ANALISIS POLIMORFISME GEN Growth Differentiation Factor 9 (GDF-9) dan HUBUNGANNYA DENGAN KEBERHASILAN INSEMINASI BUATAN pada SAPI PO
Teks penuh
Gambar
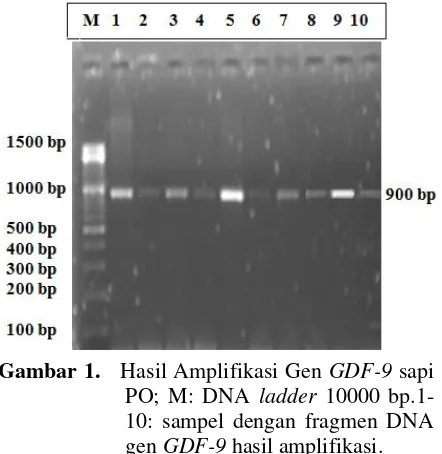
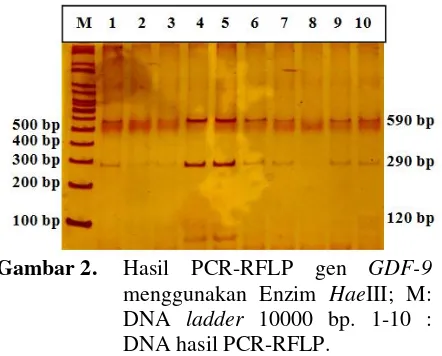

Dokumen terkait
Dengan ini saya menyatakan bahwa skripsi berjudul Evaluasi Keberhasilan Pelaksanaan Program Inseminasi Buatan (IB) pada Sapi Potong di Kota Sawahlunto adalah benar karya saya
PENGETAHUAN PETERNAK TENTANG PEMAHAMAN KETERKAITAN GEJALA BERAHI DENGAN KEBERHASILAN INSEMINASI BUATAN PADA SAPI DI KECAMATAN LENGAYANG KABUPATEN PESISIR SELATAN SUMATERA
Tujuan penelitian ini adalah untuk mengetahui tingkat keberhasilan dan faktor-faktor yang mempengaruhi tingkat keberhasilan Inseminasi Buatan (IB) pada sapi potong
Dari kesimpulan dapat disimpulkan bahwa tidak ada perbedaan keberhasilan inseminasi buatan antara sapi Bali dara dengan sapi Bali yang pernah beranak, ditinjau dari
Program yang dikembangkan untuk mendukung peningkatan populasi, mutu dan produksi sapi salah satunya adalah Inseminasi buatan (IB).Penelitian ini adalah: 1)
Berdasarkan hasil penelitian, dapat disimpulkan bahwa Inseminasi Buatan yang dilakukan pada sapi Brahman Cross dengan interval waktu 0-4 jam memiliki nilai
vi EVALUASI KEBERHASILAN INSEMINASI BUATAN PADA SAPI PERAH BERDASARKAN PENGALAMAN DAN UMUR INSEMINATOR YANG BERBEDA DI KECAMATAN LEMBANG RIFQI JIHADI ASYRAF NIM.. 210220111
Peran Faktor Peternak dan Inseminator Terhadap Keberhasilan Inseminasi Buatan Pada Sapi Potong di Kecamatan Kota Bangun.. Jurnal Peternakan Lingkungan
