KARYA TULIS
DINAMIKA SPORULASI
CENDAWAN MIKORIZA
ARBUSKULA
Oleh:
Dr. Delvian, SP.MP.
NIP. 132 299 348
JURUSAN KEHUTANAN
FAKULTAS PERTANIAN
UNIVERSITAS SUMATERA UTARA
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
2006
KATA PENGANTAR
Puji dan syukur Penulis panjatkan ke hadirat Allah SWT atas rahmat dan hidayah-Nya sehingga penulis dapat menyelesaikan tulisan tentang Dinamika Sporulasi Cendawan Mikoriza Arbuskula ini dengan baik.
Tulisan ini berisi informasi tentang dinamika sporulasi cendawan mikoriza arbuskula. Banyak informasi yang menyebutkan bahwa perkembangan cendawan mikoriza arbuskula sangat dipengaruhi oleh pertumbuhan tanaman inangnya danm faktor lingkungan. Dengan kata lain perkembangan cendawan mikoriza mempunyai pola musiman atau bersifat seasonal. Pada sisi lain banyak studi tentang keanekaragaman cendawan mikoriza ini hanya dengan satu kali eksplorasi lapangan dan tidak memperhatikan faktor musim atau iklim. Dalam tulisan ini coba dibahas dengan data-data hasil eksplorasi lapangan tentang pola sporulasi cendawan mikoriza yang bersifat musiman ini.
Penulis berharap tulisan yang sederhana ini dapat bermanfaat sebagai bahan bacaan bagi para mahasiswa yang berminat dan dapat menjadi salah satu sumber referensi dalam melakukan penelitian dalam bidang yang berkaitan.
Akhirnya, pada kesempatan ini Penulis ingin menyampaikan terima kasih kepada semua pihak yang telah memberikan bantuannya dalam penulusuran bahan tulisan ini.
Medan, Juli 2006
Penulis
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
DAFTAR ISI
Halaman KATA PENGANTAR DAFTAR ISI I. PENDAHULUAN 1II. DINAMIKA SPORULASI CENDAWAN MIKORIZA ARBUSKULA 5 III. ULASAN 11 IV. KESIMPULAN 17 DAFTAR PUSTAKA 21
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
DINAMIKA SPORULASI
CENDAWAN MIKORIZA ARBUSKULA
DELVIAN
Departemen Kehutanan Fakultas Pertanian
Universitas Sumatera Utara
Jl. Tri Darma Ujung No. 1 Kampus USU Padang Bulan
M e d a n
e-mail :
dvilly6@yahoo.co.uk
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
PENDAHULUAN
Cendawan mikoriza arbuskula dapat ditemukan hampir pada semua ekosistem di dunia, bahkan lebih dari dua per tiga spesies tanaman yang ada di dunia membentuk simbiosis dengan cendawan ini (Fitter dan Merryweather, 1992). Keberadaan dan keanekaragaman CMA dalam ekosistem dapat meningkatkan keanekaragaman tanaman. Menurut van Der Heijden at el. (1998) keanekaragaman CMA adalah faktor utama yang memelihara keanekaragaman tanaman dan fungsinya dalam ekosistem. Selain itu interaksi CMA dengan mikroba tanah lainnya dapat mengatur fungsi ekosistem seperti keanekaragaman, produktivitas dan variabilitas tanaman. Hal ini menunjukkan betapa pentingnya peranan CMA dan fungsinya dalam pengelolaan lingkungan.
Mempelajari keanekaragaman CMA cukup rumit dan kekhususan inangnya adalah salah satu aspek lain dari keanekaragaman CMA. Jika kekhususan inang dari CMA kecil dalam komunitas yang beragam maka keanekaragaman CMA akan sangat besar. Sebaliknya jika kekhususan ini sangat tinggi maka akan mengurangi keanekaragaman CMA dalam suatu ekosistem (Abbott dan Gaezy, 1994). Di samping itu stabilitas atau perubahan komunitas CMA juga sangat penting dalam menentukan keanekaragaman dan keberlanjutan dari tanaman yang obligat mikotrofik. Lebih lanjut, keberadaan CMA pada suatu ekosistem ditentukan oleh komposisi dan keberadaan vegetasi yang menjadi inangnya.
Keberadaan dan peranan CMA telah dipelajari secara ekstensif pada beberapa komunitas tanaman, seperti di hutan tropis (Pacioni, 1986) dan bukit-bukit pasir (Puppi, 1986 ; Louis, 1990 ; Semones dan Young, 1995 ; Siguenza et
al., 1996), tetapi informasi tentang ekologi CMA masih sangat kurang.
Faktor-faktor yang mempengaruhi ketahanan CMA, perkecambahan spora, kolonisasi
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
akar, dan pembentukan spora CMA akan menentukan keberadaan dan keanekaragaman CMA di alam. Oleh karena itu studi yang mengarah pada faktor-faktor yang mempengaruhi perkembangan CMA masih perlu terus dikembangkan. Pemahaman akan hal ini sangat penting dalam pemanfaatan CMA guna meningkatkan produksi pertanian dalam arti luas.
Menurut Johnson et al. (1982) sporulasi CMA terjadi sebagai rsepon terhadap fluktuasi pertumbuhan akar, akan tetapi produksi spora mungkin meningkat setelah periode pertumbuhan akar yang ekstensif atau penuaan dan proses senescen tanaman inang. Perbedaan tanaman inang dan kesuburan tanah akan memberikan pengaruh yang berbeda terhadap sporulasi setiap spesies (Hayman, 1975; McGraw dan Hendrix, 1984). Di samping itu di beberapa habitat ditemukan bahwa pembentukan spora CMA bersifat musiman (Bargett et al., 1999; Mohammad et al., 1998; Puppi et al., 1986; Siguenza et al., 1996).
Umumnya studi tentang variasi musiman dalam populasi CMA didasarkan pada jumlah spora yang diisolasi (Abbott dan Gazey, 1994). Apakah puncak produksi spora terjadi pada musim semi-musim panas atau musim panas-musim gugur tampaknya berhubungan dengan iklim dan tanaman tetapi penurunan jumlah spora yang nyata terjadi selama musim dingin. Selanjutnya inokulum yang mampu bertahan selama musim dingin akan segera mengkolonisasi akar tanaman pada musim semi. Jumlah spora meningkat selama musim pertumbuhan dan kemudian menurun dengan berlalunya musim semi.
Saif (1977) dalam Hetrick (1984) membuat model siklus biologi CMA pada tanaman semusim yang dibagi menjadi 3 fase. Fase pertama atau lag fase, berhubungan dengan perkecambahan spora CMA yang diikuti oleh penetrasi ke dalam akar yang merupakan proses awal kolonisasi. Selama fase
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
ini jumlah spora tinggi sebaliknya persentase kolonisasi masih rendah. Pada fase kedua, kolonisasi menyebar dalam kortek akar dan fase ini ditandai dengan kelimpahan arbuskula dalam akar tanaman.
Fase ketiga atau fase konstan, berhubngan dengan fase reproduktif tanaman inang. Selama fase ini kolonisasi berada pada level konstan dan umumnya terjadi pembentukan vesikula. Pada saat yang sama jumlah spora yang terbentuk di daerah perakaran meningkat. Ini adalah bentuk pertahanan hidup CMA dalam keadaan tidak aktif, sampai pembentukan asosiasi mikoriza yang baru (Puppi et al., 1986).
Banyak studi yang menunjukkan bahwa perkembangan CMA dipengaruhi oleh musim atau bersifat musiman. Tidak sedikit pula yang melaporkan bahwa perkembangan CMA tidak bersifat musiman. Gay et al. (1982) mempelajari pengaruh musim terhadap kolonisasi CMA pada Abronia
umbellata dan Camissonia californica. Hasilnya menunjukkan kolonisasi CMA
berkurang selama atau sesudah periode pembungaan, yaitu antara April – Juni untuk Abronia umbellata dan Mei – Nopember untuk Camissonia californica.
Penelitian lain yang dilakukan Saif dan Khan (1975) dan Bethlenfalvay
et al. (1982) mempelajari penghentian pertumbuhan CMA berhubungan dengan
fase reproduktif tanaman. Selama musim dingin CMA umumnya ditemukan dalam akar tanaman sementara jumlah spora di daerah perakaran lebih sedikit, dan sebaliknya pada musim panas dimana pembentukan spora akan meningkat.
Hasil yang berbeda dilaporkan oleh Sparling dan Tinker (1978) untuk padang rumput di dataran tinggi, dimana kolonisasi CMA tidak berfluktuasi dengan musim. Persentase kolonisasi CMA pada rumput di latitud rendah (Pond
et al., 1984; Puppi et al., 1986) dan pada semua seri bukit pasir di laut Adriatik
(Pacioni, 1986) adalah tetap sepanjang tahun.
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
Rozema et al. (1986) menyatakan bahwa distribusi CMA di salt marsh di Netherlands lebih dipengaruhi oleh filogeni tanaman inang (famili tanaman bermikoriza versus tidak bermikoriza) daripada posisi spasial di lapangan. Sedangkan Johnson-Green et al. (1995) melaporkan bahwa distribusi dan aktivitas CMA lebih berhubungan dengan perbedaan fenologi pertumbuhan akar, bukan faktor iklim atau tanah.
Belum ada informasi tentang pola sporulasi CMA di daerah tropis khususnya di ekosistem hutan pantai, apakah juga bersifat musiman mengingat untuk daerah tropis tidak adanya perbedaan musim yang cukup tegas. Diduga setiap jenis CMA mempunyai pola pembentukan spora dan kolonisasi yang berbeda sebagai respon terhadap perubahan musim.
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
DINAMIKA SPORULASI
CENDAWAN MIKORIZA ARBUSKULA
Dinamika sporulasi CMA dapat dilihat dengan melakukan pengukuran jumlah spora dan penentuan jenis CMA yang ada. Untuk dapat melihat dinamika sporulasi CMA maka pengamatan/pengukuran harus dilakukan secara periodik dalam waktu satu tahun. Di samping itu perlu juga dihitung persentase kolonisasi yang terjadi pada tanaman inang di lapangan karena besarnya persentase kolonisasi juga dipengaruhi oleh kondisi musim dan ada hubungannya dengan proses pembentukan spora atau sporulasi.
Pada kondisi basah atau banyak hujan umumnya persentase kolonisasi meningkat dan pembentukan spora baru berkurang. Hal ini disebabkan karena kelembaban tanah yang tinggi pada kondisi basah akan merangsang perkecambahan spora dan terbentuknya kolonisasi dengan tanaman inang. Sebaliknya pada kondisi kering atau sedikit hujan pembentukan spora baru akan meningkat dan persentase kolonisasi akan menurun. Kondisi kering akan merangsang pembentukan spora yang banyak sebagai respon alami dari CMA serta upaya untuk mempertahankan keberadaannya di alam.
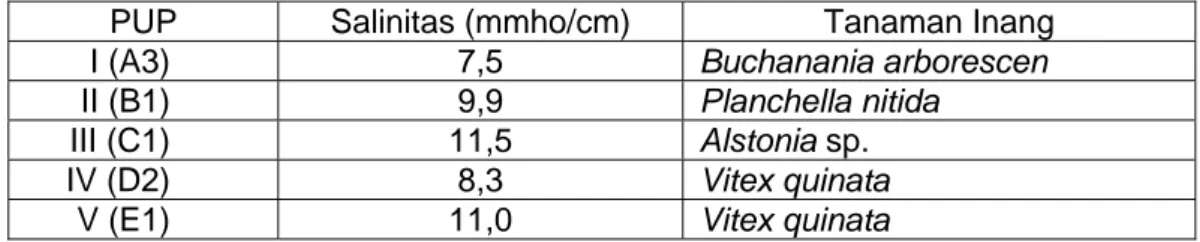
Dalam studi ini jenis tanaman sampel untuk pengukuran persentase kolonisasi dan kepadatan spora pada setiap PUP berbeda, kecuali pada PUP IV dan V. Data selengkapnya disajikan dalam Tabel 1 berikut.
Tabel 1. Jenis tanaman sampel dan tingkat salinitas setiap PUP
PUP Salinitas (mmho/cm) Tanaman Inang
I (A3) 7,5 Buchanania arborescen
II (B1) 9,9 Planchella nitida
III (C1) 11,5 Alstonia sp.
IV (D2) 8,3 Vitex quinata
V (E1) 11,0 Vitex quinata
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
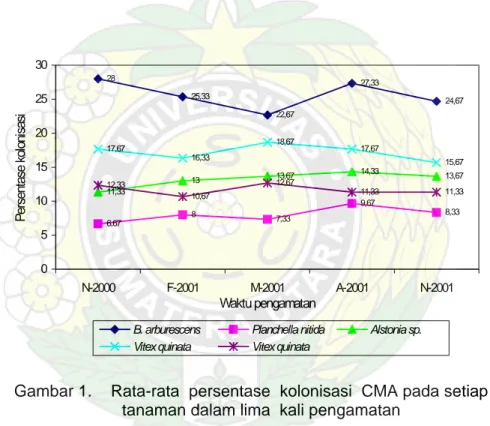
Persentase kolonisasi dan kepadatan spora CMA bervariasi dan berfluktuasi pada setiap tanaman dalam setiap pengambilan contoh tanah. Setiap tanaman mempunyai pola fluktuasi yang berbeda yang tidak menunjukkan suatu kecenderungan tertentu terhadap perbedaan waktu pengamatan. Akan tetapi variasi persentase kolonisasi dan kepadatan spora dipengaruhi oleh tingkat salinitas tanah. CMA. Hasil penghitungan persentase kolonisasi dan kepadatan spora CMA pada setiap tanaman disajikan pada Gambar 1 dan Gambar 2 sedangkan data jumlah curah hujan pada Gambar 3.
28 25,33 22,67 27,33 24,67 6,67 8 7,33 9,67 8,33 11,33 13 13,67 14,33 13,67 17,67 16,33 18,67 17,67 15,67 12,33 10,67 12,67 11,33 11,33 0 5 10 15 20 25 30 N-2000 F-2001 M-2001 A-2001 N-2001 Waktu pengamatan Per sentas e k ol oni sas i
B. arburescens Planchella nitida Alstonia sp.
Vitex quinata Vitex quinata
Gambar 1. Rata-rata persentase kolonisasi CMA pada setiap tanaman dalam lima kali pengamatan
Pada B. arburescens persentase kolonisasi cenderung menurun dengan berkurangnya curah hujan kecuali pada pengamatan keempat (A-2001) dengan persentase kolonisasi sebesar 27,33%. Sementara itu persentase kolonisasi CMA untuk Vitex quinata pada PUP IV dan PUP V mempunyai pola yang hampir sama. Puncak kolonisasi terjadi pada pengamatan ketiga (M-2001) masing-masing sebesar 18,67% dan 16,67%.
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
Pada Alstonia sp. persentase kolonisasi cenderung tetap tanpa perubahan yang cukup berarti dengan tingkat kolonisasi berkisar antara 11,33-14,33%. Tanaman Planchella nitida mempunyai persentase kolonisasi terkecil dibandingkan empat jenis tanaman lainnya dan fluktuasi kolonisasi yang terjadi pada setiap pengamatan relatif rendah (6,67-9,67%).
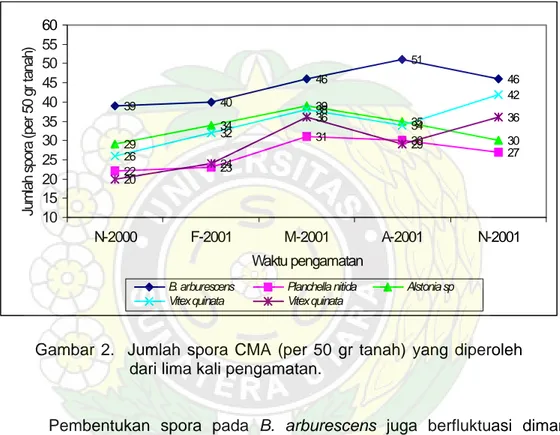
39 40 46 51 46 22 23 31 30 27 29 34 39 35 30 26 32 38 34 42 20 24 36 29 36 10 15 20 25 30 35 40 45 50 55 60 N-2000 F-2001 M-2001 A-2001 N-2001 Waktu pengamatan
Jumlah spora (per 50 gr tanah)
B. arburescens Planchella nitida Alstonia sp
Vitex quinata Vitex quinata
Gambar 2. Jumlah spora CMA (per 50 gr tanah) yang diperoleh dari lima kali pengamatan.
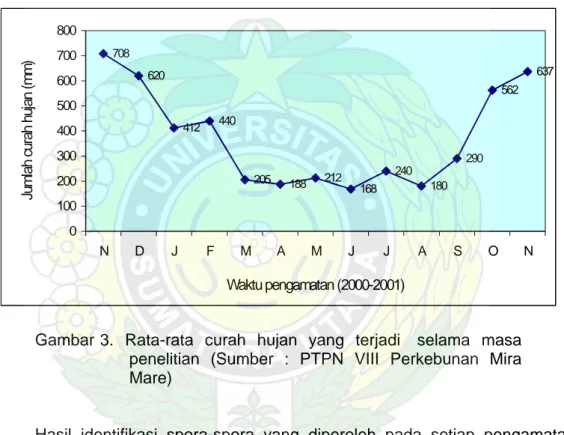
Pembentukan spora pada B. arburescens juga berfluktuasi dimana pembentukan spora cenderung meningkat dengan berkurangnya curah hujan dan jumlah spora terbanyak diperoleh pada pengamatan ke-empat (51 spora per 50 gr tanah). Dilihat dari perubahan musim (curah hujan) produksi spora CMA terjadi pada periode kering dengan curah hujan berkisar antara 212-440 mm (Gambar 3).
Untuk tanaman Vitex quinata jumlah spora terbanyak juga diperoleh pada pengamatan ketiga masing-masing 38 spora dan 36 spora per 50 gr tanah. Sedangkan pada Alstonia sp. produksi spora CMA berfluktuasi dengan waktu
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
pengamatan dimana jumlah spora terbanyak diperoleh pada pengamatan ke-tiga (M-2001), yaitu 39 spora per 50 gr tanah. Dari Gambar 14 terlihat bahwa pembentukan spora CMA cenderung meningkat dengan berkurangnya curah hujan, meskipun fluktuasi peningkatannya bervariasi antar jenis tanaman. Tidak demikian halnya dengan pembentukan spora pada Planchella nitida dimana jumlah spora terus meningkat dengan perubahan waktu pengamatan.
708 620 412 440 205 188 212 168 240 180 290 562 637 0 100 200 300 400 500 600 700 800 N D J F M A M J J A S O N Waktu pengamatan (2000-2001)
Jumlah curah hujan (mm)
Gambar 3. Rata-rata curah hujan yang terjadi selama masa penelitian (Sumber : PTPN VIII Perkebunan Mira Mare)
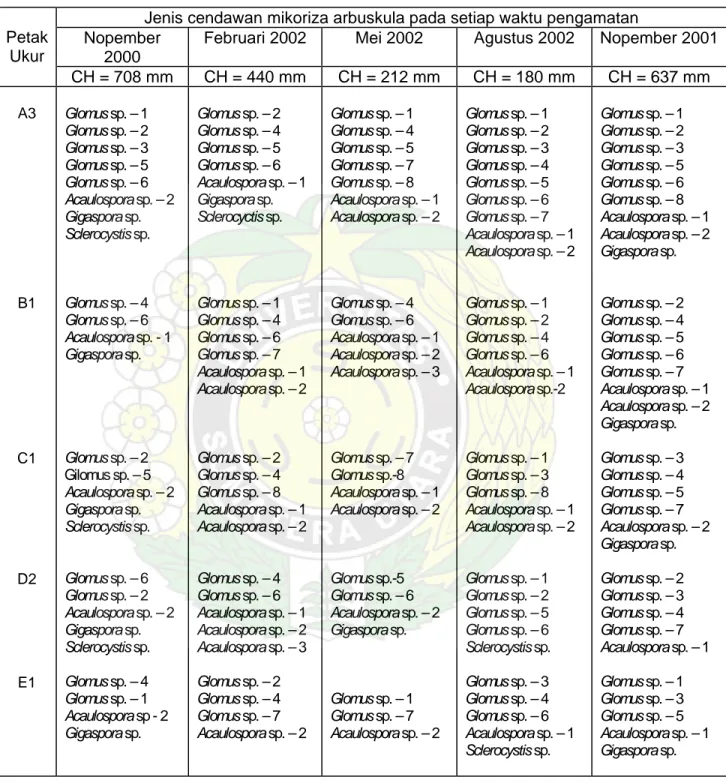
Hasil identifikasi spora-spora yang diperoleh pada setiap pengamatan menunjukkan terjadinya perubahan tipe spora yang terbentuk, seperti terlihat pada Tabel 2. Dari lima PUP dengan lima kali pengamatan tampak tidak ada satu pun PUP yang mempunyai komposisi tipe CMA yang persis sama. Setiap kali pengamatan jenis spora CMA yang diperoleh berbeda dengan jenis CMA yang diperoleh pada pengamatan sebelumnya.
Jumlah tipe spora yang diperoleh pada setiap PUP berbeda, baik antar PUP dalam waktu pengamatan yang sama maupun dalam PUP yang sama tetapi
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
Tabel 8. Dinamika jenis cendawan mikoriza arbuskula yang diperoleh pada setiap petak ukur permanen dalam lima kali pengamatan
Jenis cendawan mikoriza arbuskula pada setiap waktu pengamatan Nopember
2000
Februari 2002 Mei 2002 Agustus 2002 Nopember 2001 Petak Ukur CH = 708 mm CH = 440 mm CH = 212 mm CH = 180 mm CH = 637 mm A3 B1 C1 D2 E1 Glomus sp. – 1 Glomus sp. – 2 Glomus sp. – 3 Glomus sp. – 5 Glomus sp. – 6 Acaulospora sp. – 2 Gigaspora sp. Sclerocystis sp. Glomus sp. – 4 Glomus sp. – 6 Acaulospora sp. - 1 Gigaspora sp. Glomus sp. – 2 Gilomus sp. – 5 Acaulospora sp. – 2 Gigaspora sp. Sclerocystis sp. Glomus sp. – 6 Glomus sp. – 2 Acaulospora sp. – 2 Gigaspora sp. Sclerocystis sp. Glomus sp. – 4 Glomus sp. – 1 Acaulospora sp - 2 Gigaspora sp. Glomus sp. – 2 Glomus sp. – 4 Glomus sp. – 5 Glomus sp. – 6 Acaulospora sp. – 1 Gigaspora sp. Sclerocyctis sp. Glomus sp. – 1 Glomus sp. – 4 Glomus sp. – 6 Glomus sp. – 7 Acaulospora sp. – 1 Acaulospora sp. – 2 Glomus sp. – 2 Glomus sp. – 4 Glomus sp. – 8 Acaulospora sp. – 1 Acaulospora sp. – 2 Glomus sp. – 4 Glomus sp. – 6 Acaulospora sp. – 1 Acaulospora sp. – 2 Acaulospora sp. – 3 Glomus sp. – 2 Glomus sp. – 4 Glomus sp. – 7 Acaulospora sp. – 2 Glomus sp. – 1 Glomus sp. – 4 Glomus sp. – 5 Glomus sp. – 7 Glomus sp. – 8 Acaulospora sp. – 1 Acaulospora sp. – 2 Glomus sp. – 4 Glomus sp. – 6 Acaulospora sp. – 1 Acaulospora sp. – 2 Acaulospora sp. – 3 Glomus sp. – 7 Glomus sp.-8 Acaulospora sp. – 1 Acaulospora sp. – 2 Glomus sp.-5 Glomus sp. – 6 Acaulospora sp. – 2 Gigaspora sp. Glomus sp. – 1 Glomus sp. – 7 Acaulospora sp. – 2 Glomus sp. – 1 Glomus sp. – 2 Glomus sp. – 3 Glomus sp. – 4 Glomus sp. – 5 Glomus sp. – 6 Glomus sp. – 7 Acaulospora sp. – 1 Acaulospora sp. – 2 Glomus sp. – 1 Glomus sp. – 2 Glomus sp. – 4 Glomus sp. – 6 Acaulospora sp. – 1 Acaulospora sp.-2 Glomus sp. – 1 Glomus sp. – 3 Glomus sp. – 8 Acaulospora sp. – 1 Acaulospora sp. – 2 Glomus sp. – 1 Glomus sp. – 2 Glomus sp. – 5 Glomus sp. – 6 Sclerocystis sp. Glomus sp. – 3 Glomus sp. – 4 Glomus sp. – 6 Acaulospora sp. – 1 Sclerocystis sp. Glomus sp. – 1 Glomus sp. – 2 Glomus sp. – 3 Glomus sp. – 5 Glomus sp. – 6 Glomus sp. – 8 Acaulospora sp. – 1 Acaulospora sp. – 2 Gigaspora sp. Glomus sp. – 2 Glomus sp. – 4 Glomus sp. – 5 Glomus sp. – 6 Glomus sp. – 7 Acaulospora sp. – 1 Acaulospora sp. – 2 Gigaspora sp. Glomus sp. – 3 Glomus sp. – 4 Glomus sp. – 5 Glomus sp. – 7 Acaulospora sp. – 2 Gigaspora sp. Glomus sp. – 2 Glomus sp. – 3 Glomus sp. – 4 Glomus sp. – 7 Acaulospora sp. – 1 Glomus sp. – 1 Glomus sp. – 3 Glomus sp. – 5 Acaulospora sp. – 1 Gigaspora sp.
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
USU Repository©2006
waktu pengamatan yang berbeda. Secara umum dalam lima kali pengamatan jumlah tipe spora pada PUP A3 selalu lebih banyak daripada empat PUP lainnya.
Jika diamati perubahan tipe spora yang terdapat pada suatu PUP antar waktu pengamatan tampak frekuensi munculnya suatu tipe spora sangat bervariasi. Ada tipe spora yang dapat ditemukan pada setiap waktu pengamatan, yaitu Acaulospora sp.-2 dan ada yang hanya ditemukan dalam satu kali pengamatan, yaitu Acaulospora sp.-3.
Perubahan tipe spora CMA yang ada pada setiap pengamatan tampaknya berhubungan dengan jumlah spora yang diperoleh pada setiap PUP dan waktu pengamatan. Bertambahnya tipe spora CMA yang diperoleh akan diikuti oleh peningkatan jumlah spora yang terbentuk. Jika kita lihat pada PUP A3, pada pengamatan pertama (N-2000) diperoleh 8 jenis CMA dengan 32 spora per 50 gr tanah. Kemudian pada pengamatan kedua (F-2001) diperoleh 10 jenis CMA dengan 58 spora per 50 gr tanah. Dari kedua pengamatan tersebut diperoleh penambahan 4 jenis baru, yaitu Glomus sp.-3, Glomus sp.-4, Glomus sp.-5, dan Acaulospora sp.-1, tetapi kehilangan dua jenis, yaitu Glomus sp.-8 dan
Gigaspora sp. Dari penambahan jenis CMA ini diperoleh penambahan 26 spora
baru.
Dari lima kali pengamatan pada lima PUP diperoleh total 13 tipe spora. Tipe spora Glomus adalah yang paling dominan, yaitu 8 tipe spora diikuti oleh
Acaulospora dengan 3 tipe spora serta Gigaspora dan Sclerocystis
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
USU Repository©2006
ULASAN
Keanekaragaman jenisCMA (dilihat dari tipe spora yang terbentuk) di hutan pantai cukup tinggi, dimana pada satu individu pohon yang berbeda ditemukan 3-10 tipe spora. Total tipe spora yang ditemukan dalam lima kali pengamatan adalah 13 tipe spora, terdiri atas 8 tipe spora Glomus, 3 tipe spora Acaulospora dan masing-masing 1 tipe spora Gigaspora dan Scelrocyctis.
Seperti diketahui bahwa CMA mempunyai kisaran jenis inang yang sangat luas (Smith dan Read, 1987) dan tidak ada kekhususan inang untuk membentuk simbiosis (Abbott dan Gazey, 1994). Dengan demikian dapat dibayangkan betapa besar keanekaragaman CMA yang mungkin ada di hutan pantai. Terlebih lagi jika dilihat data keanekaragaman ini hanya diperoleh dari 1 jenis inang dalam luasan areal yang kecil (5x20 m) dengan radius 0-60 m dari garis pantai. Di samping itu interval waktu antar pengamatan juga masih cukup lebar, yaitu 3 bulan.
Keanekaragaman tipe spora CMA selalu berubah dengan perubahan waktu pengamatan, jenis inang dan tingkat salinitas (ditunjukkan oleh letak petak ukur). Hal ini menunjukkan bahwa keanekaragaman CMA dipengaruhi oleh faktor lingkungan dan tanaman inang (Johnson-Green et al., 1995; Siguenza et al., 1996). Faktor lingkungan di sini dibedakan menjadi tingkat salinitas tanah dan musim (curah hujan).
Secara umum salinitas berpengaruh negatif terhadap keanekaragaman tipe spora yang ada (lihat bab Status dan Kelimpahan Cendawan Mikoriza Arbuskula di Hutan Pantai Berdasarkan Gradien Salinitas). Perubahan musim (curah hujan) juga mempengaruhi komposisi tipe spora yang ditemukan pada suatu PUP.
Pengaruh perubahan musim ini berhubungan dengan aktivitas tanaman inang dan CMA itu sendiri (McGraw dan Hendrix, 1984). CMA adalah simbion obligat sehingga semua faktor yang mempengaruhi tanaman inang juga akan mempengaruhi CMA sebagai simbionnya (Smith dan Read, 1997). Kondisi terbaik bagi pertumbuhan dan perkembangan inang akan memberikan pertumbuhan dan perkembangan terbaik bagi CMA.
Di samping itu sebagai individu setiap CMA mempunyai faktor intrinsik (Ocampo et al., 1986) yang akan mempengaruhi responya terhadap perubahan musim. Hal ini ditunjukkan oleh frekuensi munculnya suatu tipe spora dengan perubahan musim. Ada tipe spora yang selalu ditemukan dalam setiap pengamatan yang menunjukkan bahwa aktivitas tipe spora ini tidak dipengaruhi oleh perubahan musim, tetapi ada juga yang hanya muncul dalam satu kali pengamatan saja. Hal ini mempertegas bahwa pengaruh perubahan musim terhadap aktivitas CMA tergantung pada tipe spora (faktor intrinsik). Hasil ini sejalan dengan penelitian yang dilakukan Sparling dan Tinker (1978), Pond et al. (1984), Puppi et al. (1986), dan Pacioni (1986).
Hasil penelitian ini mungkin belum dapat memberikan gambaran pasti tentang potensi keanekaragaman CMA di hutan pantai. Akan tetapi hasil ini lebih komprehensif karena data diperoleh dari lima kali pengamatan dan data yang ada juga menunjukkan adanya perubahan jumlah dan tipe spora CMA pada setiap pengamatan. Selama ini informasi tentang keanekaragaman CMA pada suatu ekosistem atau tegakan atau individu pohon diperoleh hanya dari satu kali pengamatan, seperti yang dilakukan oleh Ervayenri (1998) dan Ekamawanti (1999) pada ekosistem gambut, Purwanto (1999) pada ekosistem hutan pantai, Maryadi (2002) pada tegakan jati, dan Silviana et al. (1999) pada rizosfir manggis. Hall
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
(1984) menyatakan bahwa jumlah dan jenis CMA yang dihasilkan setiap tahunnya mungkin tidak sama dan ada kecenderungan satu atau beberapa genus CMA sangat terbatas penyebarannya. Oleh karena itu sporokarp atau spora yang terkumpul dari satu wilayah dalam suatu waktu tertentu mungkin tidak mewakili seluruh spora yang ada dari jenis CMA yang ada pada wilayah tersebut.
Dari lima kali pengamatan selama kurun waktu satu tahun diperoleh hasil bahwa persentase kolonisasi dan produksi spora bervariasi dan berfluktuasi pada setiap tanaman dan petak ukur. Variasi persentase kolonisasi dan jumlah spora antar jenis tanaman tampaknya menunjukkan pola yang tidak sama terhadap perubahan musim (curah hujan). Hasil ini menunjukkan bahwa dinamika kolonisasi dan pembentukan spora atau sporulasi lebih berhubungan dengan tanaman inang daripada perubahan musim. Menurut Johnson-Green et al. (1995), jika simbiosis antara CMA dan inangnya adalah sesuatu yang penting maka waktu pertumbuhan terbaik dari tanaman inang adalah merupakan puncak aktivitas CMA.
Jika melihat respon kolonisasi pada setiap jenis tanaman terhadap waktu pengamatan tampak bahwa pola perkembangan kolonisasi setiap tanaman berbeda. Menurut Puppi et al. (1996) kolonisasi CMA pada tanaman berhubungan dengan situasi tanaman inang sebagai responnya terhadap kondisi lingkungan. Umumnya pada daerah dengan salinitas tinggi kolonisasi CMA sangat jarang dan kalupun terjadinya persentasenya rendah. Pada sisi lain Abbott dan Gazey (1994) menyatakan bahwa kolonisasi CMA lebih tinggi pada kondisi ketersediaan air cukup. Pada keadaan ini persentase dan kecepatan perkecambahan spora meningkat (Ocampo et al., 1986) sehingga kolonisasi pada akar tanaman juga akan meningkat. Adanya curah hujan yang tinggi pada daerah bersalinitas tinggi diharapkan akan dapat meningkatkan kolonisasi akar (Puppi et al., 1996; Abbott dan Gazey,
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
1994). Ada dua pendekatan untuk hal ini, yaitu curah hujan yang tinggi diharapkan akan mengurangi konsentrasi garam dalam tanah. Menurut Young (1976), air hujan dapat dengan cepat membuat garam-garam pada tanah tercuci. Berkurangnya kadar garam dalam tanah ini diharapkan akan mengurangi hambatan bagi proses kolonisasi akar. Selain itu adanya air yang cukup dari curah hujan akan membantu proses perkecambahan spora CMA (Clark, 1997) sehingga juga akan dapat meningkatkan kolonisasi akar.
Dari hasil penelitian ini secara umum tidak terjadi fluktuasi persentase kolonisasi CMA yang besar dengan perubahan musim (curah hujan), kecuali pada tanaman B. arburescens dimana persentase kolonisasi cenderung menurun dengan berkurangnya curah hujan. Tidak adanya respon persentase kolonisasi yang tegas dengan perubahan curah hujan ini mungkin berhubungan dengan masalah contoh akar tanaman yang diamati. Contoh akar tanaman diambil dari tanaman yang berbeda pada setiap periode pengamatan, meskipun dari jenis yang sama. Perbedaan asal akar tanaman ini diduga menjadi alasan mengapa fluktuasi persentase kolonisasi akar terhadap perubahan curah hujan tidak tegas.
Seperti halnya kolonisasi akar, pembentukan spora atau sporulasi CMA juga dipengaruhi oleh faktor lingkungan, tanaman inang dan cendawan lain. Menurut Bardgett et al. (1999) dalam banyak kasus faktor-faktor yang merangsang atau menghambat proses kolonisasi akar akan juga merangsang atau menghambat pembentukan spora CMA. Akan tetapi walaupun kolonisasi akar dan pembentukan spora mempunyai hubungan yang erat, menurut Abbott dan Gazey (1994) kedua fenomena ini tidak dapat dikatakan selalu mempunyai hubungan yang positif.
Dalam penelitian ini ada kecenderungan peningkatan jumlah spora dengan berkurangnya jumlah curah hujan. Menurut Lewis (1986) fluktuasi kelembaban
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
tanah dapat mempengaruhi pembentukan spora atau sporulasi, mungkin karena hifa eksternal dipengaruhi secara drastis daripada hifa di dalam kortek akar. Sedangkan Hernandez et al. (1986) menyatakan bahwa cekaman air pada tanaman bermikoriza akan menginduksi peluruhan (senescen) miselia CMA, keadaan ini akan memacu pembentukan spora lebih awal.
Banyak penelitian di daerah temperate yang menghubungkan produksi spora CMA dengan fenologi tanaman inang. Menurut Siguenza et al. (1996) selama fase perkembangan buah jumlah karbon yang tersedia bagi CMA akan berkurang hal ini menyebabkan produksi spora atau kolonisasi akan menurun. Produksi spora paling tinggi terjadi pada akhir musim pertumbuhan (Dehne, 1986), dan kecenderungan ini telah dilaporkan oleh banyak peneliti (Hayman, 1970 ; Sutton dan Barron, 1972 ; Ebbers et al., 1987).
Untuk daerah tropis, seperti Indonesia, pembentukan spora CMA pada tanaman tahunan mungkin tidak dapat dikaitkan secara langsung dengan fenologi tanaman inang ataupun perubahan musim. Sulit untuk menentukan fenologi yang tegas dari tegakan yang sudah stabil sehingga untuk mencari hubungan antara fenologinya dengan perkembangan CMA cukup rumit. Begitu juga dengan pengaruh musim terhadap perkembangan CMA, dimana tidak adanya perbedaan musim yang tegas seperti halnya di daerah dengan empat musim akan menyulitkan dalam mempelajari hubungan musim dengan produksi spora.
Meskipun banyak penelitian yang melaporkan bahwa cekaman air akan merangsang pembentukan spora CMA, belum dapat disimpulkan bahwa kondisi kering akan selalu menghasilkan spora yang lebih banyak. Penelitian Sieverding dan Toro (1988) memberikan fenomena lain dimana mereka mempelajari pengaruh cekaman terhadap tujuh jenis CMA (Acaulospora longula, A. myriocarpa,
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
Entrophospora colombiana, Glomus fasciculatum, G. manihotis, G. occultum, dan Scutellospora heterogama). Pada akhir penelitian jumlah total spora yang terbentuk
dari semua jenis CMA yang digunakan menurun secara signifikan pada kondisi kering, kecuali pada S. Heterogama yang relatif meningkat dengan perlakuan pengeringan. Hal ini terjadi karena perlakuan kering akan menurunkan produksi bahan kering tanaman, maka produksi spora CMA juga akan menurun.
Dari hasil-hasil penelitian yang ada tampak bahwa pengaruh curah hujan terhadap pembentukan spora sangat tergantung dari jenis CMA yang ada, dalam hal iini faktor intrinsik CMA lebih berperan (Ocampo et al., 1986). Mungkin terdapat perbedaan dalam kebutuhan air bagi setiap CMA untuk tahapan perkembangannya sehingga hubungan antara ketersediaan air tanah dengan pembentukan spora belum dapat disimpulkan secara lebih tegas.
Adanya perubahan tipe spora CMA dalam setiap pengamatan menunjukkan bahwa setiap jenis CMA membentuk spora pada saat yang berbeda, tergantung fenologi dan responnya terhadap tanaman inang. Di samping itu hal ini memberikan informasi bahwa keanekaragaman CMA pada suatu lokasi tergantung pada jenis CMA yang bersporulasi pada saat tersebut, meskipun belum menggambarkan keanekaragaman CMA yang sesungguhnya.
Dengan demikian apabila kita ingin mengetahui keanekaragaman CMA pada suatu tegakan atau ekosistem maka harus dilakukan pengamatan secara periodik. Dengan demikian peluang untuk mendapatkan semua jenis CMA yang ada pada suatu tegakan atau ekosistem akan lebih besar, sehingga informasi yang kita dapatkan tentang keanekaragaman CMA yang ada lebih lengkap.
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
KESIMPULAN
Dari hasil penelitian ini dapat diambil kesimpulan sebagai berikut:
1. Keanekaragaman CMA di hutan pantai cukup tinggi dimana pada satu individu pohon terdapat 3-10 tipe spora CMA.
2. Glomus adalah tipe spora CMA yang paling banyak ditemukan pada ekosistem hutan pantai.
3. Terdapat dinamika sporulasi CMA di hutan pantai sehingga untuk mengetahui keanekaragaman CMA yang ada harus dilakukan pengamatan secara periodik.
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
DAFTAR PUSTAKA
Abbott LK dan Gazey C. 1994. An ecological view of the formation of VA mycorrhizas. Plant and Soil 159 : 69-78
Bardgett RD, Lovell RD, Hobbs PJ, dan Jarvis SC. 1999. Seasonal changes in soil microbial communities along a fertility gradient of temperate grasslands. Soil Biology and Biochemistry 31 : 1021-1030
Bethlenfalvay GJ, Pakovsky RS, Brown MS, dan Fuller G. 1982. Mycrotrophic growth and mutualistic development of host plant and fungal endophyte in an endomycorrhizal symbiosis. Plant Soil 68 : 43-54
Clark RB. 1997. Arbuscular mycorrhizal adaptation, spore germination, root colonization, and hoast plant growth and mineral acquisition at low pH. Plant and Soil 192 : 15-22
Dehne HW. 1986. Influence of VA mycorrhizae on host plant physiology. Di Dalam : Gianinazzi-Pearson V dan Gianinazzi S (Eds). Physiological and genetical aspect of mycorrhizae. Proceeding of the 1st Europens Symposium on Mycorrhizae. Hal. 431-435
Ebbers BC, Anderson RC dan Liberta AE. 1987. Aspect of the mycorrhizal ecology of prairie dropseed Sporobolus heterolepis (Poaceae). Am. J. Bot. 74 : 564-573
Ekamawanti HA. 1999. Biodiversity of Arbucular mycorrhizal fungi in peat ecosystems in West Kalimantan. Di Dalam : Smith FA, Kramadibrata K, Simanungkalit RDM, Sukarno N, dan Nuhamara ST (Eds.) Mycorrhizas in sustainable tropical agriculture and forest ecosystems. Proceedings of International Conference on Mycorrhiza. Bogor, Indonesia. Hal. 77-84 Ervayenri. 1998. Studi keanekaragaman dan potensi inokulan cendawan mikoriza
arbuskula (CMA) di lahan gambut (studi kasus di Kabupaten Bengkalis, Propinsi Riau). Tesis. Program Pascasarjana IPB. Bogor.
Fitter AH dan Merryweather JW. 1992. Why are some plants more mycorrhizal than others? An ecological enquiry. ?. Di Dalam : Read DJ, Lewis DH, Fitter AH, dan Alexander IJ (Eds). Mycorrhizas in ecosystems. C.A.B. International. Hal. 26-36
Gay pe, Grubb PJ dan Hudson HJ. 1982. Seasonal changes in the consentrations of nitrogen phosphorus and potassium and in the density of mycorrhiza in biennial and matrix-forming perennial species of closed chalkland turf. J. Ecol. 70 : 571-593
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
Giovannetti M dan Mosse B. 1980. An evaluation of technique for measuring vesicular-arbuscular mycorrhizal infection in roots. New Phytol 84 : 489-500
Hayman DS. 1970. Endogone spore numbers in soil and vesicular arbuscular mycorrhiza in wheat as influenced by season and soil treatment. Trans. Br. Mycol. Soc. 54 : 53-63
Hayman DS. 1975. The occurrence of mycorrhizas in field crops as affected by soil fertility. Di dalam : Sanders FE, Mosse B dan Tinker PB (Eds.). Endomycorrhizas. Academic Press. New York. Hal. 495-509
Hernandez AP, El-Sharkawy, Sieverding E, dan Toro S. 1986. influence of water stress on growth and formation of VA mycorrhiza of 20 cassava cultivars. Di Dalam : Gianinazzi-Pearson V dan Gianinazzi S (Eds). Physiological and genetical aspect of mycorrhizae. Proceeding of the 1st Europens Symposium on Mycorrhizae. Hal. 717-720
Hetrick BAD. 1984. Ecology of Vesicular-Arbuscular Mycorrhiza Fungi. pp. 35-56. In : Powell CL and Bagyaraj DJ. (Eds). Vesicular-Arbuscular Mycorrhiza. CRC Press. Inc. Boca Raton. Florida.
Johnson CR, Menge JA, Schwab S, dan Ting IP. 1982. Interaction of photoperiod and vesicular-arbuscular mycorrhizae on growth and metabolism of sweet orange. New Phytol. 90 : 665-673
Johnson-Green PC, Kenkel NC dan Booth T. 1995. The distribution and phenology of arbuscular mycorrhizae along an inland salinity gradient. Can. J. Bot. 73 : 1318-1327
Kormanik PP dan McGraw AC. 1982. Quantification of VA mycorrhizae in plant root. Di Dalam : N.C.Schenk (Ed.) Methods and principles of mycorrhizae research. The American Phytop. Soc. 46 : 37-45
Lewis D.H. 1986. Inter-relationships between carbon nutrition and morphogenesis in mycorrhizas. Di Dalam : Gianinazzi-Pearson V dan Gianinazzi S (Eds). Physiological and genetical aspect of mycorrhizae. Proceeding of the 1st Europens Symposium on Mycorrhizae. Hal. 85-100
Louis I. 1990. A mycorrhizal survey of plant species colonizing coastal reclaimed land in Singapore. Mycologia 82 (6) : 772-778
Maryadi F. 2002. Status dan keragaman CMA di bawah tegakan klonal jati (Tectona grandis L.f) pada umur 4-7 tahun di kebun benih klonal padangan. Skripsi. Fakulta Kehutanan Institut Pertanian Bogor. 53 hal.
McGraw AC dan Hendrix JW. 1984. Host and fumigation effects on spore population densities of species of endogonaceous mycorrhizal fungi. Mycologia. 76 : 122-131
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
Ocampo JA, Cardona FL dan El-Atrach F. 1986. Effect of root extracts of non host plants on VA mycorrhizal infection and spore germination. Di Dalam : Gianinazzi-Pearson V dan Gianinazzi S (Eds). Physiological and genetical aspect of mycorrhizae. Proceeding of the 1st Europens Symposium on Mycorrhizae. Hal. 721-724
Pacioni G. 1986. Sporulation of the VAM fungi stimulated by water stress in natural conditions. Di Dalam : Gianinazzi-Pearson V dan Gianinazzi S (Eds). Physiological and genetical aspect of mycorrhizae. Proceeding of the 1st Europens Symposium on Mycorrhizae. Hal. 713-716
Puppi G, Tabacchini P, Riess S, dan Sanvito A. 1986. Seasonal pattern in mycorrhizal associations in maritime sand dune system (Castelporziano, Italy). Di Dalam : Gianinazzi-Pearson V dan Gianinazzi S (Eds). Physiological and genetical aspect of mycorrhizae. Proceeding of the 1st Europens Symposium on Mycorrhizae. Hal. 245-249
Purwanto A. 1999. Studi hubungan salinitas dengan kelimpahan cendawan mikoriza arbuskula (CMA) pada lahan hutan pantai dan hutan mangrove di cagar alam Leuweung Sancang, Kabupaten Garut, Jawa Barat. Skripsi Jurusan Manajemen Hutan. Fakultas Kehutanan IPB. Bogor. 33 hal. Rozema J, W. ARP, Van Esbroek M, Broekman R, Punte H, dan Schat H. 1986.
Vesicular arbuscular mycorrhiza in salt marsh plants in response to soil salinity and flooding and the significance to water relations. Di Dalam : Physiological and genetical aspect of mycorrhizae. Proceeding of the 1st Europens Symposium on Mycorrhizae. Hal. 657-660.
Saif SR dan Khan G. 1975. The influence of season and stage of development of plant on Endogone mycorrhiza of field-grown wheat. Can J. Microbiol 21 : 1020-1024
Semones SW dan Young DR. 1995. VAM association in the shrub Myrica cerifera on a Virginia, USA barrier island. Mycorrhiza 5 : 423-429
Siguenza C, Espejel I dan Allen EB. 1996. Seasonality of mycorrhizae in coastal sand dunes of Baja California. Mycorrhiza 6 ; 151-157
Sieverding E and Toro TS. 1988. Influence of Soil Water Regimes on VA Mycorrhiza. V. Performance of Different Fungal Species with Cassava. J. Agron. Crop Sci. 161 : 322-332.
Silviana, Gunawan AW dan Kramadibrata K. 1999. Biodiversity of arbucular mycorrhizal fungi in the rhizospheres of mangosteen. Di Dalam : Smith FA, Kramadibrata K, Simanungkalit RDM, Sukarno N, dan Nuhamara ST (Eds.) Mycorrhizas in sustainable tropical agriculture and forest ecosystems. Proceedings of International Conference on Mycorrhiza. Bogor, Indonesia. Hal. 97-100
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006
Smith SE and Read DJ. 1997. Mycorrhizal symbiosis. Second edition. Academic Press. Harcourt Brace & Company Publisher. London. pp. 32-79
Sparling GP dan Tinker PB. 1978. Mycorrhizal infection in Pennine grassland I Level infection in the field. J. Appl. Ecol. 15 : 943-950
Sutton JC dan Barron GL. 1972. Population dynamics of Endogone spores in soil. Can. J. Bot. 54 : 326-333
Sylvia DM and Schenck NC. 1982. Effect of Post-colonization Treatments on Sporulation of Vesicular-Arbuscular Mycorrhizal Fungi. Phytopathology. 72 : 950-957.
van der Heijden MGA, Klironomos JN, Ursic M, Moutoglis P, Streitwolf-Engel R, Boller T, weimken A, dan Sanders IR. 1998. Mycorrhizal fungal diversity determines plant biodiversity, ecosystems variability and productivity. Nature 5 : 69-72
Wetzel PR dan van der Valk A. 1996. Vesicular-arbuscular mycorrhizae un prairie pothole wetland vegetationin Iowa and North Dakota. Can. J. Bot. 74 : 883-890
Young A. 1976. Tropical soil and soil survey. Cambridge University Press. Cambridge.
Delvian: Dinamika Sporulasi Cendawan Mikoriza Arbuskula, 2006