Oseana, Volume XIV, Nomor 1 : 27–36, 1989. ISSN 0216–1877
FORAMINIFERA oleh
Ricky Rositasari 1) ABSTRACT
FORAMINIFERA. As unicelular animals, foraminifera have specific charac- teristics such as the capability to build a hard protective shell which is secreted by the animal and to adapt in the various kind of environments. This group of animals has high species diversity and widely distributed both geographically and geologi- cally. These animals can be used as indicator in many aspects. Recent foraminifera are used as indicators in many oseanographic aspects such as chemical, physical and geological and their fossils can also be used as paleoecologic indicators.
PENDAHULUAN
Salah satu keunikan dari foraminifera adalah kemampuannya untuk melangsung-kan proses fisiologis dalam sek tunggalnya yang memiliki ukuran 1 µm - 2 mm sesem-purna proses fisiologis pada organisme ber-tulang belakang kecuali pada organel per-kembangbiakannya. Keistimewaan lainnya adalah kemampuannya untuk membentuk arsitektur serta komposisi kimia yang rumit pada pembentukan cangkangnya sebagai ha- sil sekresi sel. ANDERSON & ALLAN (1978) menyebutkan bahwa kemampuan un-tuk membenun-tuk arsitektur serta menyusun komposisi cangkang yang berbeda pada se-tiap jenis, dikontrol oleh kode genetika yang mempengaruhi proses fisiologisnya.
Alasan para ahli perminyakan pada awal perang dunia pertama menggunakan
analisis foraminifera dalam kegiatan eks-plorasi adalah karena foraminifera memiliki bagian tubuh yang keras dengan ukurannya yang kecil, sehingga dengan mudah terawet-kan di dalam sedimen. Selain itu sebaran geograflsnya yang luas serta kisaran umurnya yang panjang memungkinkan foraminifera untuk merekam informasi yang sangat ber-harga tentang sejarah bumi, sehingga hanya dengan menggunakan sejumlah kecil contoh para ahli sudah dapat menganalisis dan meng-interpretasi korelasi regional antara satu tempat dengan tempat lainnya, serta sifat oseanografi dan ekologi pada masa lalu. Di samping itu hasil penelitian terakhir me-nunjukkan bahwa foraminifera dapat juga di-pakai sebagai indikator untuk mengetahui keadaan suatu lingkungan yang sedang meng-alami tekanan.
BIOLOGI FORAMINIFERA Sistematika
Dalam sistematik tata nama, forami-nifera termasuk dalam filum Protozoa kare- na organisme ini bersel satu, dan termasuk pada kelas Sarcodina karena sistem pergerak-annya terdiri dari kaki semu (pseudopodia).
Ordo foraminifera terbagi lagi menjadi 5 anak ordo yaitu Allogramiina, Textulariina, Fusulinina, Miliolina dan Rotaliina. Menurut HAQ & BOERSMA (1984) pembagian anak ordo tersebut didasarkan pada :
1. Mikrostruktur dan komposisi dinding cangkang.
2. Susunan kamar dan dinding tambahan. 3. Bentuk dan modifikasi mulut (apertura). 4. Bentuk kamar.
5. Kebiasaan dan tempat hidup. 6. Karakteristik protoplasma. 7. Perubahan ontogeni. 8. Proses perkembangbiakan 9. Sebaran geologi.
Anak ordo Allogramiina merupakan kelompok foraminifera yang mempunyai morfologi paling sederhana, yaitu terdiri dari 1 sel dengan bentuk dan struktur cangkang yang sederhana pula. Contoh dari anak ordo ini sangat jarang ditemukan karena cangkang nya tersusun dari bahan tektin yang mudah rusak. Sedangkan jenis-jenis yang memiliki cangkang pasiran hanya sebagian kecil saja.
Anak ordo Fusulinina merupakan fora-minifera yang berukuran relatif besar dengan susunan kamar yang sangat kompleks, se-hingga kelompok ini biasa disebut foramini-fera bentonik besar. Kisaran hidupnya dalam kala geologi hanya sampai Triasik (200 juta tahun yang lalu). Jadi pada masa resen (0– 0,23 juta tahun yang lalu) jenis-jenis ini ti-dak dapat ditemukan lagi.
Jenis-jenis dari anak ordo Textulariina memiliki kamar lebih dari satu, dindingnya ada yang terdiri dari satu lapisan atau lebih. Struktur dinding terdiri dari tektin dengan lapisan pasiran, kristal butir-butir kalsit de-ngan atau tanpa lapisan pasiran. Kamar-ka-mar tersusun dalam satu jajar, terputar atau terpilin.
Ciri morfologi pada anak ordo Milio-lina adalah jumlah kamar banyak, tersusun secara terputar atau terpilin (Gambar 1), mu-lut tunggal atau jamak dengan atau tanpa gigi. Dinding cangkang terdiri dari kristal kalsit dengan kenampakan seperti porselen atau dengan tambahan bahan pasiran.
Jenis-jenis foraminifera yang termasuk dalam anak ordo Rotaliina memiliki kamar lebih dari satu, sebagian besar memiliki din-ding yang berlapis-lapis (Gambar 1). Susun- an kamar pada sebagian besar jenisnya ter-pulir (trochospiral).
Sel dan isinya
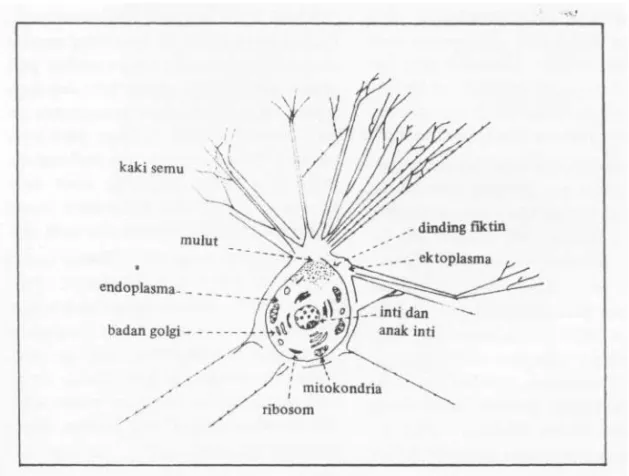
Seperti jenis-jenis Protozoa lainnya, foraminifera terdiri dari satu sel yang di-lengkapi oleh organel-organel yang berfungsi seperti organ pada organisme bertulang be-lakang. Badan golgi merupakan organel yang mengatur sekresi sel, mitokondria ber-fungsi unruk pernafasan, ribosom yang mengandung RNA merupakan tempat sin-tesa protein dan vakuola yang merupakan tempat pengaturan cairan dan gas dalam protoplasma. Gambar 2 memperlihatkan sel foraminifera hidup secara skematis. Pada umumnya sel foraminifera memiliki satu inti atau lebih dengan anak-anak inti yang berfungsi sebagai cadangan protein.
Untuk hidupnya, foraminifera mem-butuhkan nutrisi yang berasal dari alga, di-atom, krustasea, kimah, jenis mereka sen-diri dan serpihan bahan organik (MURRAY 1973). Berdasarkan jenis makanannya maka
irisan melintang Susunan Kamar Milioliina lapis bagian luar lapis rongga kamar
Gambar 1. Dinding cangkang pada anak ordo Rotaliina dan Milioliina (HAYNESS 1981).
^ kaki semu mulut endoplasma___ badan golgi
foraminifera dapat digolongkan sebagai her-bivora, karnivora maupun omnivora. Fora-minifera menangkap makanan dengan meng-gunakan kaki semu yang menjulur-julur ke segala arah melalui pori-pori atau mulut. Makanan yang tertangkap akan melekat dan langsung dicerna, sisa jasad dari mangsanya kemudian dilepas.
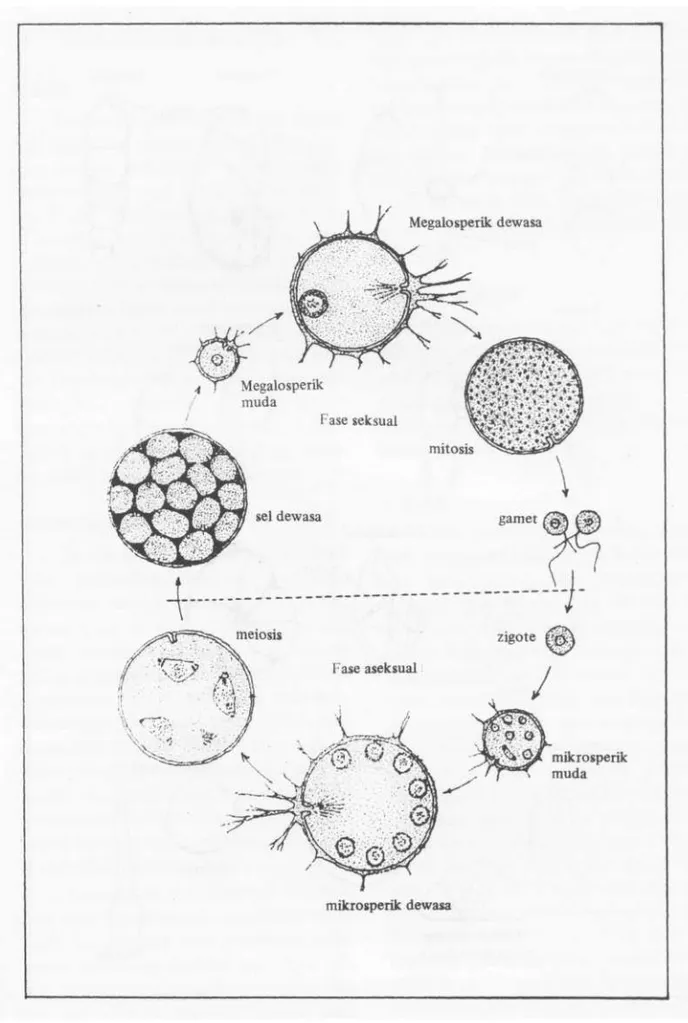
Dalam pergerakannya foraminifera bentonik menggunakan kaki semu yang ke-luar melalui mulut, sehingga dapat merayap dan bergerak turun-naik pada substratnya. Pada jenis-jenis planktonik pergerakan dila-kukan pada sepanjang kolom air, sebagai akibat perubahan kimia pada protoplasma yang menyebabkan terjadinya perubahan dalam kandungan gas di dalam sel. Fora-minifera bentonik memiliki dua fase dalam daur perkembangannya yaitu seksual dan aseksual (Gambar 3). Namun demikian be-berapa peneliti seperti HAQ & BOERSMA (1984), CUSHMAN (1955) dan ALBANY (1979) menduga perkembangbiakan yang utama pada foraminifera adalah secara asek-sual. Dugaan tersebut didasarkan pada ke-nyataan bahwa jumlah individu yang memiliki ciri morfologi megalosperik sebagai hasil perkembangbiakan aseksual sangat melimpah.
Foraminifera memiliki dua macam per-tumbuhan yaitu perper-tumbuhan periodik dan nonperiodik. Pertumbuhan nonperiodik ter-jadi pada organisme yang memiliki satu ka-mar pada cangkangnya, seperti jenis-jenis pada anak ordo Allogromiina. Sedangkan pertumbuhan periodik terjadi pada organis-me yang organis-memiliki banyak kamar pada cang-kangnya seperti jenis-jenis pada anak ordo Rotaliina, Textulariina, Miliolina dan Fusuli-nina. Pertumbuhan periodik adalah proses pertambahan ukuran cangkang sebagai aki-bat dari proses presipitasi dan akresi
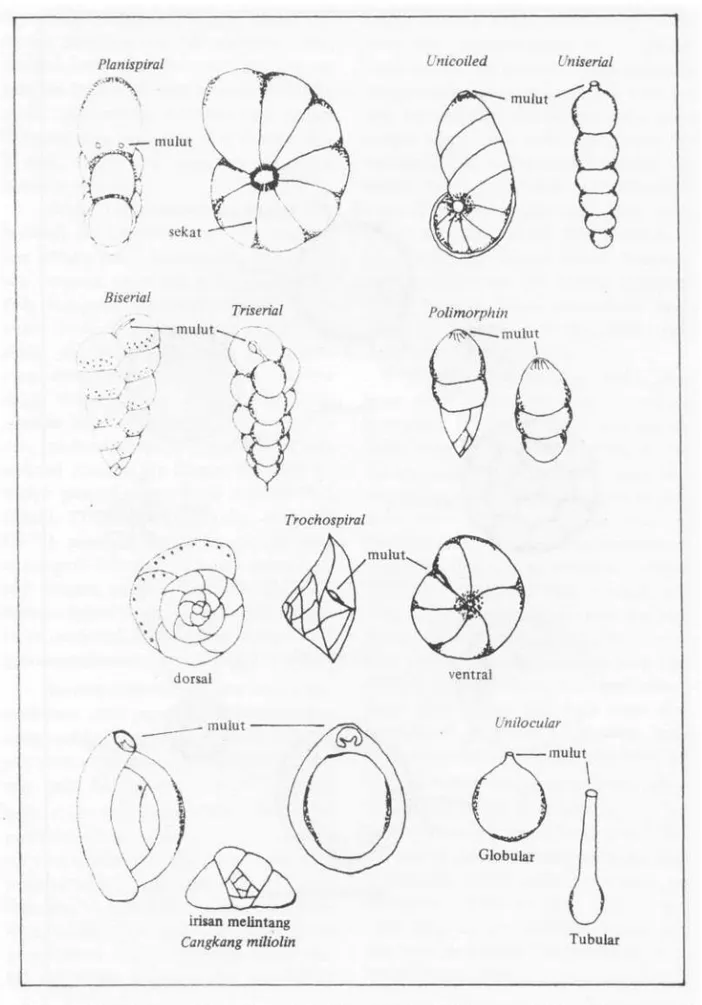
bahan-bahan mineral untuk membentuk kamar yang bam. Sebelum proses ini berlangsung foraminifera akan menyelimuti cangkangnya dengan material-material yang ada di sekitar-nya dan menghentikan seluruh aktivitassekitar-nya sampai kamar baru terbentuk. Bentuk dan susunan kamar pada foraminifera sangat ber-variasi, secara garis besar dapat dibedakan menjadi dua yaitu unilokular yang hanya terdiri dari satu kamar dan multilokular yang terdiri dari banyak kamar. Kamar-ka-mar ini dipisahkan oleh dinding tambahan. Bentuk susunan kamar multilokular dapat dibagi lagi menjadi enam tipe, seperti yang dapat dilihat pada Gambar 4.
Cangkang foraminifera secara garis besar dapat dibagi dua yaitu pasiran dan gampingan. Cangkang pasiran tersusun dari bahan-bahan yang terkandung dalam subs-tratnya seperti spikula, pasir, mika atau pecahan-pecahan cangkang organisme lain. Sedangkan cangkang gampingan terbentuk dari hasil akresi dan presipitasi protoplasma. Cangkang gampingan ini terbagi lagi menjadi dua, yaitu gampingan yang memiliki pori dengan kenampakan seperti kaca dan gam-pingan tak berpori dengan kenampakan se-perti porselen. Setiap cangkang pada fora-minifera memiliki mulut yang berfungsi se-bagai jalan keluar bagi kaki semu serta gamet-gamet pada saat berlangsung proses perkembangbiakkan. Bentuk dan letak mu-lut pada setiap cangkang berlainan seperti yang dapat dilihat pada Gambar 4. Pada beberapa marga terdapat hiasan yang berupa gigi pada mulutnya seperti pada marga Quin- queloculina. Selain mulut, gigi dan pori, komponen taksonomi lain adalah hiasan yang dapat berupa jalur-jalur, cucuk, duri, dan tonjolan-tonjolan yang tersebar secara beraturan atau tidak.
Megalosperik dewasa
mikrogperik dewasa
mikrosper ik muda
Planispi ral Unicoile d Uniseria l mulut Biserial - mulut Triserial Polimorphin - mulut i Trochospiral dorsal ventral . mulut irisan melintang Cangkang mUiolin Unilocular mulut Tubula r mulu
EKOLOGI FORAMINIFERA Sebaran
Sebaran geografis foraminifera sangat luas, mulai dari perairan tawar sampai per-airan dengan kandungan salinitas tinggi, tetapi sebagian besar dari jenis-jenisnya hi-dup di laut. Tipe lingkungan perairan terten-tu tercirikan oleh jenis-jenis yang berbeda, tergantung pada tingkat adaptasi yang di-milikinya. Hasil penelitian MURRAY (1973) menunjukkan bahwa daerah perairan pasang-surut didominasi oleh Elphidium, Quinque-loculina, Ammonia dan beberapa jenis lain-nya. Daerah terumbu karang didominasi oleh Peneroplis, Amphistegina. Calcarina dan be-berapa jenis gampingan lainnya. Daerah delta didominasi oleh Bolivina, Nonionella, Briza-lina dan beberapa jenis khusus lainnya. Lingkungan Biotik
Di dalam daur makanan mahkluk hidup, foraminifera dapat berperan sebagai konsumen tingkat pertama atau kedua, karena selain memakan diatom dan alga mereka juga memakan kimah, krustasea dan jenis-jenis mereka sendiri. Di lain pihak fora-miniferapun banyak dikonsumsi oleh ikan, ekinodermata dan jenis-jenis fauna lain yang mengambil makanan dengan cara mengisap. Selain sebagai hewan pemangsa dan yang di-mangsa, foraminifera juga banyak digunakan sebagai tuan rumah dari beberapa organisme parasit seperti bakteri, nematoda, amuba ser-ta jenis-jenis mereka sendiri.
Berdasarkan cara hidupnya foramini-fera dapat dibagi menjadi foraminiforamini-fera plank- tonik dan bentonik. Jenis planktonik hidup secara mengapung di dekat permukaan air, dan jenis bentonik hidup dengan melekatkan diri pada substrat. Jenis-jenis substrat yang biasa didiami adalah pasir, lempung, rumput laut, cangkang pelesipoda, gastropoda dan
koral. Cara hidup lain yang biasa dilakukan foraminifera adalah dengan mengadakan sim-biose bersama alga. Cangkang foraminifera berfungsi sebagai rumah kaca bagi alga, dan alga berfungsi untuk mensintesa nutrisi bagi foraminifera. Bentuk simbiose seperti ini menyebabkan beberapa foraminifera terlihat seperti berwarna hijau, coklat atau merah, sesuai dengan jenis alga yang hidup di dalam cangkangnya.
Lingkungan Abiotik
Komponen-komponen lingkungan abi-otik yang berperan dalam kehidupan fora-minifera ada yang berpengaruh secara lang-sung ada juga yang tidak langlang-sung.
Komponen abiotik yang berpengaruh secara langsung adalah :
1. Suhu
Pada umumnya foraminifera merupa-kan organisme poikilotermik (suhu tubuh-nya sama dengan suhu lingkungantubuh-nya). Beberapa jenis foraminifera memiliki tole-ransi yang besar terhadap perubahan tem-peratur lingkungan dan beberapa jenis lain tidak memiliki toleransi tersebut. Pada orga-nisme bentonik perubahan suhu siang-malam dan perubahan musim akan berpengaruh pa- da sebarannya. Perkembangbiakkan biasanya berlangsung pada temperatur optimum. BRADSHAW (dalam MURRAY 1973) telah mengamati batas toleransi suhu pada Am-monia beccarii tepida, pada suhu 10°C orga-nisme ini tidak membentuk kamar baru, suhu 24°C – 30°C merupakan suhu yang paling optimum untuk berkembang biak, sedangkan suhu 35°C merupakan suhu letal bagi jenis ini Setiap jenis foraminifera me-miliki batas toleransi yang berlainan seperti Spirilina vivipara yang mati pada suhu 39°C dan Bolivina compacta mati pada suhu 41 °C.
2. Salinitas
Sebagian besar foraminifera bentonik merupakan stenohalin atau memiliki kemam-puan yang terbatas dalam mentolelir peru-bahan salinitas seperti pada Rotaliella hete-rocaryotica yang hanya dapat bertahan pada salinitas 23,5‰ – 37,0‰. Beberapa jenis foraminifera mempunyai kemampuan adap-tasi yang besar terhadap perubahan salinitas seperti jenis Ammonia beccarii tepida yang memiliki toleransi salinitas dari 2‰ sampai 41‰, sehingga jenis ini dapat ditemukan di perairan yang bersalinitas rendah maupun tinggi. Jenis-jenis yang beradaptasi terhadap salinitas rendah, sangat jarang ditemukan pada salinitas normal karena perubahan sali-nitas akan berpengaruh pada densitas air dan tekanan osmotik pada sel foraminifera. 3. Kandungan Oksigen
LUTZE (dalam MURRAY 1973) me-nyebutkan bahwa foraminifera tidak ditemu-kan pada perairan anoksigenik seperti di cekungan dalam perairan Baltik, tetapi be-lum ada penelitian yang dapat memastikan bahwa oksigen merupakan faktor pembatas bagi foraminifera.
4. Tekanan air
BRADSHAW (dalam MURRAY 1973) telah mengadakan pengamatan terhadap da-ya tahan Ammonia beccarii tepida pada te-kanan 1000 atmosfir. Ternyata jenis ini hanya dapat bertahan dalam waktu yang sangat singkat. Ini memperlihatkan bahwa foraminifera perairan dangkal tidak dapat hidup di lingkungan abisal karena mereka telah beradaptasi dengan baik pada ling-kungan hidupnya.
Faktor abiotik yang berperan secara tidak langsung adalah:
1. Kekeruhan air
Kekeruhan biasanya dijumpai di dae-rah pantai dan sekitarnya. Kekeruhan yang tinggi akan mengurangi penetrasi cahaya dan salinitas yang merupakan salah satu faktor pembatas bagi foraminifera.
2. Intensitas cahaya matahari
Cahaya matahari tidak berpengaruh langsung terhadap kehidupan foraminifera tetapi mempengaruhi proses fotosintesa plankton dan alga yang merupakan makan-an bagi foraminifera.
3. Kedalaman air
FUNNEL (dalam MURRAY 1973) menyebutkan bahwa kemungkinan besar ke-dalaman air bukan merupakan faktor pemba-tas bagi foraminifera. Diduga yang menjadi faktor pembatas adalah beberapa parameter yang berkaitan erat dengan perubahan ke-dalaman seperti tekanan air, densitas, pene-trasi cahaya, suhu, kandungan oksigen dan karbon dioksida.
4. Zat hara dalam perairan
Jenis-jenis foraminifera yang paling di-pengaruhi oleh zat hara adalah jenis-jenis yang hidup di daerah aliran tempat pem-buangan limbah. Akan tetapi kandungan zat hara yang tinggi pada suatu perairan selalu identik dengan kandungan oksigen yang rendah, karena oksigen banyak digunakan untuk dekomposisi bahan organik. Diduga bahwa foraminifera lebih tergantung pada kandungan oksigen daripada terhadap kan-dungan zat hara. Jenis-jenis yang jumlah individunya meningkat di sekitar daerah tercemar, diduga merupakan jenis yang telah beradaptasi pada kandungan oksigen rendah.
FORAMINIFERA SEBAGAI INDIKATOR LINGKUNGAN Dari hasil analisis data foraminifera dapat dihitung beberapa koefisien yang ber-guna sebagai indikator beberapa sifat oseano- grafi, seperti nilai perbandingan jumlah indi-vidu yang hidup dengan yang mati (L/D). Semakin rendah nilai perbandingan tersebut menunjukkan semakin tingginya tingkat ke-cepatan sedimentasi, kekeruhan, kekuatan arus, serta rendahnya kandungan zat hara dan oksigen. Dan semakin tingginya nilai L/D menunjukkan terdapatnya intensitas ca-haya yang baik, kecerahan air tinggi, serta tingginya kandungan zat hara dan oksigen. Tingginya nilai perbandingan antara jumlah individu yang bercangkang pasiran dengan gampingan (A/C) menunjukkan kadar sali-nitas dan keasaman yang tingginya serta kandungan kalsium karbonat rendah. Nilai perbandingan antara jumlah individu plank-tonik dan individu benplank-tonik (P/B) yang ting- gi menunjukkan bahwa daerah tersebut merupakan perairan yang dalam dan terbu-ka, sedangkan rendahnya nilai perbandingan ini menunjukkan bahwa daerah tersebut me-rupakan perairan dangkal yang dikelilingi oleh daratan.
Indikasi lain yang diperlihatkan fora-minifera sebagai cerminan keadaan lingkung- an adalah perubahan morfologi, keragaman dan kepadatan. Pada perairan yang bersa-linitas rendah, diversitas foraminifera akan rendah dengan ukuran cangkang yang me-ngecil dan menipis. Hal ini terjadi akibat adanya penurunan sekresi kalsium karbonat dari dalam protoplasma.
Pada lingkungan yang tercemar, jenis-jenis yang beradaptasi akan memperlihat-kan peningkatan kepadatan di sekeliling zona suspensi limbah, sebaliknya di dalam zona suspensi sendiri tidak terdapat
minifera hidup. Di dalam kumpulan fora-minifera hidup kepadatan anak ordo Rota-liina delapan kali lebih besar daripada ke-padatan anak ordo Textulariina, tetapi di dalam kumpulan foraminifera yang sudah mati terjadi hal yang berlawanan. Hal ini diduga disebabkan karena terlarutnya cang-kang gampingan yang telah kosong akibat keasaman air yang tinggi. (BANDY, INGE & RESIG dalam MURRAY 1973). Perubah-an lain yPerubah-ang diperlihatkPerubah-an oleh beberapa jenis foraminifera adalah terjadinya per-kembangan cangkang yang abnormal. Pe-ningkatan kepadatan foraminifera yang sangat mencolok di sekeliling zona suspensi limbah disebabkan melimpahnya fitoplank-ton sebagai produsen yang langsung meng-gunakan zat hara, dan kemudian fitoplank-ton yang melimpah ini akan dikonsumsi oleh foraminifera. Penggunaan fosil fora-minifera sebagai indikator lingkungan purba, sepenuhnya didasarkan pada karakteristik dan perubahan yang diperlihatkan oleh organisme-organisme resen yang ada saat ini. Parameter lingkungan purba yang dapat ditentukan dengan menggunakan foramini-fera sebagai indikator adalah iklim, sifat oseanografi, geografi, suhu, umur, korelasi regional dan keadaan ekologis dari suatu perairan laut.
DAFTAR PUSTAKA
ALBANY, A.D. 1979. Recent shallow water Foraminifera from New South Wales. The Australian Marine Science Association, Australia : 54 pp.
ANDERSON, O.R. and ALLAN W.H. BE. 1978. Recent Advances in Foraminifera fine structure research. In : Foraminifera. (R.H. HEADLEY and C.G. ADAMS, Eds.) Acad. Press., London : 10–25.
CUSMAN, J.A. 1955. Foraminifera their classification and economic use. Harvard University Press., Massachusetts: 683 pp. HAQ, B.U. and A. BOERSMA. 1984.
In-troduction to marine Micropaleontology. Elsevier Biomedical., New York: 376 pp.
HAYNESS, J.R. 1981. Foraminifera. Mac- millan Publishers, LTD. London : 555 pp. MURRAY, J.W. 1973. Distribution and
ecology of living benthic Foraminifera. Crane, Russak and Co, Inc. New York: 525 pp.