Molecular cloning of a novel water channel from rice: its products
expression in
Xenopus
oocytes and involvement in chilling
tolerance
Le-gong Li
1, Shi-fang Li
2, Yan Tao, Yoshichika Kitagawa *
Laboratory of Plant Genetic Engineering,Biotechnology Institute,Akita Prefectural Uni6ersity,Ogata,Akita010-0444,Japan
Received 15 March 1999; received in revised form 28 October 1999; accepted 1 December 1999
Abstract
Water channel proteins, aquaporins, play a fundamental role in transmembrane water movements in plants. We isolated rice cDNA,rwc1, by screening a rice (Oryza sati6acv. Josaeng Tongil) cDNA library using a conserved motif of aquaporins. Like
other aquaporin genes,rwc1 encodes a 290-residue protein with six putative transmembrane domains. The derived amino acid sequence of RWC1 shows high homology with PIP1 (plasma membrane intrinsic protein 1) subfamily members, which suggest it is localized in the plasma membrane. Injection of its cRNA intoXenopusoocytes increased the osmotic water permeability of the oocytes 2 – 3 times. Northern analysis showed that rice aquaporin genes are expressed in rice seedling leaves and roots, but that it disappeared from the root 6 h after osmotic stress began and that the transcript level remained low for about 24 h, then recovered. The time course of rice aquaporin gene-expression under osmotic stress was correlated with time course of turgor transition in plant. On the other hand, the levels of rice aquaporin gene-transcripts in leaves under chilling and recovery temperature depend on the pretreatment of mannitol for short time. This variation of the transcripts shown that rice aquaporin genes may play an important role in response to water stress-induced chilling tolerance. © 2000 Elsevier Science Ireland Ltd. All rights reserved.
Keywords:Rice; Water channel; Water stress; Chilling tolerance;Xenopusoocyte
www.elsevier.com/locate/plantsci
1. Introduction
Transmembrane water flow is a fundamental process of life. Although water permeability as a biophysical feature of cell membrane, the molecu-lar pathway of transmembrane water movement remained unknown until the discovery of
aquapor-ins [1]. The recent discovery that plants express numerous aquaporins in both the plasma mem-brane and the tonoplast has changed our view of how plant cells regulate transmembrane water movement [2,3]. Water channel proteins (aquapor-ins) belong to the major intrinsic protein (MIP) superfamily that permit the passage of specific molecules through biological membranes. Since the first aquaporin (AQP1) was identified in hu-man erythrocytes [1], hu-many more have been iso-lated from various organisms, including bacteria, plants, and animals [3]. In plants, many MIP genes have been isolated. They are encoded by several gene families and are hydrophobic integral mem-brane proteins that range in apparent molecular
mass from 23 to 31 kDa [4]. Since g-tonoplast
intrinsic protein (g-TIP) was first recognized as a
plant aquaporin [5], more different genes have
The nucleotide sequence data reported appeared in EMBL, Gen-Bank and DDBJ Nucleotide sequence Database under the accession number AB009665.
* Corresponding author.
E-mail address:[email protected] (Y. Kitagawa)
1Permanent address: Shanghai Institute of Plant Physiology,
Chi-nese Academy of Sciences, Shanghai 200032, People’s Republic of China.
2Present address: Institute of Plant Protection, Chinese Academy
of Agricultural Sciences, 2 West Yuanmingyuan Road, Beijing 100094, China. Le-gong Li and Shi-fang Li contributed equally to this work.
been identified [6]. Their products are located in the tonoplast and plasma membrane. Sequence comparisons have shown a high homology be-tween plant aquaporins; all published sequences are clearly of TIP or plasma membrane intrinsic protein (PIP) members. These relationships even extend to the PIP1 and PIP2 subfamilies, origi-nally introduced by Kammerloher et al. [7]; easily classified as PIP1 or PIP2 based on specific arrays of amino acids at the N- and C-termini.
Although DNA sequences with high homology to the aquaporin genes have been identified in several plant species [4], the water permeability and function haven’t been determined, only a few of the gene products have been characterized [6 – 11]. Their roles in response to various physiologi-cal and stress conditions warrants further research. Here we report the isolation of the first water channel gene to be identified in rice, the activity of its water permeability, and the possible role in response to osmotic and chilling stress.
2. Materials and methods
2.1. Plant material, osmotic and chilling treatments
Wasetoitsu (Oryza sati6a cv. Josaeng Tongil) is
a less chilling-tolerant hybrid rice variety devel-oped in the Republic of Korea. Seedlings and culture cells were prepared as described previously [12]. For treatment with mannitol and NaCl, rice seedlings at the three leaf-stage were transferred to
fresh liquid Hoagland’s medium [5 mM Ca(NO3)2,
5 mM KNO3, 2 mM MgSO4, 0.025 mM FeSO4
-EDTA in tap water] containing 0.25 M mannitol or 0.15 NaCl, at 25°C. For chilling, after mannitol treatment the seedling was washed with tap water, and transferred to fresh Hoagland’s medium, incu-bated for 24 h at 4°C and then transferred to 25°C for 24 h.
2.2. cDNA library construction and screening
Total RNA was extracted from leaves that had been treated for 30 min with 0.5 M mannitol. A
cDNA library was constructed in the NotI site of
lgt10. Aliquots of the cDNA were used as a
template for the polymerase chain reaction (PCR) with designed primers corresponding to the
se-quences surrounding the Asn-Pro-Ala motif of aquaporins [5]. A PCR product of 393 bp was subcloned into the pT7Blunt vector (Novergen, USA) and sequenced on both strands using ABI373 Applied Biosystem (USA). This DNA
fragment was radioactive labeled with [a32P]dCTP
by random DNA labeling (Takara, Japan) and
was used as a hybridization probe to screen 3×
105 plaques from the cDNA library. A positive
plaque was isolated and the insert of the phage
DNA was subcloned into a pBluescript KS+
ac-cording to the manufacturer’s protocol
(Strata-gene, USA). The subcloned cDNA, rwc1, was
sequenced and analyzed using the GENETYX
Ver-sion 8 (Software Development Co., Japan).
2.3. Northern analysis
Total RNA was prepared as described by
Koga-Ban et al. [13], and 5 mg of total RNA was
fractionated on a 1.2% denaturing formaldehyde agarose gel. The RNA was transferred to a
Hy-bond™-N+ membrane (Amersham, UK) using
200SSC [14]. Prehybridization and hybridization were performed using a rapid-hybridization buffer, as suggested by the manufacturer (Amersham).
The probe was the full-length rwc1 sequence
la-beled with 32P by the random primer method
(Takara, Japan).
2.4. Competiti6e PCR
Transcription levels of rwc1 were specifically
determined by the competitive PCR method [15]. Competitor lambda RNA (448 bases) against rice actin mRNA (accession number X15863 [16]) was made from a lambda DNA fragment placed in
between the rice actin 5%primer (5%
-AGAGCTAC-GAGCTTCCTGATGGAC-3%) and 3% primer
(5%- GAGAGATGCCAAGATGGATCCTCC - 3%)
regions. A SP6 promoter (5%
-ATTTAGGTGA-CACTATAGAATAC-3%), induced upstream of
the 5%actin primer, was used to synthesize RNA
using SP6 polymerase according to the
manufac-ture’s protocol (Takara, Japan). Competitor
lambda RNA (530 bases) againstrwc1 mRNA was
prepared in the same manner from a lambda DNA
fragment that was spaced by rwc1 5%(5%
-ATCTA-CAACAAGGACCATGCCTGGA-3%) andrwc1 3%
(5%- ATTACACGATTGAGTTGTTCAGGGT - 3%)
pro-moter. Using the rwc1 or actin-primers, fragment
sizes of 333 bp for actin and 280 bp for rwc1 are
obtained by RT-PCR using total RNA. The reac-tion mixture contained 10 ng of sample RNA,
105– 109 copies of competitor lambda RNA, 10
pmol of random 9 mer primer, 1 U of AMV reverse transcriptase, 4 U of RNase inhibitor, and
4 mM dNTP mixture in 4 ml of PCR buffer (10
mM Tris – HCl, pH 8.3, 50 mM KCl and 5 mM
MgCl2). cDNA synthesize was allowed to proceed
for 30 min at 42°C, followed by denaturation for 5
min at 99°C. The 4 ml cDNA solution was then
mixed with 16ml of PCR reaction mixture
contain-ing 1 – 4 pmol each of the particular 5%and 3%
primers, and 0.5 U of Taq polymerase in PCR
buffer. Samples were amplified by 30 cycles at 94°C for 0.5 min, 55°C for 0.5 min, and 72°C for 1 min. An aliquot of each reaction mixture was subjected to electrophoresis on 3% NuSive agarose gel.
2.5. In 6itro synthesis of rwc1 cRNA
The rwc1 gene which was cut from the plasmid
withBamHI was inserted into theBglII site of the
Xenopus expression construct pXbG/ev-1; a
pSP64T-derived pBluescript-type vector into
which Xenopus b-globin 5%and 3%untranslated
sequences were inserted [1]. Recombination of the plasmid was checked by sequencing. Capped RNA
transcripts ofrwc1 were synthesized in vitro using
T3 RNA polymerase and linearized recombinant
plasmids containing rwc1 cDNA after digestion
withBamHI (Stratagene). After ethanol
precipita-tion, the synthesis products were suspended in
diethylpyrocarbonate-treated H2O at a final
con-centration of 1 mg/ml.
2.6. Microinjection of cRNA into oocytes and water permeability analysis
Mature oocytes (1.2 – 1.3 mm diameter, stage V
and VI) were isolated from adult Xenopus la6is
[17,18] and stored in Barth’s buffer containing
Na-penicillin (10 mg/ml) and streptomycin (10 mg/
ml). The follicular cell layer was removed by
incu-bation with 2 mg/ml collagenase (Boehringer,
Germany) in Barth’s buffer for 2.0 – 2.5 h at 25°C with gentle continuous agitation. Defolliculated oocytes were injected with 46 nl of cRNA (1
mg/ml) or water using an automatic injector
(Nanoject; Drummond). The oocytes were incu-bated for 2 days at 18°C in Barth’s buffer before water permeability measurements.
Individual oocytes were transferred from
Barth’s buffer (200 mosm) to a three-fold dilution of Barth’s buffer with distilled water (70 mosm) at 20°C. Oocyte swelling was checked using an area Colony Analyzer (CA-7, Toyo Sottuki, Japan). The area covered by the oocytes was measured at 5-s intervals; the total cell volume was calculated from the cell area. The relative oocyte volume
(V/V0) was calculated from the relative oocyte
area (A/A0) using the equation V/V0=(A/A0)
3/2.
The osmotic water permeability coefficient (Pf,
cm/s) was calculated from oocyte surface area
(S=0.045 cm2), initial oocyte volume (V
0=9×
10−4 cm), water volume (Vw=18 cm3/mol), and
the initial rate of oocyte swelling, d(V/V0)/dt,
using the equation Pf=V0d(V/V0)/dt[SVw
-(osmout−osmin)], where osmout is 70 mosm and
osmin is 200 mosm.
3. Results and discussion
3.1. Isolation of rwc1 from the rice cDNA library
We used the most conserved sequences of the aquaporins to design our PCR primers, and used the cDNA library as the template. The 393-bp product showed high sequence homology to PIP1c from Arabidopsis, and it was used to probe the
same cDNA library. The 1.1-kb fragment of rwc1
contained a 49-bp 5%-untranslated sequence
pre-ceding an initiation site consensus sequence. An 870-bp open reading frame was followed by a
3%-untranslated sequence containing a
polyadeny-lation consensus sequence (accession no.
AB009665).
3.2. Deduced structure of RWC1 and homology comparisons
Analysis of the Genbank database showed that
rwc1 is a novel member of the aquaporin family.
domains as well as the functionally important B and E loops. Hydropathy analysis indicated that RWC1 has six putative bilayer-spanning domains and five connecting loops, of which loops B and E were also hydrophobic (Fig. 1b). Both hydropho-bic loops, which may form opposite leaflets, contained the NPA motif (Asn-Pro-Ala) that is present in all aquaproins (Fig. 1b),
how-ever RWC1 did not contain the cysteine
residue that confers mercury sensitivity to AQP1 [19]. In addition, the phosphorylation site seen in PM28A (Ser-274) [20] was not identified in RWC1; however, the predicted topology of RWC1 contained some distinctive features. One putative cAMP or cGMP-dependent protein kinase phos-phorylation motif (RKLS) was identified at residues 128 – 131. Two consensus sequences (SER, SSK) for protein kinase C phosphorylation were present near the N-terminal region (Fig. 1a), which was longer than the corresponding domain
in E. coli [21] and mammalian aquaporins; some
domains may have functions that are specific to plants.
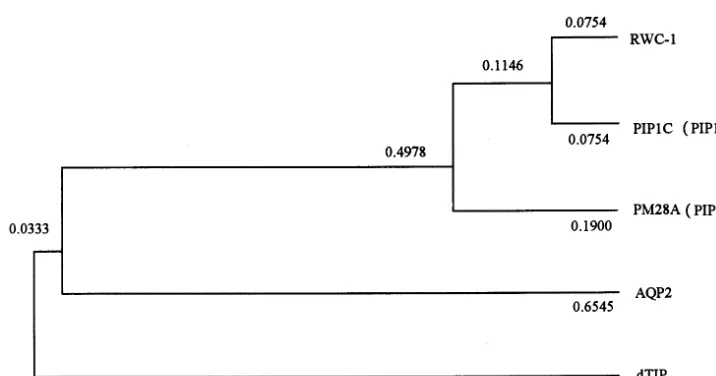
Nucleotide sequence comparisons between rwc1
and other genes analysis are shown in Fig. 1C.
Related sequences were detected in human, Ara
-bidopsisand spinach genes. The phylogenetic
anal-ysis indicates thatrwc1 belongs to the MIP-related
genes and is more closely related to PIP1 of Ara
-bidopsis than to PM28A of spinach. We have
classified rwc1 as a member of the PIP1 subfamily.
The deduced amino acid sequence of RWC1
dif-fers significantly from that of g-TIP, which is
located in the tonoplast, so RWC1 may be located in plasma membrane.
3.3. Rice aquaporin genes response to osmotic
6ariation and in6ol6e into water stress-induced chilling tolerance
The expression of rice aquaporin genes was determined by Northern blotting using total RNA isolated from leaves and roots of rice seedlings at
the three-leaf stage and full-length rwc1 as the
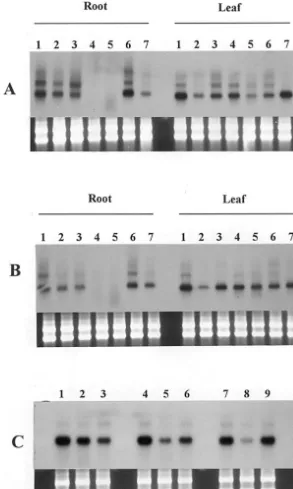
probe. Rice aquaporin-mRNAs were detected in both leaves and roots (Fig. 2a and b). When the seedlings were treated with 0.25 M mannitol or 0.15 M NaCl, the transcript level changed. Tran-script level in root had declined drastically 6 h after the stress began and remained low for about 24 h. After that period, the transcript level of aquaporin genes increased again to at least the pre-stress level (Fig. 2A and B). The response of rice aquaporin genes to osmotic stress is similar to response of MipA and MipC genes in ice plants [11]; expression of aquaporin genes is suppressed under osmotic stress. Beside aquaporin gene-sig-nal, there are several signals in Fig. 2A and B, but
Fig. 2. Northern blot analysis of rice aquaporin-mRNA, stress response and tissue specificity. Total RNA fractions were prepared from leaves and roots, and then aliquots (5mg) of them were applied to northern analysis. The gels were stained with ethidium bromide to detect rRNAs (indicated below). (A) Mannitol treatment. 1, control; 2, 0.25 M mannitol for 1 h; 3, 0.25 M mannitol for 3 h; 4, 0.25 M mannitol for 6 h; 5, 0.25 M mannitol for 24 h; 6, 0.25 M mannitol for 48 h; 7, 0.25 M mannitol for 72 h. (B) NaCl treatment. 1, control; 2, 0.15 M NaCl for 1 h; 3, 0.15 M NaCl for 3 h; 4, 0.15 M NaCl for 6 h; 5, 0.15 M NaCl for 24 h; 6, 0.15 M NaCl for 48 h; 7, 0.15 M NaCl for 72 h. (C) Chilling treatment (leaf). 1, control; 2, 4°C for 24 h; 3, 4°C for 24 h, then transferred to 25°C for 24 h; 4, 0.25 M mannitol for 3 h; 5, 0.25 M mannitol for 3 h, then incubated at 4°C for 24 h; 6, 0.25 M mannitol for 3 h, then incubated at 4°C for 24 h and transferred to 25°C for 24 h; 7, 0.25 M mannitol for 6 h; 8, 0.25 M mannitol for 6 h, then incubated at 4°C for 24 h; 9, 0.25 M mannitol for 6 h, then incubated at 4°C for 24 h and transferred to 25°C for 24 h.
not in Fig. 2C. It is presumed that washing condi-tion of RNA blot of Fig. 2A and B was not optimal.
to 0.25 – 0.5 M mannitol over a short time before chilling treatment [12]. When Wasetoitsu seedlings pretreated with 0.25 M mannitol for 3 and 6 h were incubated at 4°C for 24 h and then trans-ferred to 25°C for 1 day, the expression of the aquaporin genes in the leaves showed fluctuation. Mannitol pretreatment for short time is helpful to down-regulate the expression of the aquaporin genes under chilling stress, and improve its expres-sion during the period in recovering (Fig. 2C). When the pretreatment time with mannitol are increased, the transcription levels of the aquaporin genes in leaves are to be lower at low temperature,
but the transcription levels are recovered signifi-cantly after the seedling transferred to 25°C (Fig. 2C). As we know, water channel is a bi-directional pore for water efflux and influx. The driving forces behind water movement are hydraulic or osmotic in nature [2]. One aspect of the chilling harm to plant is lost water. At low temperature, lower expression of aquaporin genes is beneficial to keep a suitable status of water; a fast recovery is also necessary for adaptation to standard physiological process. Characterization of the expression of aquaporin genes is just right to match the plant request. It is presumed that aquaporin genes may play an important role in response to water-stress induced chilling tolerance.
The multiplicity of MIP genes are in plants as reported before [22], it leads us to presume that many similar genes exist in rice; some may be precisely regulated by water stress, whereas other are probably not affected. In this Northern experi-ment we could not determine the specific
expres-sion of rwc1, because we used full-length rwc1 as
the probe. Next we determined the specific
expres-sion ofrwc1 in rice roots under osmotic stress and
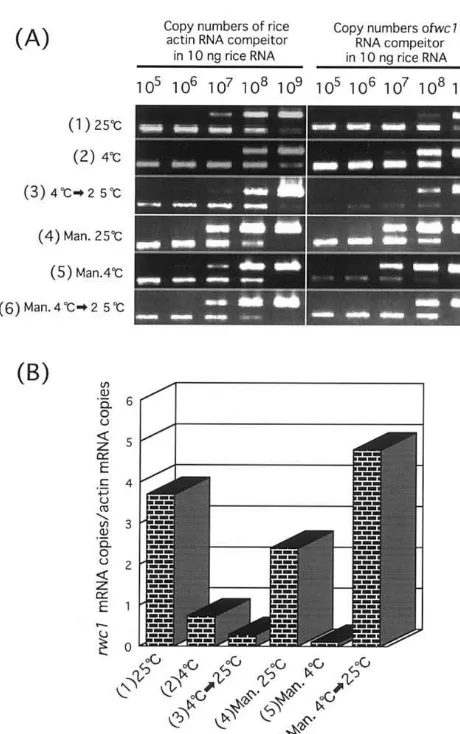
chilling stress by competitive PCR using specific
primers for the 3%-untranslated rwc1 region. The
agarose gel results are shown in Fig. 3A, and the
calculated numbers of rwc1 transcripts per actin
transcript are shown in Fig. 3B. The specific
ex-pression of rwc1 mRNA under osmotic and
chill-ing stress is essentially similar to the results of Northern hybridization shown in Fig. 2C, and
suggests that rwc1 in particular is involved in
responses to water stress-induced chilling
tolerance.
3.4. Analysis of the water transport function of RWC1
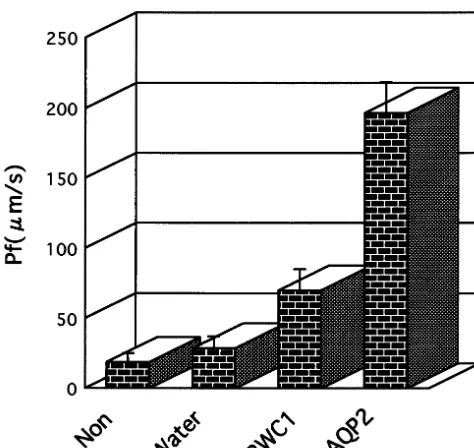
The water channel activity of RWC1 was
as-sayed by injecting rwc1 complementary RNA
(cRNA) into Xenopus oocytes. A cRNA that was
synthesized in vitro from the cDNA, which had
been cloned in vector pXbG/ev-1. cRNA of aqp2
[23] and rwc1 synthesized using pXbG/ev-l, were
injected into oocytes. After 3 days of incubation in isotonic Barth’s buffer, the oocytes were
trans-ferred to a hypotonic buffer (1/3 Barth’s buffer),
and the relative increase in cell volume, a conse-quence of water uptake, was measured [17]. The results indicate that RWC1 has water channel
Fig. 4. Expression ofrwc1 cRNA and cell membrane perme-ability. Oocytes were injected with cRNA orF water as indi-cated. Oocyte swelling was as described in Section 2. Values are the mean9S.D. of 7 – 10 oocytes (stippled bars).
Acknowledgements
We thank Dr K. Fushimi and M. Kuwahara of Tokyo Medical and Dental University for kindly
providing the AQP2 gene and pXbG/ev-1 vector.
This work was supported by Grant-in-Aid for Scientific Research from the Ministry of Educa-tion, Science and Culture (Japan), for Special Sci-entific Research on Agriculture, Forestry and Fisheries, and by Grants from the Sumitomo Foundation (Japan) and Ciba-Geigy Foundation (Japan) for the Promotion of Science; and Na-tional nature and science foundation: 39670074 (China).
References
[1] G.M. Preston, T.P. Carroll, W.B. Guggino, P. Agre, Appearance of water channels in Xenopus oocytes ex-pressing red cell CHIP28 protein, Science 256 (1992) 385 – 387.
[2] M.J. Chrispeels, C. Maurel, Aquaporins: the molecular basis of facilitated water movement through living plant cells?, Plant Physiol. 105 (1994) 9 – 13.
[3] C. Maurel, Aquaporins and water permeability of plant membranes, Annu. Rev. Plant Physiol. Plant Mol. Biol. 48 (1997) 399 – 429.
[4] A.R. Scha¨ffner, Aquaporin function, structure, and ex-pression: are there more surprises to surface in water relations, Planta 204 (1998) 131 – 139.
[5] C. Maurel, J. Reizer, J.I. Schroeder, M.J. Chrispeels, The vacuolar membrane protein r-TIP creates water specific channels inXenopusoocytes, EMBO J. 12 (1993) 2241 – 2247.
[6] A. Weig, C. Deswarte, M.J. Chrispeels, The major intrin-sic protein family of Arabidopsis has 23 members that form three distinct groups with functional aquaporins in each group, Plant Physiol. 114 (1997) 1347 – 1357. [7] W. Kammerloher, U. Fisher, G.P. Piechottka, A.R.
Scha¨ffner, Water channels in the plant plasma membrane cloned by immunoselection from a mammalian expres-sion system, Plant J. 6 (1994) 187 – 199.
[8] M.J. Daniels, E. Mirkov, M.J. Chrispeels, The plasma membrane ofArabidopsis thalianacontains a mercury-in-sensitive aquaporin that is a homolog of the tonoplast water channel protein TIP, Plant Physiol. 106 (1994) 1325 – 1333.
[9] M.J. Daniels, F. Chaumont, T.E. Mirkov, M.J. Chrispeels, Characterization of a new vacuolar mem-brane aquaporin sensitive to mercury at a unique site, Plant Cell 8 (1996) 587 – 599.
[10] C.H. Oppermann, C.G. Taylor, M.A. Conkling, Root-knot nematode-directed expression of a plant root-spe-cific gene, Science 263 (1994) 221 – 223.
[11] S. Yamada, M. Katsuhara, W.B. Kelly, C.B. Michalowski, H.J. Bohernert, A family of transcripts encoding water channel proteins: tissue-specific
expres-activity in oocytes (Fig. 4). Our osmotic water permeability indicated that the RWC1 protein led to a swelling rate 2 – 3 times higher than that of water-injected oocytes. However, the water
permeability of the oocytes injected with rwc1
cRNA was not as great as that of oocytes
in-jected with aqp2 cRNA. Why did RWC1 show
weaker water channel activity? There are several possible reasons. One is that the phosphorylation sites and the other residues of the carboxyl-ter-minal cytoplasmic domain of RWC1 do not clar-ify function as restraints, as they do in animals. The second possibility is that, as for MIP in animals, a specific membrane environment may be required to improve the translation of the
rwc1 mRNA [24].
sion in the common ice plant, Plant Cell 7 (1995) 1129 – 1142.
[12] Y. Kitagawa, K. Yoshizaki, Water stress-induced chilling tolerance in rice; putative relationship between chilling tolerance and Ca2+ flux, Plant Sci. 137 (1998) 73 – 85.
[13] Y. Koga-Ban, M. Abe, Y. Kitagawa, Alteration in gene expression during cold treatment of rice plants, Plant Cell Physiol. 32 (1991) 901 – 905.
[14] J. Sambrook, E.F. Fritsch, T. Maniatis, Molecular Cloning: a Laboratory Manual, second ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989.
[15] G. Gilliland, S. Perrin, K. Blanchard, F. Bunn, Analysis of cytokine mRNA and DNA: Detection and quantita-tion by competitive polymerase chain reacquantita-tion, Proc. Natl. Acad. Sci. USA 87 (1990) 2725 – 2729.
[16] K.S. Reece, D. McElroy, R. Wu, Genomic nucleotide sequence of four rice (Oryza sati6a) actin genes, Plant Mol. Biol. 14 (1990) 621 – 624.
[17] R. Zhang, K.A. Logee, A.S. Verkman, Expression of mRNA coding for kidney and red cell water channels in
Xenopus oocytes, J. Biol. Chem. 265 (1990) 15375 – 15378.
[18] R. Zhang, A.S. Verkman, Water and urea permeability properties of Xenopus oocytes: expression of mRNA from toad urinary bladder, Am. J. Physiol. 260 (1991) C26 – C34.
[19] G.M. Preston, J.S. Jung, W.B. Guggino, P. Agre, The
mercury-sensitive residue at cysteine 189 in the CHIP28 water channel, J. Biol. Chem. 268 (1993) 17 – 20. [20] I. Johansson, M. Karisson, V.K. Shukla, M.J.
Chrispeels, C. Larsson, P. Kjellbom, Water transport activity of the plasma membrane aquaporin PM28A is regulated by phosphorylation, Plant Cell 10 (1998) 451 – 459.
[21] G. Calamita, K. Bettina, M. Bonhivers, R. Bishai, E. Bermer, P. Agre, Regulation of theEscherichia coliwater channel geneaqp2, Proc. Natl. Acad. Sci. USA 95 (1998) 3613 – 3627.
[22] S. Yamada, T. Komori, P.N. Myers, S. Kuwata, T. Kubo, H. Imaseki, Expression of plasma membrane wa-ter channel genes under wawa-ter stress inNicotiana excel
-sior, Plant Cell Physiol. 38 (1997) 1226 – 1231.
[23] K. Fushimi, S. Uchida, Y. Hara, Y. Hirata, F. Marumo, S. Sasaki, Cloning and expression of apical membrane water channel of rat kidney collecting tubule, Nature 361 (1993) 549 – 552.
[24] S.M. Mulders, G.M. Preston, P.M.T. Deen, W.B. Gugg-ino, C.H. van Os, P. Agre, Water channel properties of major intrinsic protein of lens, J. Biol. Chem. 270 (1995) 9010 – 9016.
[25] J.B. Mariaux, C. Bockel, F. Salamini, D. Bartels, Desic-cation- and abscisic acid-responsive genes encoding ma-jor intrinsic proteins (MiPs) from the resurrection plant
Craterostigma plantagineum, Plant Mol. Biol. 38 (1998) 1089 – 1099.