1
UJI KOMPETISI INTERSPESIFIK ANTARA Trichogramma chilonis Ishii DAN Trichogramma japonicum Girnult
(HYMENOPTERA: TRICHOGRAMMATIDAE)
DALAM TELUR Chilo auricilius Dudgeon (LEPIDOPTERA: PYRALIDAE)
Dwi Anggun Putri Solikha1, Fatchur Rohman2, dan Susilowati3
Program Studi Biologi, FMIPA, Universitas Negeri Malang Jl. Semarang 5, Malang, Indonesia
E-mail: anggunputri030695@gmail.com
ABSTRAK: Uji kompetisi interspesifik antara Trichogramma chilonis dan Trichogramma japonicum dalam telur Chilo auricilius telah dilakukan dengan tujuan untuk mengetahui kompetisi
interspesifik antara T. chilonis dan T. japonicum ketika keduanya didedahkan pada kelompok telur yang sama. Penelitian ini memberikan informasi mengenai teknik yang tepat untuk mengaplikasikan T. chilonis dan T. japonicum sehingga upaya pengendalian C. auricilius pada perkebunan tebu menjadi optimal. Penelitian ini merupakan penelitian eksperimental menggunakan Rancangan Acak Lengkap (RAL) dengan 5 macam perlakuan dan diulang 10 kali. Data yang diperoleh berupa jumlah imago Trichogramma chilonis dan Trichogramma japonicum yang muncul dan larva Chilo auricilius yang menetas, kemudian dianalisis menggunakan uji Kruskal-Wallis dan dilanjutkan dengan uji Dunn. Hasil penelitian menunjukkan bahwa asosiasi T.
chilonis dan T. japonicum (PB) pada waktu bersamaan menunjukkan tingkat parasitasi yang
rendah, hal ini diakibatkan oleh adanya kompetisi fisik antara imago T. chilonis dan T. japonicum untuk memperebutkan inang telur C. auricilius. Pada saat asosiasi T. chilonis dan T. japonicum diberikan jeda waktu satu jam dan diacak parasitoid primernya maka tingkat parasitasi telur C.
auricilius menjadi tinggi. Trichogramma japonicum diduga berperan sebagai hyperparasitoid
karena T. japonicum tetap muncul pada telur yang sebelumnya telah terparasit oleh T. chilonis.
Trichogramma japonicum mampu berkompetitif dalam memperebutkan nutrisi telur C. auricilius
ketika T. chilonis dan T. japonicum diasosiasikan dengan jeda waktu satu jam.
Kata Kunci: Trichogramma chilonis Ishii, Trichogramma japonicum Girault, kompetisi interspesifik, telur Chilo auricilius Dudgeon.
ABSTRACT: Test of interspecific competition between Trichogramma chilonis and Trichogramma japonicum in eggs Chilo auricilius been conducted to understand the interspecific
competition between T. chilonis and T. japonicum when both of them exposing the same egg of C.
auricilius. This study provides information on the proper technique for applying T. chilonis and T. japonicum so that efforts to control C. auricilius on the sugar cane to be optimal. This study was
an experimental study using a Completely Randomized Design (CRD) with 5 kinds of treatment and repeated 10 times. The data obtained is number of adult T. chilonis and T. japonicum emerging and C. auricilius larvae that hatch, then analyzed using Kruskal-Wallis test followed by Dunn test. The results showed that the association T. chilonis and T. japonicum (PB) at the same time showed a low level of parasitism, this is caused by their physical competition between imago T. chilonis and T. japonicum to fight over the host eggs C. auricilius. The parasitism rate on egg of C.
auricilius be high when T. chilonis and T. japonicum is associated with a lag time of one hour and
randomized primary parasitoid. Trichogramma japonicum might as hyperparasitoid because T.
japonicum oviposition on the eggs that had previously exposed by T. chilonis. Trichogramma japonicum is able to be competitive in getting the nutrition of eggs C. auricilius when T. chilonis
and T. japonicum is associated with a lag time of one hour.
Keywords: Trichogramma chilonis Ishii, Trichogramma japonicum Girault, interspecific competition, eggs of Chilo auricilius Dudgeon.
2
PENDAHULUAN
Tebu (Saccharum officinarum) merupakan salah satu komoditas tanaman perkebunan yang dimiliki oleh Indonesia. Perkebunan tebu terdapat pada 5 (lima) provinsi di Indonesia, yaitu Jawa Timur, Jawa Tengah, Lampung, Jawa Barat dan Yogyakarta (Pusat Data dan Sistem Informasi Pertanian, 2013). Hal penting yang harus diperhatikan dari peningkatan mutu dan kualitas hasil produksi tebu yaitu pembuatan bibit unggul dan pengendalian hamanya. Hama yang umumnya menyerang perkebunan merupakan serangga dari ordo Lepidoptera (Huffaker & Messenger, 1989). Hama kebun tebu dapat dibedakan berdasarkan bagian tanaman yang diserang. Hama yang sering ditemukan adalah hama penggerek. Hama penggerek menyerang bagian tanaman tebu tertentu yaitu penggerek yang menyerang bagian pucuk dan penggerek yang menyerang batang. Hama penggerek yang menyerang batang tebu adalah penggerek bergaris (Chilo sacchariphagus), penggerek berkilat (Chilo auricilius), penggerek abu-abu (Eucosma scistaceana), penggerek kuning (Chilotraea infuscatella), penggerek jambon (Sesamia
inferens), dan penggerek batang tebu raksasa (Phragmatocea castanea) (Pusat Penelitian
Perkebunan Gula Indonesia, 2008). Penggerek batang berkilat C. auricilius dan penggerek batang bergaris C. sacchariphagus menjadi hama utama tanaman berusia 3 bulan yang mengakibatkan penurunan bobot batang dan pertumbuhan ruas menjadi tidak normal (Meidalima, et al., 2012). Apabila bobot batang tebu mengalami penurunan maka hasil produksi juga akan berkurang, sehingga jumlah pendapatan dari produksi gula juga akan berkurang. Oleh karena itu, dibutuhkan teknik pengendalian tertentu untuk mengatasi hama penggerek pada tanaman tebu. Pengendalian hama yang diharapkan yaitu pengendalian yang bersifat alami, efisien, berkelanjutan, tidak mencemari lingkungan, serta tidak mengganggu dan merusak keragaman hayati. Pengendalian tersebut disebut dengan pengendalian hayati (biological
control) salah satunya yaitu dilakukan dengan menggunakan musuh alami. Soula, et al. (2003)
menegaskan pula bahwa musuh alami dibutuhkan untuk program pengendalian yang lebih baik karena sesuai dengan sistem bioekologis.
Agen hayati yang sering dijadikan sebagai pengendali hama adalah genus Trichogramma dan Trichogrammatoidea (Ordo Hymenoptera) (Clausen 1940; Nagarkatti & Nagaraja, 1977). Menurut Nagaraja (1978), Alba (1989), Meilin, et al. (2000), Trichogrammatoidea merupakan agen pengendalian hayati yang memparasiti serangga pada fase telur, sehingga genus ini sangat dikenal sebagai parasitoid telur. Genus Trichogramma dan Trichogrammatoidea umumnya memparasiti telur ordo Lepidoptera. Serangga yang termasuk dalam genus Trichogramma yaitu
Trichogramma chilonis Ishii dan Trichogramma japonicum Girault. Keduanya telah diketahui
dapat memparasiti telur penggerek batang tebu (C.auricilius) (Herlina, 2010; Sudarsono, 2011), sehingga tidak menutup kemungkinan keduanya dapat memparasiti kelompok telur C. auricilius yang sama atau juga disebut dengan peristiwa multiparasitisme. Pada saat terjadi multiparasitisme keduanya dapat melakukan interaksi kompetisi antara T. chilonis dan T.
japonicum. Driesche, et al. (2008) mengemukakan bahwa risiko introduksi parasitoid di lokasi
baru adalah serangan dari organisme/musuh alami lokal, baik berupa gangguan dari parasitoid yang lain (superparasitoid dan hiperparasitoid), predator (khususnya terhadap imago parasitoid),
3 maupun mikroba entomopatogen (virus, bakteri atau fungi). Serangan oleh musuh alami lokal berpengaruh terhadap keberhasilan introduksi parasitoid di lapangan. Berdasarkan alasan tersebut, maka penting untuk diadakan penelitian lebih lanjut mengenai interaksi kedua spesies
T. chilonis dan T. japonicum apabila keduanya berada pada inang, waktu parasitasi, kondisi, dan
lingkungan yang sama. Penelitian ini mencoba untuk menguji kompetisi interspesifik T. chilonis dan T. japonicum dalam telur Chilo auricilius.
METODE
Penelitian dilakukan pada bulan Februari-Maret 2016 di Laboratorium Ekologi 109 Universitas Negeri Malang (UM) dan Laboratorium Quality Control (QC) PG. Pesantren Baru, Jengkol, Kediri. Penelitian ini merupakan penelitian eksperimental. Rancangan penelitian yang digunakan adalah Rancangan Acak Lengkap (RAL), menggunakan 5 macam perlakuan dengan 10 kali ulangan. Adapun 5 macam perlakuan yang digunakan yaitu,
PT.C = perlakuan pendedahan tunggal T. chilonis artinya pendedahan hanya oleh
Trichogramma chilonis selama satu jam,
PT.J = perlakuan pendedahan tunggal T. japonicum artinya pendedahan hanya oleh
Trichogramma japonicum selama satu jam.
PB = perlakuan pendedahan bersamaan T. chilonis & T. japonicum artinya pendedahan oleh parasitoid Trichogramma japonicum dan Trichogramma chilonis secara bersamaan selama satu jam,
C J = perlakuan pendedahan bergantian T. chilonis T. japonicum artinya pendedahan terlebih dahulu oleh Trichogramma chilonis, satu jam kemudian diganti dengan
Trichogramma japonicum,
J C = perlakuan pendedahan bergantian T. japonicum T. chilonis artinya pendedahan terlebih dahulu oleh Trichogramma japonicum, satu jam kemudian diganti dengan
Trichogramma chilonis.
Objek penelitian yang digunakan dalam penelitian adalah 240 betina imago
Trichogramma chilonis dan 240 betina imago Trichogramma japonicum yang berasal dari Pusat
Pelayanan Agens Hayati (PPAH) Tani Makmur, Beji, Pasuruan dan PG. Pesantren Baru, Jengkol, Kediri. Telur Chilo auricilius berasal dari PG. Pesantren Baru, Jengkol, Kediri. Jumlah seluruh telur Chilo auricilius yang digunakan sebanyak 50 kelompok telur atau 1000 butir telur. Adapun prosedur penelitian yang dilakukan yaitu pemeliharaan Trichogramma chilonis dan
Trichogramma japonicum, pembuatan piasan telur Corcyra cephalonica, pengembangbiakan dan
perbanyakan Trichogramma chilonis dan Trichogramma japonicum, pemeliharaan dan pengembangbiakan Chilo auricilius, pemanenan telur Chilo auricilius, pengujian kompetisi
Trichogramma chilonis dan Trichogramma japonicum pada telur Chilo auricilius, dan
pembedahan telur Chilo auricilius.
Pemeliharaan, pengembangbiakan dan perbanyakan T. chilonis dan T. japonicum dilakukan dengan menggunakan media inang telur Corcyra cephalonica. Telur yang digunakan sebagai inang merupakan telur yang telah di UV selama 15 menit agar telur tidak dapat menetas.
4 Inang telur tersebut kemudian dipias diatas kertas karton ukuran 2x8. Pemiasan dilakukan dengan menaburkan telur diatas kertas karton ukuran 2x8 yang sudah diberi lem pada salah satu bagian ujungnya. Setelah telur menempel dan lem mengering, piasan diberi keterangan waktu dan spesies Trichogramma yang memparasit, kemudian dimasukkan ke dalam botol biakan yang telah berisi T. chilonis atau T. japonicum.
Chilo auricilius dipelihara dan diperbanyak menggunakan media sogolan tebu (tebu yang
berumur 3-4 bulan). Media sogolan tebu yang digunakan sebagai media yaitu sogoloan tebu yang sudah disterilisasi dengan autoclave. Media biakan disusun dalam tabung erlenmayer kemudian diinokulasikan telur C. auricilius ke bagian dinding tabung. Telur di pelihara ± 3 minggu hingga waktu pemanenan. Pada saat pemanenan, C. auricilius dipanen dan dipisahkan berdasarkan fase dalam siklus hidupnya pada tempat yang berbeda.
Pengujian kompetisi Trichogramma chilonis dan Trichogramma japonicum pada telur
Chilo auricilius dilakukan sesuai dengan ketentuan perlakuan pendedahan yaitu pendedahan
tunggal T. chilonis, pendedahan tunggal T. japonicum, pendedahan bersamaan T. chilonis dengan
T. japonicum, pendedahan bergantian dengan parasitoid primer T. chilonis, dan pendedahan
bergantian dengan parasitoid primer T. japonicum. Hasil uji didukung dengan bukti pembedahan telur Chilo auriclilius umur 6 dan 8 hari pada perlakuan pendedahan bersamaan T. chilonis dengan T. japonicum, pendedahan bergantian dengan parasitoid primer T. chilonis, dan pendedahan bergantian dengan parasitoid primer T. japonicum.
Data yang diperoleh dari penelitian ini berupa jumlah imago Trichogramma chilonis dan
Trichogramma japonicum yang mucul dan larva Chilo auricilius yang menetas. Hasil data yang
telah diperoleh kemudian dilakukan analisis statistik menggunakan software SPSS 16.0 for
windows. Data dianalisis dengan menggunakan uji nonparametrik yaitu uji Kruskal-Wallis
kemudian dilanjutkan dengan Uji Dunn untuk mengetahui perbedaan rerata jumlah spesies T.
chilonis, T japonicum dan C. auricilius pada setiap perlakuan.
HASIL DAN PEMBAHASAN
Hasil penelitian menunjukkan bahwa nilai signifikasi yaitu 0,000 (<0,05) yang berarti macam perlakuan pendedahan parasitoid (PB, PT.C, PT.J, C→J, dan J→C) berpengaruh terhadap jumlah imago Trichogramma chilonis dan Trichogramma japonicum yang muncul dan larva Chilo auricilius yang menetas. Pada setiap perlakuan dapat dibandingkan nilai rerata jumlah imago Trichogramma chilonis dan Trichogramma japonicum yang muncul dan larva
Chilo auricilius yang menetas, sehingga dapat diketahui perlakuan yang paling efektif. Perlakuan
efektif dapat ditentukan dengan menghitung dan membandingkan jumlah larva C. auricilius yang menetas, semakin rendah larva C. auricilius yang menetas maka semakin efektif perlakuan yang digunakan. Jumlah larva C. auricilius yang menetas bergantung dengan kemampuan Trichogramma saat memparasit, semakin tinggi tingkat parasitasi suatu spesies Trichogramma maka jumlah larva C. auricilius yang menetas lebih sedikit sehingga usaha pengendalian semakin efektif. Hal ini dipertegas oleh pernyataan Herlinda (2004) & Herlina (2010), yang menyatakan bahwa semakin tinggi tingkat parasitasi maka semakin efektif usaha pengendalian
5 hayati. Tingginya tingkat parasitasi pada perlakuan pendedahan tunggal dipengaruhi oleh ketersediaan inang, daya parasitasi spesies parasitoid, kondisi lingkungan dan keberadaan musuh alami lain (Herlina, 2010).
Tingkat parasitasi T. chilonis pada telur C. auricilius lebih tinggi dibandingkan dengan tingkat parasitasi T. japonicum. Tingkat parasitasi T. chilonis yang tinggi juga telah dilaporkan oleh Li (1994) yang menyebutkan bahwa T. chilonis merupakan agen hayati yang paling sukses dalam mengendalikan populasi penggerek batang tebu di Filipina, India, dan Taiwan. Sedangkan tingkat parasitasi T. japonicum dapat kategorikan cukup rendah, hal ini teramati dari banyaknya jumlah larva C. auricilius yang menetas. Perbandingan tingkat parasitasi T. chilonis dan T.
japonicum diperlihatkan pada Tabel 1.
Pendedahan bersamaan (asosiasi) T. chilonis dan T. japonicum akan menurunkan tingkat parasitasi T. chilonis terhadap C. auricilius. Pada saat telur C. auricilius hanya didedahkan dengan T. chilonis maka jumlah telur yang terparasit sangat tinggi, namun ketika telur C.
auricilius didedahkan dengan T. chilonis dan T. japonicum secara bersamaan maka jumlah
terparasit sangat rendah. Menurunnya tingkat parasitasi T. chilonis terhadap C. auricilius mungkin dapat terjadi karena keduanya saling berkompetisi untuk memperebutkan inang, sehingga menurunkan daya serang terhadap telur C. auricilius. Irwan (1992) menyatakan bahwa kompetisi dapat berbentuk perebutan langsung terhadap inang yang terbatas (resource
competition) atau saling menyakiti antar kedua individu secara langsung dengan kekuatan fisik
(interference competition). Kompetisi diantara dua atau lebih spesies organisme terhadap sumber daya akan menimbulkan efek yang merugikan kedua belah pihak. Driesche, et al. (2008) menyebutkan bahwa serangan musuh alami lain akan mempengaruhi keberhasilan introduksi parasitoid. Musuh alami lokal merupakan salah satu ancaman untuk upaya adaptasi musuh alami yang diintoduksikan (Herlina, 2010).
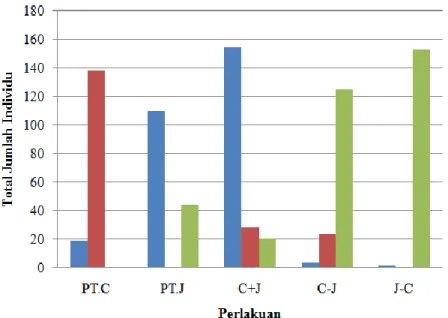
Jeda waktu parasitasi dua parasitoid berpengaruh terhadap keberhasilan pengendalian hayati. Pendedahan bersamaan (asosiasi) T. japonicum dengan T. japonicum akan menurunkan tingkat parasitasi terhadap telur, namun pendedahan T. chilonis dan T. japonicum dengan jeda satu jam dapat meningkatkan jumlah parasitasi telur. Hal ini dapat diamati pada perlakuan pendedahan bergantian T. chilonis dan T. japonicum dalam Tabel 1 dan Gambar 1.
Tabel 1. Nilai Rerata (Mean) Jumlah Imago T.chilonis, T. japonicum, dan Larva C. auricilius pada Perlakuan Pendedahan T. chilonis dan T. japonicum
Perlakuan Pendedahan Spesies yang Muncul
T. chilonis T. japonicum C. auricilius
Tunggal (PT) T.chilonis 45,50
e
7,00a 23,40a
T. japonicum 14,00a 24,75c 38,15b
Bersamaan (PB) 27,90d 17,15b 40,60b
Bergantian dengan parasitoid primer
T.chilonis 26,10c 37,50d 13,55a
T. japonicum 14,00b 41,10d 11,80a
Catatan:
6 Perlakuan pendedahan bergantian dengan parasitoid primer T. chilonis (C→J) dan pendedahan bergantian dengan parasitoid primer T. japonicum (J→C) memiliki tingkat parasitasi yang relatif sama tinggi, hal ini dapat diamati dari jumlah larva C. auricilius yang menetas sangat sedikit. Perlakuan pendedahan bergantian dengan parasitoid primer T. chilonis (C→J) dan pendedahan bergantian dengan parasitoid primer T. japonicum (J→C) dianggap sama baik untuk diterapkan dalam pengendalian C. auricilius. Pada perlakuan pendedahan bergantian dengan parasitoid primer T. chilonis (C→J), imago spesies T. chilonis, T. japonicum dan larva C. auricilius berhasil hidup, namun pada perlakuan pendedahan bergantian dengan parasitoid primer T.
japonicum (J→C), hanya imago T. japonicum dan larva C. auricilius yang berhasil hidup
sedangkan imago T. chilonis tidak muncul (Gambar 1). Hal tersebut mungkin terjadi karena T.
chilonis mampu mendeteksi telur yang sudah terparasit oleh T. japonicum, sedangkan T. japonicum tidak mampu mendeteksi telur yang sudah terparasit atau mungkin juga dapat
mendeteksi namun tetap meletakkan telurnya di dalam inang telur yang sudah terparasit (hyperparasitoid).
Gambar 1. Perbedaan Macam Spesies dan Jumlah Individu yang Muncul pada Perlakuan Pendedahan Telur C. auricilius oleh T. chilonis dan T. japonicum
Keterangan:
Telur yang sudah terparasit dapat terdeteksi karena adanya penandaan (marker) telur oleh parasitoid primernya. Penandaan yang dilakukan ada dua yaitu penandaan internal dan eksternal (Vinson, 1997). Penandaan eksternal dapat dideteksi dengan menggunakan antena, sedangkan penandaan internal dideteksi dengan mengintroduksikan ovipositor betina (Nufio & Papaj, 2001). Penandaan eksternal digunakan untuk menghindari terjadinya superparasitisme,
: Total jumlah larva C. auriclius : Total jumlah imago T.chilonis : Total jumlah imago T. japonicum
7 sedangkan penandaan internal sangat penting digunakan dalam mengurangi terjadinya hyperparasitisme (Vinson, 1976). Vinson (1997) menambahkan bahwa Trichogrammatidae menguji inang telurnya dengan menyentuhkan antena diatas permukaan telur untuk menguji kualitas dan kuantitas telur, sehingga dapat memastikan jumlah telur yang akan diintoduksikan.
Pada penelitian sebelumnya diinformasikan bahwa beberapa familia Hymenoptera seperti Platygastridae, Formicidae, Anthophoridae dan Apidae umumnya memberikan penanda pada telur yang sudah diparasiti dengan memproduksi substansi lipidic dari kelenjar Dufour (Rosi, et
al., 2001). Substansi lipidic yang diproduksi dari kelenjar Dufour berfungsi sebagai peringatan
tanda bahaya (Gökçen, 2002). Substansi lipidic mungkin juga diproduksi oleh familia Trichogrammatidae ordo Hymenoptera misalnya T. japonicum, ini dapat dibuktikan dengan tidak adanya kemunculan T. chilonis pada perlakuan pendedahan bergantian dengan parasitoid primer
T. japonicum (J→C). Trichogramma chilonis diduga mampu mendeteksi substansi lipidic yang
dikeluarkan oleh T. japonicum, sehingga T. chilonis tidak meletakkan telurnya ke dalam inang.
Trichogramma chilonis mungkin juga dapat memproduksi substansi lipidic untuk
memberi penanda pada inang yang sudah diparasiti, namun penanda yang diberikan oleh T.
chilonis tidak terdeteksi dengan baik oleh T. japonicum. Hal tersebut dapat teramati pada
perlakuan pendedahan bergantian dengan parasitoid primer T. chilonis (C→J).
Gambar 2. Hasil Pembedahan Telur C. auricilius yang telah Terparasit T. chilonis dan T.
japonicum; A. Perlakuan Pendedahan Bersamaan (PB), 1) Dua telur T. chilonis usia 6 hari, 2) T. japonicum usia 8 hari; B. Perlakuan Pendedahan Bergantian dengan
Parasitoid Primer T. chilonis (C→J), 1) Kulit Telur C. auricilius, 2) Dua T. chilonis usia
8 hari; C. Perlakuan Pendedahan Bergantian dengan Parasitoid Primer T. japonicum
(J→C), 1) T. japonicum usia 8 hari, 2) Dua T. japonicum usia 8 hari
A B C 1 2 1 2 1 2
8 Imago T. japonicum tetap muncul pada perlakuan pendedahan bergantian dengan parasitoid primer T. chilonis (C→J), dan jumlah kemunculan imago T. japonicum jauh lebih tinggi jika dibandingkan dengan jumlah kemunculan T. chilonis. Telur T. japonicum yang tetap diletakkan dalam inang (telur) yang sudah terparasit oleh T. chilonis akan mengakibatkan adanya kompetisi nutrisi oleh kedua embrio T. chilonis dan T. japonicum di dalam inang. Trichogramma
japonicum yang muncul dari inang yang sebelumnya telah diparasiti oleh T. chilonis
membuktikan bahwa T. japonicum mampu mengalahkan persaingan nutrisi dengan T. chilonis. Pada penelitian ini, T. japonicum mungkin berperan sebagai Hyperparasitoid karena T.
japonicum mampu meletakkan telur pada inang yang sudah terparasit dan mampu mengalahkan
kompetisi dengan T. chilonis di dalam inang. Peluang kemenangan T. japonicum pada kompetisi juga dapat terjadi karena jeda parasitasi antara parasitoid primer dan parasitoid sekunder yang terlalu dekat (1 jam).
Perkembangan embrio T. chilonis dan T. japonicum didalam satu telur dibuktikan dengan membedah telur pada perlakuan pendedahan bersamaan T. chilonis dan T. japonicum (PB), pendedahan bergantian dengan parasitoid primer T. chilonis (C→J), dan pendedahan bergantian dengan parasitoid primer T. japonicum (J→C), dengan usia parasitasi 6 dan 8 hari. Hasil pembedahan ditampilkan pada Gambar 2. Pada telur yang dibedah belum ditemukan adanya dua spesies di dalam satu telur. Dua spesies tidak ditemukan di dalam satu inang telur diduga karena adanya kompetisi kedua spesies ketika masih pada awal perkembangan telur, terdapat satu telur dari satu jenis spesies yang mampu berkompetisi untuk memperoleh nutrisi paling tinggi.
Pengambilan nutrisi dilakukan pada saat Trichogramma masih dalam fase larva. Larva berada pada fase aktif dalam siklus hidup Trichogramma. Pada saat berada pada fase aktif tersebut dibutuhkan tambahan nutrisi berupa protein dari yolk telur C. auricilius. Telur serangga umunya telah dicukupi kebutuhan nutrisinya sampai menetas menjadi larva sehingga telur dapat diletakkan pada semua tempat, sedangkan telur parasitoid Trichogramma diletakkan didalam telur inang yang spesifik untuk memenuhi kebutuhan nutrisi. Kebutuhan nutrisi dibutuhkan pada masa perkembangan Trichogramma. Fase aktif Trichogramma berada di dalam inang telur sehingga pada saat keluar dari inang telur, Trichogramma telah menjadi imago.
Hasil penelitian memiliki kemiripan hasil dengan penelitian Earl & Graham (1984) yaitu salah satu dari spesies parasitoid yang berkompetisi akan tereliminasi dan tidak muncul. Penelitiannya mempelajari kompetisi antara T. remus dan Chelonus insularis (Hymenoptera: Braconidae) pada telur Spodoptera exigua (Lepidoptera: Noctuidae) dengan menguji kompetisi keduanya dan mengacak parasitoid primernya. Hasil dari penelitian tersebut menyebutkan bahwa
Chelonus insularis dieliminasi oleh T. remus, sehingga tidak ditemukan Chelonus insularis.
Meskipun demikian, tidak disebutkan bahwa peristiwa itu adalah hyperparasitisme, tetapi antara larva T. remus dan larva Chelonus insularis terjadi kompetisi.
Penelitian ini memberikan informasi bahwa teknik yang tepat dalam memanfaatkan T. chilonis dan T. japonicum untuk mengendalikan hama penggerek tebu yaitu dengan cara melepaskan T.
9
PENUTUP Simpulan
Perlakuan pendedahan tunggal hanya oleh T. chilonis (PT.C) menunjukkan tingkat parasitasi yang tinggi jika dibandingkan dengan perlakuan pendedahan tunggal hanya oleh T.
japonicum (PT.J). Pada saat T. chilonis dan T. japonicum didedahkan bersamaan, tingkat
parasitasi rendah, jumlah larva C. auricilius yang menetas tinggi, jumlah T. chilonis dan T.
japonicum yang muncul rendah. Pendedahan T. chilonis dan T. japonicum bergantian dengan
jeda satu jam dapat meningkatkan parasitasi terhadap telur C. auricilius. Trichogramma
japonicum muncul lebih dominan muncul pada perlakuan pendedahan bergantian dengan jeda
satu jam.
DAFTAR RUJUKAN
Alba, M.C. 1989. Use of Natural Enemies for Controlling Sugarcane Pests in the Philippines. Pusat Penelitian Pertanian Nasional, Tsukaba, Jepang, (Online), diakses 23 september 2015.
Clausen. 1940. Enthomophagus Insect. First Edition. New York. Mc Graw Hill Book.
Driesche, R. V, Center, T., & Hoddle, M. 2008. Control of Pests and Weeds by Natural Enemies:
An Introduction to Biological Control. Blackwell Publishing, UK, (Online), diakses 24
September 2015.
Earl, S.L., & Graham, H.M. 1984. Interaction between Chelonus insularis Cresson and
Telenomus remus Nixon, Parasitoid of Spodoptera exigua (Hubner). Southwest Entomol,
(Online), 9: 326-333, diakses 26 September 2015.
Gökçen, O. A., Morgan, E. D., Dani, F. R., Agosti, D., & Wehner, R. 2002. Dufour Gland Contents of Ants of the Cataglyphis bicolor Group. Journal of Chemical Ecology, (Online), Volume 28, Issue 1, pp 71-87, diakses 12 Maret 2015.
Herlina, L. 2010. Introduksi Parasitoid, Sebuah Wacana Baru dalam Pengendalian Hama Kutu Putih Pepaya Paracoccus marginatus Di Indonesia. Jurnal Litbang Pertanian, (Online), 30(3), 2011, diakses 03 Oktober 2015.
Herlinda, S. 1995. Kajian Trichogrammatoidea bactrae-bactrae Nagaraja (Hymenoptera:
Trichogrammatidae), Parasitoid Telur Etiella zinckenella Treitschke (Lepidoptera: Pyralidae). Tesis S2 Institut Pertanian Bogor tidak diterbitkan, Bogor.
Herlinda, S., Thalib, R., Saleh, R.M. 2004. Perkembangan dan Preferensi Plutella xylostella L. (Lepidotera: Plutellidae) pada Lima Tumbuhan Inang. Jurnal Hayati, (Online), 130-134, diakses 12 Januari 2016.
Huffaker, C.B & Messenger, P.S. 1989. Teori dan Praktek Pengendalian Biologis. Penerjemah Soeprapto Mangoendihardjo. UI Press, Jakarta.
Irwan, Z.D. 1992. Prinsip-Prinsip Ekologi dan Organisasi: Ekosistem, Komunitas dan
Lingkungan. Jakarta: Bumi Aksara.
Li, Li Ying. 1994. Worwide use of Trichogramma for Biological Control on Differeent Crops: a
Survey. In: Wagnberg E, Hassan SA (eds) Biological Control with egg parasitoid. CAB
International (Online), Wallingford, pp 37-54, diakses 27 Februari 2016.
Meidalima, D., Herlinda, S., Pujiastuti, Y., & Irsan, C. 2012. Pemanfaatan Parasitoid Telur,
Larva, dan Pupa untuk Mengendalikan Penggerek Batang Tebu. Universitas Sriwijaya.
10 Meilin, A., Hidayat, P., Buchori, D., & Kartosuwondo, U. 2000. Parasitoid Telur pada Hama
Kubis Plutella xylostella (L) (Iponomeutidae). Buletin Hama dan Penyakit Tumbuhan,
(Online), 12(1): 21−26, diakses 24 September 2015.
Nagaraja, H., 1978. Studies on Trichogrammatoidea (Hymenoptera: Trichogrammatidae).
Oriental Ins., (Online), 12: 489-530, diakses 24 September 2015.
Nagarkatti, S. & Nagaraja, H. 1977. Biosystemic of Trichogramma and Trichogrammatoidae species. Annu. Rev. Entomol., (Online), 22: 157-176, diakses 02 Oktober 2015.
Nufio C. R., & Papaj D. R. 2001. Host Marking Behavior in Phytophagous Insects and Parasitoids. Entomol Exp Appl., (Online), 99: 273-293, diakses 27 Februari 2016.
Pusat Data dan Sistem Informasi Pertanian. 2013. Komoditi Tebu, (Online), diakses 25 November 2015.
Pusat Penelitian Perkebunan Gula Indonesia (P3GI). 2008. Konsep Peningkatan Rendemen untuk
Mendukung Program Akselerasi, (Online), diakses 21 Oktober 2015.
Rosi, MC., Isidoro, N., Colazza, S., & Bin, F. 2001. Source of The Host Marking Pheromone in The Egg Parasitoid Trissolcus basalis (Hymenoptera: Scelionidae). J Insect Physiol, (Online), 47: 989-995, diakses 27 Februari 2016.
Soula, B., Goebel, F.R., Caplong, P., Karimjee, H., Tibere, R., & Tabone, E. 2003.
Trichogramma chilonis (Hymenoptera: Trichogrammatidae) as a Biological Control Agent of Chilo sacchariphagus (Lepidoptera : Crambidae) in Reunion Island: Initial Field Trials, (Online), diakses 05 September 2015.
Sudarsono, H. 2011. Kajian Beberapa Karakteristik Biologi Penggerek Batang Tebu Berkilat
Chilo auricilius dan Parasitoidnya (Trichogramma chilonis). Seminar Hasil Penelitian &
Pengabdian Kepada Masyarakat Unila, (Online), ISBN 978–979‐8510‐22‐9, diakses 28 September 2015.
Vinson, S. B. 1976. Host Selection by Insect Parasitoids. Annu Rev Entomol, (Online), 21: 109-133, diakses 27 Februari 2016.
Vinson, S. B. 1997. Comportamento de Seleção Hospedeira de Parasitóides de Ovos, com
Enfase na Família Trichogrammatidae. Piracicaba: FEALQ, p., (Online), 67-120, diakses