Omikusu kaiseki o mochiita bunshi kuraudingu koka, oyobi gan yakuzai taisei eno tankyu
Bebas
8
0
0
Teks penuh
(2) オミクス解析を用いた分子クラウデイング効果,及びがん薬剤耐性への探究. 環境情報学部4 年 山久保純. 要旨 本報告では, システムバイオロジー的概念に基づくオミクス解析から生命現象へのアプローチ を 利 用 し た 研 究 に ついて学 部 中 に 行 っ た テーマであ る 「 分子クラウデイング効果が大腸菌の中心炭 素代謝反応に与える影響」, 「 卵巣がんにおける薬剤耐性機構の解明に向けたプロテオーム及び リ ン 酸 化 プ ロ テ オ ー ム 解 析 」 の2 つ の 内 , 前 者 の み を 述 べ る . 細胞内の分子混雑状態が生体分子のふるまいに影響を与える分子クラウデイング効果の一つとし て, 酵 素 に お け る 熱 安 定 性 の 向 上 が 明 ら か に さ れ て い る . 分 子 混 雑 状 態 で は , い く つ か の 酵 素 に. おいて,熱 に 対 す る 恒 常 性 が 増 加 す る と 先 行 研 究 で 報 告 さ れ ているが , こ れ ら は 単 独 の 酵 素 反 応 のみであり,代謝反応のような連続反応においては不明である.. そ こ で 本 研 究 で は in v it r o にて擬. 似的に再現した分子混雑状態での代謝反応をメタボローム解析から調べることで,分子クラウデ イングが与える代謝反応への影響を明らかにすることを目指した. 結 果 , ペントースリン酸経路の _ 速 酵 素 で あ る G lu co se 6-phosphate dehydrogenase. (G 6 P D H )が分子クラウデイング効果によって,. 高温状態下でも活性を保ち, ペントースリン酸経路上流の反応を維持させることが示唆された. キ ー ワ ー ド :分子クラウ デ イ ン グ ; メ タ ボ ロ ミ ク ス , 癌 , 薬 剤 耐 性 , プ ロ テ オ ミ ク ス. 143.
(3) 1•序論 1.1分子クラウデイング 細 胞 内 は 多 く の 生 体 分 子 の 混 在 に よ っ て 非 常 に 高 濃 度 と な っ て い る .こ の よ う な 細 胞 内 環 境 は 高 濃 度 と い う よ り も , む し ろ 混 雑 し て い る と 表 現 さ れ る ( E llis W a/.,2 0 0 1 ) . 細 胞 内 は 非 常 に 特 殊 な 環 境を有しており,分子混雑状態の他にも,細胞内領域における局在化や,細胞ごとに存在する分 子 や ふ る ま い が 異 な る と い っ た 不 均 一 性 な ど が 観 察 さ れ る .そ し て こ れ ら が 複 雑 に 作 用 す る 細 胞 内 環 境 は 細 胞 内 に 含 ま れ る 生 体 分 子 に い く ら か 制 約 を 与 え て い る と さ れ て い る .そ の 中 で も 分 子 混雑状態が分子のふるまいに影響を与える分子クラウデイングは分子動力学に基づくシミュレーシ ヨンと数々の生化学実験から,生体反応においてその効果が実証されている( M i n t o n が 乂 ,2 0 01 ). 分子クラウデイングは込み合いによる拡散速度の低下や占有面積の減少による排除体積効果から 平 衡 定 数 を 変 化 さ せ , 酵素活性 を 増 加 さ せ る こ と や , タンパク質の折りたたみ,熱安定性を向上さ せることが知られている( E l i s 以 a/., 2 0 0 1 ) . こ れ ら 分 子 ク ラ ウ デ イ ン グ の 効 果 が , D N A ポリメラー ゼ の D N A 結 合 反 応 に 寄 与 す る こ と ( Z im m e rm a n (Stefan e以 /., 2 0 1 3 ) も 報 告 さ れ て い る .. d. W . , 1 9 8 7 ) や, 細 胞 増 殖 の 制 御 に 関 わ る こ と. しかし, 生 化 学 実 験 の 多 く は 分 子 混 雑 状 態 と は ほ ど 遠 い 低. 濃 度 の 緩 衝 溶 液 内 で 行 わ れ る た め , 例 え ば 大 腸 菌 の 細 胞 内 溶 液 で は , タ ン パ ク 質 , R N A などによ って 300〜 400 g / L と 高 濃 度 に な っ て い る の に 対 し て , 一 般 的 な in v itro 実 験 で 使 用 さ れ る 溶 液 の 多 く が 1〜1 0 g / L 程 度 と 非 常 に 低 濃 度 と な っ て い る ( E lis W a / .,2001) . 混 雑 し た 細 胞 内 に お い て は , 生 体 分 子 が 複 雑 な 相 互 作 用 を す る こ と で は じ め て そ の 機 能 を 発 揮 す る ケ ー ス も あ る と 想 定 さ れ , 希釈さ れ た 溶 液 内 で は 完 全 に 再 現 で き な い 生 体 反 応 が あ る と 考 え ら れ る . 分 子 ク ラ ウ デ イ ン グ の 効 果 を in v itro 実 験 に て 調 べ た 生 化 学 実 験 の ほ と ん ど が , 反 応 液 に ク ラ ウ デ イ ン グ 剤 と 呼 ば れ る 不 活 性 高 分 子. を添加することによって,細胞内環境の一つである分子混雑を擬似的に再現している.. しかし, 高. 濃 度 に 含 ま れ る 不 活 性 高 分 子 が 生 体 反 応 に 特 異 的 な 相 互 作 用 を 及 ぼ し ,混 み 合 い 以 外 の 効 果 を 引 き 起こすことが懸念されるなど,細胞内の分子クラウデイングを適切に再現できていないという懐疑 的な意見も已然として存在する( P h i l l i p が <3/., 2 0 1 3 ) . また B a r r y ら は 使 用 す る ク ラ ウ デ イ ン グ 剤 の 形 状 に よ っ て 得 ら れ る 結 果 が異なることを報告しており,球 状 に 比 べ て , ポリマー状のクラウデイン グ剤は粘度が高くなりやすいことから,拡散速度の低下によって酵素活性が下がると述べている ( B a r r y 以 《 /., 2 0 0 6 ) . し か し ク ラ ウ デ イ ン グ 剤 を 用 い た 実 験 に よ る 報 告 は 多 く ,例 え ば N o r r is らは in v itro 実 験 か ら ペ ン ト ー ス リ ン 酸 経 路 の G 6 P D H に お け る 酵 素 反 応 が , 分 子 ク ラ ウ デ イ ン グ に よ っ て 酵. 素 活 性 に 影 響 を 受 け て い る こ と を 報 告 し て お り ( N o rris er a/., 2 011 ), 特 に 反 応 溶 液 を 高 温 度 に し た 異 環 境 下 で は そ の 効 果 を 顕 著 に 観 察 で き る な ど 興 味 深 い 結 果 を 述 べ て い る .そ の 他 い く つ か の 酵 素反応機構においても同様の実験によって分子クラウデイングが酵素反応に影響を与えていること はいくつも報告されている( M o ra n -Z o rz a n o W a /., 2 0 0 7; V o p e l 以 《 /., 2 0 1 2 ).. 1.2 目的 in v itro 実 験 に お い て , ク ラ ウ デ イ ン グ 剤 を 用 い た 実 験 か ら 分 子 ク ラ ウ デ イ ン グ が 酵 素 活 性 に 影 響. を及ぼすことがこれまで報告されてきた( T o t a n id < 2 0 0 8 ).. In v itr o 実 験 系 で の 溶 液 濃 度 を 細 胞 内. に近づけることで,影響を受けるとされた酵素反応は,細胞内での活性により近いと想定されて いる.. しか し , 細 胞 内 の 酵 素 反 応 は 複 数 の 反 応 が 連 続 し て 起 こ る よ う な 代 謝 の 一 部 で あ り , それ. ぞ れ の 代 謝 経 路 に お い て の 反 応 速 度 を 決 め る の は , 反 応 速 度 の 遅 い 律 速 酵 素 で あ る .こ の 律 速 酵 素が,代謝経路の反応速度を決めているため, それ以外の酵素における反応速度は律速酵素と同. 144.
(4) 程 度となり,例え律速酵素以外の酵素活性が増減しても経路全体に影響を及ぼすことは少ないと 考 え ら れ る .こ れ ま で の 先 行 研 究 の 多 く は 単 独 の 酵 素 反 応 に お け る 影 響 の み を 報 告 し て い る も の が多く,代謝経路全体において分子クラウデイングがどのような影響を及ぼしているのかは明らか に な っ て い な い .そ し て 分 子 ク ラ ウ デ イ ン グ が 及 ぼ す 影 響 を シ ス テ ム と し て 理 解 す る に は , 一つ の 酵 素 だ け で は な く ,連 続 す る 酵 素 反 応 に よ る 代 謝 経 路 の 現 象 を 網 羅 的 に 俯 瞰 す る 必 要 が あ る .以 上 のことから本研究では,分子クラウデイングが代謝反応にどのような影響を与えるのか明らかにす ることを目的とした.. また分子クラウデイングはタンパク質の正しい折りたたみや,熱安定性に寄. 与 す る と さ れ , 上 述 し た N o r r is ら の 研 究 に お い て も 分 子 ク ラ ウ デ イ ン グ は , 酵 素 が 失 活 す る よ う な 高 温 状 態 に お い て も 活 性 を 安 定 に 保 つ こ と が で き る と 報 告 し て い る (N o rris W a / . ,2011 ) . このことか ら 代 謝 反 応 で も 様 々 な 摂 動 に 対 し て , 同 じ よ う な こ と が 起 こ る と 考 え た . つ ま り 熱 な ど ,タンパク 質の構造を変化させ,生体反応を妨げるような摂動に対して,分子クラウデイングがロバスト性に 寄与するという仮説を立てた. 本研究では分子クラウデイング状態での代謝反応をメ夕ボローム解析によって観察するための実 験系を構築し,分子クラウデイングの効果を観測できることを確かめた.. そ の後, in v it r o にて再構. 成 さ れ た 代 謝 反 応 系 を 分 子 ク ラ ウ デ イ ン グ 状 態 で 行 い , 分 子 ク ラ ウ デ イ ン グ の 効 果 に つ い て 調べ た .本 報 告 で は 構 築 さ れ た 実 験 系 を 用 い た 分 子 ク ラ ウ デ イ ン グ 状 態 に お け る メ 夕 ボ ロ ー ム 解 析 に ついて述べる.. 2 対象と手法 2 .1 大 腸 菌 由 来 酵 素 群 の 抽 出. クラウディング状態における代謝解析を行うために,使用する酵素群を大腸菌から抽出した. 菌 株 は B W 25113を 用 い て 培 養 , 抽 出 を 行 っ た . L B 寒 天 培 地 に 対 象 の 大 腸 菌 を 植 菌 , 37°Cで ー 晚 培 養 し た 後 ,増 殖 し た コ ロ ニ ー を l% c a s a m in o a c id 入 り の M 9 液 体 合 成 培 地 に て 植 菌 , 3 7 ° C で 前 培 養 を 行 っ た • O D 6〇 〇 = 0 .3 6 で 大 腸 菌 培 養 液 を M 9 液 体 合 成 培 地 に O D 6〇 〇 = 0 .0 1 に な る よ う 植 菌 し た . O D 600= 0 .3 4 お よ び O D 600= 0 .6 8 で 培 養 液 を 遠 心 分 離 し , 菌 体 を 回 収 し た .回 収 し た 菌 は そ れ ぞ れ M O P S B u ffe r で 懸 濁 後 , 超 音 波 破 砕 機 に て 菌 体 を 破 壊 , 遠 心 分 離 後 の 上 清 に I m M P M S F を 加 え た .. 溶液を濾過処理( 1 0 K 限 外 濾 過 フ ィ ル タ ー )に て 濃 縮 後 , M O P S B u ffe r を 添 加 し , さ ら に 濃 縮 し た . 液 量 の 半 量 の 100 % グ リ セ ロ ー ル 溶 液 を 添 加 し , こ れ ら 異 な る 増 殖 相 の 菌 体 か ら 得 た 抽 出 液 を 混 合 した . 得 ら れ た 大 腸 菌 由 来 抽 出 酵 素 群 は B ra d fo rd 法 に て 定 量 を 行 っ た . 2 .2 分 子 混 雑 条 件 下 で の メ 夕 ボ ロ ー ム 解 析. 大腸菌由来酵素群を用いた代謝反応をクラウディング状態で行い, キヤピラリー電気泳動時間 飛 行 型 質 量 分 析 (C E -T O F M S ) による中間代謝物の一斉測定から分子クラウディングが代謝反応に 与 え る 影 響 を 調 べ た . 既 に 大 腸 菌 由 来 抽 出 酵 素 群 を 用 い た 代 謝 反 応 を in v it r o に て 行 い , 代 謝 産 物 をC E - T O F M S に て 測 定 す る 実 験 系 は 慶 應 義 塾 大 学 政 策 メ デ ィ ア 研 究 科 斎 藤 菜 摘 講 師 に よ っ て 構 築 され て い る .. こ の た め 本 実 験 で は 構 築 さ れ た 実 験 系 を 応 用 し た . 反 応 溶 液 ( 500 p M A T P , 500. N A D , 500 p M N A D P , 500 M A D P , M O P S buffer ( p H 7 . 2 ) , 100 g /m L 大 腸 菌 由 来 抽 出 酵 素 群 , M i l l i - Q ) を. 調 製 し , ク ラ ウ デ イ ン グ 剤 と し て P olyethylene g ly c o l 6000 ( P E G 6 0 00 ) を 1 0 % w / v で 添 加 , 及 び 未 添 加 の 条 件 で そ れ ぞ れ 実 験 を 行 っ た . 反 応 溶 液 は 37°C , 50 °C で 3 0 m i n の プ レ イ ン キ ュ ベ ー シ ョ ン を 行 つ た.. 1 m M G lu co se 6-phosDhate ( G 6 P ) を 添 加 す る こ と で 酵 素 反 応 を 開 始 , 反 応 開 始 後 1 0 ,2 0 , 30 m in. 145.
(5) で そ れ ぞ れ サ ン プ リ ン グ を 行 っ た 後 ,C E _ T O F M S 測 定 用 の 内 部 標 準 化 合 物 を 含 む 水 溶 液 に て 反 応 溶 液 を 5倍 に 希 釈 , 限 外 濾 過 処 理 ( 1 0 K 限 外 濾 過 フ イ ル タ ー )を 行 っ た .作 成 し た 試 料 は 陰 イ オ ン モ 一ドにて測定を行った. 一連の反応は解糖系とペントースリン酸経路のみでの反応を想定した.. 3 結果と議論 3.1 分子クラウディング効果による代謝反応への影響 分子クラウディング状態におけるメタボローム解析の結果を代謝経路上にマッピングした( 図 1 )• まず, ど の 反 応 条 件 に お い て も 3 -P h o s p h a g ly ce rate P hosphoenolpyruvate (P E P ),. (3 P G ),. P yru vate ( P Y R ) は 観 測 で き て い な い .. 2 -P h o s p h a g ly c e r a te. (2 P G ),. これは P Y R か ら 下 流 の 経 路 に 代. 謝 物 が 流 れ て し ま っ て い る こ と や , G ly c e ra ld e h y d e 3-phosphate (G 3 P ), D ih y d ro x y a c e to n e phosphate ( D H A P ) 間の酵素であるT P Iが反応を行えていない可能性が考えられる.. さらに測定前に,反応液. を5倍 に 希 釈 し た た め に 低 濃 度 の 代 謝 物 が 検 出 で き て い な い 可 能 性 も あ る . G 3 P よ り 下 流 の 代 謝 物 が観測されないことの決定的な理由は分かっていない. 37て イ ン キ ュ ベ ー シ ョ ン 後 の 代 謝 反 応 で は ク ラ ウ デ ィ ン グ 剤 を 含 む こ と に よ る 変 化 は み ら れ な か. った( 青 線 ) • これ ま で G lu co se 6-phosphate dehydrogenase ( G 6 P D H ) 酵 素 反 応 実 験 で み ら れ た よ う な 分 子 ク ラ ウ デ ィ ン グ 効 果 が 代 謝 経 路 に お い て も 確 認 で き る と 予 想 し て い た が , 変イ匕はみられなかっ た.. これは連続反応による代謝反応は活性測定試験のように最大速度での反応を行っておらず;基. 質の量によって反応速度が制限されるため,G 6 P D H のような律速酵素においても単独酵素反応で みられるような分子クラウディングの効果は観察できず; 代謝反応ではその影響を受けにくいので はと考える. 次 に 5 0 °C の プ レ イ ン キ ュ ベ ー シ ョ ン 後 の 代 謝 反 応 で は P E G 6 0 0 0 を 含 ま な い 場 合 , 3 7 て の プ レ イ ン キ ュ ベ ー シ ョ ン と 比 べ て G 6 P の 消 費 速 度 が 低 下 し て い る 他 , D H A P , 6 -P h o s p h o g lu c o n a te (6 P G ), R ib u lose 5 -p h o s p h a te (R u 5P ),. R ibo se 5_ p h o p h a te (R 5P ) の 量 が 低 下 し て い る ( 赤 線 , 実 線 ) .こ れ は 高. 温度の状態が各酵素の活性を低下させたためであると考えられる.. しかし, 1 0 % w / v の P E G 6 0 0 0 を. 含 む 場 合 に お い て 6 P G ,R u 5 P の 濃 度 は 37°C プ レ イ ン キ ュ ベ ー シ ョ ン と 同 じ よ う な 挙 動 を 示 し て お り, ペ ン ト ー ス リ ン 酸 経 路 上 流 に 関 し て 代 謝 反 応 が 高 温 状 態 で も 維 持 さ れ て い る こ と が 観 察 さ れ た ( 赤 色 , 点 線 ) . これは N o r r is ら の 先 行 研 究 で 示 し た よ う に , G 6 P D H が 分 子 ク ラ ウ デ ィ ン グ の 効 果 に. よ っ て 熱 に 対 し て も 活 性 を 保 つ こ と が で き た た め で あ る と 考 え る . N o r r is ら は 酵 母 由 来 の G 6 P D H を 使用しているため,大腸菌由来のG 6 P D H においても同様に分子クラウディング効果を単独の酵素 ア ッ セ イ か ら 検 証 し た と こ ろ 同 様 の 結 果 を 得 る こ と が で き た ( 付 録 図 S 1 ) . ま た 同 様 に R u 5 P も3 7 て プレインキュベーションの場合と同じ挙動を示していることから,6PG, Ru5P間の反応における酵 素 ( 6 P G D H ) に お い て も 分 子 ク ラ ウ デ ィ ン グ の 影 響 を 受 け て い る こ と が 予 想 さ れ た . 6 P G D H におい て も 分 子 ク ラ ウ デ ィ ン グ 効 果 を 調 べ た が , G 6 P D H の よ う な 効 果 は 観 察 さ れ な か っ た こ と か ら (付 録 図 S 2 ), 律 速 酵 素 で あ る G 6 P D H の 酵 素 活 性 が 高 温 状 態 に お い て も 安 定 し て 反 応 を 行 え た こ と で 後 の Ru5Pの生産にも影響したと考えられる.. 当 実 験 に お い て P E G 6 0 00 を 1 0 % w /v含 む 溶 液 内 で 行 わ れ た 代 謝 反 応 に ペ ン ト ー ス リ ン 酸 経 路 上 流 に お け る 高 温 状 態 に お け る 代 謝 反 応 の 恒 常 性 が み ら れ た .こ れ は 分 子 ク ラ ウ デ イ ン グ の 効 果 に よ る も の で は な く , P E G そ の も の が 持 つ 性 質 で あ る 可 能 性 も あ る が , 不 活 性 分 子 で あ る P E G がもたら す効果が実際の細胞内で起こっていることも十分考えられ. 146.
(6) Q IC. 細細綱綱糊. PE660CD 0% , 37X. Im. づト PEG60®3 0%, SCTC 诚. P K 6000 i 〇 A 37X. G6PDH. PEG60CK) 10%, SOX H嫌{_約 }. I. mmn. pgk. 1,3-BP6 — ^ 3P 6/2P 6. m. m. 図 1 分子クラウデイング状態における時系列変動代謝. (min) , 縦 軸 は 濃 度 〇iM) を 表 し て い る . そ れ ぞ れ の 条 件 に お い て 時 系 列 ご と に 測 定 し た 物 質 を 代 謝 経 路 上 に マ ッ ピ ン グ し た . 青 色 . 実 線 :3ア C プ レ イ ン キ ュ ベ ー シ ョ ン , ク ラ ウ デ イ ン グ 剤 を 含 ま な い . 青 色 • 点 線 :37°Cプ レ イ ン キ ュ ベ ー シ ョ ン , ク ラ ウ デ イ ン グ 剤 を 含 む 赤 色 • 実 線 • 50°Cプ レ イ ン キ ュ ベ ー シ ョ ン , ク ラ ウ デ イ ン グ 剤 を 含 ま な い . 赤 色 • 点 線 :50て プ レ イ ン キ ュ ベ ー シ ョ ン , ク ラ ウ デ イ ン グ 剤 を 含 む . 横軸は反応時間. 4 結論 本 研 究 で は , 分 子 ク ラ ウ デ イ ン グ が 代 謝 反 応 に 与 え る 影 響 を 明 ら か に す る た め , in v itr o に再構成 し た 代 謝 反 応 を 高 濃 度 の 不 活 性 高 分 子 中 で 行 い ,C E -T O F M S に よ る 測 定 を 行うことで分子クラウ デイングの効果を観察する実験系を構築した.. そ し て メ 夕 ボ ロ ー ム 解 析 の 結 果 , 1 0 % w /v P E G 溶 液. で は 3 7 T プ レ イ ン キ ュ ベ ー シ ョ ン に お い て 代 謝 反 応 に 変 化 が な く , 連 続 反 応 の 代 謝 に お い て 37°C の 至 適 温 度 で は 分 子 ク ラ ウ デ イ ン グ の 影 響 は な い こ と が 示 唆 さ れ た . 50°Cプレインキュベーション で は 1 0 % w/ v P E G を 含 む こ と で , ペ ン ト ー ス リ ン 酸 経 路 上 流 に お い て 代 謝 反 応 は 低 下 せ ず ; 維持さ れ る な ど , 分 子 ク ラ ウ デ イ ン グ と 思 わ れ る 効 果 が 観 察 さ れ た .今 回 の 実 験 で こ の よ う な 現 象 を 観 察. 147.
(7) できたのはPEGの み で あ る た め , 混 雑 状 態 に よ る も の で は な く ,PEGがもつ性質である可能性も考 え ら れ る .ま た メ 夕 ボ ロ ー ム 解 析 を 用 い て 分 子 ク ラ ウ デ イ ン グ の 効 果 を 観 察 で き る 実 験 系 を 構 築 し たことで,今回実験の対象とした解糖系,ペントースリン酸経路以外の代謝反応経路においても同 様に実験を行い,分子クラウデイングによって代謝反応に影響を及ぼすような酵素の探索をするこ と が で き る .ま た 今 回 は 代 謝 反 応 を 阻 害 す る 例 と し て 熱 を 与 え た が , 上 述 に お け る 仮 定 を 証 明 す るためにはpHの 変 化 な ど 熱 以 外 に も 様 々 な 摂 動 に 対 する分子クラウデイングの影響を観察するこ とは興味深い. ま た 後 日 談 で は あ る が , こ の 研 究 が 終 了 し た 後 の 2 0 1 3 年 7 月 に Cheemeng ら は 構 築 し た 人 口 細 胞 に おいて分子クラウデイング環境を再現し,D. NAポリメラーゼ反応に様々な摂動を与えたところ,. 摂動に対して混雑状態がロバスト性に寄与することを報告している( C heemeng 以 a /.,2 0 1 3 ) . つ ま り 本研究で示そうとした代謝反応においては,分子クラウデイングがあらゆる摂動に対してロバスト 性に寄与するという仮説を,彼らは. D N A ポ リ メ ラ ー ゼ 反 応 か ら そ の 一 端 を 証 明 し た .本 研 究 で は. これを代謝反応において同様の効果がみられることを示せればと思う.. 謝辞 卒業プロジェクトにおける研究は,学部中のアドバイザーである佐伯憲和氏,斎藤菜摘氏,森 大 氏 の 多 大 な る サ ポ ー ト に よ っ て 行 わ れ て い る .ま た 所 属 し た. E -Cellグ ル ー プ , プ ロ テ オ ー ム グ ル. ー プ の 方 々 に も 様 々 な ア ド バ イ ス を 頂 い た .山 岸 広 太 郎 氏 を は じ め と す る 山 岸 学 生 プ ロ ジ ェ ク ト の 方 々 に は 研 究 費 の 援 助 を し て 頂 い た .こ れ ら の 方 々 に は こ の 場 を お 借 り し て 感 謝 の 意 を 示 し た い . 最後にすばらしい研究環境を与えてくださった冨田勝氏には深く感謝したい.. 参考文献 Derham , B . K . & Harding, J. J. The effect of the presence oi globular proteins and elongated polymers on enzvme activity. Biochim. Biophys.Acta 1764, 1000-6 (2006). Ellis, R . J. Macromolecular crowding: obvious but underappreciated. Trends Biochem. Sci. 26, 5 9 7 - 6 0 4 (2001). Minton, a P. The influence oi macromolecular crowding and macromolecular cominement on biochemical reactions in physiological media. J. Biol. C h e m . 276, 1 0 5 7 7 - 8 0 (2001). Mordn -Zorzano, M . T . et a l . Escherichia coli AspP activity is enhanced by macromolecular crowding and by both glucose-1,6-bisphosphate and nucleotide-sugars. FEBS Lett. 581, 1 0 3 5 -4 0 (2007). Phillip, Y . & Schreiber, G . Formation of protein complexes in crowded environments-from in vitro to in vivo. FEBS Lett. 587, 1 046-52 (2013). Tan, C ., Saurabh, S ., Bruchez, M . P., Schwartz, R . & Leduc, P. Molecular crowding shapes gene expression in synthetic cellular nanosystems. Nat. Nanotechnol. 8, 6 0 2 - 8 (2013). Totani, K ., Ihara,Y ., Matsuo, I. & Ito,Y . Effects of macromolecular crowding on glycoprotein processing enzymes. J.A m . C h e m . Soc. 130, 2 1 0 1 - 7 (2008). V 6 pel, T . & Makhatadze, G . I. Enzyme activity in the crowded milieu. PLoS One 7, e3 9 4 1 8 (2012). Zimmerman , S . B . & Harrison, B . Macromolecular crowding increases binding of D N A polymerase to D N A : an adaptive effect.Proc . N atl . A cad . Sci . U . S . A . 8 4 , 1 8 7 1 - 5 (1987).. 148.
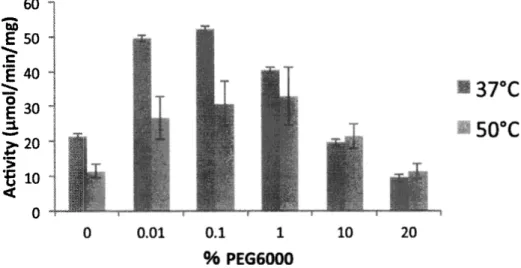
(8) ■ { 3 E / u i E / l o}l ^ y > l p <. n. 60 50 40 30 20 1 0 = L. 1 図 S I G 6 P D H 活性が受ける分子クラウデイングの影響. 横軸は各クラウディング剤の濃度 ( W. / V ) . 左の図の縦軸は G 6P D H の酵素 活 性 ( nmol/min/mg ),青色が3アC ,赤色が50てでプ. レインキュベーションした場合における活性をそれぞれ表している.誤差範囲は3回の測定結果における標準誤差. (Standard Error;S.E .)をボす.. 100 0 8 0 6 0 4. 3 {3E/UIIU/IOE > >. ♦ *. 0 2. € <. 0. 図 S2 6 P G D H 活性が受ける分子クラウディンクの影響 横軸は各クラウディング剤の濃度( w /v) . 左の図の縦軸は6P G D H の酵素活性卬 mol/min/mg ),黒が3ァC ,灰色が50てでプ レインキュベーションした場合における活性をそれぞれ表している.誤差範囲は3 回の測定結果における標準誤差. (Standard Error;S.E .)を不す.. 149.
(9)
Gambar
Dokumen terkait
Taman Nasional Kepulauan Seribu mempunyai sumber daya alam yang khas yaitu keindahan alam laut dengan ekosistem karang yang unik seperti terumbu karang, ikan
Salah satu nematoda parasit tanaman yang menyebabkan kerugian pada beberapa tanaman khususnya tanaman hortikultura seperti seledri dan tomat adalah Nematoda Puru Akar
Berbeda dengan nelayan kabupaten Cilacap, nelayan di PPSC biasa menggolongkan musim penangkapan menjadi tiga yaitu: musim puncak terjadi pada bulan Juli sampai dengan bulan
Adapun tujuan dari Penelitian ini adalah: (1) mengidentifikasi terjemahan Terminologi Keimigrasian ke dalam Bahasa Inggris dalam Undang-Undang Nomor 6 Tahun 2011 tentang
Informasi yang diperoleh melalui FGD tersebut menjelaskan hasil yang diperoleh melalui angket, dimana mayoritas responden siswa (87,6%) memandang bahwa peran yang
Prinsip kerjanya mencari beda fase dari echo signal yang diterima oleh dua belahan transducer (port- starboard phase pulse, dan fore-aft phase pulse), selain dapat mengukur in
Karena efeksamping menyebar melalui interface program, maka penting untuk mengecek semua aliran data dari modul ke modul (atau ke dunia luar) untuk memastikan
Diagram disamping menunjukan bahwa kondisi MR= MC titik E tercapai pada saat out put Q*,karena AC < P,dari output perusahaan laba sebesar BE per unit ,jika output lebih kecil
