (Campbell, 2008).
2.6 Pengertian transduksi sinyal
Pada tahap ini molekul sinyal memiliki bentuk yang komplamenter dengan situs reseptor yang melekat disitu seperti anak kunci dalam gembok atau
4 substrat dalam situs katalitik suatu enzim. Molekul sinyal berprilaku seperti ligan, istilah molekul yang berikatan secara spesifik dengan molekul lain, seringkali yang berukurakan besar. Pengikatan ligan menyebabkan protein reseptor mengalami perubahan bentuk. Umumnya efek pengikatan ligan menjadi agregasi kedua atau lebih mengaktivasi reseptor lain berinteraksi dengan molekul lainnya (Campbell, 2008).
2.7 Pengertian respon sinyal
Pada tahapan ini sinyal yang ditrandusikan menyebabkan aktivitas selular seperti glikogen fospolirase, penyusunan ulang sitoskeleton ataupun aktivasi gen-gen spesifik dalam nukleus (Campbell, 2008).
5 BAB III
PEMBAHASAN
3.1 Proses penuaan dan program kematian sel tumbuhan 3.1.1.Proses Penuaan
Bagian akhir dari proses perkembangan dari dewasa sampai hilangnya pengorganisasian dan fungsi disebut penuaan. Selama masa pertumbuhan, dengan bertambahnya umur suatu tumbuhan, akan diikuti pula dengan proses penurunan kondisi yang mengarah kepada kematian organ atau organisme (Salisbury,1995). Bagian akhir dari proses perkembangan, dari dewasa sampai hilangnya pengorganisasian dan fungsi disebut senesen atau penuaan. Sel-sel yang telah berdifferensiasi pada dasarnya mempunyai masa hidup terbatas, sehingga penuaan akan dialami oleh semua sel pada saat yang berbeda-beda. Selama proses penuaan, pada tingkat sel terjadi penyusutan struktur dan rusaknya membran seluler (Fiter & Hay, 1998). Tipe-tipe penuaan (senescence) yang dijumpai dalam tumbuhan dapat dikelompokkan sebagai berikut:
• Penuaan yang meliputi keseluruhan tubuh tumbuhan (overall senescence).Akar dan bagian tanaman di atas tanah mati semua tumbuhan mati sesudah menyelesaikan semua. satu siklus kehidupannya
• Penuaan yang meliputi hanya bagian tumbuhan di atas tanah (top senescence).Bagian tubuh di atas tanah mati, sedangkan bagian tumbuhan yang berada di dalam tanah tetap hidup
• Penuaan yang meliputi hanya daun–daunnya (Deciduous senescence). tumbuhan menggugurkan semua daun-daunnya, sementara organ tubuh lain tetap hidup
• Penuaan yang meliputi hanya daun-daun yang terdapat di bagian bawah suatu tumbuhan (Progessive Senescence). Tumbuhan hanya menggugurkan daun-daunnya yang terdapat di bagian bawah saja.
6 Proses penuaan daun dimulai pada tumbuhan umur 44 hari yang ditandai dengan menurunnya kandungan klorofil. Contohnya saja pada tanaman padi, pemberian kalsium mempengaruhi proses penuaan. Pemberian kalsium tersebut mempengaruhi kandungan klorofil, jumlah kloroplas, ukuran kloroplas dan struktur kloroplas pada proses penuaan daun padi (Fiter & Hay, 1998). Terdapat tiga tahapan dalam proses penuaan, yaitu :
a. Fase inisiasi
Penandaan permulaan metabolisme baru. Pada daun senesen terjadi beberapa metabolisme baru seperti, katabolisme klorofil, sintesis amida, kegiatan transport, pembentukan glukosa dari lipid melalui glukoneogenesis dan sintesis protein de novo. Katabolisme atau metabolisme pigmen klorofil terjadi melalui jalur kompleks enzim yang melibatkan beberapa kompartemen di dalam sel dan gejala yang paling mudah diamati adalah hilangnya warna hiaju daun dari waktu ke waktu.
Perubahan keadaan menjadi redox. Terjadi perubahan keadaan redox dimana pada daun dewasa terjadi transfer elektron untuk membentuk NADPH dimana NADPH melepas elektron ke asam askorbat dengan mereduksi glutathione (GSH), sedangkan pada daun senesen, produksi NADPH berkurang, menyebabkan meningkatnya rasio glutathione teroksidasi menjadi GSH dan dehidro askorbat menjadi askorbat.
Pada tingkat molekuler, terjadi aktivasi jalur transduksi sinyal oleh berbagai stimulus tersebut (hormonal, lingkungan, perkembangan, dan patologis) mengakibatkan fase inisiasi yang mencakup aktivasi dan inaktivasi dari berbagai macam gen untuk menginduksi terjadinya senesen.
7 b. Fase reorganisasi
Aktivasi Jalur penyelamatan. Nitrogen dan sulfur organik dikeluarkan oleh daun senesen darimana glutamine dan asparagin diekspor dari daun yang mengalami senesen. Perubahan metabolime dari autotrof menjadi heterotrof. Kebutuhan energi untuk metabolisme, mobilisasi dan proses lainnya menyebabkan meningkatnya energi yang diambil dari proses heterotrof dimana asam amini sebagai sumber karbon.
Detoksifikasi. Terjadinya metabolisme pembentukan asam salisilat dan phenol yang dibentuk secara tidak langsung melalui metabolisme fenilpropanoid. Rediferensiasi organel yang reversible. Merupakan fase transisi di mana terjadi pengendalian rediferensiasi dari struktur sel dan pengaturan kembali posisi material.
c. Fase terminal/ akhir
Terjadi akumulasi antibiotic ketika terjadinya daun senesen dimana berfungsi untuk mencegah masuknya pathogen yang secara oportunis memanfaatkan daun senesen sebagai rute infeksi. Terjadinya pelepasan radikal bebas, eliminasi sisa hasil metabolisme dan hilangnya integritas dan viabilitas sel secara irreversible dimana terjadi autofage dan pembesaran vakuola di sitoplasma dimana membran tidak pecah atau pembentukan autofage vakuola. Integritas dari membran subseluler dan pengaturan kompartemen dari jalur-jalur biokimia tetap dipertahankan hingga terjadinya kematian sel.
3.1.2. Grafik Pola Penuaan
Pertumbuhan tanaman mula-mula lambat, kemudian berangsur-angsur lebih cepat sampai tercapai suatu maksimum, akhirnya laju tumbuh menurun. Apabila digambarkan dalam grafik, dalam waktu tertentu maka akan terbentuk kurva sigmoid (bentuk S). Bentuk kurva sigmoid untuk semua tanaman kurang lebih tetap, tetapi penyimpangan dapat terjadi sebagai akibat
8 variasi-variasi di dalam lingkungan. Ukuran akhir, rupa dan bentuk tumbuhan ditentukan oleh kombinasi pengaruh faktor keturunan dan lingkungan (Wilkins, 1989).
Kurva sigmoid yaitu pertumbuhan cepat pada fase vegetatif sampai titik tertentu akibat pertambahan sel tanaman kemudian melambat dan akhirnya menurun pada fase senesen. Kurva menunjukkan ukuran kumulatif sebagai fungsi dari waktu. Tiga fase utama biasanya mudah dikenali, yaitu fase logaritmik, fase linier dan fase penuaan. Pada fase logaritmik ini berarti bahwa laju pertumbuhan lambat pada awalnya, tapi kemudian meningkat terus. Laju berbanding lurus dengan ukuran organisme. Semakin besar organisme, semakin cepat ia tumbuh. Pada fase linier, pertambahan ukuran berlangsung secara konstan. Fase penuaan dicirikan oleh laju pertumbuhan yang menurun, saat tumbuhan sudah mencapai kematangan dan mulai menua. Laju pertumbuhan relative (relative growth rate) menunjukkan peningkatan berat kering dalam suatu interval waktu dalam hubungannya dengan berat asal. Dalam situasi praktis, rata-rata pertumbuhan laju relative dihitung dari pengukuran yang di ambil pada waktu t1 dan t2 (Wilkins, 1989) Kurva pertumbuhan berbentuk S (sigmoid) yang ideal. Tiga fase utama biasanya mudah dikenali: fase logaritmik, fase linier, dan fase penuaan. Pada fase logaritmik, ukuran (v) bertambah secara eksponensial sejalan dengan waktu (t). Ini berarti bahwa laju pertumbuhan (dv/dt) lambat pada awalnya, tapi kemudian meningkat terus. Pada fase linier, pertambahan ukuran berlangsung secara konstan. Fase penuaan dicirikan oleh laju pertumbuhan yang menurun saat tumbuhan sudah mencapai kematangan dan mulai menua.
9 Gambar 3.1.2. Grafik sigmoid pada penuaan sel tumbuhan
sumber : Wilkins, 1989
3.1.3. Aspek-aspek metabolik penuaan dan pengaruh faktor penuaan 3.1.3.1. Aspek metabolik senesen
Pada tahap sel, penuaan berjalan dengan terjadinya penyusutan struktur dan rusaknya membrane subseluler. Di duga bahwa vakuola bertindak sebagai lisosom, mengeluarkan enzim-enzim hidrolitik yang akan mencerna materi sel yang tidak diperlukan lagi. Penghancuran tonoplas telah menyebabkan enzim-enzim hidrolitik dibebaskan kedalam sitoplasma. Sementara itu bagian dalam struktur kloroplas dan mitokondria mengalami penyusutan sebelum membrane luarnya dirusak. Rupanya proses degradasi yang terjadi pada organel, dimulainya sama seperti yang terjadi pada sel.
Perubahan yang jelas telah terjadi pada metabolisme dan kandungan dalam organ yang mengalami penuaan. Telah terjadi pengurangan DNA, RNA, protein, ion-ion anorganik dan berbagai macam nutrient organic. Fotosintesis berkurang sebelum senesen dimulai dan ini mungkin disebabkan menurunnya permintaan akan hasil fotosintesis. Segera setelah itu klimakterik dalam respirasi terlihat, dan nitrogen terlarut meningkat sebagai akibat dirombaknya protein(Campbel,dkk.,1993).
10
3.1.3.2. Pengaruh faktor pertumbuhan
Sitokinin dapat menghilangkan atau memperlambat proses penuaan. Mekanisme kerja sitokinin dalam proses ini masih belum jelas, tetapi ada petunjuk dari percobaan Mothes yang menunjukkan bahwa setetes sitokinin yang diberikan pada daun, telah menyebabkan terjadinya mobilisasi nutrien organic dan anorganik menuju ke daerah sekitar daun yang diberi sitokinin. Tapi masih belum jelas, apakah peningkatan nutrisi sebagai penyebab langsung permudaan kembali (rejuvenation) atau sitokinin penyebab terjadinya beberapa peristiwa yang menghasilkan permudaan kembali dan mobilisasi nutrisi (Campbel,dkk.,1993).
Tidak semua tumbuhan memberikan respon terhadap hormon yang sama. Sitokinin lebih efektif dalam menahan penuaan pada tumbuhan basah, sedangkan giberelin lebih efektif menahan senesen pada Taraxacum officinale dan Fraxinus. Kadar giberelin endogen akan turun dengan cepat selama senesen pada daun. Auksin (IAA dan 2,4-D) dapat menghalangi senesen pada tumbuhan tertentu. Etilen adalah hormon yang secara jelas merangsang kuat senesen pada banyak jaringan (Soerodikoesoemo,1993). Beberapa faktor luar dapat menghambat atau mempercepat terjadinya senescence, misalnya :
1. Penaikan suhu, keadaan gelap, kekurangan air dapat mempercepat terjadinya senescence daun
2. Penghapusan bunga atau buah akan menghambat senescence tanaman 3. Pengurangan unsur-unsur hara dalam tanah, air, penaikan suhu, berakibat menekan pertumbuhan tanaman yang berarti mempercepat senescence
3.1.4. Program Kematian Sel
Senescence adalah suatu bentuk kematian sel terprogram (programmed cell death/ PCD) ,Secara luas didefinisikan sebagai sebuah proses di mana kematian sel yang dikendalikan oleh organisme sebagai bagian dari proses pertumbuhan dan perkembangan, termasuk di dalamnya mekanisme
11 inisiasi dan eksekusi. PCD memerlukan energi dan biasanya diatur oleh serangkaian gen-gen (Hopkins,2009).
Dua contoh dari PCD antara lain kematian sel akibat senesen dan akibat respon hipersensitivitas. Sense merupakan proses kematian terprogram yang sangat lambat dari suatu jaringan pada akhir dari waktu hidupnya. Sedangkan akibat dari hipersensitivitas, PCD berlangsung lebih cepat untuk mempertahankan diri dari penyakit yang diakibatkan patogen (Hopkins,2009 ).
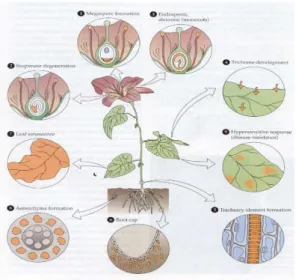
Gambar 3.1.4. Siklus hidup tumbuhan Sumber : Lodish,dkk.,2000
Selama siklus hidup tumbuhan, kematian sel terprogram terjadi pada saat:
Pembentukan gamet, termasuk pembentukan megaspora di bunga (1). Perkembangan embrio, misalnya degenerasi suspensor pada bunga (2). Degenerasi jaringan pada biji dan buah. Misalnya kematian terprogram pada endosperm (3). Perkembangan jaringan dan organ tanaman. Misalnya: pada perkembangan trikoma (4), pembentukan elemen trakeari (tracheary element/ TE) (5), dan pada tudung akar (6). Senesen pada daun (7). Tanaman merespon sinyal lingkungan pada pembentukan aerenkim (8), Tanaman merepon sinyal pathogen yang merupakan respon hipersensitivitas untuk dapat resisten terhadap penyakit (9).
12 3.2 Mekanisme pembesaran dan pemanjangan sel tumbuhan
3.2.1 Pembelahan sel
Pembelahan sel adalah suatu proses dimana material seluler dibagi kedalam dua sel anak. Pembagian sel tumbuhan, seperti sel eukariotik, terjadi dalam dua langkah: mitosis atau kariokinesis, yang hasil dalam penciptaan dua inti baru, dan sitokinesis, pembagian berikutnya dari sitoplasma untuk membuat dua sel anak yang terpisah. Antara divisi namun, masing-masing sel anak harus meningkatkan jumlah sitoplasma seperti membesar, meniru DNA-nya, dan mempersiapkan diri untuk divisi berikutnya. Urutan kejadian ini dikenal sebagai siklus sel.
3.2.2 Siklus sel
Siklus sel merupakan proses perkembangbiakan sel yang memperantarai pertumbuhan dan perkembangan makhluk hidup. Siklus sel dapat digambarkan sebagai empat tahap yang berbeda: fase mitosis (M), fase sintesis DNA (S), dan dua fase intervensi atau ''celah'' (G1 dan G2) (Gambar 3.2.2). Fase M adalah fase divisi di mana kromosom dikondensasikan dan morfologi khas mereka menjadi terlihat dengan mikroskop cahaya. Sepanjang tiga fase lainnya, secara kolektif disebut sebagai interfase, kromosom sepenuhnya tidak terlihat dengan mikroskop cahaya. Selama fase S, replikasi DNA mengarah pada pembentukan dua salinan identik dari kromosom. Pada fase G2, kromosom mulai memadat dan sel akan mempersiapkan diri untuk proses yang diperlukan pada pemindahan kromosom terpisah selama mitosis.
(Ruddon, 2007).
Waktu yang diperlukan untuk menyelesaikan siklus sel sangat bervariasi, tergantung pada jenis sel dan berbagai isyarat perkembangan, tetapi panjang fase S dan fase M tetap kira-kira konstan untuk jenis sel tertentu. G1 adalah fase yang paling variabel dan dapat menjelaskan sebagian besar dari rentang hidup sel.
Agar perkembangan untuk melanjutkan secara tertib, waktu dan tingkat pembelahan sel dan karena itu masuk ke dalam siklus sel harus
13
dikontrol. Kontrol yang diperlukan dicapai melalui interaksi yang rumit dari berbagai kinase, fosfatase, dan protease yang merespon baik sinyal intrinsik dan eksternal. Pusat kontrol siklus sel adalah aktivitas dari cyclin-dependent kinase (CDKs). CDKs terdiri dari dua subunit-satu fungsi subunit sebagai katalisator dan yang lainnya berfungsi untuk mengaktifkan kompleks. Subunit mengaktifkan adalah, protein tidak stabil kecil yang disebut cyclin, dinamakan demikian karena ketika mereka pertama ditemukan di Xenopus telur, diamati bahwa selama setiap siklus sel konsentrasi meningkat terus dari nol dan kemudian tiba-tiba runtuh. Pada tumbuhan, setidaknya ada empat kelas siklin (A, B, D, dan H) dan asosiasi CDKs dengan spesifik siklin merupakan faktor regulasi kunci. Sama pentingnya adalah ketersediaan siklin sebagai tingkat dari berosilasi pengaktif subunit selama siklus sel karena sintesis diatur dan degradasi. Aktivasi CDKs juga membutuhkan fosforilasi residu c treonin spesifik (Thr160) pada subunit katalitik oleh kinase CDK-mengaktifkan (CAK), sedangkan fosforilasi lainnya treonin dan tirosin residu (Thr14, Tyr15) menghambat aktivitas CDK. Enzim fosfatase yang tersedia untuk menghapus gugus fosfat penghambatan pada waktu yang tepat. Dua situs utama kontrol CDK tampak di G1 ke S atau G2 ke M transisi. Hal ini diasumsikan bahwa fungsi kompleks CDK aktif adalah untuk memfosforilasi dan dengan demikian mengaktifkan protein lain yang terlibat dalam sintesis DNA (S fase) atau inisiasi mitosis (fase M).
CDK inhibitor (CKI) juga memainkan peran penting dengan menangkap pembelahan sel bila diperlukan, seperti pada akhir program perkembangan atau dalam menanggapi isyarat hormonal atau lingkungan. Operasi terutama di G1 ke S transisi, sebuah CKI akan mengikat dengan kompleks CDK aktif untuk mencegah dari fosforilasi substrat. Ketika kondisi tepat untuk siklus sel untuk melanjutkan, CKI dapat dihapus oleh proteolisis. Pertama, CKI ditandai untuk degradasi oleh lampiran ubiquitin. Ubiquitin CKI kemudian terdegradasi, yang CDK kompleks diaktifkan kembali, dan G1 untuk S transisi diperbolehkan untuk melanjutkan.
14 Gambar 3.2.2. Fase siklus sel
Sumber : Campbell et al.,1999
3.2.3 Sitokinesis
Sitokinesis adalah proses pembagian sitoplasma kepada dua sel anak hasil pembelahan. Sitokinesis pada sel hewan relatif sederhana, yang melibatkan penyempitan membran sel yang uang muka ke tengah sampai satu sel menjadi dua sel. Keberadaan dinding sel mencegah pola yang sama dari pembelahan sel pada sel tumbuhan. Sel tumbuhan bukannya harus membentuk membran baru dan matriks ekstraselular baru, termasuk lamella tengah dan dua dinding utama baru, dalam sel hidup. (Hopkins, 2009)
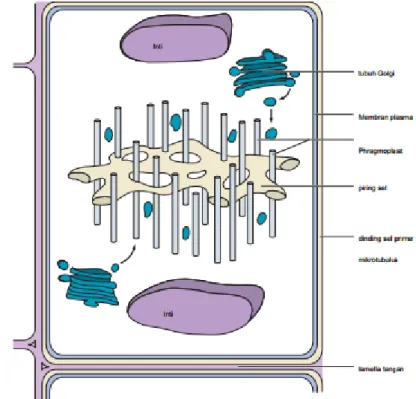
Pembentukan dinding baru dimulai pada mitosis akhir atau fase M dari siklus sel, setelah dua set kromosom telah dipisahkan dan bergerak menuju kutub yang berlawanan dalam sel dan membran nuklir baru telah mulai terbentuk. Pada titik ini, spindel mitosis muncul dan mikrotubul yang telah terbentuk sampai membongkar spindle dan kemudian berkumpul kembali untuk membentuk gugus interdigitating mikrotubulus berorientasi tegak lurus terhadap bidang dari dinding seberang yang baru (Gambar 3.2.3). Disebut phragmoplast, mikrotubulus ini berfungsi untuk mengarahkan pergerakan vesikel sekretorik kecil, berasal dari dekat kompleks Golgi, ke wilayah ekuator sel. Vesikula mulai mengumpulkan di tengah sel di mana mereka menyelaraskan sepanjang bidang ekuator dan mulai menyatu dengan satu sama lain. Agregat yang dihasilkan dari vesikel disebut piring sel.
15
Sebagai vesikel yang baru tiba terus menambah pelat sel, piring tumbuh luar ke segala arah sampai tepi terkemuka membuat kontak dengan membran plasma yang ada di sekitar sel induk. Fusi dari membran piring dengan membran sel induk efektif melengkapi pemisahan dua sel bersaudara. Interior dari pelat-antara dua membran yan diisi dengan zat pectic dijalankan oleh vesikel Golgi dan yang sekarang membentuk lamella tengah antara sel anak. Sitokinesis akan lengkap ketika kompleks selulosa synthase dimasukkan ke dalam membran baru dan masing-masing deposito sel anak dinding primer baru yang berdekatan dengan lamella tengah.
Gambar 3.2.3 The phragmoplast dan plat sel dalam sel tanaman pemisah.
Sumber : Hopkins, 2009
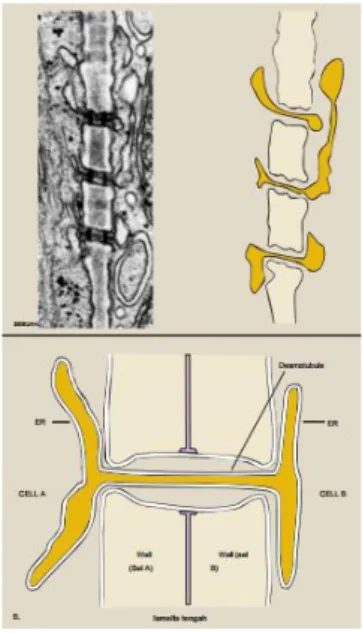
3.2.4 Plasmodesmata
Plasmodesmata adalah saluran sitoplasma yang memanjang untuk penghubung prtotoplas sel yang berdekatan. Ketika bentuk pelat sel, fusi membran tidak lengkap. Hal ini membuat beberapa lokasi di mana kontinuitas sitoplasma dipertahankan antara sel anak. Selulosa yang
16
ditetapkan dan meningkat tembok baru di ketebalan, koneksi ini membentuk saluran membran-terbungkus disebut plasmodesmata. (Hopkins, 2009)
Gambar 3.2.4. Plasmodesmata
Sumber : Hopkins,2009
Plasmodesmata ukurannya tidak besar diameter sekitar 60 nm tetapi ada sejumlah ukuran yang lebih besar dari mereka. Frekuensi diperkirakan berada di kisaran 0,1-10,0 μ m - 2 dari dinding sel, meskipun ada kecenderungan untuk plasmodesmata yang dikelompokan ke dalam daerah sekitar oval disebut medan primer berlubang. Plasmodesmata cukup kecil untuk menghalangi pertukaran organel antara sel-sel tapi cukup besar untuk memungkinkan difusi molekul zat terlarut kecil, termasukinfeksi virus RNA dan transkripsi tanaman faktor, melalui lengan sitosol. Plasmodesmata sehingga memberikan ukuran kontinuitas membran dan sitosol antara sel-sel dan memungkinkan untuk kontrol supracellular lebih program-program pembangunan. (Hopkins, 2009)
Sambungan dari protoplas tetangga melalui plasmodesmata menciptakan pekerjaan net- sitoplasma terus menerus, disebut sebagai simplas, seluruh tanaman. Dengan cara yang sama, apoplast terdiri dari perlu terus ruang ekstraselular atau ruang non sitoplasmik. Apoplast terdiri dari
17
dinding yang saling berhubungan sel, ruang antar, dan dewasa, tak hidup jaringan pembuluh darah. Konsep simplas dan apoplast sangat berguna ketika mengingat pergerakan air dan zat-zat terlarut seluruh tanaman.
3.2.5 Dinding sel dan pertumbuhan sel
Meskipun dinding primer tidak sangat tebal, jaringan penguncian antarnya brils selulosa mikrofi, xyloglycans silang, dan protein struktural menganugerahkan luar kekuatan dan kekakuan pada dinding. Kekuatan ini dan kekakuan dapat mempertahankan bentuk sel dan hubungan struktural dengan sel tetangga dan, pada akhirnya, dukungan dari seluruh pabrik. Dengan demikian sel tumbuh menghadapi tindakan penyeimbangan halus. Ini harus, pada saat yang sama, kekuatan utama dan integritas struktural dari dinding sementara sisanya cukup untuk memberikan ruang untuk protoplas berkembang. (Hopkins)
Hampir semua sel eukariotik, baik hewan dan tumbuhan, mengandung tiga dimensi, jaringan interkoneksi dari protein berserat disebut sitoskeleton. Sitoskeleton memainkan peran penting dalam menentukan organisasi sitoplasma dan bentuk sel, dan dalam pembelahan sel, pertumbuhan, dan diferensiasi. Sitoskeleton sel tumbuhan terdiri dari dua elemen yang berbeda: mikrotubulus dan mikrofilamen. Mikrotubulus adalah batang panjang sekitar 24 nm diameter dan hollow core sekitar 12 nm diameter. Mereka dirakit dari subunit dari protein globular yang disebut tubulin, yang memiliki massa molekul sekitar 100.000 dalton (100 kD) dan terdiri dari dua polipeptida globular ( α- tubulin dan β- tubulin). 1 Mikrotubulus terbentuk ketika tubulin subunit secara spontan merakit diri ke dalam rantai panjang yang disebut protofilament. Protofilamen kemudian berbaris lateral untuk membentuk dinding mikrotubulus (lihat Gambar 3.2.5). Mikrofilamen yang benang yang solid terdiri dari aktin, juga protein globular. Dua rantai subunit aktin merakit diri secara heliks untuk membentuk mikrofilamen sekitar 6 nm diameter. Syarat sitoskeleton adalah pilihan yang disayangkan karena menyiratkan
18
struktur kaku dengan fungsi statis. Sebaliknya, sitoskeleton sangat dinamis. mikrotubulus khususnya yang terus-menerus dirakit, dibongkar, dan disusun kembali sebagai sel membelah, membesar, dan membedakan. Mikrotubulus membentuk gelendong mitosis, yang memainkan peran yang signifikan dalam pergerakan kromosom selama pembelahan sel. Mikrotubulus juga menentukan orientasi dan lokasi dinding sel baru antara sel anak, dan deposisi selulosa dalam dinding sel tumbuh. (Hopkins, 2009)
Mikrofilamen muncul untuk mengontrol arah aliran sitoplasma, mengalir terus menerus dari partikel sitoplasma dan organel sekitar pinggiran sel. Bentuk keseluruhan mikrofilament atau bundel paralel berorientasi ke arah aliran sitoplasma. Mikrofilamen juga terlibat dalam pertumbuhan tabung serbuk sari. Ketika serbuk sari biji-bijian mulai tumbuh mengembangkan ekstensi tubular yang tumbuh ke bawah stigma dari bunga dan berfungsi untuk memberikan inti laki-laki untuk telur. Pertumbuhan tabung hanya di ujung, dan vesikel yang mengandung prekursor dinding sel dipandu melalui sitoplasma ke ujung tumbuh oleh jaringan mikrotubulus. Pentingnya sitoskeleton dalam mengatur dan mengkoordinasikan sifat dinamis dari sel tumbuh hanya mulai dihargai. Teknik untuk studi tanaman sitoskeleton sel berkembang cepat, dan kita bisa mengharapkan kemajuan yang menarik di masa depan.
19 Gambar 3.2.5 Mikrotubulus dan ratapan mikrofil
Sumber : Hopkins,2009
Kekuatan dan kekakuan pada dinding. Kekuatan ini dan kekakuan mempertahankan bentuk sel dan hubungan struktural dengan sel tetangga dan, pada akhirnya, dukungan dari seluruh pabrik. Dengan demikian sel tumbuh menghadapi tindakan penyeimbangan halus. Ini harus, pada saat yang sama, utama-tain kekuatan dan integritas struktural dari dinding sementara sisanya suf fi sien liat untuk memberikan ruang untuk protoplas berkembang.
Pertumbuhan sel didorong oleh serapan ai dan terbatas denngan kekuatan dan kekakuan dari dinding sel, karena sebagian besar volume setiap sel adalah air, berikut bahwa untuk sel meningkatkan volume itu harus mengambil air dari atas. Jika misalnya, sel bermandikan sebuah isotonik larutan manitol atau zat terlarut serupa yang tidak dapat masuk ke dalam sel, sel juga tidak akan mengambil air dan tidak akan tumbuh. Kekuatan pendorong untuk pembesaran sel adalah serapan air. Ingat bahwa sel-sel mengambil air dengan proses osmosis. Konsentrasi zat terlarut tinggi dari protoplasma dan vacuolar getah mengurangi potensi air dari sel ke titik di mana air berdifusi ke dalam sel. Bukan berarti untuk mengkompensasi, sel
20
dikelilingi oleh sumber air murni terus membengkak tanpa batas waktu atau setidaknya sampai tekanan internal melebihi kekuatan tarik membran. Konsekuensi dari situasi seperti ini jelas menunjukkan ketika sel-sel darah merah mamalia ditempatkan dalam air. Sel-sel cepat membengkak sampai semburan membran plasma, melepaskan isinya ke dalam media. Kebanyakan sel-sel hewan menghindari peristiwa osmotik seperti dengan menggunakan energi metabolisme untuk mengeluarkan baik zat terlarut atau air dan dengan demikian mempertahankan tekanan keseimbangan. sel tumbuhan telah menemukan solusi mereka yang berbeda mengelilingi membran plasma dengan dinding sel yang kuat, lebih atau kurang kaku. Tekanan turgor (tekanan positif) dikembangkan sebagai protoplas memperluas mendorong dinding naik sampai menyeimbangkan tekanan osmotik negatif dari protoplas tersebut. Pada saat itu potensi air sel mendekati nol dan tidak lebih bersih Pertumbuhan-akan air serapan-atau sel terjadi. Ini adalah kekuatan dan kekakuan dari dinding sel yang memaksakan restriksi kritis pada kapasitas sel tanaman untuk tumbuh. Dengan demikian, untuk sel untuk meningkatkan ukuran, kekuatan dan kekakuan dari dinding sel harus dimodifikasi untuk mengurangi potensi air sel, mengizinkan serapan air, dan, akibatnya, memungkinkan sel untuk memperbesar.
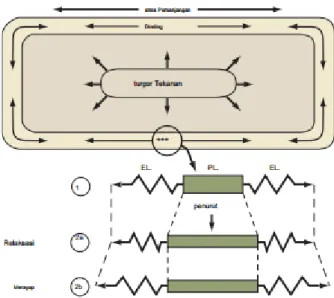
Perpanjangan dinding sel membutuhkan proses pelonggaran pada dinding yang memungkinkan di dinding menyerah pada turgor tekanan.
Setiap kenaikan volume sel membutuhkan peningkatan yang sesuai di daerah permukaan dinding di sekitarnya, atau ekstensi dinding. Peneliti tahu bahwa ekstensi dinding terkait dengan turgor tekanan-ini telah didemonstrasikan secara empiris. Misalnya, ketika tekanan turgor secara eksperimental berkurang, tingkat ekspansi sel juga menurun. Selanjutnya, ekstensi dinding dan pertumbuhan tidak terjadi di sel-sel pada tekanan yang sangat rendah atau nol turgor, meskipun sel-sel tetap metabolik rangsangan pertumbuhan aktif dan sesuai yang hadir. Adanya saling ketergantungan ini perpanjangan dinding dan tekanan turgor dapat diringkas oleh hubungan sederhana berikut:
21
dV / dt = m (P - Y) (17.1)
Istilah dV / dt adalah perubahan (d) volume (V) dari sel dari waktu ke waktu (dt). dV / dt demikian cara sederhana untuk mengungkapkan tingkat pertumbuhan sel. Y adalah ambang batas yield, atau Tekanan turgor minimum yang diperlukan untuk ekspansi sel terjadi. Istilah m adalah dinding diperpanjang, sebuah konstanta proporsionalitas antara tingkat pertumbuhan dan tekanan turgor (P) yang melebihi ambang batas yield. Dinding diperpanjang adalah ukuran kuantitatif dari kapasitas dinding untuk ireversibel meningkatkan luas permukaannya. Menurut hubungan ini, pertumbuhan sel tergantung terutama pada jumlah dimana tekanan turgor melebihi ambang batas yield. Hal ini jelas bahwa sel tumbuh dihadapkan dengan peran saling bertentangan con- tekanan turgor. Di satu sisi, tekanan turgor menentang penyerapan terus air, yang merupakan kekuatan pendorong untuk ekspansi sel. Pada saat yang sama, tekanan turgor mempromosikan ekstensi dinding ireversibel dan pembesaran sel. Bagaimana sel menyelesaikan flik on ini? Jawaban atas pertanyaan ini disediakan oleh konsep relaksasi stres. Stres relaksasi di dinding merupakan pusat proses pertumbuhan sel. tekanan turgor berkembang karena sel dinding-selulosa mikro fi brils silang dengan xyloglucans menolak deformasi sebagai protoplas mencoba untuk memperluas. Kekuatan dari protoplas memperluas mendorong dinding sehingga menghasilkan stres (didefinisikan sebagai kekuatan / satuan luas) dalam dinding. pertumbuhan sel tampaknya ini diciptakan ketika tekanan ini santai oleh peristiwa dinding-melonggarkan yang menyebabkan unsur-unsur beban di dinding untuk menjadi lebih negatif), diikuti oleh penyerapan pasif air. Dalam fluks air pada gilirannya akan meningkatkan volume sel, memperluas dinding sel, dan cenderung untuk mengembalikan kedua stres dinding dan tekanan turgor. Proses pertumbuhan sel demikian dilihat sebagai penyesuaian terus menerus tekanan turgor melalui relaksasi stres dalam rangka untuk menyeimbangkan peran
22
saling bertentangan dalam penyerapan air dan ekstensi dinding sel.
Gambar 3.2.6. Sebuah model untuk relaksasi stress di dinding sel tumbuhan Sumber : Hopkins,2009
3.3 Mekanisme persepsi dan transduksi signal dalam proses perkembangan tumbuhan.
Perkembangan suatu tanaman diatur oleh berbagai sinyal yang bekerja pada beberapa tingkat. Sinyal pada tumbuhan bisa disebut dengan hormon. Hormon adalah utusan kimia yang memungkinkan sel untuk berkomunikasi satu sama lain. Beberapa hormon pada tanaman seperti auksin, giberelin, sitokinin, asam absisat, etilen, dan brassinoteroids dikenal mendukung atau menghambat perkembangan pada tumbuhan baik tanggapan perkembangan secara tunggal maupun kombinasi. Terdapat juga faktor intraseluler lain seperti turgor, status mineral yang dapat menyebabkan sel memodifikasi metabolisme serta perkembangannya. (Hopkins, 2009)
Terdapat juga faktor eksternal seperti cahaya, suhu, gravitasi dan predasi serangga yang memiliki dampak pada suatu tumbuhan. Terdapat juga faktor lingkungan lain seperti kelembaban tanah, kelembaban nutrisi dan patogen. (Hopkins, 2009).
23 3.3.1 Sinyal Persepsi dan Transduksi
Sinyal presepsi yang diikuti oleh beragam peristiwa biokimia disebut juga dengan transduksi sinyal yang menentukan respon sel. Dalam menanggapi suatu sinyal, sel pada suatu tanama memiliki reseptor. Dalam menanggapinya diperlukan suatu mekanisme yang mengkonversi atau transduksi sinyal untuk perubahan kimia atau metaboisme sel. Semua sinyal perkembangan terbagi dalam suatu urutan yaitu persepsi sinyal, transduksi dan tanggapan. (Hopkins, 2009)
Signal transduksi pada tingkat sel mengacu pada pergerakan sinyal dari luar sel ke dalam. Gerakan sinyal dapat sederhana, seperti yang terkait dengan molekul reseptor dari kelas asetilkolin. Interaksi ligan, memungkinkan sinyal akan disahkan dalam bentuk gerakan ion kecil, baik masuk atau keluar dari sel. Gerakan-gerakan ini ion mengakibatkan perubahan potensial listrik dari sel-sel yang, pada gilirannya, menyebarkan sinyal sepanjang sel. Sinyal transduksi lebih kompleks melibatkan kopling dari interaksi reseptor-ligan terhadap peristiwa intraseluler banyak. Peristiwa tersebut termasuk phosphorylations oleh kinase tirosin dan / atau serin / kinase treonin. (Hopkins, 2009)
3.3.2 Sistem Reseptor G-Protein
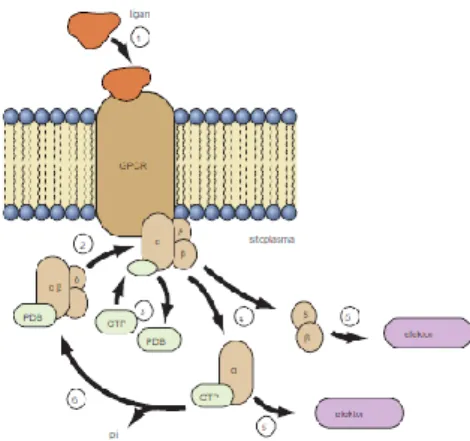
Reseptor pada hewan dan tumbuhan terdapat reseptor yang digabungkan di dalam sel untuk mengikat GTP(disebut dengan G-Protein) dan menghidrolisis protein. Reseptor yang berinteraksi dengan G-Protein semuanya memiliki struktur yang tercirikan dari 7 Transmembran. (Hopkins, 2009)
Transduksi sinyal dimulai ketika ligan ( hormon atau molekul peraturan kecil lainnya) berikatan dengan GPCR pada permukaan sel luar. Ligan mengikat menyebabkan molekul GPCR untuk mengubah
24 konformasi, atau bentuknya, sehingga sekarang memiliki afinitas yang lebih tinggi untuk G protein. G protein mengikat ke permukaan sitoplasma dari reseptor dan G α subunit pertukaran GDP untuk molekul guanosin trifosfat (GTP). G protein kini aktif, atau '' pada. '' Pada langkah berikutnya, protein G yang diaktifkan memisahkan dari reseptor dan G α subunit memisahkan dari G gabungan βγ subunit. Setiap G α dan G βγ subunit sekarang bebas untuk mengaktifkan protein efektor. Efektor adalah enzim-enzim yang pada gilirannya mengontrol jumlah utusan ondary sek- diproduksi atau yang mengatur laju yang ion melintasi membran. Contoh molekul efektor termasuk calmodulin, phospholipases, dan protein kinase dan lipid yang dibahas di bagian berikut. G α subunit juga memiliki aktivitas GTPase, sehingga ketika G α subunit telah menyelesaikan tugasnya, GTP dihidrolisis terhadap PDB dan fosfat anorganik. Berikut hidrolisis GTP, G α subunit memisahkan dari protein efektor dan bebas untuk bergabung kembali dengan G βγ subunit dan dengan demikian reformasi kompleks heterotrimeric tidak aktif. Selama GPCR itu sendiri tetap diaktifkan, dapat berubah pada beberapa protein G, sehingga memperkuat aliran sinyal ke molekul efektor. (Hopkins, 2009)
Gambar 3.3.2. Transduksi sinyal pada sistem reseptor G-Protein Sumber : Hopkins, 2009
25 3.3.3 Prinsip dasar sinyal kinase, fosfolipase, ion kalsium dan transkripsi
Reseptor di permukaan sel dapat mengenali berbagai sinyal ekstrasel dan diperbanyak sebagai kaskade intrasel, sinyal ekstrasel ini disebut sebagai primary messenger. Sinyal intrasel yang terjadi akibat perubahan sinyal dari permukaan memerlukan second messenger untuk berbagai aktivitas intrasel. Kebanyakan multiplikasi sinyal intrasel memakai protein dan enzim fosforilisasi atau defosforilisasi dengan mengaktivasi berbagai kinase atau fosfatase. (Kurniawan, 2015).
Spesies utama second messenger pada tanaman termasuk enzim protein kinase, ion kalsium, dan turunan fosfolipid. Protein kinase dan kinase kaskade telah terlibat dalam beragam jalur transduksi sinyal tanaman. Protein kinase adalah enzim-enzim yang mengaktifkan protein lain dengan katalis forforilasi. (Hopkins, 2009)
Fosfolipid merupakan unsur utama pembentuk membran lipid, selain mengandung asam lemak dan alkhol, juga mengandung residu asam fosfat, dan degradasi fosfolipid yang terdapat dalam sel (Peter A. Mayes, 2003).
Lipid muncul sebagai kelas utama pada second messenger. Pada second messenger dihasilkan fosfolipase. Fosfolipase termasuk enzim hidrolitik. (Maharani, 2012). Dimana peranan fosfolipase adalah menghidrolisis fosfolipid. Fosfolipase terdiri atas fosfolipase A1, fosfolipase A2, fosfolipase C, dan fosfolipase D. Masing-masing enzim ini bekerja secara spesifik. (Hopkins, 2009)
Proses regulasi berbagai proses fisiologi pada tanaman seperti perpanjangan dan pembelahan sel, sekresi dan aktivitas berbagai enzim, aksi hormon, dan tanggapan tropik melibatkan ion kalsium. Suatu ion kalsium akan bekerja secara efektif pada second messenger apabila ia memiliki konsentrasi yang rendah dan di bawah kontrol metabolik. (Hopkins, 2009)
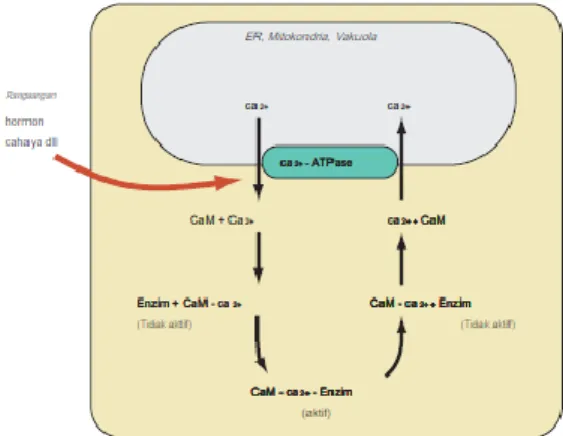
26 Sinyal Ca2+ adalah salah satu sistem sinyal utama di dalam sel. Sinyal Ca2+ ini berfungsi untuk mengatur banyak proses seluler. Sinyal Ca2+ dapat memicu kehidupan baru pada saat proses pembuahan, juga mengontrol banyak proses selama pertumbuhan, begitu satu sel membelah/berdiferensiasi, sinyal akan mengatur hampir semua aktivitas proses seluler, yang menentukan bagaimana metabolisme dan sekresi. Ada juga efek merugikan untuk sinyal Ca2+ ini, naiknya konsentrasi yang berlebihan dapat menyebabkan kematian sel, baik dalam cara yang terkendali atau sel terprogram (apoptosis) atau proses yang menghasilkan nekrotik.(Kurniawan, 2015)
Prinsip utama reseptor kalsium dalam sel tumbuhan dan hewan adalah kalmodulin, yang sangat dilestarikan, protein mana-mana yang dapat diisolasi dari berbagai tanaman yang lebih tinggi, ragi, jamur, dan ganggang hijau. bisa ditemukan di hampir setiap sel eukariotik dan membentuk hampir 1% dari semua total massa protein. Kalmodulin berfungsi sebagai reseptor Ca2+ intrasel dan mengatur proses yang melibatkan Ca2+. Kalmodulin terbentuk dari protein yang kuat, satu rantai polipeptida dengan empat tempat ikatan Ca2+ dengan afinitas tinggi. (Kurniawan, 2015)
Gambar 3. Kalsium sebagai secondary message. Pertukaran kalsium antara vakuola dan sitosol dapat diatur oleh hormon atau
27 Sumber : Hopkins, 2009
Pada reseptor yang ditemukan intra seluler dan pada ligan mengikat bermigrasi ke inti di mana-reseptor kompleks ligan langsung mempengaruhi transkripsi gen. Reseptor intraselular dan fungsi dalam nukleus sebagai faktor transkripsi mereka sering disebut sebagai reseptor nuklear.
Beberapa sinyal tanaman, seperti fitokrom pigmen yang sensitif terhadap cahaya dan beberapa hormon, by-pass reseptor membran plasma dapat mendukung reseptor nuklear dan intervensi langsung dalam ekspresi gen. Transkripsi merupakan pengkopian daerah pengkode pada DNA menjadi RNA untai tunggal (Loodish, 2000).
Dalam kasus setidaknya tiga sinyal molekul-fitokrom dan dua hormon auxin dan giberelin-bukti menunjukkan adanya faktor transkripsi itu, dengan tidak adanya sinyal, merepresi ekspresi gen sasaran.
28 BAB IV
PENUTUP 4.1 Simpulan
Berdasarkan pembahasan diatas, dapat disimpulkan bahwa :
1. Penuaan merupakan bagian dari program kematian sel. Tipe tipe penuaan yang dijumpai pada tumbuhan adalah overall senescence, top senescence, deciduous senescence, Progessive Senescence
2. Pembelahan sel adalah suatu proses dimana material seluler dibagi kedalam dua sel anak. Siklus sel merupakan proses perkembangbiakan sel yang memperantarai pertumbuhan dan perkembangan makhluk hidup 3. Semua sinyal perkembangan terbagi dalam suatu urutan yaitu persepsi
sinyal, transduksi dan tanggapan. Mekanisme presepsi dan transduksi sinyal adalah Reseptor di permukaan sel dapat mengenali berbagai sinyal ekstrasel dan diperbanyak sebagai kaskade intrasel, sinyal ekstrasel ini disebut sebagai primary messenger. Sinyal intrasel yang terjadi akibat perubahan sinyal dari permukaan memerlukan second messenger untuk berbagai aktivitas intrasel. Kebanyakan multiplikasi sinyal intrasel memakai protein dan enzim fosforilisasi atau defosforilisasi dengan mengaktivasi berbagai kinase atau fosfatase.
4.2 Saran
1. Sebaiknya kita terlebih dahulu membaca buku referensi yang dasar, sebelum beranjak menggunakan buku dengan isi yang lebih kompleks 2. Sebaiknya kita juga menguasai bahasa inggris sehingga tidak kesulitan
dalam mempelajari buku dengan bahasa inggris
3. Sebaiknya mahasiswa mengetahui kerangka materi terlebih dahulu sebelum membuat suatu makalah
29 DAFTAR RUJUKAN
Campbel, Reece dan Mitchel. 1993. Biologi Jilid 2. Erlangga. Jakarta.
Campbell, N.A,. J.B.Reece, & L.G. Mitchell. 2005. Biologi. Edisi 5. Jilid 3. Terjemahan dari: Biology. 5th ed. Oleh Manalu, w. Jakarta: Erlangga
Campbell Dan Reece. 2008. Biology edisi 8. Jakarta : Erlangga.
Fiter.A.H, Hay.R.K.M. 1998. Fisiologi Lingkungan Tanaman. Gajah Mada University Press : Yogyakarta.
Hannahs, Greg. 1997. Plasmodesmata. http//:biology.kenyon.edu
Hopkins, G. William and Huner, A P Norman. 2009. Introduction to Plant Physiology. USA : Jhon Wiley & Sons, Inc
Kurniawan, S. N. 2015. Homeostasis Ca2+ Intraselular. Jurnal MNJ. Vol.01, No.01
Lodish H, Baltimore D, Berk A, Zipursky SL, Matsudaira P, Darnell J. 2000. Molecular cell biology.6th ed. New York : Scientific American Books, Inc. Peter A, 2003. Biokimia Harper Edisi 5. Jakarta: alih bahasa Andry Hartono Ruddon, R.W. 2007. Cancer Biology.Edisi Keempat. Michigan: Oxford
University Press. Halaman 4-5, 12.
Salisbury,F.B. 1995. Fisiologi Tumbuhan Jilid 3. Institut Tekhnologi Bandung : Bandung
Soerodikoesoemo. 1993. Anatomi dan Fisiologi Tumbuhan. Universitas Terbuka, Depdikbud : Jakarta