Latar Belakang
Indonesia memiliki lahan gambut tropika terluas di dunia. Noor dan Heyde (2007), menyatakan luas lahan gambut Indonesia adalah 20,6 juta hektar. Luas tersebut berarti sekitar 50% dari luas seluruh lahan gambut tropika (40 juta hektar) atau sekitar 10,8% dari luas daratan Indonesia. Jika dilihat penyebarannya, lahan gambut sebagian besar terdapat di Sumatra (sekitar 35%), Kalimantan (sekitar 30%), Papua (sekitar 30%) dan Sulawesi (sekitar 3%) seperti pada Tabel 1.
Tabel 1. Perkiraan luas lahan gambut di Indonesia menurut beberapa sumber.
Penulis/Sumber Penyebaran Gambut (Juta ha.) Total Sumatra Kalimantan Papua Lainnya
Driessen (1978) 9,7 6,3 0,1 - 16,1
Puslittanak (1981) 8,9 6,5 10,9 0,2 26,5
Euroconsult (1984) 6,84 4,93 5,46 - 17,2 Soekardi dan Hidayat (1988) 4,5 9,3 4,6 < 0,1 18,4
Deptrans (1988) 8,2 6,8 4,6 0,4 20,1
Subagyo et al. (1990) 6,4 5,4 3,1 - 14,9
Deptrans (1990) 6,9 6,4 4,2 0,3 17,8
Nugroho et al. (1992) 4,8 6,1 2,5 0,1 13,5* Radjagukguk (1993) 8,25 6,79 4,62 0,4 20,1 Dwiyono dan Rachman (1996) 7,16 4,34 8,40 0,1 20,0 *Tidak termasuk gambut yang berasosiasi dengan lahan salin dan lahan lebak (2,46 juta ha). Sumber: Noor dan Heyde (2007).
Hutan rawa gambut tropis sedang mengalami tekanan berat dari kegiatan pengembangan pertanian/silvikultur dan penebangan. Kecenderungan penutupan lahan di Asia Tenggara dikembangkan dari perubahan antara tahun 1985 dan tahun 2000. Selama periode ini, lahan gambut mengalami laju deforestasi rata-rata sebesar 1,3% per tahun, nilai tertinggi ditemukan di Kalimantan Timur (2,8%/tahun), terendah di Papua (0,5%/tahun) (Hooijer et al., 2006).
Pada saat ini introduksi mikoriza merupakan teknologi yang tidak bisa ditawar lagi untuk meningkatkan keberhasilan rehabilitasi hutan dan lahan terdegradasi di Indonesia (Santoso et al., 2007). Kendala utama yang dihadapi dalam kegiatan rehabilitasi lahan gambut adalah sifat unik gambut dapat dilihat dari sifat kimia dan fisiknya. Lahan gambut memiliki sifat yang spesifik antara satu dengan yang lainnya. Menurut Najiyati et al. (2005), sifat kimia gambut yang lebih merujuk pada kondisi kesuburannya yang bervariasi, tetapi secara umum memiliki kesuburan rendah. Hal ini ditandai dengan tanah yang masam (pH rendah), ketersediaan sejumlah unsur hara makro (K, Ca, Mg, P) dan mikro (Cu, Zn, Mn, dan Bo) rendah, mengandung asam-asam organik beracun, serta memiliki Kapasitas Tukar Kation (KTK) yang tinggi tetapi Kejenuhan Basa (KB) rendah. Sifat fisik gambut yang perlu dipahami antara lain menyangkut kematangan, warna, berat jenis, porositas, kering tak balik, subsidensi, dan mudah terbakar.
Fungi mikoriza arbuskula (FMA) mempunyai pengaruh positif terhadap pertumbuhan dan proses fisiologi pada tanaman. Rosliani et al. (2006), menyatakan pengaruh menguntungkan dari fungi mikoriza arbuskula terhadap pertumbuhan tanaman sering dihubungkan dengan peningkatan serapan hara yang tidak tersedia terutama fosfor (P).
Mikoriza tidak hanya berkembang pada tanah berdrainase baik, tapi juga pada lahan tergenang. Bahkan pada lingkungan yang sangat miskin atau lingkungan yang tercemar limbah berbahaya, fungi mikoriza masih memperlihatkan eksistensinya. Salah satu bentuk lingkungan yang mencerminkan keadaan demikian dapat ditemui pada tipe tanah Histosol atau yang lebih umum disebut tanah gambut (Hanafiah, 2004).
Rehabilitasi lahan gambut dengan aplikasi mikoriza dapat diterapkan, apabila telah didapat isolat mikoriza yang dapat bersimbiosis baik dengan tanaman yang akan dikembangkan. Penelitian terdahulu telah mengeksplorasi keanekaragamanan mikoriza pada beberapa lahan gambut. Namun setiap lahan gambut memiliki sifat yang spesifik dan ditumbuhi dengan jenis tanaman yang berbeda, sehingga menyebabkan FMA lokal yang didapat berbeda juga. Dengan demikian dibutuhkan penelitian keanekaragaman mikoriza, guna dapat direkomendasikan dan diaplikasi untuk rehabilitasi lahan tersebut.
Tujuan
Untuk mengetahui keanekaragaman dan kepadatan spora FMA pada tegakan karet dan tegakan sawit di ekosistem lahan gambut Desa Telaga Suka, Kecamatan Panai Tengah, Kabupaten Labuhan Batu.
Kegunaan Penelitian
Memberikan informasi mengenai keanekaragaman FMA pada lahan gambut di Desa Telaga Suka, Kecamatan Panai Tengah, Kabupaten Labuhan Batu dan sebagai rekomendasi untuk pemanfaatan tipe FMA yang ditemukan untuk rehabilitasi lahan gambut tersebut.
TINJAUAN PUSTAKA
Kondisi Lahan Gambut
Lahan gambut merupakan lahan yang berasal dari bentukan gambut beserta vegetasi yang terdapat diatasnya, terbentuk di daerah yang topografinya rendah dan bercurah hujan tinggi atau di daerah yang suhunya sangat rendah. Tanah gambut mempunyai kandungan bahan organik yang tinggi (>12% karbon) dan kedalaman gambut minimum 50 cm (Rina et al., 2008).
Secara umum definisi tanah gambut adalah tanah yang jenuh air dan tersusun dari bahan tanah organik, yaitu sisa- sisa tanaman dan jaringan tanaman yang melapuk dengan ketebalan lebih dari 50 cm. Dalam sistem klasifikasi baru (taksonomi tanah), tanah gambut disebut sebagai Histosols (histos = jaringan) (Noor dan Heyde, 2007).
Menurut Agus dan Subiksa (2008), berdasarkan tingkat kematangannya gambut dibedakan menjadi:
• Gambut saprik (matang) adalah gambut yang sudah melapuk lanjut dan bahan asalnya tidak dikenali, berwarna coklat tua sampai hitam, dan bila diremas kandungan seratnya < 15%.
• Gambut hemik (setengah matang) adalah gambut setengah lapuk, sebagian bahan asalnya masih bisa dikenali, berwarma coklat, dan bila diremas bahan seratnya 15 – 75%.
• Gambut fibrik (mentah) adalah gambut yang belum melapuk, bahan asalnya masih bisa dikenali, berwarna coklat, dan bila diremas >75% seratnya masih tersisa.
Tanah Gambut dan Permasalahannya
Kesuburan tanah gambut dipengaruhi oleh kedalaman dan lapisan mineral di bawah gambut. Makin tebal gambut makin miskin lapisan atasnya. Gambut yang terbentuk di atas endapan pasir kuarsa lebih miskin dari gambut yang terbentuk di atas endapan liat. Menurut Noor (2001), secara kimiawi sifat tanah gambut yang utama adalah kemasaman tanah, ketersediaan hara tanah, kapasitas tukar kation, kejenuhan basa, kadar asam organik tanah, kadar pirit atau sulfur. Sifat-sifat kimia tanah ini sangat penting dalam penentuan jenis dan cara-cara pengelolaan hara dan pupuk dalam budidaya tanaman pertanian.
Lahan gambut mempunyai potensi yang cukup baik untuk pengembangan tanaman kehutanan maupun perkebunan. Namun dalam pengembangannya terdapat beberapa kendala seperti ketebalan dan kematangan gambut, bobot isi (BD) sangat rendah, kemasaman tanah, miskin unsur hara makro (K, Ca, Mg, P) dan mikro (Cu, Zn, Mn, dan Bo) serta keracunan asam-asam organik dan/atau pirit yang teroksidasi. Kelebihan air yang umum terjadi (seperti adanya banjir/genangan dalam jangka waktu yang lama pada musim hujan) harus dikendalikan menurut kebutuhan tanaman. Masalah-masalah tersebut, menyebabkan keberhasilan tumbuh tanaman menjadi sangat rendah atau bahkan mengalami kegagalan (Wibisono et al., 2005).
Selain itu, tanah gambut juga memiliki nilai Kapasitas Tukar Kation (KTK) yang tinggi tetapi Kejenuhan Basa (KB) rendah sehingga menyebabkan pH tanah rendah dan sejumlah pupuk yang diberikan ke dalam tanah relatif sulit terserap oleh akar tanaman. Pada umumnya lahan gambut tropis memiliki pH antara 3 - 4,5 dimana gambut dangkal mempunyai pH lebih tinggi (pH 4,0–5,1)
dari pada gambut dalam (pH 3,1–3,9). Kandungan Al pada tanah gambut umumnya rendah sampai sedang dan semakin berkurang seiring dengan menurunnya pH tanah. Sebaliknya, kandungan besi (Fe) cukup tinggi. Kandungan N total termasuk tinggi, namun umumnya tidak tersedia bagi tanaman, oleh karena rasio C/N yang tinggi (Wibisono et al., 2005).
Mikoriza
Suatu bentuk hubungan yang saling menguntungkan antara akar tanaman dan fungi disebut mikoriza. Dalam Bahasa Yunani kata mikoriza berarti fungi akar, yang dikemukakan oleh Frank pada tahun 1885 untuk menggambarkan asosiasi simbiotik antara akar tanaman dan fungi (Manoharacary et al., 2009). Mikoriza adalah suatu struktur sistem perakaran yang terbentuk sebagai manifestasi adanya simbiosis mutualisme antara cendawan (Myces) dan perakaran (Rhizo) tumbuhan tingkat tinggi (Setiadi, 2001).
Sedikitnya tujuh jenis asosiasi mikoriza yang berbeda telah dikenali, menyertakan kelompok fungi yang berbeda dan tanaman inang dan bentuk pola asosiasi yang berbeda. Adapun asosiasi tersebut sebagai berikut:
1. Vesikula Arbuskula Mikoriza (VAM), di mana fungi Zygomysetes ini memproduksi arbuskula, hifa, dan vesikula di dalam sel korteks akar.
2. Ektomicoriza (ECM), dimana fungi basidiomycetes dan fungi lainnya membentuk suatu mantel yang menyelubungi sekeliling akar dan jaringan hartig diantara sel akar.
3. Mikoriza Anggrek, dimana fungi memproduksi kumparan hifa di dalam akar atau batang tanaman anggrek-anggrekan.
4. Ericoid Mikoriza, merupakan kumparan hifa diluar sel yang membatasi akar rambut tanaman, pada tanaman ordo Ericales, dan
5. Ektendo, Arbutoid, dan Monotropoid, dimana asosiasinya mirip asosiasi ektomikoriza, namun memiliki perbedaan pada fitur anatominya (Brundett et al., 1996).
Menurut Turk et al. (2006), pembagian mikoriza yang dibedakan berdasarkan morfologi dan fisiologinya yakni endomikoriza dan ektomikoriza. Ektomikoriza ditandai dengan suatu sarung pelindung yang melingkupi akar, seringkali menembus hingga sel epidermis dan sel awal korteks dan hifa fungi biasanya menginfeksi akar tanaman hutan pada wilayah sub-tropis. Sedangkan endomikoriza seperti Vesikula Arbuskula Mikoriza (VAM), fungi tidak membentuk selubung. Fungi ini menginfeksi sistem perakaran tanaman budidaya, secara umum dan biasanya menginfeksi beberapa lapisan terluar korteks akar. Hifa fungi VAM menembus sel individu dan membentuk arbuskula di dalam sel dan vesikula di luar sel inang.
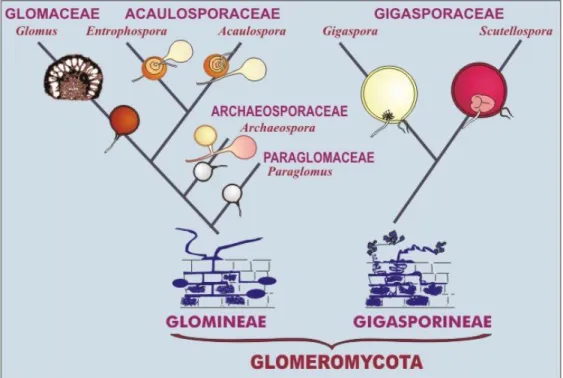
Klasifikasi Fungi Mikoriza Arbuskula
Pengenalan dan pengelompokan dalam spora mikoriza vesikular arbuskula saat ini dilakukan lebih didasarkan kepada struktur subselular dengan verifikasi teknologi molekular, mikoriza vesikular arbuskula dikelompokkan ke dalam ordo
Glomales, sub ordo Glomineae dan Gigasporineae. Glomineae terdiri dari empat
famili (Glomaceae, Acaulosporaceae, Aracheosporaceae dan Paraglomaceae). Sementara Gigasporineae terdiri dari lima famili yaitu Ehtrophospora,
Aracheospora, Paraglomus, Gigaspora dan Scutellspora. Salah satu karakteristik
dan reproduksi spora, reaksi melzer, keberadaan struktur subselular diantaranya
spore wall dan germinal wall, asesoris, serta struktur mikoriza yang terbentuk
dalam akar (Fakuara, 1988).
Fungi mikoriza arbuskula adalah salah satu tipe fungi mikoriza dan termasuk ke dalam golongan endomikoriza. Fungi mikoriza arbuskula termasuk ke dalam kelas Zygomycetes, dengan ordo Glomales yang mempunyai 2 sub-ordo, yaitu Gigasporineae dan Glomineae. Gigasporineae dengan famili Gigasporaceae mempunyai 2 genus, yaitu Gigaspora dan Scutellospora. Glomaceae mempunyai 4 famili, yaitu famili Glomaceae dengan genus Glomus dan Sclerocystis, famili Acaulosporaceae dengan genus Acaulospora dan
Entrophospora, Paraglomaceae dengan genus Paraglomus, dan Archaeosporaceae
dengan genus Archaeospora (Delvian, 2005). Klasifikasi dan perkembangan fungi mikoriza arbuskula ini dapat dilihat pada Gambar 1.
Gambar 1. Phylogeny perkembangan dan taksonomi ordo Glomales sumber : http://invam.caf.wvu.edu/Myc-_info/Taxonomy/classification.htm
Menurut Brundett et al. (1996), bagian-bagian penting spora yang digunakan dalam mengidentifikasi fungi Glomalen (VAM) adalah perkembangan spora, susunan spora, bentuk spora, ukuran spora, warna spora, ornamen spora, lapisan dinding spora dan reaksi pewarnaan, isi spora, germinasi spora hifa tanah, dan struktur asosiasi spora dengan hifa tanah.
Struktur Umum Fungi Mikoriza Arbuskula
Struktur FMA meliputi hifa eksternal, hifa internal, spora, arbuskula atau vesikula. Infeksi fungi hanya pada korteks primer sehingga tidak menyebabkan kerusakan pada jaringan akar. Proses infeksi dimulai dengan pembentukan apresorium pada permukaan akar oleh hifa eksternal, dan selanjutnya hifa akan menembus sel-sel korteks akar melalui rambut akar atau sel epidermis. Hifa dari FMA tidak bersekat, hifa ini terdapat diantara sel-sel korteks akar dan becabang-cabang di dalamnya, tetapi tidak sampai masuk ke jaringan stele. Di dalam sel-sel yang terinfeksi terbentuk gelung hifa atau cabang-cabang hifa kompleks yang dinamakan arbuskula (Moose, 1981).
Mikoriza vesikula arbuskula membentuk struktur karakteristik khusus yang disebut arbuskel dan vesikel. Arbuskel membantu dalam mentrasfer hara (terutama fosfat) dari tanah ke sistem perakaran (Rao, 2004). Arbuskula merupakan hifa bercabang halus yang dibentuk oleh percabangan dikotomi yang berulang-ulang sehingga menyerupai pohon dari dalam sel inang (Pattimahu, 2004).
Vesikel merupakan struktur cendawan yang berasal dari pembengkakan hifa internal secara terminal dan interkalar, kebanyakan berbentuk bulat telur, dan berisi banyak senyawa lemak sehingga merupakan organ penyimpanan cadangan
makanan dan pada kondisi tertentu dapat berperan sebagai spora atau alat untuk mempertahankan kehidupan cendawan. Tipe FMA vesikel memiliki fungsi yang paling menonjol dari tipe cendawan mikoriza lainnya. Hal ini dimungkinkan karena kemampuannya dalam berasosiasi dengan hampir 90 % jenis tanaman, sehingga dapat digunakan secara luas untuk meningkatkan probabilitas tanaman (Pattimahu, 2004).
Spora terbentuk pada ujung hifa eksternal. Spora ini dapat dibentuk secara tunggal, berkelompok atau di dalam sporokarp tergantung pada jenis cendawannya. Perkecambahan spora sangat sensitif tergantung kandungan logam berat di dalam tanah dan juga kandungan Al, kandungan Mn juga mempengaruhi pertumbuhan miselium. Spora dapat hidup di dalam tanah beberapa bulan sampai beberapa tahun. Namun untuk perkembangan FMA memerlukan tanaman inang. Spora dapat disimpan dalam waktu lama sebelum digunakan lagi (Mosse, 1981).
Infeksi FMA ditandai dengan produksi dan pembengkakan dinding interior intraseluler vesikel yang diyakini berfungsi sebagai organ penyimpan cadangan makanan dan formasi intraseluler tersebut membentuk formasi berupa arbuskula. Arbuskula dipertimbangkan menjadi struktur primer termasuk secara langsung dalam transfer unsur hara antara fungi simbion dengan tanaman inang. Walaupun secara umum hal tersebut berlaku pada FMA endofit, namun Gigaspora spp. hanya ditemukan arbuskula (Brown dan King, 1991).
Mosse (1981), mengamati bahwa struktur yang dibentuk pada akar-akar muda adalah arbuskul. Bertambahnya umur menyebabkan arbuskul berubah menjadi suatu struktur yang menggumpal dan cabang-cabang pada arbuskul lama kelamaan tidak dapat dibedakan lagi. Pada akar yang telah dikolonisasi oleh FMA
dapat dilihat berbagi arbuskul dewasa yang dibentuk berdasarkan umur dan letaknya. Arbuskul dewasa terletak dekat pada sumber unit kolonisasi tersebut.
Distribusi dan Ekologi Fungi Mikoriza Arbuskula
Mikoriza terdistribusi dalam jangkauan ekosistem yang sangat luas, namun memiliki pola penyebaran yang berbeda antar tipe mikoriza berdasarkan bioma, tipe tanah, dan keterbatasan sumber daya (Read 1983 dalam Bardgett 2008). Fungi mikoriza arbuskula mulai ditemukan pada profil tanah sekitar kedalaman 20 cm. Tetapi walaupun demikian juga, masih terdapat pada kedalaman 70-100 cm. FMA tersebar secara aktif (tumbuh dengan mycelium dalam tanah) dan tersebar secara pasif dimana FMA tersebar dengan angin, air atau mikroorganisme dalam tanah (Coyne, 1999).
Menurut Octavitani (2010), banyak faktor biotik dan abiotik yaang menentukan perkembangan FMA. Faktor-faktor tersebut antar lain suhu, tanah, kadar air tanah, pH, bahan organik tanah, intensitas cahaya dan ketersediaan hara, serta logam berat dan fungisida.
Hasil Penelitian Keanekaragaman FMA pada Lahan Gambut
Keanekaragaman FMA telah diteliti di beberapa lahan gambut di Indonesia. Keanekaragaman FMA di Kecamatan Pollung Kabupaten Humbang Hasundutan tipe spora yang ditemukan adalah jenis Glomus spp. dengan jumlah 15 jenis dan jenis Acaulospora spp. sebanyak 4 jenis (Lumban Gaol, 2007). Pada lahan gambut di Kalimantan Barat dengan ekosistem alami dan ekosistem yang terganggu atau diolah, spesies yang ditemukan adalah Glomus, Acaulospora,
lokasi penelitian sedangkan Acaulospora lebih produktif pada lokasi yang terganggu (Ekamawanti, 1997).
Sementara itu Ervayenri et al. (1997), melaporkan dalam penelitiannya bahwa kelimpahan spora tertinggi diperoleh pada tanah dengan vegetasi pertanian dan vegetasi produksi kehutanan dibandingkan dengan lahan yang ditumbuhi vegetasi alami (hutan alam). Keadaan ini diduga disebabkan karena adanya gangguan struktur tanah terhadap pola olah tanah yang dilakukan, sedangkan pada hutan alam gambut, sama sekali tidak ada dilakukan pola olah tanah yang dapat menyebabkan kerusakan pada struktur tanah.
Hasbi (2005), melaporkan pada gambut fisiografi datar dengan ketinggian permukaan tanah berkisar 1 – 1,5 meter di atas permukaan laut, pada tingkat kematangan sapric, memiliki pH berkisar 2,85 keberadaan fosfor sebesar 27,19 ppm. Hasil identifikasi spora menunjukkan bahwa terdapat dua kelompok cendawan yang secara morfologis diklasifikasikan ke dalam genus Glomus dan
Acaulospora. Dari kedua genus tersebut umumnya genus Glomus yang paling
dominan dijumpai yang berasosiasi dengan tanaman berturut-turut dari yang tertinggi yaitu nenas, sawi, pepaya, kangkung dan terong.
Variasi jumlah spora yang diamati di lahan gambut Pontianak, dengan tanaman budidaya yang dijadikan sebagai tanaman sampel, diduga akibat kondisi tanaman dan lingkungan tumbuh serta dapat pula disebabkan oleh faktor kemampuan infeksi dari CMA menyangkut populasi spora yang berkembang. Perbedaan lokasi dan ke dalaman rizosfir juga menyebabkan perbedaan genus yang ditemui. Genus Glomus dijumpai hampir pada semua lokasi dan tanaman sampel yaitu nenas, sawi, pepaya, kangkung dan terong kecuali bayam. Genus
Acaulospora hanya ditemukan pada tanaman sawi, pepaya dan kangkung. Hal ini
menunjukkan bahwa genus Glomus mempunyai kemampuan adaptasi dengan jenis tanaman budidaya yang lebih luas jika dibandingkan dengan genus