Directory UMM :Data Elmu:jurnal:A:Agricultural & Forest Meterology:Vol106.Issue4.2001:
Teks penuh
Gambar
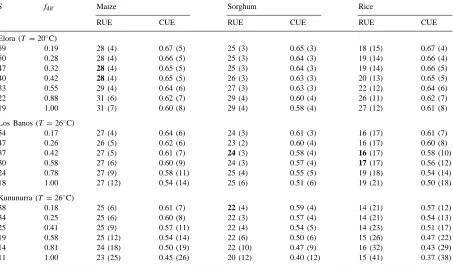
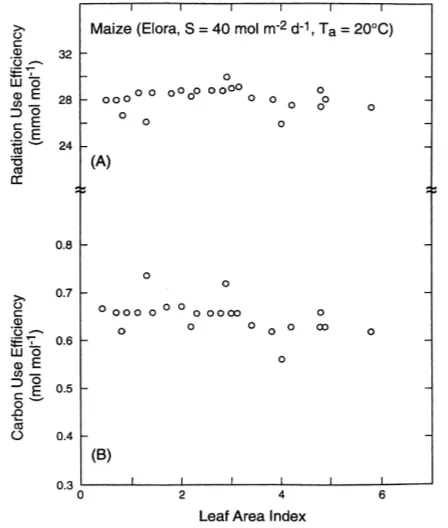
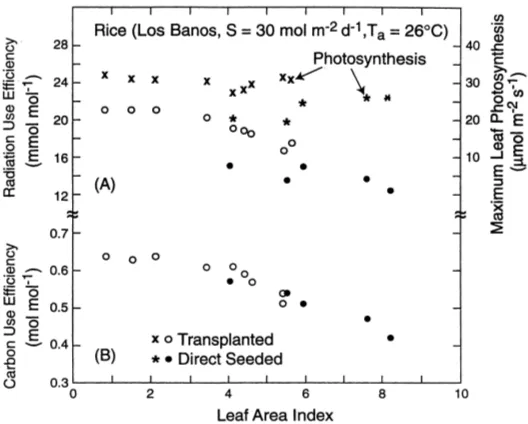
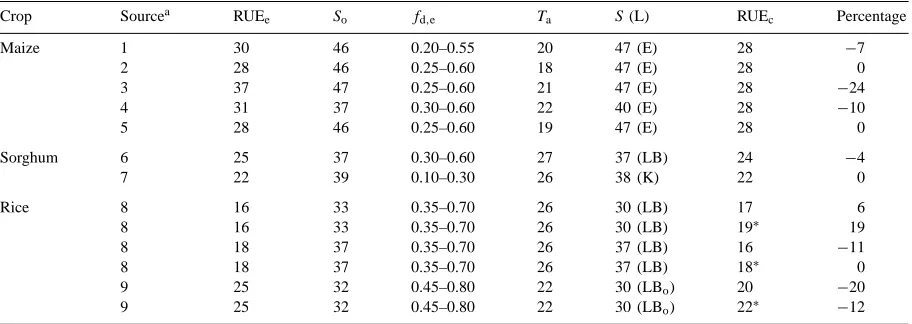
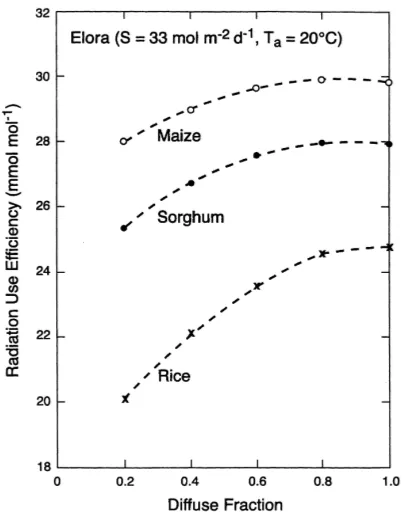
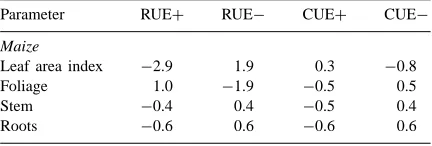
Dokumen terkait
Annual course of daily mean (from top to bottom) leaf conductance, transpiration rate, net photosynthetic rate and water use efficiency of dominant and suppressed shoots in the
Sensitivity analysis investigating the effects of altered soil and vegetation optical properties on daily sums (sun rise till sun set) of vegetation net radiation (R Nveg ), latent
Contrary to the electrical analogy, where soil and vegetation controls on evaporation are reproduced with the help of empirical surface resistances (allowing for the calculation of
It was found that using measured values to initialize the aerosol optical depth in the retrieval of surface global irradiance, the sur- face albedo increased by about 0.02 on
The vertical distribution of sources and sinks within the canopy is derived from direct measurements of both the NH 3 concentration profile and the turbulence within the canopy,
Micrometeorological measurements above the canopy indicated that gaseous Cl compounds were emitted for most of the time, and this was supported by the source/sink distributions
The EXAMINE project, funded by the European Commission, the UK Department of the Environment, Transport and the Regions, and other national organi- zations, has taken just such
We present experimental results on the radiation environment of the Scots pine canopy and using these data, the differences between various spatial and temporal averaging procedures
