www.elsevier.nlrlocateraqua-online
The effect of growing level and growing method on
winter mortality, Mikrocytos roughleyi, in diploid
and triploid Sydney rock oysters, Saccostrea
glomerata
Ian R. Smith
a,), John A. Nell
a, Robert Adlard
ba
NSW Fisheries, Port Stephens Research Centre, Taylors Beach, NSW 2316, Australia
b
Queensland Museum, PO Box 3300, South Brisbane, Queensland 4101, Australia
Accepted 8 November 1999
Abstract
The effect of growing height on mortality of Sydney rock oysters, Saccostrea glomerata, caused by Australian winter mortality Mikrocytos roughleyi was evaluated on a commercial intertidal growing lease on the Georges River, NSW, Australia. The mortality level of 35.1"8.4% at normal growing height, fell to 9.3"0.9% when oysters were cultured 300-mm higher, and increased to 52.4"0.2% at 300-mm lower in the tidal range. No significant growth penalty was evident at the higher growing level, and no relationship could be discerned between the mortalities at different growing levels and either glycogen content or condition index. The concern among
Ž
oyster farmers, that M. roughleyi may cause higher mortality levels among single seed
cultch-.
less diploid and triploid S. glomerata, compared with the same oysters grown on sticks, was not supported in a trial at two intertidal growing levels at Merimbula, NSW. These findings imply that under the conditions tested, considerable protection against M. roughleyi can be gained by raising the growing height over winter in southern NSW growing areas, and that the growth advantages of triploid S. glomerata can be utilized without an increased mortality level caused by this disease. Further trials in other growing areas, and during a severe epizootic of M. roughleyi, are required
)Corresponding author. Tel.:q61-2-49821232; faxq61-2-49821107.
Ž .
E-mail address: [email protected] I.R. Smith .
0044-8486r00r$ - see front matterq2000 Elsevier Science B.V. All rights reserved. Ž .
before these findings can be generalized to southern to mid-NSW.q2000 Elsevier Science B.V.
All rights reserved.
Keywords: Oysters; Mortality; Triploidy; Mikrocytos; Parasite; Culture
1. Introduction
Ž . Ž
The Sydney rock oyster, Saccostrea glomerata Gould, 1850 formerly S.
commer-.
cialis; Buroker et al., 1979; Anderson and Adlard, 1994 , is cultured intertidally by the
Ž .
stick and tray method in estuaries along the eastern seaboard of Australia Nell, 1993 , from around latitude 278S to 37.58S. Across the southern portion of the range of culture, Port Stephens to the NSW-Victorian border, the species is susceptible to Australian
Ž .
winter disease Lauckner, 1983 known locally as ‘winter mortality’. This may kill up to 80% of oysters in a local area, generally in their third winter and just before they reach
Ž . Ž .
market size Lauckner, 1983 . Roughley 1926 described the signs of the disease and
Ž .
associated it with low temperatures. Wolf 1976 suggested high salinity as a
contribut-Ž .
ing factor. Farley et al. 1988 named and described the aetiological agent, Mikrocytos
roughleyi, a protistan parasite of uncertain taxonomic affinity.
Growing level within the intertidal range was identified as a contributing factor to the
Ž . Ž . Ž .
severity of winter mortality. Roughley 1926 , Wolf 1967 and Nell and Smith 1988 provided anecdotal evidence that mortalities could be reduced by raising the growing level. Oyster farmers currently deal with the disease by relaying their oysters to upstream leases before May when infestation generally begins, andror by selling those oysters large enough to be marketed before the end of July and the onset of mortality
ŽNell and Smith, 1988 . The latter option is not available to many farmers from southern.
growing areas and losses can be high in bad years.
Triploid S. glomerata exhibited increased growth rates over that of their diploid
Ž .
siblings under commercial culture conditions Nell et al., 1994; Hand et al., 1998a .
Ž .
Triploid production in hatcheries currently produces single seed cultchless oysters, which require mesh trays or similar methods for growout. Oyster farmers have expressed the belief that trayed diploid oysters are more susceptible to winter mortality than oysters on sticks, and feared that the single seed triploids might similarly suffer higher mortalities. For triploids to be commercially successful in NSW, it is necessary that their resistance to winter mortality, when grown on trays, is at least as great as that of wild-caught oysters cultivated using the standard tarred stick method.
Ž .
Smith 1991 found no difference between Sydney rock oyster diploids and triploids on trays in their response to winter mortality in the Georges River. The work of Hand et
Ž .
al. 1998b , however, showed triploidy conferred greater survival when exposed to winter mortality compared with sibling diploids at some sites, and no difference at others.
2. Materials and methods
2.1. Mortalities at fiÕe growing leÕels on the Georges RiÕer
Space on a commercial oyster lease in Woolooware Bay, Georges River, NSW
Ž X X
.
33859 S, 151812 E was provided by an oyster farmer. The tarred wooden rails used to
Ž .
support trayed oysters in the traditional growing method Malcolm, 1987 were modified to provide five growing levels at height intervals of 150 mm, centred on the intertidal
Ž
growing level normally used by oyster farmers in Woolooware Bay slightly below
.
mid-tide within the maximum range of 2 m . Oyster sticks, when laid out by oyster farmers in Woolooware Bay, are set at the second highest level used in this experiment, i.e., normal tray level plus 150 mm.
Commercial 1.8=0.9 m, tarred timber and galvanised wire mesh oyster trays were divided into two equal compartments. Two trays at each level provided four replicate compartments at each of the five growing levels.
Ž
Three-year-old S. glomerata of approximately ‘plate’ grade 45 to 50 g whole
.
weight were obtained from local oyster farmers, and were randomly mixed before allocation to tray compartments at a stocking rate of 190 oysters per compartment
Ž 2.
235rm . The trays were covered with a light plastic netting to minimise oyster losses resulting from wave action.
Ž . Ž
The experiment ran from 23 April 1986 autumn to 12 January 1987
mid-.
summer . Oysters that died at the start as a result of culling damage were replaced with live oysters held for this purpose in trays adjacent to the experimental oysters.
Mortalities were determined monthly by counting and removing dead shells; a sample of six oysters from each replicate was weighed, the condition index of the pooled meats determined and analysed for glycogen content. Glycogen content was assayed by the
Ž . Ž .
enzymatic method of Kepler and Decker 1974 . The condition index CI was calcu-lated as:
dry weight of oyster meat g
Ž .
=100 CIsshell cavity volume ml
Ž
.
Cavity volumes were determined by subtracting the weight in-air of the oyster’s shell
Ž .
from the weight in-air of the whole oyster Lawrence and Scott, 1982 . This method is valid because the effective density of cavity contents is close to 1 grml, providing the oysters have not gaped and lost cavity fluid. Meat weights were determined by drying them to a constant weight at 908C.
2.2. Growing method and leÕel on diploid and triploid S. glomerata at Merimbula
Juvenile sibling diploid and triploid S. glomerata were obtained from the same batch
Ž .
used in the commercialisation trials of Hand et al. 1998a . Parent stock had been spawned at the Port Stephens Research Centre’s Mollusc Hatchery in February, 1994. Triploidy was then induced by blocking meiosis 2 polar body extrusion with
cyto-Ž . Ž
chalasin B Nell et al., 1996 resulting in 88% triploidy measured using the flow
.
Ž .
Spat were reared in mesh cylinders Holliday et al., 1993 , in Tilligerry Creek, Port Stephens to a size suitable for the experiment. Triploid and diploid oysters for the
Ž
experiment were dry graded through 18 mm and retained on 14-mm mesh diagonal
.
measurement .
Ž X
The experiment used commercial oyster racks at Merimbula Lake, NSW 36855 S,
X .
149855 E from June 1994 to October 1996. Racks were located at the high salinity, seaward end of the oyster growing area, where exposure to M. roughleyi is high, and the experiment was serviced at approximately 3-month intervals to remove fouling organ-isms.
Ž .
Treatments comprised two growing systems stick and tray , two growing levels and two ploidy types, providing eight treatment combinations, each replicated four times. Each replicate was stocked with 400 oysters.
The ‘stick’ growing system used standard tarred hardwood oyster sticks 25-mm square section=2-m long in frames of five sticks nailed at 130-mm intervals to half
Ž .
stick cross members near each end Nell, 1993 . The sticks were used to support oyster spat glued to the sides at 50-mm intervals using a non-acid curing silicone plumbers’ sealant. This system models closely the traditional commercial stick growing method of
Ž .
wild oyster spat caught on tarred sticks Nell, 1993 with the exception that oysters were glued at regular spacings rather than randomly settled from the wild.
The ‘tray’ growing system used standard single seed trays of 1.8=0.9-m tarred hardwood timber frames, subdivided into 12 compartments, and the bottom covered with
Ž .
9-mm diagonal polyethylene mesh. A second tray was used as a lid. Half the tray, six compartments, were stocked with oysters for the experiment, the remaining six were stocked with oysters used to provide samples to test for the presence of the M. roughleyi parasite.
Frames of sticks and trays were supported on a commercial oyster growing rack of
Ž .
tarred hardwood posts and rails Nell, 1993 . Two growing level treatments were used. Standard growing level, at a little under half tide was the same as other commercial oyster leases in the area. The second growing level was set up adjacent to the first but
Ž .
300 mm below the standard level to increase exposure to winter mortality Wolf, 1976 .
Ž .
Shell height umbo to ventral valve margin and whole weight were measured for 400 oysters randomly taken from each ploidy type before the sticks and trays were stocked at the start of the experiment. At termination of the experiment in October, 1996, shell height and whole weight were measured for 50 oysters from each replicate. Mortality levels during the final winter were assessed by counting live and dead oysters in each replicate at the termination of the experiment in October 1996. Percentage mortalities
Ž .
were calculated as number of deadrnumber of deadqnumber of alive =100. In June and October 1996, oyster samples were taken and forwarded to Brisbane for
Ž
M. roughleyi diagnosis by the Hemacolor stained imprint method Adlard and Lester,
.
1995 .
2.3. Statistical methods
2.3.1. Mortality at fiÕe growing leÕels on the Georges RiÕer
Ž .
Ž .
variance Sokal and Rolf, 1981 in the statistical software package, STATGRAPHICS,
Ž .
version 5 Statistical Graphics, Rockville, MD .
2.3.2. Growing method and leÕel on diploid and triploid S. glomerata at Merimbula
All data were analysed using multiple ANOVA in the Statgraphics package above. For the measurements on samples of spat taken at the start of the experiment, homogeneity of variances was confirmed for shell height for triploids and diploids using
Ž .
Cochran’s test Cs0.507, P)0.05 . Weights were log transformed to satisfy
require-Ž .
ments for homogeneity Cs0.501, P)0.05 .
Ž
Weight data were expressed as weight increase, final mean whole weight for each
. 0.5
replicate-initial mean weight . Percentage mortality data were arcsin x transformed.
Ž
Homogeneity was confirmed for shell height and whole weight Cs0.354, Cs0.256,
.
respectively: P)0.05 . Mortalities remained slightly heterogeneous following
transfor-Ž .
mation Cs0.467, Ps0.029 . This is, however, a minor departure from homogeneity
Ž .
and ANOVA is sufficiently robust to overcome such departures Underwood, 1981 .
3. Results
3.1. Mortality leÕels at fiÕe growing leÕels on the Georges RiÕer
At the termination of the experiment, an inverse relationship was apparent between
Ž .
cumulative mortality levels and growing level Table 1, Fig. 1 . The mean cumulative
Ž . Ž .
mortality level of oysters at the highest level 9.3% was significantly lower P-0.05
Ž .
than at normal tray level 35.1% and this in turn was significantly lower than at the
Ž .
lowest growing level 52.4% . Mortalities at intermediate growing levels were consistent with this trend after the August count when a sharp increase in mortalities was recorded
Ž .
for all but the highest growing level Fig. 1 .
Measurements of whole weight, dry meat condition index and glycogen content were
Ž .
not significantly different between the growing level treatments P)0.05, Table 1 .
Table 1
Effect of growing level on the physiology and mortality levels of Sydney rock oysters S. glomerata Growing level September 1986
Whole Condition Glycogen Cumulative Cumulative
U U U
Ž . Ž . Ž . Ž .
weight g index gr100 g dry meat mortalities % mortalities %
UU UU
to Sept. 1986 to Jan. 1987
a a a a a
Data are expressed as mean"SD. Within columns, means, which do not have a common superscript,
Ž .
differ significantly P-0.05 . UU
For analysis of variance an arcsin x0.5transformation was used and Tukey’s test was used to compare
Ž .
Fig. 1. The effect of deviation of standard growing height on cumulative mortality levels % of the Sydney rock oyster S. glomerata in Woolooware Bay, NSW from April 1986 to January 1987.
Data for the five growing levels were pooled for dry meat condition index and for glycogen content to reveal overall trends, which followed the usual seasonal pattern
ŽNell et al., 1994 . These parameters are tabulated for the September sampling Table 1. Ž .
at the time when mortalities were rapidly increasing, and showed no aberrations that might be associated with the mortalities.
3.2. Growing method and leÕel on diploid and triploid S. glomerata at Merimbula
The April 1995 visit revealed losses of oysters from the high level diploid stick, and the high level triploid stick treatments. The leaseholder reported pelicans roosting as the cause. Numbers at termination were markedly reduced for these treatments.
Table 2
The proportion of Sydney rock oysters S. glomerata infested with the winter mortality parasite M. roughleyi at Merimbula Lake, NSW
These trials began on June, 1994.
Date of examination Treatment Number Number Proportion
ŽPloidyrculturerheight. examined infested infested %Ž .
June 1996
TriploidrTrayrLow 30 0 0
DiploidrTrayrLow 30 0 0
Oct. 1996
TriploidrTrayrHigh 20 6 30
TriploidrTrayrLow 20 0 0
DiploidrTrayrHigh 10 0 0
DiploidrTrayrLow 10 3 30
TriploidrStickrHigh 10 3 30
DiploidrStickrHigh 10 0 0
Table 3
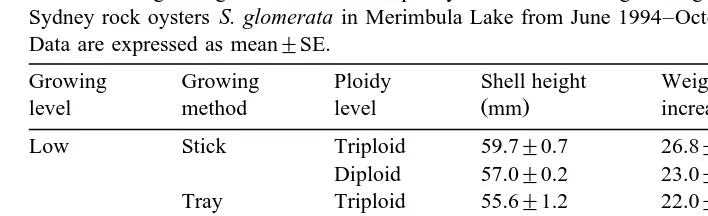
The effect of growing level, method and ploidy level on shell height, weight increase and mortality levels of Sydney rock oysters S. glomerata in Merimbula Lake from June 1994–October 1996
Data are expressed as mean"SE.
Growing Growing Ploidy Shell height Weight Mortalities
Ž . Ž . Ž .
level method level mm increase g %
Low Stick Triploid 59.7"0.7 26.8"0.4 15.6"4.7 Diploid 57.0"0.2 23.0"0.4 18.4"6.0 Tray Triploid 55.6"1.2 22.0"0.4 8.5"1.6 Diploid 53.5"0.8 18.6"0.6 16.5"3.8 High Stick Triploid 53.5"0.6 21.6"1.0 1.8"0.6 Diploid 54.1"0.7 20.4"0.5 2.5"0.5 Tray Triploid 56.0"0.8 23.0"1.1 3.3"0.5 Diploid 52.7"1.5 19.3"1.0 8.6"1.2
It is probable that fouling organisms, especially on low trays, reduced growth rates by reducing food availability. Winter mortality occurred mainly during the second winter
Ž .
and spring June–October, 1996 . Infestation of oysters with the winter mortality
Ž .
parasite, M. roughleyi was evident in the October, 1996 samples Table 2 .
Shell height did not differ with ploidy when analysed using single factor ANOVA
ŽFs0.155, P)0.05 , however, shell height was significantly affected P. Ž -0.05 by.
Ž .
ploidy, substrate and by growing level, with a significant interaction P-0.05 between
Ž .
growing method and growing level Table 3 . Whole weight increase was significantly
Ž . Ž
affected P-0.05 by ploidy and substrate, again with a significant interaction P
-.
0.05 between substrate and growing level. Triploids had a significantly greater whole
Ž .
weight increase than diploids Fs17.580, P-0.001 . Mortalities were affected
signif-Ž .
icantly P-0.05 only by growing level.
4. Discussion
The Georges River growing level experiment provides statistical confirmation of earlier suggestions that raising the growing level of S. glomerata can provide protection
Ž .
against losses attributable to M. roughleyi Roughley, 1926; Wolf, 1976 . Growth was less at the highest level but not significantly so. Farmers indicated that spring 1986 was an atypically poor season for oyster growth so growth reduction with increased height may prove greater during an average season. Further elucidation of the growth costs and resistance benefits is desirable, and should include an estimate for losses during a severe
M. roughleyi epizootic.
Easily adjustable support systems for oyster trays, baskets and sticks are now available and could provide a relatively low cost method for raising oysters in situ during the colder months in southern estuaries where moving the oysters up river or
Ž
under mangroves a traditional prophylactic which raises their intertidal height as well as
.
Condition, as exemplified by condition index and glycogen content, was not corre-lated with winter mortality. However, increased losses to winter mortality in the oysters’ third year of growth, the common occurrence of large focal necroses in gonadal tissue in
M. roughleyi infested oysters, and the greater resistance of triploids with their impaired
Ž .
gonadal development Hand et al., 1998b suggest a link between the disease and gonadal development.
In the Merimbula experiment, winter mortality occurred only during the second
Ž .
winterrspring season June–October, 1996 . There was no significant effect of growing method or ploidy on mortalities, but all low oyster treatments suffered greater rates of mortality. Diploid and triploid oysters in a parallel study carried out simultaneously
Ž .
further upstream experienced no winter mortality Hand et al., 1998a . There was, however, a similar growth advantage for triploids over diploids in this study as was
Ž .
observed by Hand et al. 1998a .
As the study demonstrated that trayed oysters are no more susceptible to winter mortality than stick-grown oysters when cultured at the same level, farmers can use tray cultivation in Lake Merimbula without increased risk of oyster kill due to winter mortality. This effect may need to be assessed in other estuaries along the NSW south coast, before it can be ascertained whether the results obtained in this study apply generally.
Acknowledgements
We thank Mr. S. Hunter, PSRC for his assistance with the Merimbula experiment and Messrs. N. Verdich, C. Boyton, and H. Wheeler who made their oyster leases and other facilities available for this study. Thanks are due to Ms R.E. Hand, PSRC, for assistance with the Merimbula experiment and Dr W.A. O’Connor, PSRC for statistical analysis of the data, both also contributed valuable editorial comments. Partial funding for this
Ž
project was provided by the Fisheries Research and Development Corporation FRDC
.
93r151 .
References
Adlard, R.D., Lester, R.J.G., 1995. Development of a diagnostic test for Mikrocytos roughleyi, the aetiological
Ž
agent for Australian winter mortality of the commercial oyster, Saccostrea commercialis Iredale and
.
Roughley . J. Fish Dis. 18, 609–614.
Anderson, T.J., Adlard, R.D., 1994. Nucleotide sequence of a rDNA internal transcribed spacer synonymy of
Saccostrea commercialis and S. glomerata. J. Moll. Stud. 60, 196–197.
Buroker, N.E., Hershberger, W.K., Chew, K.K., 1979. Population genetics of the family Ostreidae: I. Intraspecific studies of Crassostrea gigas and Saccostrea commercialis. Mar. Biol. 54, 157–169. Farley, C.A., Wolf, P.H., Elston, R.A., 1988. A long-term study of ‘‘Microcell’’ disease in oysters with a
Ž . Ž .
description of a new genus, Mikrocytos g. n. , and two new species, Mikrocytos mackini sp. n. and
Ž .
Mikrocytos roughleyi sp. n. . Fish. Bull. 3, 581–593.
Hand, R.E., Nell, J.A., Maguire, G.B., 1998a. Studies on triploid oysters in Australia: X. Growth and mortality
Ž .
Hand, R.E., Nell, J.A., Smith, I.R., Maguire, G.B., 1998b. Studies on triploid oysters in Australia. XI. Survival
Ž Ž ..
of diploid and triploid Sydney rock oysters Saccostrea commercialis Iredale and Roughley through outbreaks of winter mortality caused by Mikrocytos roughleyi infestation. J. Shellfish Res. 17, 1129–1135. Holliday, J.E., Allan, G.L., Nell, J.A., 1993. Effects of stocking density on juvenile Sydney rock oysters,
Ž .
Saccostrea commercialis Iredale and Roughley , in cylinders. Aquaculture 109, 13–26.
Ž .
Kepler, D., Decker, K., 1974. Glycogen determination with amyloglucosidase. In: Bergmeyer, H.U. Ed. , Methods of Enzymatic Analysis Vol. 3 Academic Press, New York, pp. 1127–1131.
Ž .
Lauckner, G., 1983. Diseases of Mollusca: bivalvia. In: Kinne, O. Ed. , Diseases of Marine Animals Vol. 2 Biologische Anstalt Helgoland, Hamburg, pp. 447–961.
Lawrence, N.D., Scott, G.I., 1982. The determination and use of condition index of oysters. Estuaries 5, 23–27.
Malcolm, W.B., 1987. The Sydney rock oyster. Agfact F3.1.1. Department of Agriculture, Sydney, NSW, 12 pp.
Ž .
Nell, J.A., 1993. Farming the Sydney rock oyster Saccostrea commercialis in Australia. Rev. Fish. Sci. 1, 97–120.
Nell, J.A., Cox, E., Smith, I.R., Maguire, G.B., 1994. Studies in triploid oysters in Australia: I. The farming
Ž .
potential of triploid Sydney rock oysters Saccostrea commercialis Iredale and Roughley . Aquaculture 126, 243–255.
Nell, J.A., Hand, R.E., Goard, L.J., McAdam, S.P., Maguire, G.B., 1996. Studies on triploid oysters in Australia: evaluation of cytochalasin B and 6-dimethylaminopurine for triploidy induction in Sydney rock
Ž .
oysters Saccostrea commercialis Iredale and Roughley . Aquacult. Res. 27, 101–110.
Nell, J.A., Smith, I.R., 1988. Management, production and disease interactions in oyster culture. In: Bryden,
Ž .
D.I. Ed. , Fish Diseases. Refresher Course for Veterinarians, Sydney, 1988. Proceedings 106, Post Graduate Committee in Veterinary Science. University of Sydney, NSW, pp. 127–133.
Roughley, T.C., 1926. An investigation of the cause of an oyster mortality on the Georges River, New South Wales, 1924–5. Proc. Linn. Soc. N.S.W. 51, 446–491.
Smith, I.R., 1991. The effect of growing level on growth, condition and winter mortality of Sydney rock
Ž . Ž .
oysters Saccostrea commercialis, Iredale and Roughley . In: Maguire, G.B., Nell, J.A., Smith, I.R. Eds. ,
Ž .
Mariculture Nutrition. Final Report to Fisheries Research and Development Corporation FRDC . NSW Fisheries, Brackish Water Fish Culture Research Station, Salamander Bay, NSW, pp. 112–129. Sokal, R.R., Rolf, F.J., 1981. Biometry. Freeman, New York, NY, 895 pp.
Underwood, A.J., 1981. Techniques of analysis of variance in experimental marine biology and ecology. Oceanogr. Mar. Biol. 19, 513–605.
Winer, B.J., 1971. Statistical Principles in Experimental Design. 2nd edn. McGraw-Hill, Kogakusha, Tokyo.
Ž .
Wolf, P.H., 1967. Winter mortality hits state’s oyster output. Fisherman 2 9 , 20–22.
Ž .