Comparison of models for daily light partitioning in
multispecies canopies
H. Sinoquet
a,∗, M. Rakocevic
b, C. Varlet-Grancher
c aINRA-PIAF, Site de Crouelle, 234 avenue du Brézet, F-63039 Clermont-Ferrand Cedex 2, FrancebFaculty of Agriculture, University of Belgrade, 11080 Zemun, Yugoslavia cINRA-SEPF, Domaine des Verrines, F-86600 Lusignan, France
Received 16 July 1999; received in revised form 3 December 1999; accepted 9 December 1999
Abstract
A simulation model of light partitioning in horizontally homogeneous multispecies canopies is proposed. The model is based on the Kubelka–Munk equations (KM) applied to a mixture of N vegetation components. Only two hemispherical fluxes, i.e. downwards and upwards, are considered. The exact solution of KM equations was then simplified in such a way that the model can be easily extended to multispecies canopies including several vegetation layers. The simplified KM model (KMS) was compared to two other light models dealing with mixed canopies: the more detailed model SIRASCA [Sinoquet, H., Moulia, B., Gastal, F., Bonhomme, R., Varlet-Grancher, C., 1990. Modeling the radiative balance of the components of a well-mixed canopy: application to a white clover–tall fescue mixture. Acta Oecol. 11, 469–486], and the simpler model ERIN [Wallace, J.S., 1997. Evaporation and radiation interception by neighbouring plants. Q. J. R. Meteorol. Soc. 123, 1885–1905]. All three models were applied to theoretical two-species monolayer canopies, and to actual mixed canopies, the geometry of which was retrieved from the literature. In the PAR waveband, the model KMS gave simulation results very similar to those of SIRASCA in case of contrasted canopy structures. In conditions of high leaf and soil scattering, deviations between SIRASCA and KMS outputs were higher and reached maximum values of –0.08 for erectophile species. Comparison between SIRASCA and ERIN outputs showed that ERIN largely underestimated light competition in a two-component canopy in several conditions, due to light partitioning only based on height differences between components. Simulations also showed the significant effect of the vertical distribution of leaf area on light partitioning in the case of mixtures where components had equal or different heights. Finally it appears that the model KMS could be a candidate for inclusion in growth models for multispecies canopies, since all KMS parameters have physical meaning and it is very easy to implement. ©2000 Elsevier Science B.V. All rights reserved.
Keywords: Radiation balance; PAR; Inter-cropping; Grass; Legume; Canopy structure
1. Introduction
Light partitioning is a crucial issue in multispecies canopies because light is involved in most plant re-sponses (e.g. photosynthesis, transpiration,
morpho-∗Corresponding author. Tel.:+33-4-73-62-4361; fax:+33-4-73-62-4454.
E-mail address: [email protected] (H. Sinoquet).
genesis) and the effects of light reduction on the dom-inated species may be either negative (e.g. species extinction, Caldwell, 1987) or positive (e.g. shelter from water stress, Allen et al., 1976; improvement of light use efficiency, Willey, 1979; Harris et al., 1987; Cruz, 1995). All process-based simulation models de-voted to multispecies canopies (tree/crop, McMurtrie and Wolf, 1983; e.g. crop/weeds, Kiniry et al., 1992; crop/crop, Caldwell and Hansen, 1993; grass/legume,
Soussana et al., 2000) include a sub-model of light partitioning between species. Similarly growth analy-sis methods for multispecies canopies are all based on the ones by Monteith (1972) where primary produc-tion is regarded as the combined result of light capture and use. This requires the assessment of light sharing between species. This is expressed in terms of light interception efficiency (εi) of individual component i,
namely the fraction of incident radiation which is ab-sorbed by component i (e.g. Azam-Ali et al., 1990; Cruz and Sinoquet, 1994).
All simulation models devoted to light partition-ing between species are based on the turbid medium analogy, i.e. the classical Beer’s law computing light transmission I as a negative exponential function of the downward cumulated leaf area index (LAI, L)
I =I0exp(−KL) (1)
where I0 is incident radiation and K is an extinction
coefficient. Because it deals with light transmission (i.e. non-intercepted radiation), Beer’s law cannot be used by itself to estimate light sharing between species, except if foliages occupy separated canopy spaces (e.g. tree/grass, McMurtrie and Wolf, 1983). In other cases, assessment of light partitioning needs fur-ther development. From the basic assumptions used to derive Beer’s law in vegetation canopies (i.e. small leaf size, random leaf dispersion), several authors (e.g. Rimmington, 1984; Sinoquet and Bonhomme, 1991) showed that light interception efficiency εi of
species i in a mixture of N components can be written
εi =
Eq. (2) shows that total light interception (i.e. the term in square brackets in the right member) is partitioned between species according to their contribution to total LAI (Li) weighted by their interception ability
(i.e. extinction coefficient Ki). While several models
are based on Eq. (2) (Rimmington, 1984; Ryel et al., 1990; Sinoquet et al., 1990; Wiles and Wilkerson, 1991), other models share light between species ac-cording only to the contribution to total LAI (Spitters and Aerts, 1983; Graf et al., 1990), i.e. differences in extinction coefficient are not taken into account. Eq. (2) or similar relationships have been included in light
models by different ways. The canopy may be treated as a mono-layer (Rimmington, 1984) or a multilayer medium (Spitters and Aerts, 1983; Rimmington, 1985; Graf et al., 1990). Most models applied Eq. (2) to a single hemispherical downward flux, but Ryel et al. (1990) and Sinoquet et al. (1990) distinguished a set of directions in order to apply Eq. (2) to directional fluxes. Ryel et al. (1990) and Sinoquet et al. (1990) also included multiple scattering in their models.
In contrast to the previous approach, Wallace et al. (1991) proposed a rather different way to esti-mate light sharing in two-species canopies: in the first step, light interception is computed from Beer’s law applied to (i) a two-layer canopy where Species #1 overtops Species #2 (ε1max and ε2min), and (ii)
another one where Species #2 overtops Species #1 (ε1minandε2max). These two virtual situations
corre-spond to cases of maximum dominance of one species over the other, hence in the actual situation,εi range
betweenεiminandεimax. In the following step,εi is
thus written as a linear combination ofεiminandεimax
εi =εmini (1−fi)+εimaxfi (3)
where f is an empirical function of species heights h1
and h2, e.g. (Wallace, 1997)
fi =
hi
(h1+h2)
(4)
multiple species canopies. Multilayer models, how-ever, need many more input parameters (i.e. LAI of each component in each layer) than of Wallace (1997) (i.e. total LAI and height of each compo-nent).
The first objective of this paper was to propose a simple light partitioning model for horizontally homo-geneous canopies, where parameters could be explic-itly related to canopy structure and optical properties of the leaves and the soil surface. Such a model would be useful as a sub-model in growth simulation mod-els. For this purpose, we derived the KM equations (Kubelka and Munk, 1931) for the case of a mixture of
N vegetation components, and we simplified the
solu-tion in order to obtain simple equasolu-tions for light par-titioning. As a test, we compared the simplified KM equations (KMS) to the detailed model of Sinoquet et al. (1990) on contrasting canopy structures. The second objective was to assess the requirements in structure parameters for an accurate estimation of light partitioning. For this, the detailed and simplified models were run on mono and multilayer canopies, and compared with the model of Wallace (1997).
2. Kubelka–Munk equations for multispecies canopies
2.1. Kubelka–Munk equations in a multispecies monolayer canopy
Equations were derived according to Bonhomme and Varlet-Grancher (1977). The differences with the present model are the presence of N components in the vegetation layer and taking into account only two hemispherical fluxes, i.e. downwards and upwards. The latter means that the model could only apply to overcast sky conditions. However model simula-tions have shown that daily interception is correctly estimated by the interception computed in overcast conditions, both in monocrops (Varlet-Grancher and Bonhomme, 1979) and mixtures (Sinoquet et al., 1990). The model is therefore aimed at estimating light partitioning at a daily scale, as required by growth analysis methods and most simulation models. KM equations are based on the balance of transmit-ted (T) and ascendant (A) radiation of an infinitesimal
(dimensionless) vegetation layer dz
whereσi is the scattering coefficient of leaves of
species i, i.e. the sum of leaf reflectance and transmit-tance (e.g. Goudriaan, 1977). The first term of right member of Eq. (5) accounts for radiation interception while the two last terms describe scattering of radia-tion intercepradia-tion within the small layer dz. Eq. (5) as-sumes that radiation is equally scattered upwards and downwards. Extinction coefficients Ki can explicitly
be related to foliage inclinationθiof species i, by using
values computed by Bonhomme and Varlet-Grancher (1977) (see Table 3, p. 572) in the case of a standard overcast sky (SOC, Moon and Spencer, 1942). Bon-homme and Varlet-Grancher’s extinction coefficients were related to leaf inclination as follows
K=0.988 cos
θ
2
2.4
r2=0.997 (6)
Maximum deviation between tabulated values and those computed from Eq. (6) was 2.8%. Kubelka and Munk (1931) gave the solution to Eq. (5) in the case of a turbid medium including only one compo-nent (see Bonhomme and Varlet-Grancher, 1977, for details). In the case of a mixture of N components, similar considerations can be used and lead to
α=
Constants C1 and C2 have to be derived from the
boundary conditions, i.e.At the top of the canopy (z=0),
T (0)=1 (10a)
At the ground level (z=1),
A (1)=ρsT (1) (10b)
where ρs is the soil reflectance. Rewriting Eq. (10)
from expressions of T and A given in Eq. (7) leads to a system of two equations where C1 and C2 are the
unknowns. Solving the system results in
C1=−
Transmitted and reflected fluxes (i.e. T(z) and A(z), respectively) may finally be written from Eq. (7) where parameters are given by Eqs. (8), (9), (11a), (11b) and (12). In particular, canopy transmittance Tc and
reflectanceρc are
In case of large LAI, the coefficientK′tends towards 0
and the canopy reflectance becomes that of an infinite canopyρ∞
ρ∞=α
β (15)
Canopy transmittance Tc and reflectanceρc can thus
be rewritten from Eqs. (13) and (14) in terms ofρ∞,
Eq. (17) makes clear that canopy reflectanceρcranges
fromρs toρ∞according to canopy development.
Light absorption efficiency of the whole canopy is then (e.g. Varlet Grancher et al., 1989)
ε=1−ρc−Tc(1−ρs) (18)
It can be seen from Eq. (5) that light is absorbed by component i in proportion of KiLi(1 –σi), hence the
light absorption efficiency of component i is
εi =
KiLi(1−σi)
PN
j=1KjLj 1−σj
ε (19)
2.2. Simplification of Kubelka–Munk equations (KMS) in a multispecies monolayer canopy
For dense enough canopies, neglecting exp(−K′)
with regard to exp(K′) is an approximation which
allows computations to be simplified. Eq. (11) then become
C1=0 and C2=
1
β (20)
Downward and upward fluxes may then be written from Eq. (7) as
T (z)=exp −K′z
(21a)
A (z)= α
β =ρ∞ (21b)
If the canopy includes only one component (i.e. N=1), Eq. (21) are the very same as the equations of Goudri-aan (1977). In particular, the apparent extinction coef-ficientK′=K√1−σ (see Goudriaan, 1977) (p. 26)
2.3. Simplified Kubelka–Munk equations (KMS) in a multispecies multilayer canopy
The multilayer canopy is described in terms of LAI (Lik) and mean leaf inclination (θik) of each
vegeta-tion component i in each layer k (k=1,. . ., M). Light transmission is computed from the top to the bottom of the canopy by successive applications of Eq. (21a). Light transmitted at the bottom of layer k is thus
Tk = k Y
l=1
exp −K′l
(22)
Upward fluxes at the layer boundaries are then re-cursively computed from the bottom to the top of the canopy by successive applications of Eq. (17). At ground level (i.e. the bottom of layer M),
AM =ρsTM (23)
For other layers k,
Ak =Tk
ρ∞k1−ρ∞k(Ak+1/Tk+1)+(Ak+1/
Tk+1)−ρ∞kexp −2K′k
1−ρ∞k(Ak+1/Tk+1)+ρ∞k(Ak+1/
Tk+1)−ρ∞kexp −2K′k
(24)
Eq. (24) is similar to Eq. (17) where incident radia-tion is Tk and soil reflectanceρs is replaced by the
ratio (Ak+1/Tk+1), i.e. equivalent to reflectance at the
bottom of layer k+1. Notice that the reflectance of the multilayer canopy can be computed from Eq. (24) with k=0.
Light absorption by vegetation layers and light partitioning between vegetation components are then computed from fluxes Tk and Ak by using
relation-ships similar to Eqs. (18) and (19).
3. Multispecies canopies for model application
3.1. Two-species monolayer canopies
The light models were applied to theoretical mono-layer canopies simulating well-mixed grass-legume mixtures. LAI of each species was varied between 0.25 and 5 in steps of 0.25, and all possible combinations of the two species were simulated. Leaf angle of Species
#1 and #2 were assumed to be 25 and 65◦, i.e. those
of a planophile and an erectophile species, such as numerous legumes and grasses (Ross, 1981), respec-tively. In a first series of simulation runs mimicking low scattering, leaf scattering coefficient was set to 0.20 for both species and soil reflectance was 0.15. In a second series of runs accounting for high scattering, leaf scattering coefficient and soil reflectance were set to 0.80 and 0.40, respectively.
3.2. Multispecies multilayer canopies
The light models were applied to multilayered mix-tures from canopy structure data taken from the litera-ture. The first one was a grass-legume mixture made of
Lolium perenne and Trifolium repens, where the two
components had similar heights (Faurie et al., 1996). Total LAI of the canopy ranged from 1.1 to 12.3. The plant canopy was divided into 2–6 horizontal layers of variable thickness, depending of growth stage. Leaf area index of the two species and mean leaf inclina-tion of L. perenne within each layer were computed from Faurie et al. (1996) (see Figs. 1 and 2, p. 38). Mean leaflet inclination of T. repens ranged from 25◦
at the bottom of the canopy to 45◦in the upper layer
(Soussana et al., 1995).
The second canopy was a binary mixture of
Vicia sativa and Avena sativa, (Ouknider and Jacquard,
1989). Total LAI ranged from 5.6 to 12.3. One or the other component dominated, i.e. was taller, de-pending on plant density, growth stage and nitrogen treatment. The canopy was divided into 3–6 layers of 20 cm where LAI of each component was measured (Ouknider and Jacquard, 1989) (see Figs. 4 and 5, p. 396). As leaf inclinations were not measured, they were assumed to range from the top to the bottom of the canopy: from 40 to 25◦for V. sativa and from 40
to 65◦for A. sativa, according to the rather planophile
and erectophile habit exhibited by the two species in the deeper canopy, respectively (Nichiporovitch, 1961).
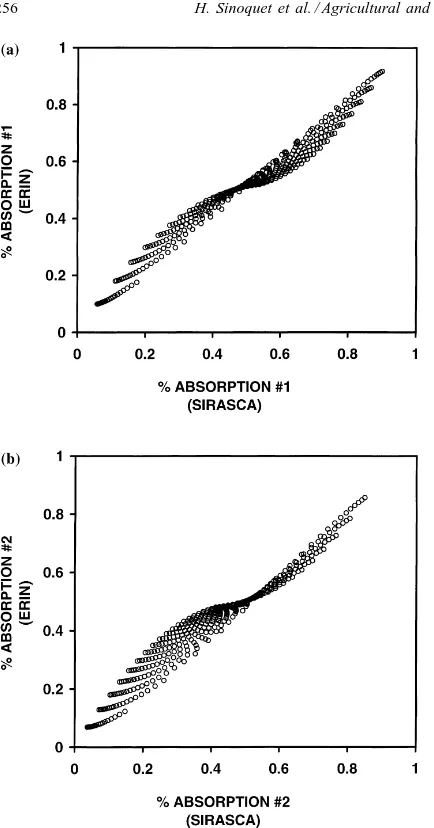
Fig. 1. Comparison between SIRASCA and ERIN models in case of monolayer canopies: fraction of absorbed radiation by planophile Species #1 (a) and erectophile Species #2 (b) under low scattering condition. See text for input parameters.
provided by Graf et al. (1990) (see Fig. 2, p. 376) for each component. The canopy was divided into four layers, the upper boundary of which corresponded to the height of the four components. The top layer only included leaf area of the dominating rice, while the bottom one consisted of a mixture of the four compo-nents. From the top to the bottom of the canopy, rice and grass weeds (i.e. Cyperaceae and other Poaceae) were attributed leaf inclinations ranging from 40 to 70◦, according to Ito et al. (1973) and Sheehy and
Fig. 2. Simulated fraction of absorbed radiation by planophile Species #1 (a) and erectophile Species #2 (b) in a monolayer canopy under low scattering conditions, as a function of species LAI, for three values of LAI of the other species: 1 (open sym-bols), 3 (grey symsym-bols), 5 (black symbols). Comparison between SIRASCA (j) KMS (d) and ERIN (m) models. See text for
input parameters.
Cooper (1973). Following Nichiporovitch (1961), leaf inclination of small dicots was assumed to be 25◦.
3.3. Model application to the multispecies canopies
Light partitioning between species in all mixed canopies described earlier was computed using SIR-ASCA (Sinoquet et al., 1990), ERIN (Wallace, 1997) and the KMS model proposed in this work.
SIRASCA was assumed to provide the reference computations, because it is a 1D version of a 3D model which has been tested against experimental data in a large number of contrasting canopies, e.g. grass–legume mixtures (Sinoquet et al., 1990; Fau-rie et al., 1996), row canopies (Sinoquet and Bon-homme, 1992; Andrieu and Sinoquet, 1993; Mabrouk et al., 1997), alley-cropping (Tournebize and Sinoquet, 1995). SIRASCA was run using LAI and mean leaf inclination of each plant species in each canopy layer, as well as leaf scattering coefficient of each species. Incident radiation was assumed to be an overcast sky obeying the SOC luminance distribution (Moon and Spencer, 1942).
ERIN was run with some modifications to the orig-inal version proposed by Wallace (1997). The origorig-inal extinction coefficients were replaced by those given by Eq. (6) (i.e. as a function of mean leaf inclina-tion), and they were weighed by√1−σ in order to account for leaf scattering (as proposed by Goudri-aan, 1977). Such changes were aimed at harmonising the models for the calculation of the extinction coeffi-cients, in order to compare the models only with regard to their ability in computing light partitioning. In the case of the monolayer canopy, the two-species were assumed to have the same height, while plant height was entered as described earlier for the multilayer canopies. ERIN was not applied to the four-component canopy.
With regard to the KM model proposed in this work, only the simplified equations as described in Section 2.3 were used, even in the case of the monolayer canopy.
4. Results
4.1. Two-species monolayer canopies
Under low scattering conditions, linear regres-sion analysis shows that the fraction of absorbed radiation computed by the KMS model is very
close to the reference model outputs, i.e. SIRASCA simulations
ε1K=1.006εS1+0.009 r2=1.000
ε2K=1.010εS2−0.023 r2=0.999
where subscript 1 and 2 refers to the planophile and erectophile species, and superscript K and S refers to KMS and SIRASCA models, respectively. The max-imum deviation between absorption value computed by the two models is 0.02 and 0.03 for the planophile Species #1 and erectophile Species #2, respectively. Mean residuals are respectively+0.012 and−0.018, indicating that light sharing between canopy compo-nents slightly favours the planophile species when computed by the KMS model.
Deviations between SIRASCA and ERIN outputs are much more important, as shown in Fig. 1 and the linear regression equations
ε1E=0.868εS1+0.081 r2=0.971
ε2E=0.864εS2−0.023 r2=0.953
where superscripts E and S refer to the ERIN and SIRASCA models, respectively. Maximum difference between light absorption computed by the models reaches 0.10 and 0.12 for Species #1 and #2, respec-tively. Fig. 1 clearly shows that outputs computed by the ERIN model are biased: residuals are not ran-domly distributed, and the scatter diagram looks like a family of curves corresponding to the combinations of the two species.
Fig. 2 shows simulated fraction of absorbed radia-tion as a funcradia-tion of LAI for each species, and for dif-ferent levels of the presence of the companion species. The deviations between SIRASCA and ERIN outputs are the lowest when the competing species exhibits a small LAI (LAI=1 in Fig. 2). When competition in-creases, ERIN underestimates the difference in light absorption by the two species, i.e. the strength of light competition.
Under high scattering conditions, linear regression analysis shows that the agreement between SIRASCA and KMS outputs is good only in the case of the planophile Species #1
ε1K=0.995εS1+0.009 r2=0.996
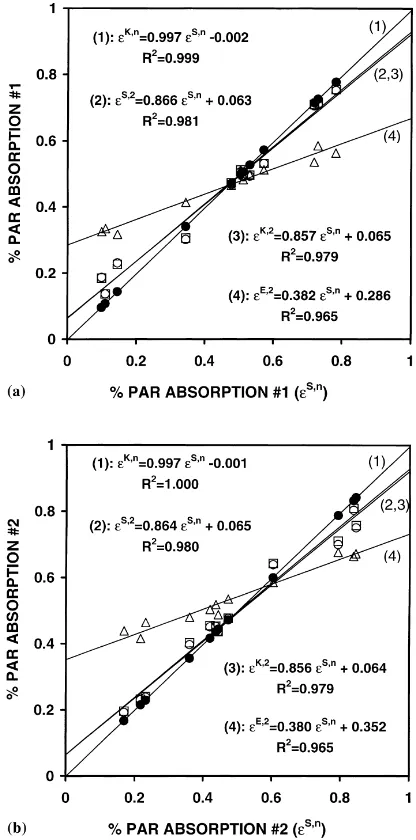
Fig. 3. Comparison between SIRASCA, KMS and ERIN models in the case of Trifolium repens–Lolium perenne canopies described by Faurie et al. (1996): fraction of PAR absorbed (ε) by Trifolium repens #1 (a) and Lolium perenne #2 (b). The mixture was simu-lated as either a multilayer (SIRASCA,εS,n, reference run; KMS:
εK,n,d) or a monolayer canopy (SIRASCA:εS,1,h; KMS:εK,1,
s; ERIN:εE,1,1). See text for input parameters.
where superscript K refers to the KMS model. KMS significantly underestimates (by 15%) light absorption by the erectophile Species #2: maximum deviation reaches –0.08 when light absorption is above 0.4. In contrast, ERIN largely overestimates light absorption
Fig. 4. Comparison between SIRASCA, KMS and ERIN models in the case of Vicia sativa–Avena sativa canopies described by Ouknider and Jacquard (1989): fraction of PAR absorbed (ε) by Vicia sativa #1 (a) and Avena sativa #2 (b). The mixture was simulated as either a multilayer (SIRASCA,εS,n, reference run; KMS:εK,n,d) or a two-layer canopy (SIRASCA:εS,2,h; KMS:
εK,2,s; ERIN:εE,2,1). See text for input parameters.
by the two species, i.e. by 50 and 28%, respectively, as can be shown from linear regression lines
ε1E=1.505ε1S+0.033 r2=0.994
The maximum deviation is 0.30 and 0.18 for Species #1 and #2, respectively.
4.2. Multispecies multilayer canopies
Fig. 3 shows the relationships between the fraction of PAR absorption computed by the three models in the T. repens–L. perenne mixture, i.e. where the height of both species was the same. The reference run was the SIRASCA simulation where the mixture was described as a multilayer canopy (according to the measurements by Faurie et al., 1996). The KMS model with the same canopy description gave results very close to the reference run: the maximum devia-tion was−0.04 and 0.01 for T. repens and L. perenne, respectively. When the canopy was described as a monolayer canopy, both SIRASCA and KMS mod-els significantly deviated from the reference model: the maximum deviations were −0.21 and −0.23, respectively. However the observed discrepancies were very similar for the two models, as suggested by the good agreement between SIRASCA and KMS outputs when applied to a monolayer canopy (Fig. 1). The monolayer models tend to smooth light compe-tition between the two species by overestimating the lowest light absorption efficiencies and
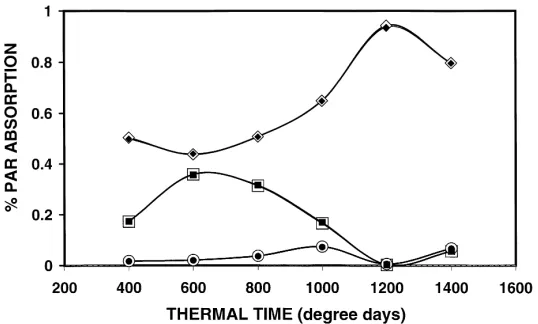
underestimat-Fig. 5. Simulated fraction of PAR absorbed by rice and two weed groups described by Graf et al. (1990), as a function of growth stage (in degree-days). Comparison between KMS (filled symbols) and Sirasca (open symbols) simulation. Rice (r), tall Cyperaceae, (j), and
other Poaceae (d). See text for input parameters.
ing the highest ones. This is a result of disregarding the vertical stratification of foliage within the canopy thickness. Deviations of ERIN outputs from the ref-erence run were highest and reached 0.24 and−0.29 for T. repens and L. perenne, respectively. The ERIN model markedly smoothed light competition between the two components. This resulted from neglecting the inter-species differences in LAI, leaf inclination and vertical distribution of leaf area.
The same general trends were found by applying the three models to the V. sativa–A. sativa mixture (Fig. 4). First PAR absorption as calculated by KMS using a multilayer description (i.e. according to the measurements by Ouknider and Jacquard, 1989) was very close to that given by the reference run. Second both SIRASCA and KMS models underestimated the strength of light competition when the mixture was represented as a two-layers canopy. Third ERIN simu-lations gave the greatest deviations from the reference run, with large underestimation of light competition between species.
are not shown since both measured and simulated val-ues ranged between 0 and 0.02).
5. Discussion
5.1. Methodology for model testing
This study was aimed at addressing the question of light partitioning between several species in a mixed canopy, especially grass–legume mixtures. The ques-tion of light partiques-tioning in well-mixed canopies (i.e. where foliages of the species are intermingled in the same canopy volume) cannot be addressed from light measurements, because there is no experimental de-sign to estimate the radiation balance of a given com-ponent in these conditions. The only way should be to use sensors attached to leaves. This was attempted to estimate light partitioning between shoots in a tree canopy (Sinoquet et al., 1997). Despite the number of sensors (60), the confidence in measurements was very low due to the large variability in leaf irradiance measurements. Moreover in case of small canopies like grass–legume mixtures, sensor settlement would be extremely difficult without disrupting canopy struc-ture. That is the reason why we chose to test the sim-ple models against simulations of the more comsim-plex SIRASCA model. Notice that Wallace (1997) did the same to test his ERIN model (but he used a smaller range of conditions). As SIRASCA is a 1D version of a radiation transfer model that has been used and tested in a number of contrasting situations (as men-tioned earlier), it was assumed to be able to serve as a reference model.
Model comparison was made from canopy structure data taken from the literature. In the reported experi-ments, LAI ranged from 1 to 12. Although this range might be regarded as a wide range, it corresponds to values that are found in natural conditions. The test was thought to be more powerful if it applies to a range of realistic values. The same applies to leaf scatter-ing coefficients, the range of which (0.2–0.8) approxi-mately corresponds to values found for leaves: 0.2 and 0.8 are typical values for the PAR and NIR wavebands, respectively. Testing the models in the PAR waveband is useful for primary production analysis (e.g. Mon-teith, 1972) while NIR domain should be included to compute the energy balance.
5.2. Use of simple models for estimating light partitioning between species
5.2.1. KMS model
The KMS model accurately summarised the de-tailed model SIRASCA under low scattering condi-tions in the case of various multispecies canopies: two-species monolayer (Fig. 1), two-species mul-tilayer (Figs. 3 and 4) and multispecies mulmul-tilayer canopies (Fig. 5). This model could therefore be used in growth analysis methods and simulation models dealing with horizontally homogeneous multispecies canopies, especially those requiring partitioning in the PAR domain. In comparison with other simple light models devoted to partitioning (see Section 1), KMS is able to adapt to a wide range of canopies and to explicitly take into account the main deter-minants of light microclimate: canopy structure (i.e. LAI and mean leaf inclination of each species in each canopy layer) and optical properties of leaves (of each species) and the soil surface. In the version proposed in this paper, the model assumes overcast sky conditions. This is because the model is aimed at simulating light partitioning between species at a daily scale. Several studies showed that light absorp-tion at a daily scale is correctly approximated by a single run in overcast conditions in both monospecies and multispecies canopies (Varlet-Grancher and Bon-homme, 1979; Sinoquet et al., 1990). In order to use the model at smaller time steps, direct beam flux should be included in the model, in a way similar to Bonhomme and Varlet-Grancher (1977) in the case of monocrops. Last but not least, the model is easily implemented since it only includes Eqs. (6) and (8) for the extinction coefficient as a function of leaf inclination and leaf optical properties, Eq. (9) for coefficients α and β, and the recursive use of Eqs. (22), (24), (18) and (19) to compute light balance and partitioning in each canopy layer.
The derivation of the KMS model made it possi-ble to bridge the Kubelka–Munk theory and the model by Goudriaan (1977), where scattering is taken into account by correcting the extinction coefficient by a factor√(1−σi). Goudriaan’s proposal avoids
Goudriaan’s model can be interpreted as a two-flux KM model (upward/downward) where exp(−K′z) is
neglected with regard to exp(K′z). Reciprocally the
KMS model proposed in this paper can be regarded as an extension of Goudriaan’s to the case of multilayer multispecies canopies.
When applied to high scattering conditions, KMS correctly simulated light absorption of planophile Species #1 but significantly underestimated light in-terception by the erectophile Species #2. When using the complete KM equations, the same behaviour was observed (data not shown). As previously pointed out by Myneni et al. (1989), the KM approach does not take into account differences in directional light in-terception. By dealing with hemispherical fluxes, the KM approach implicitly assumes that the directional distribution of radiation fluxes does not change within the canopy, while it does in real canopies. Varia-tions in extinction coefficients with beam direction depend on foliage inclination (see e.g. Ross, 1981). Planophile foliage shows less change in extinction coefficients with beam direction than erectophile fo-liage (in particular, the projection coefficient of hor-izontal leaves is unity whatever the beam direction, Ross, 1981). The large directional variations of light extinction in erectophile canopies result in modifica-tion of the direcmodifica-tional distribumodifica-tion of radiamodifica-tion fluxes within the canopy while the extinction coefficient of the canopy (see Eq. (6)) is calculated according to the radiance distribution above the canopy. In agreement with Myneni et al. (1989) this suggests that models based on the KM equations would poorly estimate the radiation balance of erectophile canopies, especially under high scattering conditions.
5.2.2. ERIN model
When applied to monolayer mixed canopies, the ERIN model correctly simulated light partitioning be-tween species in some, but not in all, conditions (Figs. 1 and 2). This is because ERIN shares light between canopy components according to their height rather than their LAI (Wallace, 1997). In case of a monolayer canopy, the two components have the same height, so that the fraction of light absorbed by component i is simply the mean value between maximal and mini-mal values (i.e.εimaxandεimin computed in case of
dominance of either one or the other species, see Eq.
(3)). This estimation is correct when the difference be-tweenεiminandεimaxis low, i.e. when the companion
species have a reduced competitive ability (namely a small LAI). The ERIN model can also give correct es-timations if the species ability to capture radiation (i.e. the term KiLi(1−σi)) is the same for the two species.
Wallace (1997) compared ERIN with SIRASCA in conditions of equal LAI and leaf inclination, and he found a good agreement between the two models. This behaviour of ERIN can also be seen in Fig. 2a: ERIN lines cross SIRASCA lines when the LAI of the two species are similar. In contrast, ERIN fails in simulat-ing light partitionsimulat-ing between species when the two species have high but different LAI (Fig. 2). Due to the equal height of the two species in the monolayer canopy, light competition as simulated by ERIN is less intense than that computed from SIRASCA (Fig. 2).
In case of high scattering conditions, ERIN greatly overestimated light absorption by the two canopy com-ponents, mostly because upward radiation fluxes are not taken into account while they largely contribute to radiation lost by the canopy (i.e. reflected to the sky).
5.3. Requirements in canopy structure parameters
Reducing the requirements for canopy structure parameters in a light model considerably simplifies the inclusion of light partitioning in crop simulation models. Due to low requirements for canopy struc-ture parameters, ERIN would be the best model for inclusion in crop simulation models. However parti-tioning light according only to height difference (Eq. (4)) leads to large underestimation of light compe-tition, i.e. between-component differences in light absorption are underestimated.
This effect also exists, but to a lesser extent, when the components have different heights. These results are not fully consistent with previous simulation re-sults, where Sinoquet and Caldwell (1995) showed that the main determinants of light partitioning in two-species mixtures were: (i) the relative height of the two components and (ii) differences in leaf incli-nation. The vertical profile of leaf area density was claimed to be of least importance. These conclusions were drawn from computations with the SIRASCA model involving theoretical vertical profiles of leaf area density. Such results would have suggested that describing the canopy as a monolayer or a two-layer canopy would be enough. In another approach, Faurie et al. (1996) proposed that light partitioning between species be related to their relative contribution to the upper canopy layer of total LAI equal to 3, because light transmission below a LAI of 3 is negligible. Al-though useful, such an approach requires knowledge of the vertical profile of leaf area in order to define the upper layer of LAI equal to 3, and the contribution of the vegetation components to this layer.
Finally, it appears that a simplified description of canopy structure leads to biased estimation of light partitioning between species. This means that a de-scription of canopy structure in terms of LAI of each component in each layer would be necessary. How-ever, current crop simulation models for multispecies canopies are unable to simulate canopy structure to the required degree.
6. Conclusion
The ERIN model was shown to give biased estima-tions of light partitioning in a number of situaestima-tions. This is because ERIN computes light partitioning only as a function of height differences while other param-eters like LAIs, leaf inclination and the vertical distri-bution of leaf area have also a significant effect.
The KMS model proposed in this paper was shown to accurately summarise the more detailed light parti-tioning model SIRASCA. Moreover KMS is flexible since it can be applied to mixed canopies including any number of vegetation layers and components. The parameters used in the model are clearly related to physical properties of the canopies, i.e. the vertical dis-tribution of LAI, leaf inclination and optical properties
of each vegetation component. The model deals with two hemispherical fluxes, which means that incident radiation is only diffuse. Such an assumption makes the model suitable for simulating light partitioning at a daily scale. A third beam flux could however be eas-ily included to simulate light partitioning at smaller time steps. At present, the model KMS appears to be a good candidate for inclusion in growth models dealing with multispecies canopies at a daily scale.
References
Allen, L.H., Sinclair, T.R., Lemon, E.R., 1976. Radiation and microclimate relationships in multiple cropping systems. In: Papendick, R.A., Sanchez, P.A., Triplett, E.B. (Eds.), Multiple cropping. ASA Special Publication 27, Madison, WI, pp. 171–200.
Andrieu, B., Sinoquet, H., 1993. Evaluation of structure description requirements for predicting gap fraction of vegetation canopies. Agric. For. Meteorol. 65, 207–227.
Azam-Ali, S.N., Matthews, R.B., Williams, J.B., Peackock, J.M., 1990. Light use, water uptake and performance of individual components of a sorghum/groundnut intercrop. Exp. Agric. 26, 413–427.
Bonhomme, R., Varlet-Grancher, C., 1977. Application aux couverts végétaux des lois de rayonnement en milieu diffusant I. Etablissement des lois et vérifications expérimentales. Ann. Agron. 28, 567–582.
Caldwell, M.M., 1987. Plant architecture and resource competition. In: Schulze, E.D., Zwölfer, H. (Eds.), Ecological Studies 61. Springer, Berlin, pp. 164–179.
Caldwell, R.M., Hansen, J.W., 1993. Simulation of multiple cropping systems with CropSys. In: Penning de Vries, F., Teng, P., Metselaar, K. (Eds.), Systems approaches for agricultural development. Kluwer Academic Publishers, Dordrecht, The Netherlands, pp. 397–412.
Campbell, G.S., Van Evert, F.K., 1994. Light interception by plant canopies: efficiency and architecture. In: Monteith, J.L., Scott, R.K., Unsworth, M.H. (Eds.), Resource capture by crops. Nottingham University Press, Nottingham, pp. 35–52. Cohen, S., Fuchs, M., 1987. The distribution of leaf area, radiation,
photosynthesis and transpiration in a shamouti orange hedgerow canopy I. Leaf area and radiation. Agric. For. Meteorol. 40, 13–144.
Cruz, P., 1995. Use of the RUE concept for analysing growth of pure and mixed tropical forage crops. In: Sinoquet, H., Cruz, P. (Eds.), Ecophysiology of tropical intercropping. INRA, Paris, pp. 319–330.
Cruz, P., Sinoquet, H., 1994. Competition for light and nitrogen during a regrowth cycle in a tropical forage mixture. Field Crops Res. 36, 21–30.
Goudriaan, J., 1977. Crop micrometeorology, a simulation study. Simulation Monographs, Pudoc, Wageningen, The Netherlands, 249 pp.
Graf, B., Gutierrez, A.P., Rakotobe, O., Zahner, P., Delucchi, V., 1990. A simulation model for the dynamics of rice growth and development 2. The competition with weeds for nitrogen and light. Agric. Syst. 32, 367–392.
Harris, D., Natarajan, M., Willey, R.W., 1987. Physiological basis for yield advantage in a sorghum/groundnut intercrop exposed to drought I. Dry matter production, yield and light interception. Field Crops Res. 17, 259–272.
Ito, A., Udagawa, T., Uchijima, Z., 1973. Phytometrical studies of crop canopies II. Canopy structure of rice crops in relation to varieties and growing stage. Proc. Crop. Sci. Soc. Jap. 42, 334–342 (in Japanese).
Kiniry, J.R., Williams, J.R., Gassman, P.W., Debaeke, P., 1992. A general process-oriented model for two competing plant species. Trans. ASAE 35, 801–810.
Kubelka, P., Munk, F., 1931. Ein Beitrag zur Optik der Farbenstriche. Z. Tech. Phys. 12, 593–601.
Mabrouk, H., Sinoquet, H., Carbonneau, A., 1997. Canopy structure and radiation regime in grapevine II. Modeling radiation interception and distribution inside the canopy. Vitis 36, 125–132.
McMurtrie, R., Wolf, L., 1983. A model of competition between trees and grass for radiation, water and nutrients. Ann. Bot. 52, 449–458.
Monteith, J.L., 1972. Solar radiation and productivity in tropical ecosystems. J. Appl. Ecol. 9, 747–766.
Moon, P., Spencer, D.E., 1942. Illumination from a non-uniform sky. Trans. Illum. Eng. Soc. 37, 707–726.
Myneni, R.B., Ross, J., Asrar, G., 1989. A review on the theory of photon transport in leaf canopies. Agric. For. Meteorol. 45, 1–153.
Nichiporovitch, A.A., 1961. On properties of plants as an optical system. Sov. Plant Physiol. 8, 536–546 (in Russian).
Ouknider, M., Jacquard, P., 1989. Variabilité des phénomènes d’interférence entre Vicia sativa L. et Avena sativa L. I. Dynamique de croisance de la vesce dans un peuplement associé de vesce-avoine. Agronomie 9, 391–400.
Rimmington, G.M., 1984. A model of the effects of interspecies competition for light on dry-matter production. Aust. J. Plant Physiol. 11, 277–286.
Rimmington, G.M., 1985. A test of a model for light interception by mixtures. Aust. J. Plant Physiol. 12, 681–683.
Ross, J., 1981. The Radiation Regime and Architecture of Plant Stands. Junk, The Hague, 391 pp.
Ryel, R.J., Barnes, P.W., Beyschlag, W., Caldwell, M.M., Flint, S.D., 1990. Plant competition for light analyzed with a multispecies canopy model 1. Model development and influence of enhanced UV-B conditions on photosynthesis in mixed wheat and wild oat canopies. Oecologia 82, 304– 310.
Sheehy, J.E., Cooper, K., 1973. Light interception, photosynthetic activity, and crop growth rate in canopies of six temperate forage grasses. J. Appl. Ecol. 10, 239–250.
Sinoquet, H., Bonhomme, R., 1991. A theoretical analysis of radiation interception in a two species plant canopy. Math. Biosci. 105, 23–45.
Sinoquet, H., Bonhomme, R., 1992. Modeling radiative transfer in mixed and row intercropping systems. Agric. For. Meteorol. 62, 219–240.
Sinoquet, H., Caldwell, R.M., 1995. Estimation of light capture and partitioning in intercropping systems. In: Sinoquet, H., Cruz, P. (Eds.), Ecophysiology of Tropical Intercropping. INRA, Paris, pp. 79–97.
Sinoquet, H., Moulia, B., Gastal, F., Bonhomme, R., Varlet-Grancher, C., 1990. Modeling the radiative balance of the components of a well-mixed canopy: application to a white clover–tall fescue mixture. Acta Oecol. 11, 469–486. Sinoquet, H., Adam, B., Rivet, P., Godin, C., 1997. Interactions
between light and plant architecture in an agroforestry walnut tree. Agrofor. Forum 8 (2), 37–40.
Soussana, J.-F., Faurie, O., Sinoquet, H., 1995. Radiation and nitrogen acquisition and use in simulated grass–clover swards. In: Sinoquet, H., Cruz, P. (Eds.), Ecophysiology of Tropical Intercropping. INRA, Paris, pp. 331–341.
Soussana, J.-F., Teyssonneyre, F., Thiéry, J.-M., 2000. Un modèle simulant les compétitions pour la lumière et pour l’azote entre espèces herbacées à croissance clonale. In: Bonhomme, R., Maillard, P. (Eds.), Fonctionnement des Plantes sous Contraintes Environnementales. INRA, Paris, in press.
Spitters, C.J.T., Aerts, R., 1983. Simulation of competition for light and water in crop–weed associations. Aspects Appl. Biol. 4, 467–483.
Tournebize, R., Sinoquet, H., 1995. Light interception and partitioning in a shrub–grass mixture. Agric. For. Meteorol. 72, 277–294.
Varlet-Grancher, C., Bonhomme, R., 1979. Application aux couverts végétaux des lois de rayonnement en milieu diffusant II. Interception de l’énergie solaire par une culture. Ann. Agron. 30, 1–26.
Varlet Grancher, C., Gosse, G., Chartier, M., Sinoquet, H., Bonhomme, R., Allirand, J.M., 1989. Mise au point: rayonnement solaire absorbé ou intercepté par un couvert végétal. Agronomie 9, 419–439.
Wallace, J.S., 1997. Evaporation and radiation interception by neighbouring plants. Q. J. R. Meteorol. Soc. 123, 1885–1905. Wallace, J.S., Batchelor, C.H., Dabeesing, D.N., Teeluck, M., Soopramanien, G.C., 1991. A comparison of the light interception and water use of plant and first ratoon sugar cane intercropped with maize. Agric. For. Meteorol. 57, 85–105. Wiles, L.J., Wilkerson, G.G., 1991. Modeling competition for light