Hubungan Kelainan Gen Globin dengan Penyakit
Hubungan Kelainan Gen Globin dengan Penyakit
Thalasemia
Thalasemia β
β
Ellisa
Ellisa
Fakultas Kedokteran Ukrida Fakultas Kedokteran Ukrida Jalan Te
Jalan Terusan Arjurusan Arjuna No.6, na No.6, JakartaJakarta Email:
Email: [email protected][email protected]
Pendahuluan Pendahuluan
Dengan berkembangnya teknologi di abad ke 20 ini, semakin berkembang pesat pula Dengan berkembangnya teknologi di abad ke 20 ini, semakin berkembang pesat pula kemajuan IPTEK di dunia, terutama pada bidang biologi molekuler, seperti gen. Adanya kemajuan IPTEK di dunia, terutama pada bidang biologi molekuler, seperti gen. Adanya kemajuan teknologi tersebut menjadikan proses replikasi, transkripsi, dan translasi dapat kemajuan teknologi tersebut menjadikan proses replikasi, transkripsi, dan translasi dapat disimak lebih lanjut. Selain itu dapat diketahui pula kelainan-kelainan gen pada proses disimak lebih lanjut. Selain itu dapat diketahui pula kelainan-kelainan gen pada proses tersebut yang sering disebut mutasi gen.
tersebut yang sering disebut mutasi gen.
Cooley pertama kali melaporkan penyakit thalasemia pada tahun 1925.
Cooley pertama kali melaporkan penyakit thalasemia pada tahun 1925.11 KataKata thalasemia berasal dari bahasa Yunani yang berarti laut dan digunakan pertama kali oleh thalasemia berasal dari bahasa Yunani yang berarti laut dan digunakan pertama kali oleh Whipple dan Bradford pada tahun 1932. Penyakit ini tersebar luar dari daerah Mediteranian Whipple dan Bradford pada tahun 1932. Penyakit ini tersebar luar dari daerah Mediteranian sampai kawasan Asia Tenggara termasuk Indonesia. Pada daerah tertentu di
sampai kawasan Asia Tenggara termasuk Indonesia. Pada daerah tertentu di Italia dan negaraItalia dan negara Mediteranian frekuensi carrier thalasemia beta mencapai 15-20%. Di Muang Thai 20% Mediteranian frekuensi carrier thalasemia beta mencapai 15-20%. Di Muang Thai 20% penduduknya mempunyai satu atau jenis lain thalasemia alfa. Secara singkat, thalasemia penduduknya mempunyai satu atau jenis lain thalasemia alfa. Secara singkat, thalasemia merupakan kelainan yang terjadi pada gen globin. Gen globin merupakan bagian dari merupakan kelainan yang terjadi pada gen globin. Gen globin merupakan bagian dari molekul hemoglobin. Kelainan pada gen
molekul hemoglobin. Kelainan pada gen globin ini dapat globin ini dapat menyebabkamenyebabkan thalasemia.n thalasemia.
Hemoglobin Hemoglobin
Hemoglobin adalah molekul yang berbentuk bulat dan terdiri atas empat subunit. Hemoglobin adalah molekul yang berbentuk bulat dan terdiri atas empat subunit.22 Tiap-tiap subunit mengandung satu gugus
Tiap-tiap subunit mengandung satu gugus hemeheme yang terkonjugasi oleh suatu polipeptida.yang terkonjugasi oleh suatu polipeptida. Heme adalah suatu derivate porfirin yang mengandung besi. Polipeptida-polipeptida itu Heme adalah suatu derivate porfirin yang mengandung besi. Polipeptida-polipeptida itu secara kolektif disebut
Pada hemoglobin manusia dewasa normal (hemoglobin A), dua jenis polipeptida dinamakan rantai α, dan masing-masing mengandung 141 residu asam amino, dan rantai β, yang masing-masing mengandung 146 residu asam amino. Jadi, hemoglobin A diberi kode α2β2. Tidak semua hemoglobin di dalam darah orang dewasa normal berupa hemoglobin A.
sekitar 2,5% hemoglobin adalah hemoglobin A2, yang rantai β -nya digantikan oleh rantai δ (α2δ2). Rantai δ juga mengandung 146 residu asam amino, namun 10 macam residu asam
amino-nya berbeda dari residu asam amino di rantai β.
Ada derivat hemoglobin A dalam jumlah kecil yang terkait erat dengan hemoglobin A satu glukosa yang menempel pada valin terminal di setiap rantai β.
Sintesis hemoglobin dibentuk dalam sitoplasma sel dalam proeritroblas dan sampai stadium retikulosit, karena ketika retikulosit meninggalkan sumsum tulang dan masuk ke dalam aliran darah, maka retikulosit tetap membentuk sedikit hemoglobin selama beberapa hari berikutnya. Pertama, suksinil-KoA, yang dibentuk dalam siklus Krebs berikatan dengan glisin untuk membentuk molekul pirol. Kemudian empat pirol bergabung membentuk m9olekul heme. Akhirnya, setiap molekul heme bergabung dengan rantai polipeptida panjang yang disebut globin, yang disintesis oleh ribosom, membentuk suatu subunit hemoglobin yang disebut rantai hemoglobin. tiap-tiap rantai ini mempunyai berat molekul kira-kira 16.000; empat dari molekul ini selanjutnya akan berikatan satu sama lain secara longgar untuk membentuk molekul hemoglobin yang lengkap.
Terdapat beberapa variasi kecil pada rantai subunit hemoglobin yang berbeda, bergantung pada susunan asam amino di bagian polipeptida. Tipe-tipe rantai itu disebut rantai alfa, rantai beta, rantai gamma, dan rantai delta. Bentuk hemoglobin yang paling umum pada orang dewasa, yaitu hemoglobin A, merupakan kombinasi dari dua rantai alfa dan dua rantai beta.
Karena setiap rantai mempunyai sekelompok prostetik heme, maka terdapat 4 atom besi dalam setiap molekul hemoglobin; masing-masing dapat beikatan dengan 1 molekul oksigen, total membentuk 4 molekul oksigen (atau 8 atom oksigen) yang dapat diangkut oleh setiap molekul hemoglobin. Hemoglobin A mempunyai berat molekul 64.458.
Sifat rantai hemoglobin menentukan afinitas ikatan hemoglobin terhadap oksigen. Abnormalitas ranatai ini dapat mengubah sifat-sifat fisik molekul hemoglobin. Contohnya pada anemia sel sabit. Pada orang normal, persentase hemoglobin di setiap sel hampir selalu berada di tingkat maksimal (sekitar 34 mg/dL). Darah mengandung rata-rata 15 gram hemoglobin per 100 mililiter (16 gram pada pria dan 14 gram pada wanita).2,3 Setiap gram hemoglobin murni mampu berikatan dengan sekitar 1,34 mililiter oksigen. Pada orang
normal, lebih dari 20 mililiter oksigen dapat diangkut dalam bentuk terikat ke hemoglobin oleh setiap 100 mililiter darah.
Kelainan hemoglobin disebabkan oleh sintesis hemoglobin abnormal dan berkurangnya kecepatan sintesis rantai globin α- atau β- yang normal (thalasemia α dan β).2,4
Hemoglobin tak stabil jarang ditemukan dan menyebabkan terjadinya anemia hemolitik kronik dengan keparahan yang bervariasi disertai hemolisis intravascular. Hemoglobin abnormal juga dapat menyebabkan terjadinya polisitemia (familial) atau methemoglobinemia kongenital.
Ada dua tipe utama kelainan herediter hemoglobin pada manusia, hemoglobinopati, yaitu pembentukan rantai polipeptida abnormal, dan thalasemia serta gangguan sejenis yaitu terdapat struktur rantai hemoglobin yang normal tetapi dihasilkan dalam jumlah yang sedikit atau tidak diproduksi karena adanya defek pada bagian pengatur gen globin.2,5Jika suatu gen abnormal yang diwariskan dari salah satu orang tua memerintahkan pembentukan hemoglobin abnormal, separuh hemoglobin dalam sirkulasi akan bersifat abnormal dan separuhnya lagi, normal. Bila gen abnormal yang identik diturunkan dari kedua orang tua individu tersebut homozigot dan semua hemoglobinnya abnormal.
Kadar hemoglobin dalam darah yang rendah dikenal dengan istilah anemia. Ada banyak penyebab anemia diantaranya yang paling sering adalah perdarahan, kurang gizi, gangguan sumsum tulang, pengobatan kemoterapi dan abnormalitas hemoglobin bawaan. Kadar hemoglobin yang tinggi dapat dijumpai pada orang yang tinggal di daerah dataran tinggi dan perokok. Beberapa penyakit seperti radang paru paru, tumor dan gangguan sumsum tulang juga bisa meningkatkan kadar hemoglobin.
Defek genetik hemoglobin merupakan kelainan genetik yang paling banyak ditemukan di seluruh dunia. Kelainan ini terdapat di daerah-daerah tropis dan subtropis, dan sebagian besar tampaknya telah terseleksi karena keadaan karier menghasilkan semacam perlindungan terhadap malaria. Thalasemia-β lebih banyak ditemukan di daerah mediterania, sedangkan thalasemia-α lebih banyak ditemukan di Timur jauh.1
Hukum Mendel
Hukum segregasi dan pemilihan independen Mendel adalah aplikasi spesifik dari aturan umum probabilitas serupa yang diaplikasikan untuk pelemparan koin atau dadu.6 Memiliki pemahaman dasar akan aturan peluang ini adalah hal penting dalam analisis genetika. Kisaran angka probabilitas adalah dari 0 sampai 1. Suatu kejadian yang pasti akan
terjadi mempunyai probabilitas 1, sedangkan suatu kejadian yang pasti tidak akan terjadi mempunyai probabilitas 0.
Genetika adalah cabang ilmu dalam biologi yang mempelajari hal-hal yang berhubungan dengan gen atau asal usul.6,7 Dalam genetika akan dipelajari bagaimana sifat keturunan (hereditas) itu diwariskan kepada anak-cucu, serta variasi yang mungkin timbul didalamnya. Sejak penemuan Gregor Mendel (1822 - 1884), pengetahuan genetika berkembang dan menjadi satu lapangan pandang ilmu dalam Biologi. Mendel berhasil mengamati suatu macam sifat keturunan (karakter) dari generasi ke generasi, dan berhasil membuat perhitungan matematika tentang sifat genetis karakter tersebut. Faktor genetisnya disebut determinant . Karena itu, Mendel disebut sebagai Bapak Genetika, yang memberi dasar pengetahuan genetika modern.Kemajuan yang paling pesat dari genetika berlangsung semenjak diakuinya asam nukleat sebagai material dasar gen untuk seluruh organisme. Dengan pengetahuan tentang asam nukleat dapat dipelajari struktur, ekspresi dan proses pewarisan gen secara kimia. Informasi genetika dikandung dalam bentuk runtunan asam amino pada rantai polipeptida protein dan enzim. Protein dan enzim merupakan senyawa-senyawa penting bagi kehidupan organisme.
Teori pertama tentang proses pewarisan yang kebenarannya dapat diterima ialah yang dikemukakan Mendel (1876). Penemuan teori ini menandai lahirnya genetika (Genetika Modern). Mendel mengemukakan teorinya berdasarkan pada hasil penelitian mengenai percobaan persilangan terhadap berbagai varietas kacang kapri ( Pisum sativum).
Kesimpulan dari percobaan Mendel, diantaranya :7
1. Hibrid, adalah hasil persilangan dua individu dengan tanda beda, memiliki sifat yang mirip dengan induknya dan setiap hibrid mempunyai sifat yang sama dengan hybrid. 2. Karakter (sifat) dari keturunan suatu hibrid selalu timbul kembali secara teratur dan
memberi petunjuk bahwa tentu ada faktor-faktor tertentu yang mengambil peranan dalam pemindahan sifat dari satu generasi ke generasi berikutnya.
3. Suatu hukum atau pola akan dapat diketahui dengan cara mengadakan banyak persilangan dan menghitung bentuk-bentuk yang berbeda seperti yang tampak dalam keturunan.
Disamping itu, Mendel juga mengemukakan 4 macam postulat dari hasil penelitiannya, yaitu:7
1. Faktor keturunan (gen) berupa benda dan selalu berpasangan pada individu diploid, bertanggung jawab pada penurunan sifat-sifat.
2. Pada gametogenesis kedua faktor tersebut berpisahan atau disegregasi sehingga setiap gamet hanya memiliki salah satu dari pasangan faktor tersebut. Kemudian dari postulat ini menjadi Hukum Mendel I atau Hukum Segregasi. Dua gen yang disatukan melalui perkawinan akan tetap mempertahankan identitasnya, dan dalam proses pembentukan gamet gen tersebut akan berpisah kembali yang dikenal dengan Hukum Segregasi.
3. Faktor keturunan (gen) ada yang bersifat dominan dan ada yang bersifat resesif, disebut Prinsip dominan-resesif .
4. Bila ada dua faktor keturunan yang diperlukan, maka pada gametogenesis terjadi pilihan bebas (independent assortment ), sehingga segala macam kombinasi mungkin terjadi dalam jumlah yang sama. Postulat ini disebut Hukum Mendel II atau Hukum Pilihan Bebas. Hukum Mendel II berlaku ketika pembentukan gamet, dimana gen sealel secara bebas pergi ke masing-masing kutub ketika meiosis disebut juga Hukum Asortasi. Dihibrid adalah persilangan dengan dua tanda atau dua sifat beda. Mendel
dapat mengambil kesimpulan bahwa anggota dari sepasang gen memisah secara bebas (tidak saling mempengaruhi) ketika berlangsung meiosis selama pembentukan gamet-gamet. Hukum Mendel II disebut juga “ Hukum Pengelompokkan Gen secara Bebas”.
Proses replikasi, transkripsi, dan translasi
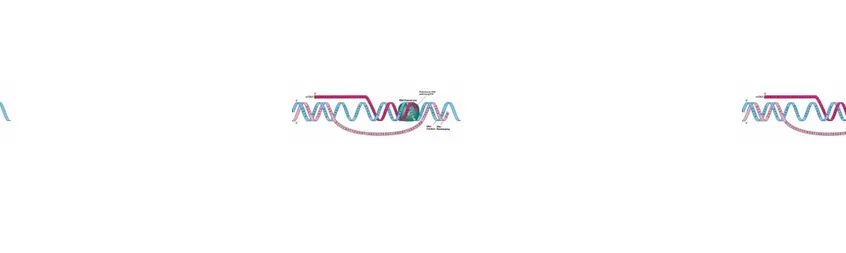
Sintesis DNA terjadi melalui proses replikasi di dalam inti sel. Replikasi (lihat gambar 1) diawali dengan terbukanya pilinan dan pemisahan rantai oleh enzim helikase atau enzim topoisomerase sehingga terbentuk dua pita ikat tunggal. Kedua pita tersebut berfungsi sebagai cetakan DNA baru yang sama dengan bantuan enzim DNA polymerase. Satu sifat DNA yang double helix yang mempengaruhi replikasi yaitu kedua DNA bersifat antiparalel artinya ikatan gula-fosfat kedua pita berlawanan arah. Jika dibayangkan untai double helix yang panjang yang hanya terdiri dari fosfat, gula pentosa, dan basa diberi nomor 1 hingga 5 terdapat gugus fosfat berikatan pada rantai C no 3 dan 5. Hasilnya terdapat dua buah pita DNA baru dengan arah 5‟→3‟. Oleh karena itu , dalam pembentukan DNA baru akan
terdapat pembentukan pita yang kontinu (5‟→3‟) tanpa terputus dan diskontinu (3‟→5‟)
terputus-putus. Pembentukannya diawali dengan pembentukan RNA primer oleh enzim primase dan diteruskan oleh DNA polymerase membentuk fragmen DNA yang disebut fragmen Okazaki. RNA primer akan digantikan DNA bersamaan dengan penyambungan fragmen Okazaki oleh enzim ligase. Akibatnya terbentuk pita DNA baru yang utuh.
Gambar 1. Replikasi DNA (sumber: www.google.com)
Proses sintesis protein terbagi atas transkripsi dan translasi (lihat gambar 2). Seperti telah diketahui bahwa DNA sebagai media untuk proses transkripsi suatu gen berada di kromosom dan terikat oleh oleh protein histon. Saat menjelang prosestranskripsi berjalan, biasanya didahului signal dari luar akan kebutuhan suatu protein atau molekul lain yang dibutuhkan untuk proses pertumbuhan, perkembangan, metabolisme, dan fungsi lain di tingkat sel maupun jaringan. Kemudian RNA polymerase II akan mendatangi daerah regulator elemen dari gen yang akan ditranskripsi. Kemudian RNA polymerase ini akan menempel (binding) di daerah promotor spesifik dari gen yang akan disintesis proteinnya, daerah promotor ini merupakan daerah consensus sequences, pada urutan -10 dan -35 dari titik inisiasi (+1) yang mengandung urutan TATA-Box sebagai basal promotor. Setelah itu polymerase ini akan membuka titik inisiasi (kodon ATG) dari gen tersebut dan mengkopi semua informasi secara utuh baik daerah axon maupun intron, dalam bentuk molekul immature mRNA (messenger RNA). Kemudian immature mRNA ini diolah pada proses splicing dengan menggunakan smaalnuclearRNA (snRNA) complex yang akan memotong hanya daerah intron, dan semua exon akan disambungkan menjadi satu urutan gen utuh tanpa non-coding area dan disebut sebagai mature mRNA. Pada tahap berikutnya, mRNA ini diproses lebih lanjut pada proses translasidi dalam ribosom, dalam tiga tahapan pokok yaitu inisiasi yang mengawali sintesis polipeptida dari kodon AUG yang ditranslasi sebagai asam
amino metionin. Secara singkat, proses ini berlangsung dengan bantuan initiation factor (IF-1. IF-2, dan IF-3) dan enzim tRNA-metionin sintetase (pada bakteri diawali oleh formylmethionine) sehingga tRNA dan asam amino metionin membentuk ikatan cognate dan bergerak ke ribosom tempat sintesis protein berlangsung. Langkah selanjutnya adalah elongasi atau pemanjangan polipeptida sesuai dengan urutan kodon yang dibawa oleh mRNA.
Pada proses elongasi ini diperlukan elongation factor complex. Seperti juga proses inisiasi enzim tRNA-amino acid snthethase berperan dalam pembentukan cognate antara tRNA dan asam amino lainnya dari sitoplasma yang sesuai dengan urutan kodon mRNA tersebut. Proses elongasi akan berhenti sampai kodon terminasi dan poly-adenyl (poly-A), dan diakhiri sebagai proses terminasi yang dilakukan oleh rho-protein. Polipeptida akan diproses sebagai molekul protein yang fungsional setelah melalui proses post translation di retikulum endoplasmik (RE) hingga tingkat jaringan.
Gambar 2. Transkripsi dan translasi (sumber: www.google.com)
Gambar 3. Proses transkripsi (sumber: www.google.com) Proses transkripsi dibagi menjadi empat tahap yaitu :5,8
1. Pra-inisiasi
Pada eukariota, RNA polymerase, untuk inisiasi transkripsi, memerlukan kehadiran sekuens promotor dalam DNA. Promotor adalah daerahc dari DNA yang tempat menempelnya RNA polymerase. RNA polymerase harus mampu mengikat promotor dengan bantuan berbagai faktor transkripsi spesifik. Perhatikan gambar 3.
Jenis yang paling umum promotor pada eukariota adalah sekuens DNA pendek yang dikenal sebagai kotak TATA. Kotak TATA, sebagai promotor, adalah situs pengikatan untuk faktor transkripsi yang dikenal sebagai TATA binding protein (TBP), yang merupakan subunit dari faktor transkripsi yang lain, yang disebut Transcription Factor II D (TFIID). Setelah TFIID mengikat mengikat ke kotak TATA melalui TBP, lima faktor transkripsi dan RNA polymerase bergabung di sekeliling kotak TATA dalam serangkaian tahapan pra-inisiasi untuk membentuk kompleks. Salah satu faktor transkripsi, DNA helikase, memiliki aktivitas helikase sehingga berfungsi memisahkan helai DNA beruntai ganda untuk menyediakan untai tunggal DNA template. Namun, hanya yang rendah, atau basal, tingkat transkripsi digerakkan oleh kompleks pra-inisiasi sendirian. Protein lain yang dikenal sebagai activator dan repressor, bersama dengan yang terkait coactivators atau corepressors, bertanggung jawab untuk modulasi tingkat transkripsi.9
2. Inisiasi
Inisiasi transkripsi pada eukariota lebih kompleks (lihat gambar 4). RNA polymerase eukariot tidak secara langsung mengenali urutan promoter inti. Sebaliknya, kumpulan protein yang disebut faktor transkripsi menengahi pengikatan RNA polymerase dan dimulainya transkripsi. Hanya setelah faktor transkripsi tertentu yang melekat pada promotor baru RNA polymerase dapat mengikatnya. Pengikatan
faktor-faktor transkripsi dan RNA polymerase pada promotor, membentuk kompleks inisiasi transkripsi.9
Gambar 4. Proses inisiasi (sumber: www.google.com) 3. Elongasi
Gambar 5. Proses elongasi (sumber: www.google.com)
Satu untai DNA, untai template (atau noncoding untai), digunakan sebagai template untuk sintesis RNA. Dalam proses transkripsi, RNA polymerase melintasi untai cetakan dan menggunakan kecocokan pasangan basa dengan DNA template untuk membuat salinan RNA. Meskipun RNA polymerase melintasi untai template dari 3‟→5‟, untai pengkodean (non template) dan RNA yang baru terbentuk juga
terjadi 5‟→3‟. Ini menghasilkan molekul RNA dari 5‟→3‟, sebuah duplikat yang sama persis pengkodean untai (kecuali yang timin digantikan dengan urasil dan nukleotida terdiri dari sebuah gula ribosa (5-karbon) sedangkan DNA telah terdeoksiribosa (kurang satu atom oksigen) dalam tulang punggung gula fosfat.
Tidak seperti replikasi DNA, transkripsi mRNA dapat melibatkan berbagai RNA polymerase pada satu cetakan DNA dan beberapa putaran transkripsi (amplifikasi mRNA tertentu), begitu banyak molekul mRNA dapat dengan cepat dihasilkan dari satu salinan gen.9 Amati gambar 5.
4. Terminasi
Berakhirnya polymerase RNA ditandai oleh disosiasi kompleks transkripsi atau terlepasnya enzim RNA polymerase beserta kofaktor-kofaktornya dari untai DNA cetakan. Begitu pula halnya dengan molekul RNA hasil sintesis. Hal ini terjadi ketika RNA polymerase mencapai urutan basa tertentu yang disebut dengan terminator.
Terminasi transkripsi dapat terjadi oleh dua macam sebab, yaitu terminasi yang hanya bergantung kepada urutan basa cetakan (disebut terminasi diri) dan terminasi yang memerlukan kehadiran suatu protein khusus (protein rho). Di antara keduanya terminasi diri lebih umum dijumpai. Terminasi diri terjadi pada urutan basa palindrom yang diikuti oleh beberapa adenine (A). Urutan palindrom adalah urutan yang sama jika dibaca dari dua arah yang berlawanan. Oleh karena urutan palindrom ini biasanya diselingi oleh beberapa basa tertentu, maka molekul RNA yang dihasilkan akan mempunyai ujung terminasi berbentuk batang dan kala ( loop).10 Lihat gambar 6.
Gambar 6. Proses terminasi (sumber: www. nayfahsai.blogspot.com)
Translasi
Tahapan-tahapan dalam proses translasi, yaitu :5,8 1. Inisisasi
Inisiasi translasi pada organism eukariotik melibatkan faktor inisiasi yang lebih banyak daripada translasi prokariotik. Faktor inisiasi translasi untuk organisme eukariot dinamai eIFs (eukaryote Initiation Factor ). Komponen pada tahap inisisasi organisme eukariot adalah kodon inisiasi (AUG), lima faktor inisiasi (eIF1, eIF2, eIF3, eIF4, eIF5), tRNA inisiator (met-tRNA), ribosom subunit 40S dan 60S, GTP dan ATP. Pada eukariot tRNA inisiator aminoasil-tRNA, GTP dan eIF2 membentuk kompleks terlebih dahulu sebelum berikatan dengan subunit 40S.9
Tahap pertama meliputi pembentukan kompleks pre-inisiasi (pre-initiation complex). Struktur ini terdiri dari subunit 40S ribosom, „ternary complex‟ yang tersusun dari faktor inisiasi eIF-2 yang terikat tRNA Met inisiator, molekul GTP, dan tiga faktor eIF-1, eIF-1A, eIF-3. Seperti pada bakteria, tRNA inisiator ini tidak mengenali kodon internal 5‟-AUG-3‟. Kompleks pre-inisiasi selanjutnya bergabung dengan u jung 5‟ mRNA. Tahapan ini memerlukan kompleks pengikatan tudung (cap binding complex), kadang-kadang disebut 4F, yang terdiri dari faktor inisiasi 4A, Eif-4E, dan 4G. faktor inisiasi 4G berfungsi sebagai jembatan antara
eIF-Hasil dari tahap ini adalah kompleks pre-inisiasi menjadi terikat pada daerah ujung 5‟ mRNA. Pengikatan ini juga dipengaruhi oleh ekor poli (A) ujung 3‟ mRNA. Interkasi ini diduga dimediasi oleh protein PADP (polyadenylate-binding protein), yang terikat pada ekor poli (A).11
Setelah kompleks pre-inisiasi mengikat ujung mRNA, kompleks ini sekarang disebut kompleks inisiasi (initiation complex), harus menggeserkan posisinya (scanning) sepanjang mRNA sampai mencapai kodon inisiasi. Daerah yang harus dipindai (scanning) ini disebut daerah leader mRNA eukariotik, panjangnya dapat beberapa puluh atau bahkan ratusan nukleotida dan seringkali mengandung daerah yang membentuk struktur tusuk konde (hairpins) dan struktur pasangan basa lain. Ada dugaan, struktur tersebut dihilangkan kombinasi faktor inisiasi eIF-4A, dan mungkin juga eIF-4B, mempunyai aktivitas helikase yang dapat memutuskan ikatan basa intramolekuler mRNA sehingga dapat melapangkan jalan kompleks inisiasi. Kodon inisiasi yang biasanya 5‟AUG3‟ pada eukariot, dapat dikenali sebab urutan ini terdapat dalam urutan consensus pendek, 5‟ACCAUGG3‟, yang dikenal sebagai consensus Koza (Kozak consensus).
Ketika kompleks inisiasi telah menduduki kodon inisiasi, subunit besar ribosom akan mengikat kompleks inisiasi ini. Seperti pada bakteria, tahap ini memerlukan hidrolisis GTP dan pelepasan faktor-faktor inisiasi. Faktor inisiasi terakhir yang terlibat pada tahap ini adalah eIF-5 (yang membantu pelepasan faktor-faktor inisiasi lain) dan eIF-6 (yang bergabung dengan subunit besar yang tidak terikat dan mencegah untuk menempel pada subunit kecil di dalam sitoplasma).
2. Elongasi
Setelah kompleks inisiasi terbentuk proses translasi dilanjutkan dengan pemanjangan rantai polipeptida (tahap elongasi). Mekanisme elongasi pada organisme eukariot mirip dengan mekanisme elongasi pada organisme prokariot. Tahap awal elongasi adalah pengikatan aminoasil-tRNA pada situr A melalui penggabungan dengan kodon kedua mRNA. Aminoasil-tRNA memasuki ribosom dengan bantuan faktor elongasi eIF-1 membentuk kompleks dengan GTP. Faktor elongasi merupakan protein yang berasosiasi terus menerus dengan ribosom selama penambahan asam amino pada rantai polipeptida.
3. Terminasi
Pemanjangan rantai polipeptida berlangsung terus menerus sampai kodon yang tidak dikenali oleh anti kodon yang dibawa oleh tRNA. Sel prokariot dengan sel eukariot juga tidak memiliki tRNA dengan anti kodon yang komplemen dengan sinyal terminasi, tetapi memiliki faktor terminasi (release factor , RF) yang mengenali sinyal terminasi sintesis protein. Pada organisme eukariot hanya terdapat satu faktor terminasi yaitu RF yang mengenali ketiga kodon stop UAG, UAA, dan UGA. RF berikatan dengan kodon terminasi pada situs A ribosom dan menstimulasi hidrolisis ikatan tRNA dan rantai polipeptida pada situs P yang menyebabkan rantai polipeptida lepas dari ribosom, tRNA dilepaskan serta subunit ribosom dan template mRNA berdisosiasi.
Pada bakteri proses transkripsi dan translasi bisa terjadi bersamaan karena DNA berada di sitoplasma dan semua proses replikasi, transkripsi, dan translasi terjadi di sitoplasma. Bahkan mRNA yang belum selesai ditranskripsi sudah mulai ditranslasi. Tetapi pada eukariot keadaannya berbeda, karena replikasi dan transkripsi terjadi di dalam inti. mRNA kemudian dibawa ke sitoplasma untuk ditranslasi.
Mutasi
DNA merupakan molekul yang mudah rusak. Namun integritasnya dilindungi oleh enzim-enzim yang sanggup memperbaiki secara cermat beberapa jenis kerusakan tertentu pada salah satu rantai polinukleotida sepanjang rantai lainnya utuh.
DNA dapat mengalami kerusakkan yang disebabkan oleh faktor kimia dan fisika seperti :
1. Sinar UV
2. Radiasi yang bersifat mengionisasi 3. Senyawa kimia yang reaktif
4. Perubahan ph 5. Suhu
Mutasi diakibatkan kerusakkan nukleotida DNA atau dari kesalahan yang tidak diperbaiki selama replikasi dapat ditranskripsi ke mRNA menyebabkan susunan abnormal dari asam amino. Mutagen adalah faktor yang merusak DNA dan menyebabkan mutasi. Mutagen dapat menyebabkan sel normal menjadi sel kanker. Mutasi pada gen dapat mengarah pada munculnya alel baru dan menjadi dasar bagi kalangan pendukung evolusi
mengenai munculnya variasi-variasi baru pada spesies. Mutasi terjadi pada frekuensi rendah di alam, biasanya lebih rendah daripada 1 : 10.000 individu. Mutasi di alam dapat terjadi akibat zat pembangkit mutasi (mutagen, termasuk karsinogen), radiasi surya maupun radioaktif, serta loncatan energi listrik seperti petir.12
Mutasi gen yaitu perubahan pasangan basa pada satu gen. Bila melibatkan satu pasang basa maka disebut mutasi titik. Mutasi titik dapat dibedakan lagi menjadi:7
a. Delesi
Delesi merupakan hilangnya sebagian segmen kromosom karena putus secara tunggal atau ganda (perhatikan gambar 7). Bagian kromosom yang terpisah bila tidak memiliki sentromer disebut Fragmen Asentrik . Fragmen asentrik tidak akan ditarik ke kutub dan hilang pada proses mitosis atau meiosis. Pada Gambar di bawah ini dapat dilihat proses delesi kromosom, pada B terbentuk kromosom cincin bila terjadi putus di bagian terminal dan kromosom dengan ujung lengket akan membentuk kromosom cincin. Pada C I bagian kromosom yang terpisah masuk di antara kromosom lain dengan putus secara tunggal dan terbentuk insersi ( C II).
Gambar 7. Delesi kromosom
(sumber :www.ghr.nlm.nih.gov/handbook/illustrations/chromosomaldeletion.jpg) b. Duplikasi
Duplikasi terjadi karena hasil pindah silang antara kromosom homolog yang tidak sebanding (sama), sehingga ada gen yang ganda (perhatikan gambar 8). Duplikasi lebih umum didapat dan efeknya tidak sejelek delesi, bahkan duplikasi kecil mungkin merupakan salah satu mekanisme evolusi.
Gambar 8. Duplikasi kromosom
(sumber : www.ghr.nlm.nih.gov/handbook/illustrations/chromosomalduplication.jpg) c. Inversi
Inversi dimulai dengan putusnya kromosom pada dua tempat (amati gambar 9). Bagian kromosom yang terpisah berputar 180o, kemudian bertaut kembali. Disini urutan gen yang mengalami inversi jadi terbalik. Misalnya kromosom dengan urutan gen ABCDEFGH setelah putus antara CD dan FG kemudian membentuk kromosom lagi dengan urutan ABCFEDGH.
Bila putusnya di salah satu pihak sentromer maka terjadi inversi parasentrik . Tetapi bila putusnya di sebelah menyebelah sentromer, maka terbentuk inversi perisentrik.
Gambar 9. Inversi kromosom
d. Insersi
Insersi adalah suatu tipe translokasi dimana potongan kromosom itu masuk di antara kromosom nonhomolog yang putus. Untuk memudahkan penafsiran proses insersi perhatikan gambar 10.
Gambar 10. Insersi kromosom
(Sumber: www.accessexcellence.org/RC/VL/GG/images/insertion.gif) e. Translokasi
Translokasi adalah suatu peristiwa berpindahnya bagian kromosom ke bagian kromosom non-homolog. Kejadian ini juga dimulai dengan putusnya kromosom pada kedua kromosom non-homolog tersebut. Translokasi resiprok yaitu perpindahan segmen kromosom timbal balik. Dari translokasi ini dapat menimbulkan translokasi homozigot dan translokasi heterozigot. Ada juga translokasi tidak seimbang dan seimbang. (lihat gambar 11 dan 12).
Gambar 11. Unbalanced translocation
(Sumber : www.ghr.nlm.nih.gov/handbook/illustrations/unbalancedtranslocation.jpg) Gambar 12. Balanced translocation
(sumber: www.ghr.nlm.nih.gov/handbook/illustrations/balancedtranslocation.jpg) Contoh mutasi titik adalah penyakit thalasemia-β (thalasemia ditandai dengan cacat pada proses sintesis subunit hemoglobin, sehingga penyakit thalasemia- β akan terjadi kalau poduksi β-globin tidak mencukupi).13 Mutasi yang terjadi dalam gen akan merubah kodon sehingga merubah asam amino yang di kode oleh kodon sehingga dapat merubah polipeptida. Namun, tidak semua mutasi yang merubah kodon akan merubah asam amino, karena adanya asam amino yang di kode oleh beberapa kodon. Mutasi gen dapat juga dibedakan berdasarkan pengaruhnya terhadap kodon, yaitu :9
1. Mutasi frameshift adalah mutasi yang disebabkan oleh penyisipan atau penghapusan dari sejumlah nukleotida yang tidak terbagi merata menjadi tiga dari sekuens DNA. Karena sifat triplet ekspresi gen oleh kodon, penyisipan atau penghapusan dapat mengganggu pembacaan, sehingga menghasilkan terjemahan yang sama sekali berbeda dari aslinya. Semakin awal dalam urutan penghapusan atau penyisipan terjadi, maka semakin mengubah protein yang dihasilkan.
2. Sebuah mutasi nonsense adalah sebuah titik dalam urutan mutasi DNA yang menghasilkan kodon stop premature, atau kodon nonsense dalam transkripsi mRNA, dan sintesis protein terhenti sebelum waktunya, dan produk protein yang dihasilkan sering nonfungsional.
3. Missense mutasi atau mutasi nonsynonymous adalah jenis mutasi titik dimana satu nukleotida yang berubah menyebabkan penggantian asam amino yang berbeda. Hal ini dapat membuat protein yang dihasilkan tidak berfungsi. Mutasi seperti ini dapat menyebabkan penyakit seperti Epidermolysis bullosa, penyakit sel sabit, dan ditengahi SODI ALS.
4. Mutasi netral adalah mutasi yang terjadi pada kodon asam amino yang akan menghasilkan asam amino yang kegunaannya berbeda, tetapi secara kimiawi mirip. Kesamaan antara keduanya mengakibatkan tidak berpengaruhnya produk protein yang dihasilkan. Sebagai contoh, perubahan dari AAA ke AGA akan menyandikan lisin, molekul yang secara kimiawi serupa dengan arginin.
5. Silent mutasi adalah mutasi yang tidak menghasilkan perubahan ke sekuens asam amino dari protein. Mutasi ini mungkin terjadi di daerah yang tidak mengkode protein atau mungkin terjadi dalam suatu kodon dalam cara yang tidak mengubah produk asam amino.
Mutasi yang paling berbahaya adalah mutasi Missense dan Nonsense. Keduanya dapat mengakibatkan protein yang dihasilkan menjadi tidak berfungsi. Salah satu contoh mutasi missense dan nonsense adalah mutasi gen reseptor insulin.
Kesimpulan
Seperti yang telah dibahas bahwa hemoglobin merupakan molekul yang berbentuk bulat dan terdiri atas empat subunit. Tiap-tiap subunit mengandung satu gugus heme yang akan bergabung dengan globin menjadi hemoglobin. Dalam pembentukannya (sintesis hemoglobin), dapat pula terjadi berbagai mutasi pada gen globin yang akan menyebabkan kelainan pada tubuh manusia. Mutasi tersebut bermacam-macam jenisnya. Salah satu contoh hasil mutasi tersebut adalah thalasemia. Thalasemia ditandai dengan cacat pada proses sintesis subunit hemoglobin, sehingga penyakit thalasemia-β akan terjadi kalau poduksi β-globin tidak mencukupi. Hal ini dapat diprediksi dengan menggunakan teori probabilitas yang berdasarkan pada hokum mendel.
Daftar pustaka
1. Soeparman, Waspadji S. Ilmu penyakit dalam jilid II. Jakarta: Balai Penerbit FKUI, 1999. 2. Ganong WF. Fisiologi kedokteran. Edisi ke-22. Jakarta: EGC, 2008.
3. Hall JE. Fisiologi kedokteran. Edisi ke-11. Jakarta: EGC, 2010.
4. Hoffbrand AV, Pettit JE, Moss PAH. Hematologi. Jakarta: EGC, 2005. 5. Roberts JA, Pembrey ME. Genetika kedokteran. Edisi ke-8. Jakarta: 2002.
6. Rikky F, Agus MH, Umar R. Mudah dan aktif belajar biologi. Bandung: Setia Purna Invers, 2007.
7. Priastini R, Hartono B. Biologi kedokteran blok 4. Jakarta: Fakultas Kedokteran Universitas Kristen Krida Wacana, 2010.
8. Price SA, Wilson LM. Hereditas, lingkungan, dan penyakit. Jakarta: EGC, 2006. 9. Gaffar S. Buku ajar bioteknologi molekul. Bandung: Universitas Padjajaran, 2007.
10. Champe, Pamela C, Harvey, Richard A, Ferrier, Denise R. Lippincott‟s illustrated reviews: biochemistry. 3rd ed. Hagerstown: Lippincott Williams & Wilkins, 2004.
11. Cox, Michael, Nelson, David R, LehningerAlbert L. Lehninger principles of biochemistry. 4th ed. San Fransisco: W. H. Freeman, 2005.
12. Burrus V, Waldor M. Shaping bacterial genomes with integrative and conjugative elements. London: Res Microbio, 2004.
13. Murray RK, Granner DK, Mayes PA, Rodwell VW. Biokimia Harper. 27th ed. Jakarta: EGC, 2009.