IV. HASlL
DAN
PEMBAHASAN
4.1. Uji Ketahanan Diferensial Beberapa Klon Karet terhadap
Beberapa
Isolat
Cendawan
C.
cassiicola
4.1.1. Ketahanan klon karet terhadap
PGDC
Tingkat ketahanan klon karet terhadap penyakit gugur daun corynespora tercennin dari tingkat keparahan penyakit yang terjadi. Makin rendah tingkat keparahan penyakit, makin tinggi tingkat ketahanan klon terhadap PGDC; sebaliknya makin tinggi tingkat keparahan penyakit, makin rendah tingkat ketahanan klon terhadap penyakit tersebut.
Data keparahan PGDC pada tiap periode pengamatan ditampilkan dalam Lampiran 5. Dengan mempergunakan data keparahan penyakit pada pengamatan terakhlr, dlbuat histogram respon ketahanan klon karet terhadap penyakit tersebut (Gambar 4.1). Dari histogram tersebut dapat diketahui bahwa respons ketahanan antar klon karet berbeda-beda terhadap isolat Ccl, Ccz, dan Cc3. Secara umum dapat dikatakan bahwa respons ketahanan klon karet PB 260, BPM 1, dan RRIC 100 paling baik dibanding dengan klon lain yang diuji, karena keparahan penyakit yang terjadi tergolong paling rendah. Sebaliknya klon- klon PPN 2444, RRIC 103, RRIM 725, dan PPN 2447 memperlihatkan tingkat keparahan penyakit paling tinggi, sehingga klon-klon tersebut dapat dimasukkan dalam kelompok klon karet yang paling rentan terhadap PGDC.
Keparahan penyakit gugur daun corynespora yang terjadi pada pengujian ini terbukti dipengaruhi oleh daerah asal isolat C. cassiicolu yang dipergunakan. Secara umum dapat dikatakan bahwa isolat C. cassiicola yang berasal dari Sumatra Selatan ( C q j menyebabkan terjadinya keparahan penyakit paling tinggi, yang menunjukkan bahwa isolat Cc3 tersebut mempunyai tingkat virulensi paling tinggi dibanding dua isolat yang lain.
Isolat C. cussiicolu yang berasal dari Jawa Tengah (Cc?) memperlihatkan tingkat virulensi paling rendah, sedangkan isolat dari Kalimantan Selatan (Ccl) mempunyai tingkat virulensi di antara kedua isolat yang disebut sebelumnya. Hasil ini mendukung pendapat sebelumnya tentang adanya ras C. cassiicola yang beradaptasi dengan kondisi geografis daerah asalnya (Situmorang et al. 1996, Rodesuchit & Kajornchalyakul 1996).
Meskipun secara umum urutan tingkat virulensi isolat C.cassiicola adalah Cc3, Cc1, dan Ccz, akan tetapi tingkat keparahan penyakit yang terjadi pada tiap klon karet yang diuji tidak selalu sesuai dengan urutan tersebut. Sebagai contoh, isolat Cc3 menghasilkan keparahan penyakit paling tinggi pada hampir semua klon karet yang diuji, kecuali pada klon karet PR 261, PR 300, dan RRIM 712. Pada klon karet PR 261 dan PR 300, keparahan penyakit tertinggi disebabkan oleh isolat Ccl, sedangkan pada klon karet RRIM 7 12, isolat Ccl dan Cc3 menyebabkan tingkat keparahan penyakit yang sama. Inokulasi dengan isolat Cc, dan Ccz juga menghasilkan keparahan penyakit yang sama pada klon karet PB 260. Disamping itu isolat paling lemah Cc? menimbulkan tingkat penyakit paling rendah pada sebagian besar klon karet yang diuji, kecuali pada klon AVROS 2037, PPN 2447, dan RRIM 725. Pada ketiga klon karet tersebut isolat Cc2 menghasilkan tingkat penyakit lebih parah dibandingkan dengan isolat Ccl. Fenomena seperti diterangkan di atas menunjukkan kemungkinan adanya interaksi antara genotipe tanaman inang dan patogen (Oka 1993, Van der Plank 1968). Bila ha1 ini benar maka sifat ketahanan tanaman karet terhadap PGDC kemungkinan besar dikendalikan secara vertikal.
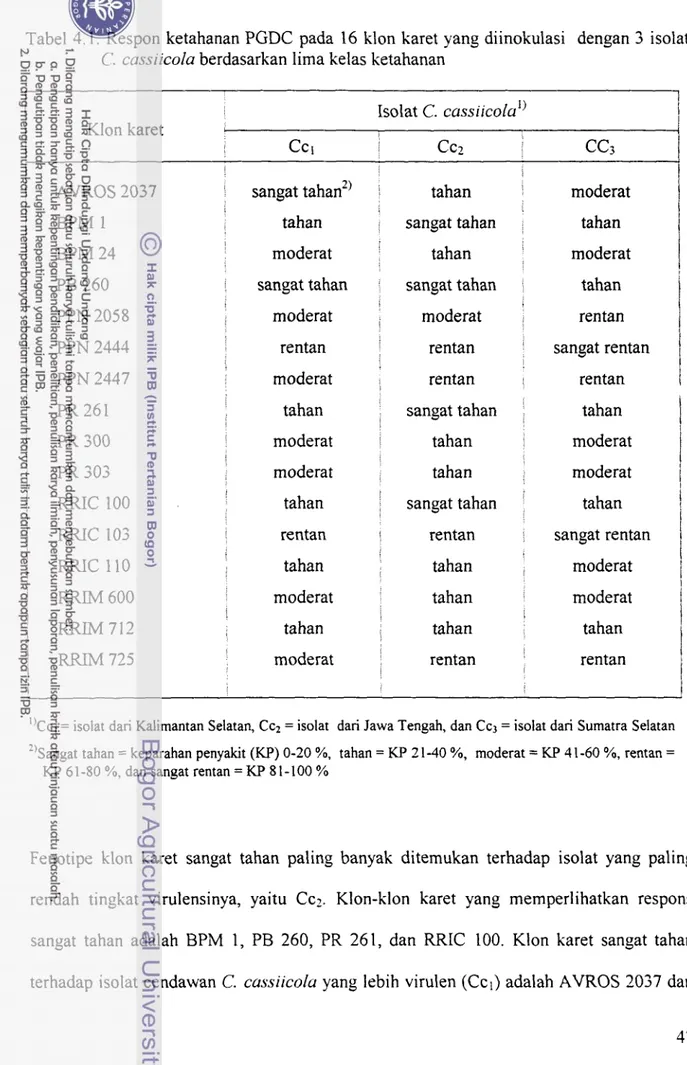
Pengelompokan klon karet berdasar lima kelas ketahanan disajikan dalam Tabel 4.1. Tabel tersebut dapat memperjelas uraian sebelumnya tentang adanya perbedaan
Tabel 4.1. Respon ketahanan PGDC pada 16 klon karet yang diinokulasi dengan 3 isolat
C. cassiicola berdasarkan lima kelas ketahanan
ip-
[solat C. cussiicolul)i
!
I Klon karet
I
I
! c c 1 Cc2 CC;
I
1
AVROS 2037 sangat tahan2' tahan mode rati
1
BPM1 tahan sangat tahan tahanBPM24 moderat tahan moderat
1
I
1 PPN 2058
I
I
PPN 2444i
sangat tahan sangat tahan tahan
moderat moderat rentan
rentan rentan sangat rentan
I
i
PPN 2447 moderat rentan rentan1
i
PR261 tahan sangat tahan tahan
i moderat moderat tahan tahan tahan sangat tahan moderat moderat tahan
1
RRIC103 rentan rentan sangat rentan1
i t
/
RRICllO tahan tahan moderat1
I 1
1
RRLM 600 moderat tahan moderat I IRRIM712
I tahan tahan tahan
I
i
1
RRIM 725 moderat rentan rentan II
i"CC, = isolat dari Kalimantan Selatan, Cc2 = isolat dari Jawa Tengah, dan Cc3 = isolat dari Sumatra Selatan *'sangat tahan = keparahan penyakit (KF') 0-20 %, tahan = KP 21-40 %, moderat = KF' 41-60 %, rentan =
KP 61-80 %, dan sangat rentan = KP 8 1 - 100 %
Fenotipe klon karet sangat tahan paling banyak ditemukan terhadap isolat yang paling rendah tingkat virulensinya, yaitu Ccz. Klon-klon karet yang memperlihatkan respons sangat tahan adalah BPM 1, PB 260, PR 261, dan RRIC 100. Klon karet sangat tahan terhadap isolat cendawan C. cussiicolu yang lebih virulen (Ccl) adalah AVROS 2037 dan
PB 260, sedangkan terhadap isolat paling virulen (Cc;) tidak ditemukan klon karet sangat tahan, tetapi ditemukan klon karet yang sangat rentan, yang tidak ditemukan terhadap isolat Ccl dan Ccz.
4.1.2. Analisis perkembangan
PGDC
Dengan data pengamatan gejala penyakit gugur daun corynespora pada minggu ke- 1 (t,) sampai dengan minggu ke-4 (tj) (Lampiran 5) dibuat grafik perkembangan penyakit seperti ditampilkan pada Gambar 4.2. Hampir semua grafik perkembangan penyakit pada klon karet yang diuji mengikuti pola sigmoid. Kurva sigmoid diawali dengan lereng landai (t, - t2) yang menunjukkan laju perkembangan penyakit lambat, diikuti lereng tajam (tz - t3) yang menunujukkan perkembangan penyakit meningkat secara eksponensial, serta dialduri dengan perkembangan penyakit yang melambat (t3 - tj). Grafik pola sigmoid mengikuti fase perkembangan penyakit yang umum terjadi di alam (Nelson & MacKenzie
1973).
Gambar 4.2 memperlihatkan pola perkembangan PGDC yang konsisten, yaitu apabila tingkat keparahan penyakit pada pengamatan minggu pertama (tl) tinggi, maka tingkat keparahan penyakit pada pengamatan minggu terakhir (b) juga tinggi (PPN 2444, PPN 2447, RRIC 103, dan RRIC 725). Demikian pula apabila tingkat keparahan penyakit pada tl rendah, maka tingkat keparahan penyakit pada
t
juga rendah (PB 260, RRIC 100, dan BPM 1). Oleh karena itu penilaian tingkat ketahanan tanaman terhadap penyakit dapat dilakukan baik pada saat awal maupun akhir perkembangan penyakit (Reynolds & Cunfer90- 70- (001
0 4 PRIzu4mww7 m R W W
WNaUI eQ1 RYCtOBllRIlllS IIADtm
-)
a':
RwifB 7u 4 8 0 4 a; A,
&
mmrn 2U7 PPNZlCl1
i
1 mpss1;
/A
I-MO m2m mm%-lo Y m, nmcqmj
sod1
-1mSOS -@so PRSOOPRa PPNaD6(1 RRY-I
= I AV zwQ
-7WmSOD --a) "'/
P R m m -1-1 a1 10=':
7;:
wtL
0 9 w O W 1 1 J 1 w J 0 1 2 3 47"
1-
2 3 U 1 2 3 4 4 F.8- ~utsddahkrokulahli(t) ww-inolaJld(t) (b) isotit Cc2(a) idat
eel
')Gmhu4.2.
C k r a f i k ~ P a D C p a d a l 6 $ l o n k a r e t y a a g d i ~ @ ~ ~ ~ t i C . - i
-
(all,
J a w
T
&
(Ccz),-
?3el-
(a)
s
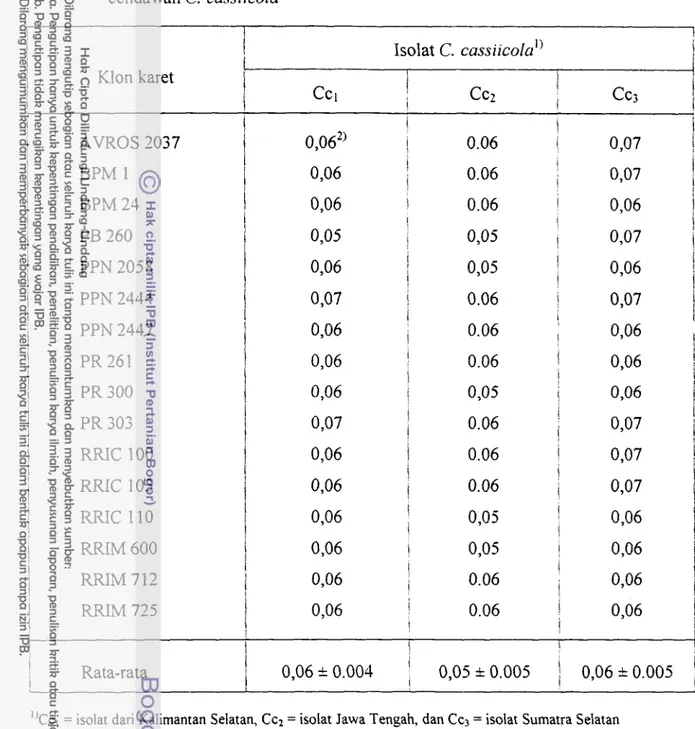
Perbandingan prakiraan laju perkembangan penyakit (r) yang ditunjukkan dengan kemiringan grafik perkembangan penyakit pada tiap klon karet yang diuji relatif sama antara klon yang satu dengan lainnya, baik yang disebabkan oleh isolat Ccl, Ccz, maupun CC; (Gambar 4.2 dan Tabel 4.2). Nilai r berkisar antara 0,05 - 0,07 unitlhari dengan r rata- rata pada Ccl 0,06
*
0.004 unithari, pada Ccz 0,05 k 0.005 unithari, sedangkan pada Cc; 0,06*
0.005 unithari. Dengan nilai r yang relatif sama maka apabila dibuat linier, grafik perkembangan penyakit gugur daun corynespora masing-masing klon karet pada setiap isolat yang diujikan akan bersifat sejajar antara klon satu dengan klon karet lainnya (Lampiran 6).Pola grafik perkembangan penyakit antar klon atau varietas tanaman dapat dipergunakan untuk mempelajari sifat ketahanan tanaman terhadap suatu penyakit. Apabila laju infeksi antara genotipe tahan dan genotipe rentan berbeda secara nyata, maka kemungkinan ketahanan tersebut bersifat horizontal. Laju infeksi pada genotipe yang tahan lebih rendah, sedangkan laju infeksi pada genotipe rentan lebih tinggi. Pada ketahanan yang bersifat vertikal, laju infeksi antar genotipe sama sehingga grafik perkembangan penyakit antara genotipe tahan dan rentan mempunyai lcemiringan yang sama pula (Nelson & MacKenzie 1973, Van der Plank 1968). Dengan mempelajari grafik perkembangan penyakit pada Gambar 4.2 dan Lampiran 6 dapat diduga bahwa sifat ketahanan tanaman karet terhadap PGDC bersifat vertikal, yaitu ketahanan tanaman yang dikendalikan oleh sejumlah kecil gen utama.
Tabel 4.2. Laju infeksi (r) PGDC pada 16 klon karet yang diinokulasi dengan 3 isolat cendawan C. cassiicola i
I
Isolat C. cassiicolc~') ! Ii
Klon karetI
i
I I Cc 1 iI
1 I I Ccz I Cc3I
I II
1 I i 11
AVROS 2037!
0,06*' ! 0.06 1 0,071
II
! , I BPM 1 0,06 I 0.06 0,07I
I
I
I
I BPM 241
0,06 0.06 ! i 0,06i
PB 260 PPN 2058 PPN 2444 PPN 2447 PR 261 PR 300 RRIC 100 RRIC 103 RRIC 110" C C ~ = isolat dari Kalimantan Selatan, CC.Z = isolat Jawa Tengah, dan C q = isolat Sumatra Selatan
2kaju infeksi ( r ) dinyatakan dalarn unithari, dihitung dari minggu kke-1 sarnpai dengan rninggu ke-4 setelah inoL-lasi.
Pada spesies tanaman yang berketahanan penyakit vertikal, adanya perubahan ras fisiologi patogen tidak mengubah laju infeksi, sehingga kemiringan grafik perkembangan penyakit tidak berubah. Bila ketahanan tanaman inang terpatahkan, maka grafik perkembangan penyakit akan dimulai pada tingkat penyakit
&
yang lebih tinggi dan berakhir pada tingkat penyakit Xt yang lebih tingg pula (Nelson & MacKenzie 1973). Dengan demikian besaran laju infeksi ( r ) tidak dapat dipergunakan untuk membedakan tingkat ketahanan tanaman terhadap suatu penyakit.Pada kasus di atas, tingkat ketahanan antar tanaman inang dapat dibedakan dengan lebih mudah apabila menggunakan perbandingan area dibawah grafik perkembangan penyakit (area under the disease progress curve, AUDPC). Tanaman yang tahan penyakit mempunyai luas AUDPC lebih kecil, sebaliknya tanaman rentan mempunyai AUDPC lebih besar (Reynolds & Neher 1997, Reynolds & Cunfer 1997). Pada Gambar 4.2 terlihat bahwa grafik klon-klon PB 260, BPM 1, dan RRIC 100 terletak di bagian bawah, sehingga secara kualitatif luas AUDPC klon-klon tersebut lebih kecil. Jadi dapat disimpulkan bahwa klon-klon tersebut lebih tahan terhadap PGDC dibanding dengan klon-klon lainnya dalam grafik tersebut. Sebaliknya klon-klon PPN 2444, RRIC 103, PPN 2447, dan RRIM 725 tergolong klon paling rentan terhadap PGDC karena secara kualitatif mempunyai luas AUDPC lebih besar. Besarnya AUDPC mencerminkan akumulasi kerusakan tanaman
inang oleh suatu penyakit, yang dipengaruhi oleh laju infeksi ( r ), periode waktu serangan ( t ) dan intensitas penyakit pada akhir pengamatan ( X, ) (Reynolds & Cunfer 1997).
4.2.
Analisis Genetik Pewarisan Sifat Ketahanan Tanaman Karet
terhadap Penyakit Gugur Daun Corynespora
4.2.1. Segregasi
Z2
keturunan tetua
TJIR
1
x PR
107
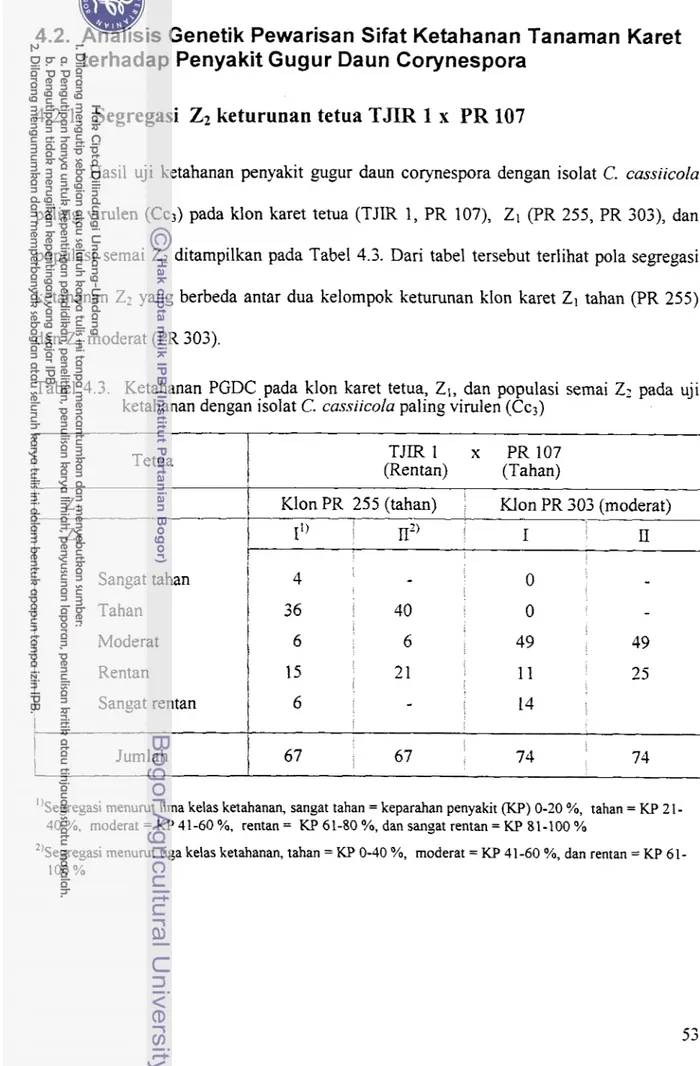
Hasil uji ketahanan penyakit gugur daun corynespora dengan isolat C. cassiicola
paling virulen (Cc3) pada klon karet tetua (TJIR 1, PR 107), Z1 (PR 255, PR 303), dan populasi semai Z2 ditampilkan pada Tabel 4.3. Dari tabel tersebut terlihat pola segregasi ketahanan Z2 yang berbeda antar dua kelompok keturunan klon karet ZI tahan (PR 255) dan Z1 moderat (PR 303).
Tabel 4.3. Ketahanan PGDC pada klon karet tetua, Z1, dan populasi semai ZZ pada uji ketahanan dengan isolat C. cassiicola paling virulen (Cc;)
Tetua (Rentan) TJIR1 x (Tahan) PR107
/
Klon PR 255 (tahan) Klon PR 303 (moderat)1
1
I" n2j III
i
I
Sangat tahanI
-
0-
I
i
1
Tahan1
36 40 0-
i
i
6 i Moderat 49 49i
Rentan Sangat rentan 1I
Jumlah 67 67 74 74i
" ~ e g r e ~ a s i menurut lima kelas ketahanan, sangat tahan = keparahan penyakit (KP) 0-20 %, tahan = KP 2 1
-
40 %, moderat = KP 4 1-60 %, rentan = KP 6 1-80 %, dan sangat rentan = KP 81-1 00 %2'~egregasi menurut tiga kelas ketahanan, tahan = KP 0-40 %, moderat = KP 41-60 %, dan rentan = KP 61- 100 %
Pola segregasi menurut lima kelas ketahanan seperti tertulis pada lajur I tersebut tidak sesuai dengan pola segregasi dua gen epistasis dengan masing-masing dua alel. Akan tetapi bila pengelompokan kelas ketahanan diubah menjadi tiga katagori seperti tersaji dalam lajur I1 dalam tabel yang sama, yaitu : (a) tahan
-
yang merupakan gabungan fenotipe sangat tahan dan tahan, (b) moderat, dan (c) rentan- yang merupakan gabungan
fenotipe rentan dan sangat rentan, maka kelas fenotipe ini mendekati model dihibrid untuk suatu sifat yang dikontrol oleh dua gen secara epistasis. Untuk memastikan kesesuaian data yang diamati dengan model yang diharapkan, tiap populasi Z2 dari Z1 yang berbeda dianalisis dan diuji dengan uji Khi kuadrat.4.2.1.1. Populasi
Z2
dari klon
Z1
PR 255
Pola segregasi Zz dari keturunan tanaman klon karet Z1 PR 255 memperlihatkan nisbah 40 : 6 : 21 untuk sifat tahan, moderat, dan rentan (Tabel 4.4). Pola segregasi ini sesuai dengan model nisbah fenotipe 9 : 3 : 4, yang menunjukkan pola segregasi fenotipe yang dikendalikan oleh dua pasang gen yang berinteraksi secara epistasis resesif (Griffiths 1993). Uji Khi kuadrat diperoleh angka Khi kuadrat htung (Xzu,,) = 4,05. Angka ini
lebih kecil dari XZlabl j % dengan derajat bebas db = 2 yang besamya 5,99. Dengan
demikian perbandingan fenotipe 9 : 3 : 4 mengikuti pola segregasi yang sesuai dengan model kalau genotipe Z I heterozigot untuk dua gen (AaBb) yang bersifat epistasis. Jadi sifat ketahanan tanaman karet terhadap PGDC kemungkinan dikendalikan oleh dua pasang gen utama yang berinteraksi secara epistasis.
Perbandingan fenotipe 9 : 3 : 4 pada populasi Z2 menunjukkan segregasi genotipe di Zz menjadi 16 genotipe, yang berasal dari pembuahan antar 4 macam kombinasi genotipe gamet yang membawa ale1 berbeda. Pembentukan gamet menjadi 4 macam 5 4
kombinasi genotipe hanya dapat tejadi bila genotipe Z1 tersusun oleh dua pasang gen yang bersifat heterosigot. Tanaman genotipe AaBb yang dibiarkan menyerbuk sendiri atau disilangkan dengan pohon heterozigot yang sama, pada saat pembentukan gamet akan menghasilkan kombinasi genotipe gamet betina dan gamet jantan dengan proporsi kombinasi genotipe masing-masing ?4 AB, % Ab, % aB, dan '/4 ab. Populasi genotipe
Zz
merupakan zigot hasil pembuahan acak antar kelompok gamet betina dan kelompok gamet jantan tersebut (Tabel 4 . 9 , sehingga model genotipe untuk tiap kelas ketahanan yang dipelajari serta frehensi kemunculannya seperti dalam Tabel 4.6.Tabel 4.4. Uji U u kuadrat populasi tanaman karet Z2 yang berasal dari klon karet Z1 PR 255
I !
/
~ e n o t i ~ e "j
Jml. Teramatii
Jurnlahdiharapkanj
X 2 hitung 2)1
! I f Tahan 40 i 3 8 i I ! I 0,11 6 12
1
Moderat ! I ! 3 ,OO1
! Rentan 2 1 17 I Jumlah 6 7 67 4,05') Tahan = keparahan penyakit (KP) 0-40 %, moderat = KP 41-60 %, dan rentan = KP 61-100 % 2) X 2
tabel 5 3.a d b 2 = 5799
Tabel 4.5. Model persilangan antar pohon klon karet Z I PR 255 bergenotipe AaBb
1
Garnet % AB % Ab % aB % ab1
1
i
G A B/
1/16 AABB 1/16 AAOb 1/16 AaBB 1/16 AaBb1
I
I 'A Ab
1
1116 AABb 1/16 AAbb 1/16 AaBb 1116 AabbI
II
1
% a B 1/16 AaBB 1/16 AaBb 1/16 aaBB 1/16 aaBbI
1
?4 ab 1/16 AaBb 1/16 Aabb 1/16 aaBb 1/16 aabb1
Tabel 4.6. Model genotipe Z2 keturunan klon karet Z1 PR 255 untuk tiap kelas ketahanan terhadap PGDC
I i
1
Fenotipe Jumlah individu Genotipe Frekuensi i! Tahan 40 AABB AABb AaBB AaBb Moderat 6 AAbb 1/16 I I Aab b Rentan 2 1 aaBB I aaBb 2/16
i
aabb 1/16 I I i i I Jumlah 67-
1 I I IJadi klon PR 255 bergenotipe heterozigot AaBb tahan terhadap PGDC. Ale1 A dominan terhadap a, serta ale1 B dominan terhadap b. Fenotipe tahan terjadi bila susunan genotipe minimum mengandung masing-masing satu ale1 dominan dari tiap gen yaitu : AABB, AABb, AaBB, atau AaBb. Hasil persilangan di atas, total frekuensi kemunculan fenotipe tahan adalah 9/16. Di dalam populasi Zz tersebut, fenotipe tahan yang diamati 40 individu dari total 67 individu yang diuji. Fenotipe moderat berdasar model genetik tersebut berfrekuensi 3/16 yaitu Aabb dan Aabb, sedangkan yang diamati dalam populasi
Zz
adalah 6 individu tanaman.Nisbah fenotipe 9 : 3 : 4 menunjukkan interaksi gen epistasis yang bersifat alel resesif a pada keadaan homozigot aa menekan ekspresi alel B dan b yang terletak pada lokus yang lain (Griffiths et ul. 1993), sehingga tanaman yang bergenotipe aa -- (aaBB, aaBb, dan aabb) akan selalu bersifat rentan terhadap PGDC. Proporsi tanaman yang
memperlihatkan sifat rentan adalah 4116, di dalam populasi Z2 klon PR 255 terdapat 21 individu tanaman rentan. Model-model genotipe tersebut di atas hams berlah pula untuk populasi segregasi keturunan pohon Z1 klon karet yang berfenotipe ketahanan moderat terhadap PGDC berikut ini.
4.2.1.2. Populasi
Z2
dari klon
karet
Z1
PR 303
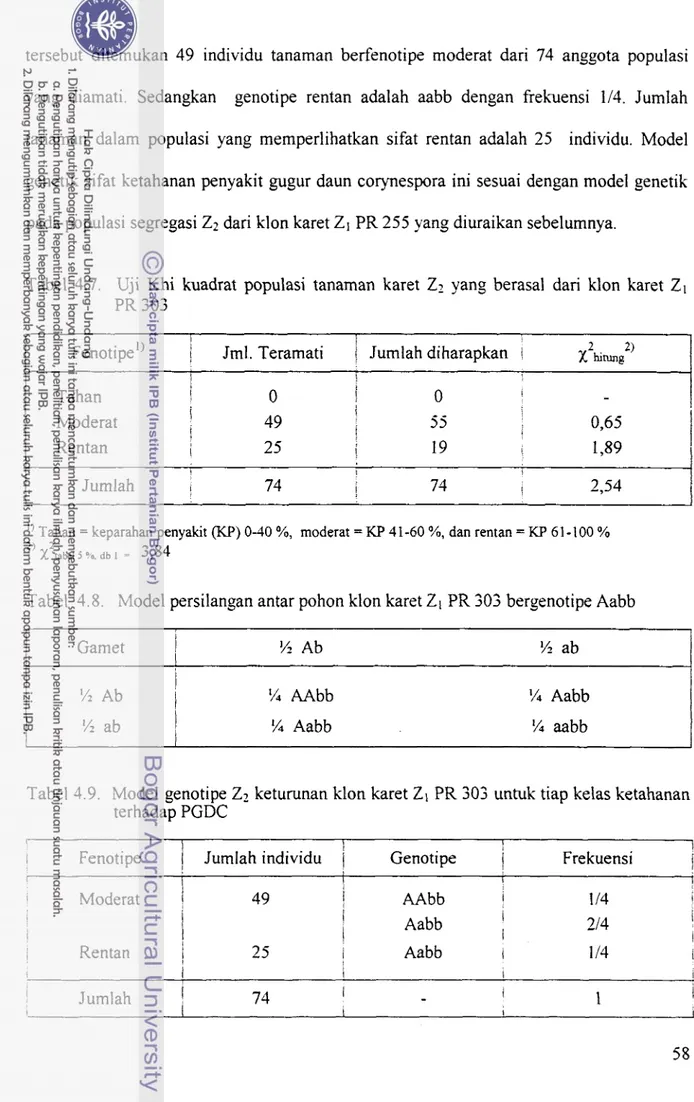
Klon karet Z1 PR 303 bersifat moderat terhadap PGDC dan kemungkinan genotipenya AAbb atau Aabb. Uji ketahanan semai karet populasi Z2 keturunan pohon Z l klon PR 303 terhadap PGDC menghasilkan dua kelompok ketahanan, yaitu kelompok moderat (49 individu) dan rentan (25 individu). Dengan hipotesis nisbah yang diharapkan 3 : 1 untuk sifat moderat : sifat rentan, menghasilkan uji Khi kuadrat dengan X'u,, = 2,54 lebih kecil dan X',hl j %, db 1 = 3,84 (Tabel 4.7), berarti bahwa hipotesis nisbah fenotipe 3 : 1 pada populasi Z2 tersebut dapat diterima. Jadi model genotipe sifat ketahanan Z1 klon karet PR 303 hams Aabb yang mengekspresikan sifat moderat dan bukan AAbb. Genotipe Aabb menghasilkan dua genotipe sel gamet Ab dan ab, sedangkan genotipe AAbb hanya menghasilkan satu macam genotipe gamet Ab. Kemungkinan genotipe Z2 keturunan klon karet ZI PR 303 dilaporkan dalam Tabel 4.8, sedangkan model genetik Zz untuk tiap kelas ketahanan serta frekuensi kemunculannya ditampilkan pada Tabel 4.9.
Dalam populasi segregasi Z2 pada Tabel 4.8 dan Tabel 4.9, genotipe yang bersifat tahan terhadap PGDC tidak mungkin dihasilkan. Genotipe yang mungkin ada hanya untuk fenotipe moderat dan fenotipe rentan. Model genotipe untuk individu moderat adalah AAbb dan Aabb dengan frekuensi kemunculan sebesar 314, dan di dalam populasi Z2
tersebut ditemukan 49 individu tanaman berfenotipe moderat dari 74 anggota populasi yang diamati. Sedangkan genotipe rentan adalah aabb dengan fiekuensi 114. Jumlah tanaman dalam populasi yang mernperlihatkan sifat rentan adalah 25 individu. Model genetik sifat ketahanan penyakit gugur daun corynespora ini sesuai dengan model genetik pada populasi segregasi Z2 dari klon karet Z1 PR 255 yang diuraikan sebelumnya.
Tabel 4.7. Uji Khi kuadrat populasi tanaman karet
Zz
yang berasal dari klon karet Z1 PR 303!
~ e n o t i ~ e " j Jml. Teramati i Jumlah diharapkan ! X 2 iutung 2 )
I I Tahan 1 0 I 0 I
-
I Moderat I ! 49 j 55 0,65 Rentan I 25 I 19 I 1,89 I I Jumlah I 74 ! 74 2,54''
Tahan = keparahan penyakit (KP) 0-40 %, moderat = KP 4 1-60 %, dan rentan = KP 6 1-
100 % 2' x ~ ~ , M j Y& tib = 3,84Tabel 4.8. Model persilangan antar pohon klon karet ZI PR 303 bergenotipe Aabb
1
Gamet
1
I/j Ab ?4 ab'/z Ab
i
I
'/4 AAbb '/4 Aabb'/t ab
1
!4 Aabb !4 aabbTabel 4.9. Model genotipe Z2 keturunan klon karet 21 PR 303 untuk tiap kelas ketahanan terhadap PGDC
1 \ I 1
i Fenotipe Jumlah individu
i
Genotipe1
Frekuensi I I i ModeratI
Rentan I II
AAbb 1 Aabb I 214 i I I Aabb I I 14 I 1 I Jumlah 1 74 I-
I 1 I4.2.2. Segregasi
Zz
keturunan dari tetua
yang
lain
Untuk memperhat model genetik tersebut di atas, maka perlu dianalisis segregasi pada populasi Z2 yang berasal dari tetua klon karet yang lainnya. Untuk keperluan tersebut digunakan populasi segregasi Zz yang berasal dari pohon klon karet Z1 PB 260, RRIC 100 dan RRIM 712. Ketiga klon tersebut masing-masing berasal dari tetua yang berbeda. Dalam pengujian ketahanan PGDC dengan isolat Cc3, klon-klon unggul anjuran tersebut tennasuk dalam katagori tahan. Sedangkan pengujian terhadap populasi semai Zz keturunan klon-klon Z1 tersebut menghasilkan segregasi ketahanan seperti tersaji pada Tabel 4.10. Dengan alasan yang sama seperti sebelumnya, pembagian segregasi Zz kc dalam lima kelas ketahanan (lajur I) pada tabel tersebut kurang sesuai, sehingga perlu kelompokkan ke dalam tiga kelas ketahanan (lajur 11). Selanjutnya analisis Khi kuadrat dilakukan untuk menguji nisbah fenotipe yang te rjadi (Tabel 4.1 1).
Tabel 4.10. Pola segregasi ketahanan tanaman karet terhadap PGDC pada tiga kelornpok populasi Zz keturunan klon karet Z1 PB 260, RRIC 100, dan RRLM 712.
I
I
Sangat tahan Kelas ketahanan Populasi Z2 I Tahan ModeratSumber populasi (pohon Z1)
PB 260 (tahan)
i
RRIC 100 (tahan)I
RRIM 7 I2 (tahan)Rentan 15 19 ! 12 14 10
Sangat rentan 4 2
i f I 16
''
Segregasi menurut lima keias ketahanan, sangat tahan = keparahan penyakit (KP) 0-20 %, tahan = KP 21-40%, moderat = KP 4 1-60 %, rentan = KP 61-80 %, dan sangat rentan = KP 8 1
-
100 %Tabel 4.1 1. Uji IS~I kuadrat populasi Z2 keturunan klon karet Z1 PB 260, RRIC 100, dan RRIM 712 Tahan Moderat Rentan Kelas ketahanan I populasi
zz"
I I 11
Jumlah tanaman 67 I 46 ! 751
t Sumber populasi (pohon Z 1)1
PB 260
/
RRIClOO RRIM7121
Nisbah harapan 7
x-hitung
tabel 5 %
''
Tahan = keparahan penyakit (KP) 0-40 %, moderat = KP 41-60 %, dan rentan = KP 61-100 %4.2.2.1. Populasi
Z2 dari
klon karet
Z1
PB 260
Populasi segregasi Z2 yang berasal dari keturunan klon karet Z1 PB 260 menunjukkan nisbah jumlah individu tahan : individu rentan adalah 48 : 19 dari total 67 nomor individu yang diuji. Sedangkan fenotipe moderat tidak ditemukan dalam populasi ini. Dengan model perbandingan fenotipe yang diharapkan adalah 3 : 1 untuk sifat tahan dan sifat rentan, hasil analisis Khi kuadrat diperoleh nilai X2u,g = OJ2, jauh lebih kecil dari X2,kl j %, J,, 2 = 3,84 (Tabel 4.1 1). Jadi populasi
Zi
hanya mungkin dihasilkan dari tanaman karet Z1 yang bergenotipe AaBB agar diperoleh perbandingan fenotipe Zz tahan : rentan = 3 : I (Tabel 4.12 dan 4.13).Tabel 4.12. Model persilangan antar pohon klon karet Z1 PB 260 yang bergenotipe AaBB
I
Gamet i !h AB % aB , II
!
Ii
% AB i % AABB % AaBBi
I % aB I 1 % AaBB 1/4 aaBBTabel 4.13. Model genotipe Z 2 keturunan dari pohon klon karet Z1 PB 260 untuk tiap kelas ketahanan terhadap PGDC
t I
I
! Fenotipe Jumlah individu Genotipe Frekuensi
I
I I I I Tahan 4 8 AABB 114
1
i AaBB 214 Ii
i
I
Moderat 0-
I II
i
Rentan 19 aaBB 1 I 4 tt
I J1
Jumlah 6 7-
1I
I IDari Tabel 4.12 dan Tabel 4.13 diketahui bahwa individu populasi Zz terdiri dari tiga macam genotipe, yaitu : AABB, AaBB untuk fenotipe tahan, dan aaBB untuk fenotipe rentan. Kelompok individu tahan mempunyai kemungkinan frekuensi 3/4, yang dalam penelitian ini berjumlah 48 individu tanaman. Sedangkan kelompok fenotipe rentan mempunyai kemungkinan frekuensi 114, yang diamati 19 individu tanaman dari 67 nomor individu yang diuji. Jadi genotipe klon karet PB 260 adalah AaBB yang bersifat tahan terhadap PGDC.
4.2.2.2. Populasi
Z2
dari klon
karet
Z1
RRIC
100
Jumlah individu dalam populasi segregasi Zz yang berasal dari tanaman Z1 klon karet RRIC 100 paling kecil dibanding populasi lainnya. Banyak benih pada populasi ini tidak dapat berkecambah karena buah karet dipanen belurn cukup masak. Dari sejumlah semai yang dapat ditumbuhkan, 46 batang diantaranya dapat diuji tingkat ketahanannya terhadap penyakit gugur daun corynespora. Pengujian tersebut rnenghasilkan data : 28 individu semai memperlihatkan sifat tahan, 4 individu tergolong moderat dan 14 individu lainnya termasuk katagori rentan. Hasil uji Khi kuadrat memperlihatkan bahwa nisbah segregasi 9 : 3 : 4 dapat diterima untuk nisbah sifat tahan : moderat : rentan (Tabel 4.11). Pola segregasi Z2 demikian hanya dapat tejadi apabila genotipe Z1 bersifat heterozigot, yaitu AaBb seperti pada segregasi keturunan klon Z1 PR 255 (Tabel 4.5). Jadi model genotipe sifat ketahanan terhadap PGDC pada klon karet RRIC 100 adalah AaBb. Model persilangan antar pohon klon Z1 RRIC 100 yang ditampilkan pada Tabel 4.14 memperlihatkan frekuensi genotipe populasi Zz yang mungkin dihasilkan dari Z1 tersebut.
Tabel 4.14. Model persilangan antar pohon klon karet Z1 RRIC 100 bergenotipe AaBb
Gamet % AB L/4 Ab % aB L/4 ab
% AB
Ab % aB !h ab
1/16 AABB 1/16 AABb 1/16 AaBB 1/16 AaBb 1/16 AABb 1/16 AAbb 1/16 AaBb 1/16 Aabb 1/16 AaBB- 1/16 AaBb 1/16 aaBB 1/16 aaBb 1/16 AaBb 1/16 Aabb 1/16 aaBb 1/16 aabb
Dari persilangan antar pohon Z1 pada Tabel 4.14 diperoleh 16 genotipe tanaman Zz. Klasifikasi berdasarkan model genetik sifat ketahanan terhadap PGDC diketahui bahiva 9/16 bagian dari populasi tersebut termasuk katagori tahan (A-B-), 3/16 bagian moderat (A-bb) dan 4/16 bagian yang lainnya tergolong rentan (aa--). Dari 46 anggota dalam populasi yang diamati, ketenvakilan untuk ketiga fenotipe ketahanan tersebut berturut- turut adalah 28,4 dan 14 individu tanaman (Tabel 4.15).
Tabel 4.15. Model genotipe 22 keturunan klon karet Z1 RRIC 100 untuk tiap kelas ketahanan terhadap PGDC
I Fenotipe Jumlah individu : Genotipe Frekuensi
1
1
Tahan 2 8 AABB 1/16I
I AABb 2/16 I 1 AaBB 21 16 AaBb 4/16 4i
j
Moderat AAbb 1/16i
Aabb Rentan 14 AaBB AaBb Aabbi
Jumlah 46-
11
1
4.2.2.3. Populasi
Z2
dari
klon
karet Z1
RRIM
712
Jumlah tanaman dalam populasi Z2 yang berasal dari keturunan pohon klon Z I RRIM 712 yang dapat diuji sifat ketahanannya terhadap PGDC adalah 75 nomor. Hasil uji ketahanan terhadap penyakit tersebut diketahui 38 nomor tanaman memiliki sifat tahan, 11
nomor tergolong moderat dan 26 nomor yang lain termasuk dalam katagori rentan. Nisbah fenotipe 9 : 3 : 4 untuk ketiga katagori ketahanan tersebut sesuai dengan pola segregasi Z2 kalau genotipe Z1 adalah heterozigot AaBb seperti pohon klon karet Z1 RRIC 100 (Tabel 4.13) dan PR 255 (Tabel 4.4). Dan frekuensi genotipe A-B- (9116) bersifat tahan, A-bb (31 16) moderat, dan aa-- (41 16) bersifat rentan (Tabel 4.16).
Tabel 4.16. Model genotipe Z2 keturunan klon karet Z1 RRIM 712 untuk tiap kelas ketahanan terhadap PGDC
1
Fenotipe Jumlah individu I Genotipe iI I i i Frekuensi
!
Tahan i 3 8 i I I i I i Moderat 11 \i
i Rentan AABB AABb AaBB AaBb AAbb Aabb AaBB AaBb Aabb I I I I JumlahI
75i
-
I I 14.2.3. 3Ienduga model genetik tetua klon karet TJIR 1 dan
PR 107
Berdasar hasil analisis segregasi pada populasi Z2 seperti diuraikan di atas diketahui bahwa model genotipe sifat ketahanan terhadap PGDC klon karet PR 255 adalah AaBb (tahan) dan genotipe PR 303 adalah Aabb (moderat). Kedua klon karet tersebut merupakan keturunan dari silangan antara tetua klon karet P1 TJIR I (rentan) dan klon P2
PR 107 (tahan). Model genetik sifat ketahanan terhadap PGDC dari kedua tetua tersebut belum diketahui.
Model genetik tetua klon karet TJIR 1 dan PR 107 tidak dapat ditentukan dengan menggunakan metode sebelumnya, mengingat tidak tersedia populasi segregasi Z2 dari masing-masing tetua tersebut. Kedua klon karet tersebut merupakan klon primer (Dijkman, 1951) yang sudah tidak ditanam lagi di kebun karena produktivitasnya tergolong rendah. Klon-klon tersebut hanya dapat ditemukan dalam bentuk koleksi di kebun entres, sehingga tidak dapat menghasilkan biji.
Untuk menentukan model genetik kedua tetua tersebut digunakan metode analisis pedigri, yaitu penelusuran silsilah dengan memperhatikan suatu sifat fenotipe yang dimiliki oleh tiap generasi yang dipelajari. Fenotipe yang dimaksud adalah sifat ketahanan PGDC pada kedua tetua dan dua individu Z1 (klon PR 255 dan PR 303). Oleh karena model genetik dan fenotipe ketahanan pada generasi 2 , telah diketahui, maka model genetik kedua tetua tersebut dapat diduga.
Fenotipe ketahanan PGDC pada PI klon TJIR 1 adalah rentan, sehlngga berdasar model sebelumnya, maka genotipe yang dimungkinkan adalah aaBB, aaBb, atau aabb. Sedangkan klon P2 PR 107 mempunyai fenotipe tahan terhadap penyakit tersebut, sehingga kemungkinan genotipenya adalah AABB, AaBB, AABb, atau AaBb. Persilangan antara kedua tetua tersebut menghasilkan beberapa genotipe, diantaranya genotipe tahan AaBb (PR 255) dan genotipe moderat Aabb (PR 303). Dengan membuat model persilangan antara T J R 1 x PR 107 serta mempelajari model genetik keturunannya, maka akan dapat ditentukan model genetik kedua tetua tersebut (Tabel 4.17).
l'abel 4.17. Model geriotipe yang dimungkinkan untuk tcti~a TJlR I dan I'R 107 dan ragaln genotipe Z I hasil persilangannya.
AaBE3 AaE3R A;~l%b Aa13B aaBI3 AaBB AaBb aaBB aaBb . ......-s.....-....--...----... ... -.--.--...- - ---.--~~.--..~~.-...~~...~. ... ...-. ~ ...
AaBB
1
Aal3B AaB:1
AaBB ad313 AaBB AaRb aaBI3 aaBb" Yang dicetak tebal menunjukkan ~iiodel genotipe ZI yang dikehendaki, model genotipc tetua yang paling mungkin bila dapat nienghasilkan dua model genotipe
Z , AaBb (I'R 255) dan Aabb (PK 303) secara bersama-sama.
1
ab AaBb -- AaBb Aal3b Aabb ... AaRb Aabb AaBb aaBb AaBb aaBbAaBb Aabb aaBb aabb
... ~
A a S b Aabb aaBb aabb
Dengan mempelajari semua kemungkinan model genotipe persilangan TJIR 1 dan PR 107, serta zuriat hasil persilangannya (Tabel 4.17), maka model genotipe tetua klon karet TJIR 1 yang paling mungkin adalah aaBb atau aabb, sedangkan untuk klon PR 107 adalah AABb atau AaBb. Model genotipe PI dan Pz lainnya tidak mungkin karena tidak dapat menghasilkan kedua genotipe Z I secara bersama.
4.3.
Analisis Penanda Molekul Sifat Ketahanan Tanaman Karet
terhadap Penyakit Gugur Daun Corynespora
4.3.1.
Penanda isozim
Penelitian ini telah menganalisis 12 sistem enzim pada tanaman karet, yaitu alkohol dehidrogenase (ADH), malat dehidrogenase (MDH), katalase (CAT), polifenoloksidase (PPO), katekol oksidase, peroksidase (PER), superoksid dismutase (SOD), kalkon isomerase, asam fosfatase (ACP), esterase (EST), aspartat aminotransferase (AAT), dan P- glukosidase (P-GLU). Dari 12 isozim yang dianalisis, terdapat 5 isozim yang meperlihatkan pola pita monomorfik, 5 isozim polimorfik, dan 2 isozim lainnya tidak memperlihatkan pita yang diharapkan (Tabel 4.18). Pita-pita yang terbentuk dalam suatu sistem enzim diasumsikan dikontrol oleh satu alel, yang penomorannya dimulai dari yang mengalami migrasi tercepat pada proses elektroforesis (Yeang et al. 1998)
Isozim yang bersifat monomorfik tidak dapat digunakan sebagai penanda suatu sifat ketahanan, karena tidak dapat membedakan antara genotipe rentan dan tahan. Atau dengan kata lain gen penyandi protein tersebut dipunyai oleh semua genotipe yang dipelajari. Isozim monomorfik yang ditemukan pada klon tanaman karet adalah : alkohol
dehidrogenase (ADH), katalase (CAT), katekol oksidase (CO), asam fosfatase (ACP), dan P-glukosidase (P-GLU).
Jenis isozim yang kemungkinan dapat dipergunakan sebagai penanda suatu sifat adalah yang polimorfik. Yang dimaksud dengan polimorfik adalah isozim yang memperlihatkan pola pita yang berbeda antar genotipe contoh yang dianalisis, sehingga dapat dipergunakan sebagai pembeda. Pada penelitian ini diketahui ada lima jenis isozim yang bersifat polimorfik, yaitu : malat dehidrogenase (MDH), peroksidase (PER), superoksid dismutase (SOD), esterase (EST) dan aspartat aminotransferase (AAT).
Tabel 4.18. Jenis isozim, jumlah pita dan polimorfisme pada klon karet Z L BPhl 1 (tahan), semai Z2-tahan dan semai &-rentan terhadap PGDC
i i Jumlah pita Isozim No. 1 t ) Polimorfisme i pita isozim z,(T)')
j
Z2(T)/
Z2(R)i
i: i 1. iAlkoholdehidrogenase(ADH) , 5 . 5'
1
51
monomorfikMalat dehidrogenase (MDH) 1 5 . 4 1 ' 5 i polimorfik
i
Katalase (CAT) 1 1 1 1 t : 1 L monomorfik
j
Polifenol oksidase (PPO)
-
i-
i-
I Katekol oksidase
Peroksidase (PER)
Superoksid dismutase (SOD) Kalkon isomerase
Asam fosfatase (ACP) Esterase (EST)
Aspartate aminotransferase (AAT) P-glucosidase (P-GLU) monomorfik polimorfik polimorfik
-
monomorfik polimorfik polimorfik monomorfi k"T = tahan, dan R = rentan
G a m k 4.3.
Pola
pita
isozim
mrtlat d e h i d m p w (MDrr)
pada
klm b e iZ r - m
f k m a i ~ d a n & - r n
P ~ p a d a i s o e i m P E R m e n g h a s i l k a n p i t a y a n g c ~ j e l a s , b a i k p d a ~
p i t i f
mupun
kutub
ne@f
( W b a r
4.4).Pola
pita
pada
kutubp i t i f
m o n ~ ~ o r & d e n g a n j u m l a h p i t a ~ ~ m p a t b u a h S e d a n g k r t n j u n l l a h p i t a p a d a
*kutub mgatif
rmtuk Z1, &-tahm dmZ r r a t m
berh&turut 4,3,dun
3.Pola
pita
padrr
Za-~ d m r n & ~ ~ ~ ~ i s o z i m P E R t i d a k d a p a o t d i ~ u n l l l t p e n a n d a
&ktk e t a h m
tanamanhrdkrhadap
PGDC.
(a)
Clambar
4.4.Pola pita isozim peroksidase
(PER)pada
klon
Bcaret Z l - h h ,&-tahan, dm
Zrrentrm PGDC.
h t u k
-bar
dapatdisajihn
mlah
gel
diliktdi
a&ww
trmsItaptinafor(Gambar
4.5).Dari
pmbartersebut berlihat
bahwajumlrth pita yang
kdmtukp d a
Z 1 - h h , Za-tahmdan
Z2-rentan batumt-tumt : 6, 4,dm
6. Polapita
pada Z1dan
Zpmtanpersis
am,
-pi
tidak
k d q m tpada
hotiprenm
makaisozim
SOD
tidak
&patdigunalaan
s&gaiAngka 1 d m 2 menunjukkan ulmgan
Gambar 4.5. Interpretasi pola pita isozim superoksid dismutase (SOD) pada klon karet Z1-tahan, semai Zz-tahan, dan &-rentan PGDC
Harnpir sama dengan SOD, pola pita isozim esterase (EST) pada klon karet Z1 dan semai Z2-rentan identik, yaitu masing-masing mempunyai 5 buah pita. Sedangkan pada semai Z2-tahan hanya mempunyai 4
buah
pita, karena tanaman ini tidak mempunyai pita nomor 4 (Gambar 4.6). Dengan alasan yang sama seperti pada SOD, isozim EST tidak dapat digunakan sebagai penanda sifat ketahanan tanaman karet terhadap PGDC.Pola pita isozim aspartat aminotransferase (AAT) tidak terlihat dengan jelas. Pita- pita yang terbentuk hanya dapat dilihat apabila gel diletakkan di atas uv transluminator. Jumlah pita yang terbentuk pada isozim tersebut berhuut-turut adalah 4,3, dan 3 untuk Z1, Zt-tahan, dan Z2-rentan. Pita pertama hanya terdapat pada Z1 (tahan), tetapi tidak ditemukan pada fenotipe tahan yang lain ( Z ~ t a h a n ) maupun Z2-rentan (Gambar 4.7). Dengan demikian isozim AAT juga tidak dapat digunakan sebagai penanda sifat ketahanan tanaman karet terhadap PGDC.
Aagka 1 dan2
lMufiiuttafi-
(a)
-
pin pita h o i m EST peda gd pati@ ) = ~ p & # t a i s o a i m B S T p l p d r r g d p e t i
-bar
4.6.Pols pita
isozim
esterase
(EST)
peda
Llon
karct
21-tahan,
semi &bhaqdan&-rentmfGDC
(a)
oambar
4.7.Pola
pita isozim
asptatadnotransferase
(AAT)peda
klon
Laret Zl-tahan, semai Zrtahan, dm &-mtaaPGDC
Isozim polifenol oksidase (PPO) dan kalkon isomerase tidak memperlihatkan pola pita isozim yang diharapkan meskipun telah dilakukan analisis berulang-ulang dengan bermacam modifikasi, baik konsentrasi substrat maupun jenis dan pH larutan penyangga yang digunakan. Penggunaan gel poliakrilamid pada proses elektroforesis mungkin dapat dicoba.
4.3.2.
Penanda
RAPD
4.3.2.1.
Uji kuantitas
dm
kualitas DNA
DNA hasil isolasi contoh daun tanaman karet klon TJIR 1 (PI), PR 107 (PI), PR 255 (Z1), semai Zz-tahan serta semai Zl-rentan dilarutkan dalam bufer TE sebagai sediaan DNA. Meskipun berat contoh daun pada saat ekstraksi DNA sama, akan tetapi jumlah DNA yang diperoleh belum tentu sama, sehingga konsentrasi larutan sediaan DNA perlu diketahui. Hal ini diperlukan untuk menghitung besar faktor pengenceran pada saat pembuatan larutan untuk reksi amplifikasi DNA dengan mesin PCR.
Hasil elektroforesis pada uji kuantitas sediaan DNA tiap tanaman contoh yang dipelajari ditampilkan dalam Gambar 4.8. Dengan membandingkan ukuran tebal pita pada DNA contoh dengan marka DNA k , maka konsentrasi tiap sediaan DNA dapat diperkirakan (Tabel 4.20). Yaitu bahwa konsetrasi sediaan DNA genotipe P1
*
530 ng/ pt, P2 i 530 ngl p t , Z,*
265 ng/ pt, Zl-tahan*
530 ng/ pt, dan Zz-rentan i 530 ng/ pC.Kualitas DNA contoh dipelajari dengan cara uji pemotongan DNA dengan menggunakan enzim restriksi EcoRI. Enzim ristriksi EcoRI akan memotong utas DNA pada situs tertentu, sehingga menghasilkan fragrnen-fragrnen DNA dengan bermacam
Bits
d i h k h n elektmhds
akan
diptrolch p h
pitay m g
m
e
&
ubm
molekul
tdaggihhgp
tcmdah ( m e t ) .Gmbar 4.8. Pert,andingan
tebal
pita DNAcontoh
tanaman
karet
yang
diuji
dmgm tebal pitaDNA
k.
Tabel
4.20.Perkrraan
k0-i
lanrEanstok
DNA
contob
tammanlawet
Pi, pz, 21,&ahsn,
b&--wur
M
1 2 3 4S
Hasil
e
l
~
tiap DNA
f
~
~0-4
b i k
p4
llaaupun ~gtidak
diptong
dmtpnemim
restriksi EmRI ditaporIran
rlalam
Gambar
4.9.Dari
gmbm tersebutteflihat
bahwasemua
DNA
contoh
y m gdiptong
h g a n e&EmRI
DNA-
DNAl(marka)
DNAPi DNAPz DNA&DNA&--
D N A & - ~ ~ I W IVol.
ujiw)
1 22
2 22
TebalpitaMM
lx
2~ 2xIX
2x 2xK m m t m i f n # f l )
530*
530*
530 k 265*
530 k 530 IGambm 4.9.
Pola
pitaDNA
contoh
h a m a n
lavet
yang
tkbk
diptong
c h g m emdm
m t c k i E
d
(a),dan
yang
dipotongdeagan
(b).
Gambr
4.10rnemperlihatkan pola
pita RAPDdari
semm
habat
klon kmt
yang
diuji, yaituPI,
P2, Z1, dan&.
Penggmmn plimer
OPN
15,OPD
03dan
OPD
11tidak
+m ~ ~ R A P D y a a g d a p a t d i ~ ~ p e m ~ S i W t r r h R n a t a u r e n t a n
tamam karet
terhadap
PGDC.
Primer
OPN
15hanya
m e n g h d h
Iitlzapita
m o m o ~
baik
pada
PI,
Pz,
21,
&-tahan
m a u p
&rmtm
Mmgprimer
OPD
03dm
OPD
11meskipun rnenghwilkan pita polimo*
a h
Mapiti-
a h yaug dapat~psrgMslolnsebagsipcnands- y a a g - p ~ ~ ~ b yaitrt
OPH
05dm
OPB 07juga
tidak
menemdm
marka RAPD
yang
d s p l t m d e d h n
t s n a m s n h t t e a a n d m i y m g r e n t a n t e r h a d s p W D C , m e s k i p l m . k e d ~ m u n p u
m m g b d b
pla
pita
polimorfik
(Gambar
4.1 1).Gambar
4.10.Pola pita
RAPD
maman kmtdengan
primerO W
IS,
OPD
11, dmOm)
03.d a m
Gambar 4.11.
Pola
pita RAPI) tammankaret
dengat
prima OPH 05,Hal
ymgmenark
ptida pggmm primerOPW
19 (Oambw 4.11).M & p
p b mtersebut
h y ameqgbdlksn
empat
pi@ tetapi
satupita
dtqm
ukutanwkitar
550bp
dapt menjadikandichrt
marka
ketahman
t4nem4nkaret
tdudapR33X.
Pita
OPW 1 9 ~ ~ 0tenlapat
smmkmshtm
pada
~ ~ Ugmotipe
E I WP2,
21
drln
Z a - e ) ,
tetapi
tidak
muncul
p d a
@pe
rentan(PI
dan
&-ratan).
Untuk
menguji
pendapat
bersebut,
pengpmm
phmrOPW
19diprims
pda
4genotipe
& - t a h dan2
genotipe
&
-mta.Hasil
rCAPD
mem*
bafiwa pitaOPW
lgsurm w u l
konsisQenpada
gadm
tduqtetapi
tidek
d
i
u
pada
genotipe
ratan (Gambar 4.12).I%qm
~ ~ ~ p i t a 1 9 s j o d a p a t d i ~ s e b a g a i p e a a n d a s i f a t E a h a n ~ O P w
lrrrret
tddqPGDC. Untuk
htjuan bmdmtmsih
diperlukan
m i a n -1h j a t
dengan
men-
p p d d tammanm t o h
yanglet&
baryak
-bar
4.12.Poh pita
RAPD
tmmmkaret
d e wpdm~t
O m
19dmgm
4.4.
Pembahasan Umum
Hasil analisis dua percobaan pertama, yaitu uji ketahanan diferensial beberapa klon karet terhadap tiga isolat C. cassiicola serta analisis genetik pewarisan sifat ketahanan tanaman karet terhadap PGDC mendukung hipotesis bahwa sifat ketahanan tanaman karet yang diuji terhadap penyakit tersebut dikendalikan secara vertikal. Pada percobaan pertama, nilai r atau laju infeksi antar klon karet yang diuji relatif tidak berbeda (Tabel 4.2). Atau dengan kata lain garis linier perkembangan penyakit antar klon sejajar satu dengan yang lainnya (Lampiran 6). Hal ini merupakan salah satu petunjuk bahwa suatu sifat dikendalikan secara vertikal oleh sejumlah kecil gen (Van der Plank 1968, Nelson & MacKenzie 1973). Meskipun demikian jumlah gen utama yang mengendalikan sifat ketahanan tersebut belum dapat diketahui. Pada penelitian kedua tentang pewarisan sifat ketahanan tanaman karet terhadap PGDC membuktikan adanya indiksasi ketahanan vertikal tersebut. Yaitu bahwa sifat ketahanan tanaman karet terhadap PGDC dikendalikan oleh dua pasang gen utama yang berinteraksi secara epistasis resesif. Sifat tahan dapat diekspresikan apabila minimum terdapat satu ale1 dominan pada kedua gen penyusunnya (A-B-). Sedangkan gen A resesif homosigot (aa) menekan ekspresi gen dominan B untuk memunculkan sifat tahan, sehingga sifat yang ditampilkan adalah rentan terhadap PGDC.
Ketahanan vertikal pada umumnya melibatkan reaksi hipersensitif (hypersensitive reaction, HR) (Agrios 1988, Oka 1993). Ketidaksesuaian antara inang dan patogen melalui mekanisme HR sering kali dikendalikan oleh sejumlah gen yang bersifat epistatik (Lamb et
al. 1989). Reaksi hipersensitif menyebabkan kematian sel secara cepat di sekitar titik infeksi sehingga nampak gejala nekrosis kecil pada jaringan daun yang terserang. Patogen
yang terdapat di dalamnya akan terisolasi sehingga tidak aktif atau mati (Goodman & Novacky 1994, Jackson & Taylor 1996, Delaney 1997, Pennell & Lamb 1997, Gilchrist 1998). Bersamaan dengan berlangsungnya proses reaksi hipersensitif, sel inang juga mensintesis senyawa fitoaleksin yang dapat membunuh patogen yang terdapat di dalarnnya. Morfologi gejala seperti terlihat pada skor 1 dalam Gambar 3.2 memperlihatkan gejala nekrosis yang mirip dengan gejala yang biasa terjadi pada reaksi hipersensitif. Dugaan bahwa mekanisme ketahanan tanaman karet terhadap PGDC melibatkan reaksi hipersensitif juga dilaporkan oleh peneliti sebelumnya (Breton et al. 2000).
Hasil penelitian ini berlawanan dengan pendapat sebelumnya yang mengatakan bahwa sifat ketahanan tanaman karet terhadap PGDC kemungkinan dikendalikan secara poligenik (Tan & Tan 1996). Konsekuensi atas sifat ketahanan yang dikendalikan secara vertikal adalah bahwa sifat ketahanan tersebut relatif mudah dipatahkan (Oka 1993, Agnos 1988, Tan & Tan 1996). Selama dekade terakhir telah banyak laporan yang menyebutkan bahwa tingkat ketahanan klon karet terhadap PGDC telah dapat dipatahkan. Klon-klon yang tadinya dikenal sangat tahan terbukti mulai dapat terserang. Gejala penurunan tingkat ketahanan klon karet terhadap PGDC terjadi baik di Indonesia, India, maupun Malaysia (Jayasinghe & Silva 1996b, Othman et ul. 1996, Situmorang et ul. 1996).
Adanya perbedaan tingkat virulensi antar isolat C. cassiicolu yang diuji pada percobaan pertama memperkuat dugaan sebelumnya tentang adanya keragaman ras fisiologi cendawan tersebut di Indonesia (Situmorang et al. 1996). Dengan demikian pemilihan klon karet yang sesuai untuk suatu daerah tidak cukup hanya berdasar karakteristik tanah dan iklim saja, tetapi juga perlu dipertimbangkan kemungkinan adanya ras-ras C. cmsiicolu yang lebih virulen.
Dari hasil percobaan kedua diketahui bahwa pemeringkatan ketahanan klon karet terhadap PGDC menjadi lima kelas ketahanan terbukti kurang tepat. Jumlah kelas ketahanan yang lebih tepat adalah tiga, yaitu tahan, moderat dan rentan. Hal ini sesuai dengan hasil analisis yang menyebutkan bahwa sifat ketahanan tanaman karet terhadap PGDC dikendalikan oleh dua pasang gen utama yang mengalami interaksi secara epistasis. Dengan demikian respon ketahanan klon karet terhadap tiga isolat C. cassiicola yang ditunjukkan pada Tabel 4.1 diperbaiki menjadi seperti pada Tabel 4.20 berikut ini.
Tabel 4.20. Respon ketahanan PGDC pada 16 klon karet yang diinokulasi dengan 3 isolat C. cassiicola berdasarkan tiga kelas ketahanan
I
I Isolat C. cassiicola')I
Klon karet 1i
BPMI BPM 24 PI3 260I
PPN 2058/
PPN 2444 PPN 2447 PR 26 1 PR 300 PR 303 RRIC 100 RRIC 103 RRIC 1 101
RRIM600 RRIM 712/
RRIM725I
aha an') tahan moderat tahan moderat rentan moderat tahan moderat moderat tahan rentan tahan moderat tahan moderat I tahan I tahan I tahan I tahan i i moderat rentan i rentan I tahan I tahan I I ! tahan I tahan I rentan 1 tahan I i tahan i tahan I rentan i moderat tahan moderat tahan rentan rentan rentan tahan moderat moderat tahan rentan moderat moderat tahan rentan"CC, = isolat dari Kalimantan Selatan, Cct = isolat dari Jawa Tengah, dan Cc3 = isolat dari Sumatra Selatan 2'Tahan = keparahan penyakit (KP) 0-40 %, moderat = KP 4 1-60 %, dan rentan = KP 6 1
-
100 %Klon-klon AVROS 2037, BPM 24, PPN 2058, PPN 2447, PR 300, PR 303, RRIC 110, RRIM 600, dan RRIM 725 pada Tabel 4.20 memperlihatkan respon yang berbeda terhadap tiga isolat C. cassiicola yang diinokulasikan. Dengan demikian klon-klon tersebut dapat dipergunakan sebagai inang diferensial untuk menguji ras fisiologi dari suatu isolat C. cassiicola. Klon-klon yang dikenal mempunyai tingkat ketahanan tinggi (BPM 1, PB 260, PR 261, RRIC 100, dan RRIM 712) menunjukkan respon ketahanan yang sama terhadap ketiga isolat yang diuji, sedangkan klon-klon yang tergolong paling rentan (PPN 2444 dan RRIC 103) menunjukkan respon kerentanan yang sama terhadap ketiga isolat C.
cassiico/a tersebut. Dengan demikian dua kelompok klon yang disebut terakhir tidak dapat dipergunakan sebagai inang diferensial.
Pada percobaan isozim tidak diperoleh penanda yang dapat digunakan untuk seleksi sifat ketahanan tanaman karet terhadap penyakit gugur daun corynespora. Meskipun pelaksanaannya cepat dan mudah, analisis isozim untuk penanda sifat ketahanan mengandung beberapa kelemahan, antara lain karena jumlah pita yang sedikit serta pita yang terbentuk pada umumnya kurang jelas. Rendahnya jumlah pita memperkecil peluang tejadinya polimorfisme antar individu tanaman yang diteliti. Pada analisis polifenol oksidase (PPO) dan kalkon isomerase bahkan tidak muncul pita yang diharapkan. Hal ini dapat disebabkan oleh dua kemungkinan. Pertama, penggunaan protokol analisis yang belum tepat sehingga perlu dicoba dengan berbagai modifikasi, baik terhadap bufer, jenis gel maupun komposisi larutan pewarna. Kedua, kemungkinan karena enzim tersebut tidak disintesis secara konstitutif oleh tanaman, sehingga tidak terjadi reaksi pewarnaan. Untuk membuktikan ha1 tersebut dapat dilakukan dengan cara membandingkan dua tanaman yang sama, yang satu diinokulasi dengan C. cmsiicolu sedang yang lainnya tidak diinokulasi.
Bila enzim tersebut merupakan bagian dari reaksi pertahanan tanaman terhadap patogen, maka pada tanaman yang diinokulasi akan terbentuk enzim yang dimaksud. Dengan demikinan pada proses pewarnaan akan terbentuk pita-pita isozim. Kelemahan lain penggunaan isozim untuk penanda suatu sifat adalah masih terbatasnya jumlah isozim yang dapat dipergunakan.
Penggunaan penanda RAPD mempunyai kelebihan dibanding isozim, antara lain tersedianya banyak primer yang dapat dipergunakan dalam reaksi amplifikasi DNA. Tersedianya banyak primer yang dapat dipergunakan akan memperbesar peluang diperolehnya penanda yang dikehendah. Pada analisis RAPD diketahui bahwa pita OPW 19 dengan ukuran sekitar 550 bp muncul secara konsisten pada fenotipe tahan, tetapi tidak ditemukan pada fenotipe rentan. Dengan demikian OPW lgjjo kemungkinan dapat digunakan sebagai penanda sifat tahan tanaman karet terhadap PGDC. Meskipun demikian ha1 tersebut masih perlu dibuktikan pada genotipe karet yang lebih luas mengingat jumlah tanaman yang dianalisis pada percobaan ini masih sangat terbatas. Disamping itu kelas ketahanan populasi tanaman b a n didasarkan pada hasil uji ketahanan terhadap isolat C.
cassiicola yang berasal dari Sumatra Selatan (CC~). Dengan demikian masih diperlukan penelitian lebih lanjut terhadap isolat-isolat virulen yang lain agar temuan ini dapat diaplikasikan dalam seleksi sifat ketahanan tanaman karet terhadap PGDC.