181
PHENACOCCUS MANIHOTI MATILE-FERRERO (HEMIPTERA:
PSEUDOCOCCIDAE), MEALYBUG INVASIF BARU DI INDONESIANila Wardani
Balai Pengkajian Teknologi Pertanian Lampung Jl. Hi. Za. Pagar Alam No. IA, Rajabasa, Bandar Lampung
ABSTRAK
Kutu putih yang berasosiasi pada tanaman ubi kayu di Indonesia ada beberapa spesies diantaranya adalah Ferrsia virgata (Cockerell), Paracoccus marginatus Williams&Granara de Willink, dan Phenacoccus manihoti Matile-Ferrero.P. manihoti merupakan hama kutu putih pada ubi kayu yang diperkirakan masuk ke Indonesia (Jawa Barat) tahun 2010. Hama ini berasal dari Amerika Selatan dan telah menyebar ke beberapa negara produsen ubi kayu seperti Afrika dan Thailand. Hama ini merupakan hama utama pada daerah-daerah tersebut dan dapat menyebabkan kehilangan hasil mencapai 80%. Di Indonesia hama ini juga telah menyebar ke beberapa daerah sentra ubi kayu seperti Lampung dan Jawa Timur. Sampai saat ini belum banyak yang mengetahui perihal dari hama ini, walaupun telah melihat gejala serangannya di pertanaman. Dalam tulisan ini akan dijelaskan mengenai morfologi, bioekologi, dan kerusakan yang ditimbulkan oleh P. manihoti.
Kata kunci: Phenacoccus manihoti, invasif, Indonesia
ABSTRACT
Mealybug on cassava plants in Indonesia there are several species include Ferrsia virgata (Cockerell), Paracoccus marginatus Williams & Granara de Willink, and Phenacoccus manihoti Matile-Ferrero. P. manihoti are pests of cassava mealybug on the expected entry into Indonesia (West Java) in 2010. This pest is native to South America and has spread to several countries cassava producers such as Africa and Thailand.It is a major pest in these areas and cause yield losses of 80%. In Indonesia, this pest has spread to some central areas of cassava as Lampung and East Java. there has not been a lot to know about this pest, although have seen symptoms of its presence in the crop. This paper will explain the morphology, Bioekologi, and the damage of P. manihoti.
182
PENDAHULUAN
Kutu tanaman dari famili Pseudococcidae sering disebut dengan mealybug atau dalam bahasa Indonesia dikenal dengan nama kutu putih. Kerusakan tanaman yang disebabkan oleh hama ini karena adanya hisapan stilet, dan beberapa jenis dapat berperan sebagai vektor virus tanaman, atau menimbulkan klorosis karena adanya saliva (air liur) yang bersifat toksik (Williams dan Willink, 1992; Ben-Dov dan Hodgson, 1997).
Kutu tanaman sering menginvasi ke daerah-daerah yang sebelumnya belum terinfestasi melalui pengiriman bahan tanaman yang tidak melalui pemeriksaaan karantina. Peledakan populasi sering terjadi apabila terbawanya kutu tidak disertai dengan musuh alaminya atau tidak terdapat musuh alaminya di daerah baru. Dan populasi kutu akan tetap stabil pada daerah baru karena adanya kontrol dari musuh alami baik predator maupun parasitoid (Williams dan Willink 1992). Dan peranan penting dari musuh alami kutu putih ini telah banyak dilaporkan di benua Amerika dan Afrika (van Driesche et al., 1990; CIAT, 1990; Neuenschwander , 2004).
Salah satu hama kutu putih yang menyerang tanaman ubikayu di Indonesia adalah P. manihoti. Kutu putih P. manihoti berasal dari Amerika Selatan dan masuk ke Afrika pada tahun 1980- an. Hama ini pertama kali memasuki kawasan Asia Tenggara yaitu di Thailand pada tahun 2009, yang menyebabkan serangan berat, kemungkinan telah menyebar ke Laos dan kamboja. Di Indonesia pertama kali ditemukan di daerah Jawa Barat pada akhir tahun 2010 (Muniappan et al. 2011, Rauf, 2011). Dalam tulisan ini akan dijelaskan mengenai morfologi, bioekologi, dan kerusakan yang ditimbulkan oleh kutu putih P. manihoti pada tanaman ubi kayu.
Kutu Putih (mealybug) pada Tanaman Ubi Kayu
Kutu putih juga dikenal dengan nama mealybug, merupakan serangga dari Ordo Hemiptera, famili Pseudococcidae. Kutu ini banyak menyerang tanaman, bersifat sedenter, dan tubuh ditutupi oleh lapisan lilin yang berwarna putih. Banyak tanaman pertanian yang berasosiasi dengan kutu putih ini.
Beberapa spesies Pseudococcidae ditemukan pada tanaman ubi kayu dan pada tanaman sekerabat lainnya (Cox dan Williams 1981). Williams dan Granara (1992) melaporkan bahwa ada 19 spesies dari Sub Ordo Sternorrhyncha, famili Pseudococcidae yaitu Ferrisia meridionalis Williams, Ferrisia terani Williams dan
183
Granara de Willink, Ferrisia virgata (Cockerell), Hypogeococcus spinosus Ferris, Nipaecoccus nipae (Maskell), Paracoccus herreni Williams dan Granara de Willink, Paracoccus marginatus Williams dan Granara de Willink, Phenacoccus gregosus Williams dan Granara de Willink, Phenacoccus helianthi (Cockerell), Phenacoccus herreni Cox dan Williams, Phenacoccus madeirensis Green, Phenacoccus manihoti Cox dan Williams, Planococcus citri (Risso), Planococcus minor (Maskell), Pseudococcus affinis (Maskell), Pseudococcus elisae, Pseudococcus mandio Williams, Pseudococcus maritimus, Puto barberi (Cockerell). Perhatian utama ditujukan pada dua spesies yang dominan ditemukan dan meyebabkan serangan berat adalah Phenacoccus manihoti dan Phenacoccus herreni (Cox dan Williams 1981). Dari sejumlah kutu putih yang menyerang tanaman ubi kayu hanya Phenacoccus herreni dan Phenacoccus manihoti, yang penting secara ekonomi.
Kutu putih P. manihoti berasal dari Amerika Selatan dan masuk ke Afrika pada tahun 1980 an. Hama ini pertama kali memasuki kawasan Asia Tenggara yaitu Thailand pada tahun 2009, yang menyebabkan serangan berat pada pertanaman ubi kayu, kemungkinan telah menyebar ke Laos dan Kamboja. Di Indonesia pertama kali ditemukan di daerah Jawa Barat pada tahun 2010 (Muniappan et al., 2011; Rauf, 2011).
Kutu putih P. manihoti, dilaporkan mempunyai inang terbatas pada ubi kayu, namun tidak tertutup kemungkinan bahwa hama ini juga mempunyai inang alternatif pada tanaman sekerabat ubi kayu seperti tanaman jarak, dan karet, karena di beberapa negara hama ini dilaporkan bersifat oligophagus. P. manihoti and P. herreni di Afrika juga menyerang Talinum triangularae Jack. (Portulacaceae) (Neuenschwander et al. 1986). Di Amerika Selatan, P. manihoti ditemukan menyerang Citrus spp. (Rutaceae) dan kedelei Glycine max (L.) Merr. (Fabaceae) dan P. herreni dilaporkan hanya pada M. esculenta (Williams dan Granara 1992). Di laboratorium, P. manihoti bisa dipelihara dan diperbanyak pada poinsettia, Euphorbia pulcherrina Wild. (Euphorbiaceae) (Boussienguet, 1984).
Perbedaan ketahanan tanaman ubi kayu terhadap P. manihoti telah dipelajari oleh Tertuliano et al. (1993). Dilaporkan bahwa ketahanan ubi kayu terhadap P. manihoti disebabkan adanya perbedaan kadar kandungan senyawa sekunder pada tanaman ubi kayu.Senyawa asam sianida merupakan senyawa sekunder yang terdapat di dalam floem tanaman ubi kayu. Floem merupakan
184
jaringan tempat kutu putih P. manihoti menghisap cairan untuk dikonsumsi sebagai nutrisi. Adanya senyawa ini akan mempengaruhi perkembangan dan reproduksi P. manihoti, dan pada akhirnya juga akan mempengaruhi tingkat ketahanan tanaman ubi kayu terhadap P. manihoti. (Catalayud et al., 1994; Fraenkel, 1969). Semakin tinggi kadar asam sianida pada ubi kayu maka semakin rentan varietas tersebut terhadap kutu P. manihoti.
Morfologi, bioekologi Kutu Putih, Phenacoccus manihoti
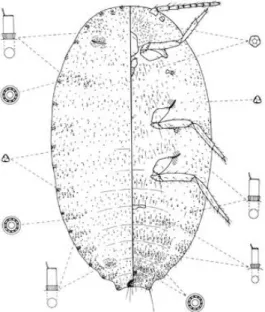
Phenacoccus manihoti merupakan hama yang hidup pada pucuk tanaman, di bawah permukaan daun atau batang dari tanaman ubi kayu. Tubuh berwarna pink yang ditutupi oleh lapisan lilin, berbentuk oval, mempunyai filamin tubuh yang pendek (James et al., 2000). Secara rinci Matile-Ferrero (1978) menyebutkan bahwa spesies P. manihoti mempunyai telur berbentuk oval, dengan warna kuning keemasan dan ditutupi oleh kantung telur (ovisac) yang terdapat pada ujung posterior dari betina dewasa. Panjang dan lebar telur berkisar antara 0.30-0.75 mm dan 0.15–0.30 mm. Tubuh nimfa terdiri dari 6 ruas pada instar pertama dan 9 ruas pada instar selanjutnya. Panjang dan lebar tubuh nimfa berturut-turut untuk instar pertama 0.40–0.75 dan 0.20–0.30 mm. Instar kedua 1.00–1.10 dan 0.50–0.65 mm. Instar ketiga 1.10–1.50 dan 0.50– 0.60 mm, imago 1.10–2.60 dan 0.50–1.40 mm. Betina dewasa berbentuk oval, berwarna merah muda (pink) dan ditutupi tepung putih berlilin, mata relatif berkembang, tungkai berkembang baik dengan ukuran yang sama (Gambar 1).
Gambar 1. Phenacoccus manihoti Matile-Ferrero, betina dewasa (Cox & Williams, 1981)
185
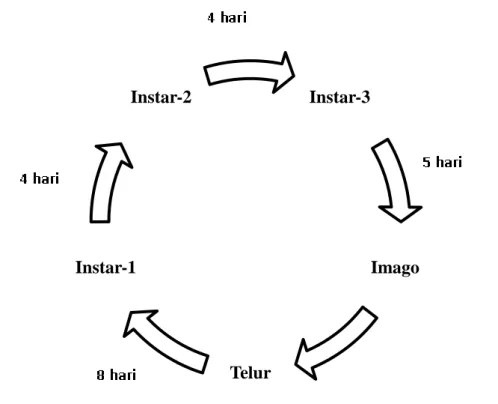
P. manihoti bersifat partenogenetik telitoki, yaitu semua keturunan yang dihasilkan adalah betina, sehingga satu ekor kutu mampu untuk manghasilkan keturunan dan jika populasi melimpah mampu menyebabkan ledakan hama di pertanaman. Dalam kondisi optimal, satu betina mampu menghasilkan 200-600 butir telur (Iheagwam, 1981; Lema dan Herren, 1985) yang terdapat dalam kantung telur di bawah daun dan disekitar tunas apikal dan lateral tanaman. Ovisak mudah menempel pada pakaian, dan dapat berfungsi untuk penyebaran jarak jauh. Telur menetas menjadi nimfa instar-1 (crawler) dan dapat berpindah ke tanaman lainnya dengan perantaraan angin. Nimfa makan pada floem batang tanaman, dan mengalami 3 instar sebelum menjadi dewasa. Di bawah kondisi laboratorium pada suhu 25 oC, untuk perkembangan dari telur menjadi dewasa membutuhkan waktu rata-rata 31-33 hari (Iheagwam dan Eluwa, 1983). Perkembangan optimal terjadi pada suhu 27 oC (Lema dan Herren 1985) dan kematian yang signifikan terjadi di bawah suhu 15 °C (Yaseen, 1982) dan di atas 33 °C (Iheagwam dan Eluwa, 1983). Perkembangan dari telur sampai dewasa membutuhkan waktu lebih kurang 21 hari (Gambar2) (Nwanze, 1977).
Gambar 2. Siklus perkembangan Phenacoccus manihoti (Nwanze, 1977)
Instar-3
Imago
Telur
Instar-1
186
Curah hujan merupakan faktor penentu dalam dinamika populasi kutu P. manihoti di lapangan (Herren dan Hennessey, 1983). Hujan dapat menyebabkan kutu hanyut secara mekanis. Perubahan populasi P.manihotisangat dipengaruhi oleh adanya stress pada tanaman karena kekeringan atau kekurangan air. Kekeringan akan meningkatkan senyawa sekunder pada tanaman ubi kayu. Senyawa ini berperan sebagai fagostimulan bagi P. manihoti, sehingga dapat meningkatkan populasinya pada tanaman. Sebaliknya pemberian pupuk organik, mulsa dan NPK dapat menurunkan serangan P. manihoti pada tanaman ubi kayu (Neuenschwander, 1994).
Di Afrika dan Amerika Selatan populasi P. manihoti meningkat pada musim kemarau (Le Ru dan Catalayud, 1994), jumlah kutu berlipat dalam waktu 7-10 minggu dari jumlah populasi kurang dari 10 individu/tanaman menjadi lebih dari 100 individu/tanaman. P. manihoti menyebabkan distorsi pada tunas terminal, daun menguning dan keriting, ruas memendek, batang menjadi stunting, dan melemah. Dengan tidak adanya musuh alami dan tindakan pengendalian lainnya, kerusakan ini dapat mengurangi hasil lebih dari 80 % (Nwanze KF, 1977). Tidak ada kultivar ubi kayu yang diketahui sepenuhnya tahan terhadap P. manihoti (Calatayud dan Le Rü, 2006). Hasil eksplorasi musuh alami asli P. manihoti telah ditemukan empat hymenoptera parasit, dua belas predator dan satu jamur entomopatogen (Lohr et al., 1990; Yaseen, 1982).
Kerusakan yang Disebabkan oleh Kutu Putih P. manihoti
Kutu putih pada ubi kayu menghisap cairan daun dan pucuk tanaman. Serangan hama dapat mengurangi panjang internode dan menyebabkan daun menjadi mengkerut dan pucuk mengerdil atau disebut bunchy tops .Serangan berat dapat menyebabkan defoliasi pada tanaman, namun hal ini jarang terjadi karena ubi kayu merupakan tanaman cepat untuk bertunas kembali. Kutu putih ini juga dapat menyebabkan distorsi pada batang (Gambar 3). Serangan lebih berat terjadi pada musim kering dibandingkan musim basah (James et al., 2000).
187
Gejala awal Gejala Bunchy top Gejala distorsi pada batang Gambar 3. Gejala serangan P. manihoti pada tanaman ubi kayu
Selama proses makan kutu putih menghisap cairan floem dan mengambil kalsium dari daun ubi kayu. Hal ini terbukti ketika daun ubi kayu dianalisis kandungan Ca, P, dan K, pada daun yang tidak terserang ditemukan kandungan Ca 32% lebih banyak dari daun yang terserang, dan tidak berbeda nyata untuk kandungan N, P, dan K. Reduksi Ca menyebabkan lemahnya daun, dinding sel sedikit mengkerut, dan daun menjadi melengkung (CIAT 1988). Pada tanaman yang terserang kutu putih juga terjadi pengurangan laju fotosintesa, pengurangan efisiensi transpirasi dan mesofil daun, dan dalam waktu yang bersamaan juga terjadi defisit air, CO2 internal dan juga pengurangan temperatur daun (CIAT 1992).
Musuh Alami Kutu Putih dan Peranannya
Menurut Charernsom and Suasa-ard (1994; 2010), beberapa parasitoid dan predator diketahui berasosiasi dengan ubi kayu (Tabel 1). Interaksi antara musuh alami dengan hama telah banyak dilaporkan, di antaranya oleh Nadel dan van Alphen (1987) dan Souissi et al.(1997) yaitu respon dari parasitoid betina pada zat kimia volatil yang dikeluarkan oleh tanaman ubikayu yang terserang kutu putih. Telah diteliti model interaksi antara P. manihoti danApoanagyrus lopezi, dan P. herreni dengan Apoanagyrus diversicornis Howard,Aenasius vexans Kerrich, Acerophagus coccois Smith (Hymenoptera: Encyrtidae).Contoh lain adalah spesies endoparasitoid untuk mengendalikan P. herreni di Amerika Selatan (Bertschy et al. 1997; 2001).
188
Tabel 1. Hama tanaman ubi kayu dan musuh alaminya
________________________________________________________________ Hama tanaman ubi kayu Serangga musuh alami
Ferrisia virgata - Acerophagus sp. (Hymenoptera:
Encyrtidae)
P. madeirensis - Allotropa sp. (Hymenoptera: Encyrtidae)
P. manihoti - Anagyrus sp. (Hymenoptera: Encyrtidae)
P. solenopsis
Pseudococcus jackbeardsleyi Serangga predator:
(Homoptera: Pseudococcidae) - Plesiochrysa ramburi (Neuroptera: Chrysopidae)
- Mallada basalis (Neuroptera: Chrysopidae - Spalgis epius (Lepidoptera: Lycaenidae) - Brumoides sp. (Coleoptera:
Coccinellidae)
- Chilomenes sexmaculatus (Coleoptera: Coccinellidae) - Micraspis discolor (Coleoptera: Coccinellidae)
- Nephus sp. (Coleoptera: Coccinellidae)
________________________________________________________________ Sumber: Charernsom and Suasa-ard (1994; 2010).
Senyawa kimia yang dikeluarkan oleh tanaman berperan penting dalam interaksi antara musuh alami dan tanaman. Senyawa kimia yang dikeluarkan oleh tanaman bisa digunakan oleh musuh alami untuk mendeteksi inang atau mangsa. Hal inisecara intensif diteliti pada serangga parasitoid maupun predator tungau (Vet dan Dicke 1992, Dicke dan Vet 1999; Cortesero et al. 2000). Namun baru sedikit informasi tentang interaksi antara serangga predator seperti coccinellids dengan hama tanaman.
Coccinellidae merupakan predator yang banyak digunakan untuk mengendalikan kutu putih (Obata 1986; Zhu et al. 1999). Penelitian pada Coccinellidae, salah satunya telah dilakukan oleh Le Ru and Makayamakosso (2001), diketahui bahwaadanya respon dari betina predator generalis ladybird Epidinocarsis flaviventris pada bau yang dikeluarkan dari interaksi ubikayu dan kutu putih. Tanaman ubi kayu sendiri dikatakan tidak atraktif dalam menarik musuh alami, tetapi dengan percobaan infestasi kutu putih pada tanaman ubi kayu, maka tanaman ubi kayu mengeluarkan senyawa volatil yang menarik betina dari coccinellids pada mikrohabitatnya.
189
DAFTAR PUSTAKA
Ben-Dov Y, Hodgson CJ. 1997. Their Biology, Natural Enemies and Control, Armoured Scale Insects, Vol 4B. Amsterdam: The nederlands.
Bertschy C, Turlings TCJ, Bellotti AC, Dorn S. 1997. Chemically-mediated attraction of three parasitoid species to mealybug-infested cassava leaves. Florida Entomologist.80: 383-395.
Bertschy C, Turlings TCJ, Bellotti AC, Dorn S. 2001. The role of mealybug-induced cassava plant volatiles in the attraction of the Encyrtid parasitoids Aenasiusvexansand Apoanagyrusdiversicornis. J Insect Behav.14: 363-371.
Boussienguet J. 1984. Bio-ecologie de la cochenille du manioc, PhenacoccusmanihotiMat.-Ferr. et de ses ennemis naturels au Gabon. TheseUniversite de Paris Jussieu, Paris, France, 100 p. inPaul-André and Bruno. 2006. Cassava–Mealybug Interactions. Instutute de Recerche Pour Le Developpement. Paris. 112 p.
Catalayud PA, Le RU B. 2006. Cassava-Mealybug interactions. Paris. 112 p. Calatayud PA, Rahbé Y, Delobe1 B, Khuong-Huu E, Tertuliano M, Le RÜ B.
1994. Infhence of secondary compounds in the floem sap of cassava on expression of antibiosis towards the mealybug Phenacoccus manihoti. Entomol exp appl. 72: 47-57.
Charernsom K,Suasaard W. 1994. A host list of natural enemies of insect pests in Thailand. Special Publication No 6.National Biological Control Research Center Kasetsart University/National Research Council of Thailand. 144 pp. (In Thai).
Charernsom K,Suasaard W. 2010. Insects of Thailand: Host and preys of Natural enemies. Special Publication No 9.National Biological Control Research Center Kasetsart University/National Research Council.714 pp. (In Thai).
CIAT 1988. Annual Report Cassava Program. CIAT Calt. Colombia CIAT 1990. Annual Report Cassava Program 1989, CIAT Calt. Colombia CIAT 1992. Annual Report Cassava Program 1987-1991, CIAT Calt.
Colombia.
Cortesero AM, Stapel JO, Lewis WJ. 2000. Understanding and manipulating plant attributes to enhance biological control. BiolCont.17: 35-49.
Cox JM, Williams DJ. 1981. An account of cassava mealybugs (Hemiptera: Pseudococcidae) with a description of a new species. Bull EntomolRes.71: 247-258.
Dicke M, Vet LEM. 1999. Plant-carnivore interactions: evolutionary and ecological consequences for plant, herbivore and carnivore. In:
190
Herbivores: between plants and predators, H. Olff, V. K. Brown & R. H. Drent (eds), Blackwell Science, Oxford, UK, p. 483-520.
Fraenkel, G., 1969. Evaluation of our thoughts on secondary plant substances. Entomol exp appl. 12: 473486.
Herren HR, Hennessey RN. 1983. Biological control and host plant resistance to control the cassava mealybug and green mite in Africa: Proceedings of an international workshop. Ibadan, Nigeria: IITA. 154 p.
Iheagwam EU. 1981. The influence of temperature on increase rates of the cassava mealybug Phenacoccus manihoti Mat.-Ferr.(Homoptera; Pseudococcidae). Rev Zool Afr 95: 959–967.
Iheagwam EU, Eluwa MC. 1983. The effects of temperature on the development of the immature stages of the Cassava Mealybug, Phenacoccus manihoti Mat-Ferr.(Homoptera, Pseudococcidae). Deut Entomol Z 30: 17– 22.
James B, Yaninek J, Neuenschwander P, Cudjoe A, Modder W, Echendu N, Toko M. 2000. Pest Control in Cassava Farms.International Institute of Tropical Agriculture. 36p.
Lema KM, Herren HR. 1985. The influence of constant temperature on population growth rates of the cassava mealybug, Phenacoccus manihoti. Entomol Exp Appl 38: 165–169.
Löhr B, Varela AM, Santos B. 1990. Exploration for natural enemies of the cassava mealybug, Phenacoccus manihoti (Homoptera: Pseudococcidae), in South America for the biological control of this introduced pest in Africa. Bull Entomol Res 80: 417–425.
Le Ru B,Makayamakasso JP. 2001. Prey habitat location by the cassava mealybug predator Exochomusflaviventris: Olfactory responses to odor of plant, mealybug, plantmealybug complex, and plant-mealybug-natural enemy complex. J Insect Behav.14: 557-572.
Le Ru B,Calatayud PA. 1994. Interactions between cassava and arthropod pests. African Crop Sci J.2: 385-390.
Matile-Ferrero D, 1978. Cassava mealybug in the People's Republic of Congo. In: Nwanze KF, Leuschner K, ed. Proceedings of the International Workshop on the cassava mealybug Phenacoccus manihoti Mat.-Ferr. (Pseudococcidae) held at INERA-M'vuazi, Bas-Zaire, Zaire, June 26-29, 1977. International Institute of Tropical Agriculture. Ibadania Niger, 29-46.
Muniappan R, Shepard BM, Watson W, Carner GR, Rauf A, Sartiami D, Hidayat P, Afun JVK, Goergen G, ZiaurRahman AKM. 2011. New Records of Invasive Insects (Hemiptera:Sternorrhyncha) in Southeast Asia and West Africa. JAgricUrban Entomol. 26(4):167-174.
191
Nadel H, van Alphen JJM. 1987. The role of host- and host-plant odours in the attraction of a parasitoid, Epidinocarsislopezi, to the habitat of its host, the cassava mealybug, Phenacoccusmanihoti. EntomolExpAppl. 45: 181-186.
NeuenschwanderP, Schulthess F,Madojemu E. 1986. Experimental evaluation of the efficiency of Epidinocarsislopezi, a parasitoid introduced into Africa against the cassava mealybugPhenacoccusmanihoti. Entomol. Exp Appl. 42: 133-138.
Neuenschwander P. 1994. Control of cassava mealybugs in Africa:lessons from a biological control project. African Crops Sci J.l2(4):369-383.
Neuenschwander P. 2004 Harnessing nature in Africa: biological pest control can benefit the pocket, health and the environment. Nature.432: 801–802. Nwanze KF. 1977. Biology of the cassava mealybugPhenacoccusmanihoti
Mat-Ferr. in the Republic of Zaire. Proceedings of the International Workshop on Cassava MealybugPhenacoccusmanihotiMat-Ferr.(Pseudococcidae). INERA, M’Vuazi-Zaire, June 26-29, IITA Press, Ibadan, Nigeria, p. 20-28. Obata S. 1986. Mechanisms of prey finding in the aphidophagous ladybird
beetle, Harmoniaaxyridis(Coleoptera: Coccinellidae). Entomophaga.31: 303-311.
Rauf A. 2011. Invasive pests. IPM CRSP Annual Report 2010-2011.
Souissi R, Nenon JP, Le Ru B. 1997. Behavioural responses of the endoparasitoidApoanagyruslopezito odours of the host and host’s cassava plants. EntomolExpAppl. 90: 215-220.
Tertuliano M, Dossou-Gbete S, Le Ru B. 1993. Antixenotic and antibiotic components of resistance to the cassava mealybug, Phenacoccusmanihoti(Hom.,Pseudococcidae), in various host-plants. Insect SciAppl.5-6: 657-665.
Van Driesche RG, Bellotti AC, Castillo JA, Herrera CJ. 1990. Estimating total losses from parasitoids for a field population of a countinously breeding insect, cassava mealybug, Phenacoccus herreni (Homoptera : Pseudococcidae) in Colombia, S.A. Florida Entomologist. 73: 133-143.
Vet LEM, Dicke M. 1992. The ecology of infochemical use by natural enemies of herbivores in a tritrophic context. Annu Rev Entomol.37: 141-172.
Williams DJ, Granara de Willink MC. 1992. Mealybugs of Central and South America. CAB International. Wallingford.Oxon, UK, 635 p.
Yaseen M. 1982. Exploration for natural enemies of Phenacoccus manihoti and Mononychellus tanajoa: the challenge, the achievements. In: Herren HR, Hennessey RN, editors. Biological control and host plant resistance to control the cassava mealybug and green mite in Africa: Proceedings of an international workshop. Ibadan, Niger: IITA. pp 81–102.
Zhu J, Cosse AA, Obrycki JJ, Boo KS, Baker TC. 1999. Olfactory reactions of the twelve-spotted lady beetle, Coleomegillamaculataand the green
192
lacewing, Chrysoperlacarneato semiochemicals released from their prey and host plant: electroantennogram and behavioral responses. J ChemEcol.25: 1163-1177.