www.elsevier.com / locate / bres
Research report
Identification of deflation-sensitive inspiratory neurons in the dorsal
respiratory group of the rat
*
Kazuhisa Ezure , Ikuko Tanaka
Department of Neurobiology, Tokyo Metropolitan Institute for Neuroscience, 2-6 Musashidai, Fuchu, Tokyo 183-8526, Japan Accepted 15 August 2000
Abstract
It has been well established that inspiratory neurons of the dorsal respiratory group (DRG) are classified into two types based on whether they receive inputs from slowly-adapting pulmonary stretch receptors (SARs) or not. Inspiratory neurons with SAR inputs are called Iband the others are called Ianeurons. In this study, we identified a novel group of inspiratory neurons in Nembutal-anesthetized, paralyzed, and artificially-ventilated rats. (1) These DRG inspiratory neurons were activated characteristically by lung deflation. (2) They were orthodromically activated by electrical stimulation of the vagus nerve at a low intensity just above the threshold for afferents from SARs or rapidly-adapting pulmonary stretch receptors (RARs). (3) The orthodromic latencies (ranged from 1.9 to 2.5 ms) indicated that they receive direct inputs from low-threshold vagal afferents. (4) Unlike Ibneurons, they hardly responded to lung inflation and never exhibited tonic firing in response to maintained lung inflation. (5) The majority (92%) of them were antidromically activated by electrical stimulation of the cervical spinal cord. These deflation-sensitive inspiratory neurons clearly form a distinct group, and their firing pattern is consistent with the hypothesis that they receive inputs from RAR afferents as well as the central inspiratory drive. The results indicating that DRG inspiratory neurons are classified into at least three groups provide new insights into the organization and role of the DRG.
2000 Elsevier Science B.V. All rights reserved.
Theme: Endocrine and autonomic regulation
Topic: Respiratory regulation
Keywords: Respiration; Nucleus tractus solitarii; Lung deflation; Inspiratory neuron; Rapidly-adapting pulmonary stretch receptor
1. Introduction the cat. However, detailed firing properties in some types of DRG neurons are not necessarily the same as those of In our previous study in the rat [15], we could find too cats. For instance, pump neurons (P-cells) [3] which few respiratory neurons in the dorsomedial medulla and receive inputs from the slowly-adapting pulmonary stretch reported that the dorsal respiratory group (DRG) that was receptors (SARs) [19] exhibit various properties which initially identified in the cat [2,3,6] might not exist in the have not been described in the cat [26]. Furthermore, firing rat. The controversy about the presence or absence of rat patterns of Ib neurons, which have combined central DRG [15,31,35] has been settled by de Castro et al. [13], inspiratory and SAR inputs, seem different from those in who showed clearly the presence of inspiratory neurons the cat [13]. Since the rat is now the most important which were located in the ventrolateral subnucleus of the experimental animal for study of the neural control of tractus solitarii (NTS). In addition, they showed that breathing, we have re-examined the firing properties of various respiration-related neurons of rat DRG had firing respiration-related neurons of the DRG.
patterns generally similar to those previously described in Lung inflation and withholding inflation are frequently-used maneuvers to examine inputs from lung stretch receptors and classify respiratory neurons [8,10]. On the
*Corresponding author. Tel.: 181-42-325-3881; fax: 1
81-42-321-other hand, lung deflation generated by either lowering a
8678.
E-mail address: [email protected] (K. Ezure). positive end-expiratory pressure (PEEP) or allowing the
K. Ezure, I. Tanaka / Brain Research 883 (2000) 22 –30 23
lungs to collapse to atmospheric pressure is a specially lactated Ringer’s solution with 5% glucose was intraven-useful maneuver to examine inputs from the rapidly-adapt- ously administered when necessary. Rectal temperature ing pulmonary stretch receptors (RARs) [11,17,19] but has was maintained within 36–378C using a heating lamp. End not been adopted for classifying respiratory neurons. In the tidal CO2 was monitored and maintained at 4–5% by present study, we found a group of inspiratory neurons adjusting tidal volume or frequency of a respirator. which were characteristically activated by deflation of the
lungs. 2.2. Recording and experimental protocol
DRG neurons were recorded extracellularly with glass
2. Materials and methods micropipettes filled with 3 M KCl saturated with Fast Green FCF dye (DC impedance, about 1.0 MV). The Experiments were conducted on 14 adult Wistar rats recording sites were marked with the dye. After fixation, (260–425 g). All experimental procedures were performed serial frozen sections of the brainstem (100 mm thick in in accordance with the Guiding Principles for the Care the frontal plane) were made and stained with cresyl violet.
and Use of Animals in the Field of Physiological Science At the beginning of each experiment, we estimated the (Physiological Society of Japan, 1988). The experiments thresholds for orthodromic activation of the second-order were reviewed and approved by the Animal Experiment relay neurons from SARs (P-cells) by vagal nerve stimula-Committee of the Tokyo Metropolitan Institute for Neuro- tion. For this purpose we sampled at least five P-cells on science. each side and measured their thresholds: these thresholds Almost all the experimental procedures, such as surgery, for P-cells are about the same as or a little higher than care of animals, vagal nerve stimulation, electrodes, neural those for the second-order relay neurons from RARs recording, antidromic stimulation, or histological recon- (RAR-cells) (see Refs. [16,17]). Stimulus intensity less struction of the brainstem, were the same as those de- than two times the mean threshold (normally less than 10 scribed previously [17,18,25,26], and are described here mA) of P-cells was routinely used for vagal nerve
stimula-only briefly. tion.
The means of generating maintained lung inflation and 2.1. Surgical procedures deflation were fully described previously [17,18]. Signals (microelectrode potentials, phrenic nerve discharge, trache-The rats were initially anesthetized with sodium pen- al pressure and blood pressure) were stored on magnetic tobarbitone (Nembutal, 70 mg / kg; i.p.); supplementary tape (DAT recorder: PC-216A, Sony Precision Technolo-doses (about 5 mg / kg per h; i.v.) were given during gy, Tokyo; sampling rate 50ms).
surgery and recording as necessary. The trachea was intubated, and cannulae placed in the femoral artery to
monitor blood pressure and in the femoral vein for drug 3. Results
administration. Bipolar cuff electrodes were attached to the
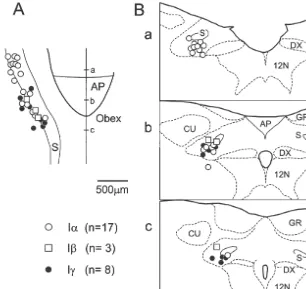
cervical vagus nerves on both sides for stimulation, but the This study was based on a total of 67 inspiratory nerves were not cut. The superior laryngeal nerve (in six neurons which were recorded from the nucleus tractus rats) and the phrenic nerve (C4 / 5 branch) on one side were solitarii (NTS) and its vicinity. Neural recordings were dissected, cut distally and the central ends were mounted made in the area extending from 1.0 mm rostral to and 1.0 on bipolar cuff electrodes for stimulation and recording, mm caudal to the obex (defined as the caudal end of the respectively. The nerves prepared for stimulation and area postrema), within 1.5 mm lateral to the midline and recording were kept in oil pools. In addition, stimulating within 1.2 mm below the dorsal surface. In 28 of the 67 electrodes were inserted bilaterally into the ventrolateral neurons their locations were marked and confirmed within funiculus at the rostral C4 level in nine rats. the NTS area (Fig. 1).
The animals were placed in a stereotaxic frame and
occipital craniotomy and partial cerebellectomy were 3.1. Deflation-sensitive inspiratory neurons conducted to expose the dorsal surface of the medulla. The
Fig. 1. Distribution of Ia, Iband deflation-sensitive (Ig) neurons in the NTS. Distribution of 28 neurons are projected onto horizontal (A) and frontal planes (B). Three planes (a,b,c) in B correspond to levels (a,b,c) shown in A. AP, area postrema; CU, cuneate nucleus; DX, dorsal motor nucleus of the vagus; GR, gracile nucleus; S, solitary tract; 12N, hypoglossal nucleus.
transient firing which was locked to deflation of the lungs. 3.2. Ia and Ib neurons
The firing started at the onset of the deflation (Fig. 3).
During control respiratory period, especially when PEEP Besides Igneurons, several distinct types of respiration-levels were lowered, their firing at the deflation phase was related neurons were found. Many P-cells and RAR-cells conspicuous (Fig. 2Ad,Bd,C). These firing patterns were were recorded from but were discarded in this study since similar to those of RAR afferents and RAR-cells [17,18]. their firing properties and distributions have been fully However, these neurons hardly responded to lung inflation described previously [17,25]. We recorded from 50 in-(Fig. 2Aa), although some weak firing at the initial part of spiratory neurons consisting of Iaand Ibneurons (Fig. 1). inflation was occasionally seen (Fig. 2Ba). Tentatively, we Ib neurons (n59) were identified by (1) their activity at call these deflation-sensitive inspiratory neurons ‘Ig neu- the inspiratory phase, (2) monosynaptic activation by rons’ (see Section 4). electrical stimulation of the vagus nerve, and (3) activity All of the Ig neurons (n517) were orthodromically evoked by lung inflation, specifically tonic activity by activated by electrical stimulation of the vagus nerve (Fig. maintained lung inflation (Fig. 4). Orthodromic latencies 2Ae). The orthodromic latencies ranged from 1.9 to 2.5 ms following vagus nerve stimulation ranged from 1.8 to 2.2 (mean6S.D. 2.160.21 ms) (Fig. 6). The intensities of ms (mean6S.D. 2.060.11 ms) (Fig. 6). Two Ib neurons stimulation for these responses were within two times the examined could not be activated antidromically from the thresholds for P-cell or RAR-cell activation (see Section spinal cord, and one of them was orthodromically activated 2). The majority (12 of 13) of the Ig neurons examined from the SLN (latency 3.0 ms).
K. Ezure, I. Tanaka / Brain Research 883 (2000) 22 –30 25
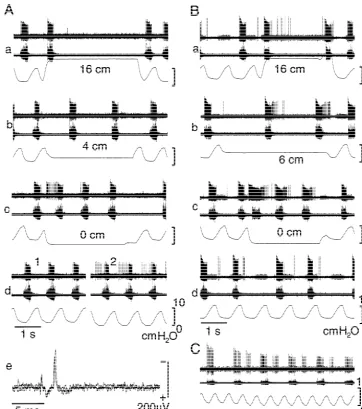
Fig. 2. Firing pattern of Igneurons. Three Igneurons (A–C) are shown. Traces are extracellular spike activity (top), phrenic nerve activity (middle), and tracheal pressure (bottom). These inspiratory neurons are hardly activated by lung inflation (Aa,Ba) but consistently activated at lung collapse to atmospheric pressure (Ac,Bc). Pressure of maintained inflation is shown on each panel by number (cmH O). These neurons are also activated at deflation2
phase of artificial ventilation (Ad,Bd,C) especially when PEEP is lowered (Ad2,C). Ad: responses at two different PEEP levels; C: responses during lowering PEEP. Igneurons are orthodromically activated by electrical stimulation of the ipsilateral vagus nerve (Ae). Ae: three superimposed traces of the responses to vagal nerve stimulation.
Since these neurons were neither Ib nor Ig neurons, they Ig neurons were located more caudally. In general, it was were all classified as Ianeurons. A few Ianeurons (5 / 41) easy to find Ia neurons but difficult to find Ib neurons, were activated orthodromically by electrical stimulation of presumably because the population of Ibneurons is small. the vagus nerve (Fig. 6). The orthodromic latencies ranged In two experiments which focused on sampling Ib neu-from 1.8 to 2.5 ms (mean6S.D. 2.160.30 ms). Two of the rons, no Ib neurons could be found. Ig neurons could be 21 Ia neurons examined projected to the contralateral found more easily than Ib neurons.
spinal cord. Electrical stimulation of the SLN orthodromi-cally activated the majority (23 / 29) of the Ia neurons
examined. The orthodromic latencies ranged from 1.8 to 4. Discussion
2.9 ms (mean6S.D. 2.360.29 ms).
Fig. 3. Expanded firing pattern of Igneurons at the lung deflation. Three Igneurons (A–C): neurons A and B are the same as neurons A and B shown in Fig. 2. The start of their firing coincides with the deflation of the lungs (shown by vertical bars). The majority of the Igneurons project
Fig. 4. Firing pattern of Ibneuron. During control respiration Ibneurons to the spinal cord (D). (D) Three superimposed traces of collision test for
exhibit firing in response to ventilator-induced lung inflation and the antidromic activation from the spinal cord (same neuron as shown in A).
inspiratory phase (Ad). Ibneurons fire tonically in response to maintained Stimuli (arrows) were applied 3 ms (1) and 2 ms (2) after spontaneous
lung inflation (Aa). Their firing locked to the inspiratory phase becomes spikes.
evident when lung inflation is maintained at lower levels (Ab,Ac). Ib
neurons are orthodromically activated by electrical stimulation of the ipsilateral vagal nerve with stable latencies. (B) Four superimposed traces of the responses to vagal nerve stimulation (same neuron as A).
inputs that activate these neurons, and (3) their functional meaning in respiration.
K. Ezure, I. Tanaka / Brain Research 883 (2000) 22 –30 27
Fig. 6. Orthodromic latencies after vagal nerve stimulation. All Iband Ig
neurons examined were orthodromically activated, but only five of 41 Ia
neurons examined were activated.
Ia neurons. Therefore, a substantial number of Igneurons were possibly intermixed in Ia neurons identified in previous studies. In the rat, de Castro et al. [13] identified 57 Ianeurons in the NTS area ventrolateral to the solitary tract. Twelve of the 57 Ia neurons were excited from the ipsilateral vagus nerve at the latency 2.560.2 ms. Eleven Ia neurons projected to the spinal cord. Therefore, it is possible that some of these Ia neurons could be Ig
neurons.
We do not think that Igneurons exists only in the rat. In fact, some Ibneurons of the cats were supposed to receive RAR inputs in early studies [3,7,23]. This hypothesis has not necessarily been denied experimentally but seems to be
Fig. 5. Firing pattern of Ianeuron. Two neurons (A,B) are shown. Their
neglected at present after monosynaptic connections
be-firing is bound to the inspiratory phase and of augmenting type within the
tween SAR afferents and Ibneurons were proved [1,4]. In
phase: firing often starts in the preceding expiratory phase. Lung inflation
addition, some Ia neurons as well as Ib neurons of cats
and deflation do not specially affect their firing in many cases (A). In a
few neurons (B), firing is rather suppressed at lung deflation (Bc) and were shown to receive excitatory inputs during lung
facilitated at middle-range inflation of the lungs (Bb). These neurons inflation [12,22]. Although the results suggested the pres-(A,B) are clearly different from Iband Igneurons and designated as Ia
ence of SAR inputs, the possibility of RAR inputs could
neurons.
properties found in rat P-cells [26] have not been noticed receive inputs from deflation-sensitive receptors that are in cat P-cells. Therefore, Ig neurons in the cat may not reported to be sensitive only to deflation [5,32]. We think necessarily be the same as those in the rats. that this third mechanism is possible but with the
reserva-tion discussed in detail below.
4.2. Afferents to Ig neurons The previous study [5] classified the ‘deflation-sensitive’ and ‘mostly deflation-sensitive’ receptors as SARs, be-We think that Ig neurons receive inputs from RAR cause these receptors adapted slowly. Based on this afferents (Fig. 7) for following reasons. Firstly, Igneurons classification it was concluded that RAR afferents in the are orthodromically activated by electrical stimulation of rat were rare [5]. We are not convinced that such deflation-the vagus nerve at a low intensity which activates SAR and sensitive receptors [5,32] are different from RARs and RAR afferents. The latencies (2.160.21 ms) of the Ig form a distinct group. We rather believe that many of the neurons are similar to those of RAR-cells (2.060.23 ms) reported deflation-sensitive receptors [5,32], even if they [18] or of P-cells (2.160.19 ms) [26] obtained under the adapt slowly, belong to (at least a subcategory of) RARs same experimental conditions. This indicates that Ig which are also called lung irritant receptors and are not neurons receive direct inputs from low-threshold vagal rare in the rat [20,32]. The confusion arises when we afferents similarly to P-cells and RAR-cells. Secondly, Ig classify receptors based solely on adaptation rate or neurons are activated by lung deflation that characteristi- adaptation index [19,33]. As clearly mentioned [11,33], cally stimulates RARs [11]. In fact, responses of Ig adaptation index alone cannot identify receptor types; in neurons to lung collapse to atmospheric pressure and to particular, application of adaptation index to responses to lowered PEEP are quite similar to those of RAR afferents deflation is too problematic [33]. During the course of the and RAR-cells [17,18]. Taken together, if we specify one study (see also Refs. [16,17]), we also recorded from of the three accepted categories of pulmonary receptors deflation-sensitive vagal afferents which initially seemed (SARs, RARs, and C-fiber endings) [11], the most plaus- insensitive to hyperinflation (about 15–20 cmH O) of the2
ible receptors that provide excitatory inputs to Ig neurons lungs. However, they responded to more intense and rapid are RARs. A question then arises as to why Ig neurons lung inflation (about 30–40 cmH O), and we designated2
hardly respond to lung inflation which normally activates them as RAR afferents. Virtually all of the deflation-RARs. We think that either one or a combination of the sensitive and fast-conducting vagal afferents are classified following three mechanisms functions. Firstly, Ig neurons as RARs in our studies (see also Refs. [20,32]). SAR and are presumably inhibited by expiratory neurons during the RAR afferents thus-defined in the rat make two distinct expiratory phase, and such inhibitory inputs are intensified groups [17,25], and both firing property and location of during lung inflation (the Hering–Breuer reflex) [8]. their second-order relay neurons, P-cells and RAR-cells Secondly, Igneurons may be inhibited by P-cells similarly respectively, also make clearly different groups [16– to RAR-cells (Fig. 7). The direct inhibitory linkage from 18,25,26]. At present, therefore, we stick to the three P-cells to RAR-cells has been identified recently [16], and categories of pulmonary receptors (SARs, RARs, and C-in fact RAR-cells do not much respond to lung C-inflation as fiber endings) established in cats, rabbits and dogs until a compared with RAR afferents. Thirdly, Ig neurons might more detailed classification of pulmonary receptors is
established.
4.3. Functional significance
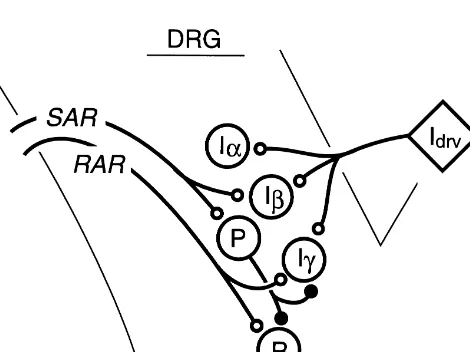
Until now we have no suitable answer to the question why the DRG includes Ia and Ib neurons. The finding of Ig neurons has made the situation more complex. The connections shown in Fig. 7 indicate that Ib neurons are involved in SAR-related pathways or pathways of the Hering–Breuer reflex whereas Ig neurons are involved in RAR-related pathways. However, SAR-related pathways and RAR-related pathways are not independent but work cooperatively via inhibitory connections arising from P-cells [16]. Furthermore, although it is not shown in Fig. 7, a close linkage between SLN inputs and Ianeurons should not be dismissed. Therefore, three types of DRG inspirat-ory neurons seem to integrate the central inspiratinspirat-ory drive and corresponding vagal inputs.
Fig. 7. Schematic representation of the results. P, P-cell; R, RAR-cell;
Since Ig neurons are activated by lung deflation it
Idrv, inspiratory drive to DRG inspiratory neurons. Connections from
K. Ezure, I. Tanaka / Brain Research 883 (2000) 22 –30 29 [6] A.L. Bianchi, Localisation et etude des neurones respiratoires
reflex [5,11,33]. We have proposed [16] the possible
bulbaires. Mise en jeu antidromatique par stimulation spinale ou
mechanisms of the Hering–Breuer deflation reflex based
vagale, J. Physiol. (Paris) 63 (1971) 5–40.
on both the inhibitory linkage from P-cells to RAR-cells [7] A.L. Bianchi, J.C. Barillot, Activity of medullary respiratory and the notion that RARs elicit inspiration-facilitating neurones during reflexes from the lungs in cats, Respir. Physiol. 25
(1975) 335–352.
reflexes to restore lung compliance to an optimal level
[8] M.I. Cohen, Neurogenesis of respiratory rhythm in the mammal,
[11,20,30]. Inflation of the lungs activates preferentially
Physiol. Rev. 59 (1979) 1105–1173.
SAR-related pathways by inhibiting RAR-related pathways [9] M.I. Cohen, How is respiratory rhythm generated?, Fed. Proc. 40 and elicits the Hering–Breuer inflation reflex. Whereas, (1981) 2372–2377.
[10] M.I. Cohen, J.L. Feldman, Discharge properties of dorsal medullary
deflation of the lungs which silences SARs activates
RAR-inspiratory neurons: relation to pulmonary afferent and phrenic
related pathways and elicits the Hering–Breuer deflation
efferent discharge, J. Neurophysiol. 51 (1984) 753–776.
reflex. This hypothesis is now expanded to include Ig
[11] H.M. Coleridge, J.C.G. Coleridge, Reflexes evoked from
trach-neurons in RAR-related pathways. In this respect, it is eobronchial tree and lungs, in: N.S. Cherniack, J.G. Widdicombe possible that spinal projections from Ig neurons target (Eds.), The Respiratory System, Handbook of Physiology, Vol. II, American Physiological Society, Washington, DC, 1986, pp. 395–
inspiratory neurons, such as phrenic motoneurons [21,27]
429.
or intercostal motoneurons [14] as shown previously for
[12] R.O. Davies, J. Metzler, D.A. Silage, A.I. Pack, Effects of lung
bulbospinal inspiratory neurons of the DRG. Furthermore, inflation on the excitability of dorsal respiratory group neurons, projections of Igneurons to the brainstem are also possible Brain Res. 366 (1986) 22–36.
[13] D. de Castro, J. Lipski, R. Kanjhan, Electrophysiological study of
since bulbospinal axons from some DRG inspiratory
dorsal respiratory neurons in the medulla oblongata of the rat, Brain
neurons have medullary collaterals [28].
Res. 639 (1994) 49–56.
If our hypothesis shown in Fig. 7 is correct, Igneurons [14] J. Duffin, J. Lipski, Monosynaptic excitation of thoracic are activated not only by lung deflation but also by motoneurones by inspiratory neurones of the nucleus tractus
solitarius in the cat, J. Physiol. (Lond.) 390 (1987) 415–431.
maneuvers which activate RARs. For instance, inhalation
[15] K. Ezure, M. Manabe, H. Yamada, Distribution of medullary
of lung irritant may activates Ig neurons, since RARs in
respiratory neurons in the rat, Brain Res. 455 (1988) 262–270.
the rat are certainly activated by lung irritant [20]. How- [16] K. Ezure, I. Tanaka, Lung inflation inhibits rapidly-adapting re-ever, such experiments have not been performed yet. At ceptor relay neurons in the rat, NeuroReport 11 (2000) 1709–1712. present, we are not certain whether RARs are homoge- [17] K. Ezure, I. Tanaka, M. Miyazaki, Inspiratory inhibition of pulmon-ary rapidly adapting receptor relay neurons in the rat, Neurosci. Lett.
neous or not. It is suggested that RARs which cause cough
258 (1988) 49–52.
and those which cause deep inspiration are different [34].
[18] K. Ezure, I. Tanaka, M. Miyazaki, Electrophysiological and
pharma-Findings of receptors which are sensitive mostly to lung cological analysis of synaptic inputs to pulmonary rapidly adapting deflation [5,32] further suggest that RARs are heteroge- receptor relay neurons in the rat, Exp. Brain Res. 128 (1999)
neous (see above). To understand the functional meaning 471–480.
[19] G.C. Knowlton, M.G. Larrabee, A unitary analysis of pulmonary
of Ig neurons, further detailed studies on pulmonary
volume receptors, Am. J. Physiol. 147 (1946) 100–114.
receptors and axonal projections of Igneurons are awaited.
[20] C.J. Lai, Y.R. Kou, Stimulation of pulmonary rapidly adapting receptors by inhaled wood smoke in rats, J. Physiol. (Lond.) 508 (1998) 597–607.
[21] J. Lipski, L. Kubin, J. Jodokowski, Synaptic action of Rbneurons
Acknowledgements
on phrenic motoneurons studied with spike-triggered averaging, Brain Res. 288 (1983) 105–118.
The authors thank Ms Yasuko Kishimoto for her expert [22] J. Lipski, A. Trzebski, L. Kubin, Excitability changes of dorsal technical assistance in histology. inspiratory neurons during lung inflations as studied by measure-ment of antidromic invasion latencies, Brain Res. 161 (1979) 25– 38.
[23] P.L. Mariono, R.O. Davies, A.L. Pack, The responses of Ibcells to
References increases in the rate of lung inflation, Brain Res. 219 (1981)
289–305.
[1] D.B. Averill, W.E. Cameron, A.J. Berger, Monosynaptic excitation [24] R.A. Mitchel, A.J. Berger, Neural regulation of respiration, Am. of dorsal medullary respiratory neurons by slowly adapting pulmon- Rev. Respir. Dis. 111 (1975) 206–224.
ary stretch receptors, J. Neurophysiol. 52 (1984) 771–785. [25] M. Miyazaki, A. Arata, I. Tanaka, K. Ezure, Activity of rat pump [2] R.V. Baumgarten, E. Kanzow, The interaction of two types of neurons is modulated with central respiratory rhythm, Neurosci.
inspiratory neurons in the region of the tractus solitarius of the cat, Lett. 249 (1998) 61–64.
Arch. Ital. Biol. 96 (1958) 361–373. [26] M. Miyazaki, I. Tanaka, K. Ezure, Excitatory and inhibitory [3] A.J. Berger, Dorsal respiratory group neurons in the medulla of cat: synaptic inputs shape the discharge pattern of pump neurons of the spinal projections, responses to lung inflation and superior laryngeal nucleus tractus solitarii in the rat, Exp. Brain Res. 129 (1999) nerve stimulation, Brain Res. 135 (1977) 231–254. 191–200.
[4] A.J. Beger, T. Dick, Connectivity of slowly adapting pulmonary [27] R. Monteau, M. Khatib, G. Hilaire, Central determination of stretch receptors with dorsal medullary respiratory neurons, J. recruitment order: intracellullar study of phrenic motoneurons, Neurophysiol. 58 (1987) 1259–1272. Neurosci. Lett. 56 (1985) 341–346.
[29] G. Paxinos, C. Watson, The Rat Brain in Stereotaxic Coordinates, types of pulmonary receptors responding to collapse, inflation, and Academic Press, New York, 1986. deflation of the lungs, Exp. Neurol. 92 (1986) 541–552.
[30] T.E. Pisarri, A. Jonzon, J.C.G. Coleridge, H.M. Coleridge, Rapidly [33] J.G. Widdicombe, Receptors in the trachea and bronchi of the cat, J. adapting receptors monitor lung compliance in spontaneously Physiol. (Lond.) 123 (1954) 71–104.
breathing dogs, Am. J. Physiol. 68 (1990) 1997–2005. [34] J.G. Widdicombe, Afferent receptors in the airways and cough, [31] K. Saether, G. Hilaire, R. Monteau, Dorsal and ventral respiratory Respir. Physiol. 114 (1998) 5–15.