IDENTIFIKASI MORFOLOGIS DAN UJI AKTIVITAS MIKROBA RUMEN
DARI HEWAN-HEWAN RUMINANSIA YANG TELAH TERADAPTASI
PADA SUBSTRAT SELULOSA DAN HEMISELULOSA
Amn
tus
THALiB,Y.WjDiAWATt, H. HAmID,danMuLYANi Balai Penelitian Ternak P.O. Box 221, Bogor 16002ABSTRAK
Telah dilakukan skrining mikroba fibrolitik melalui proses adaptasi pada substrat holoselulosa, selulosa dan hemiselulosa. Sumber mikroba diambil dari rumen sapi, kerbau, domba, kambing dan rusa. Mikroba diadaptasi pada substrat secara anaerobik pada suhu 39°C dan pH 6,9. Inkubasi dilakukan hingga diperoleh kurva plateu (produksi gas kumulatif vs waktu). Identifikasi dan populasi mikroba (bakteri dan protozoa) dilakukan setelah masa adaptasi tercapai. Selanjutnya dilakukan uji aktivitas mikroba ntu mencema substrat jerami padi dan rumput Gajah. Hasil identifikasi mikroba memperlihatkan bahwa baktei rumen dari semuajenis hewan terdiri dari bentuk-bentuk coccus, batang dan crostridial, dan bentuk rosette terdapat pada mikroba domba. Protozoa rumen dari semua jenis hewan terdiri dari entodinium sp., dan protozoa dasytricha sp. terdapat pada kambing dan rusa, poyplastron sp pada kerbau dan pada sumber mikroba domba terdapat oligoissotricha sp dan eremoplastron sp. Secara berturut-turut nilai kecernaan bahan kering (DMD)jerami padi menunjukkan=27,72; 26,96; 25,45;24,39 dan 24,34% untuk sumber mikroba rusa, kerbau kambing, sapi dan domba. Sedangkan nilai DMD rumput Gajah berturut-turut=44,07; 41,61; 45,05; 40,26; dan 40,39% untuk sumber mikroba rusa, kerbau, kambing, sapi dan domba.
Kats kunei:Bakteri, protozoa, sapi, kerbau, domba, kambing, rusa PENDAHULUAN
Kinerja sistem pencemaan rumen (pada temak ruminansia) ditentukan oleh aktivitas mikroba rumen. Sekitar 70-85% bahan pakan yang dikonsumsi temak ruminansia dicema dengan bantuan mikroba. Mikroba rumen terdiri dari golongan protozoa, bakteri dan sejumlah kecil jamur. Dari ketiga golongan mikroba rumen, bakteri adalah pencema ekstensif serat kasar di dalam sistem pencemaan rumen yakni merupakan sumber utama penghasil enzim selulolitik dan hemiselulolitik (DEMEYER, 1981). Peranan mikroba selulolitik dan hemiselulolitik sangat penting dalam proses penemmaan serat pakan.
Karakteristik interaksi suatu bakteri dengan partikel substrat yang akan dicema dapat dipengaruhi oleh bentuk morfologis dan tipe struktur dinding sel dari bakteri tersebut (LING, 1990; JoANY, 1991). Aktivitas bakteri dalam mendegradasi pratikel bahan makanan diawali oleh proses penempelan. Spesies bakteri utama yang menyerang portikel bahan makanan adalah bakteri selulolitik (yakni Ruminococcus albus, R. flavefaciens, dan bacteroides succinogenes). B. succinogenes menempel pada partikel makanan lebih kuat daripada Rumino cocci. Bakteri selulolitik lainnya adalah Butyrivibrio fibri solvens dan Clostridium lochheadii. Disamping sebagai oencema selulosa, Rumino cocci dan B. fibrisolvens juga sebagai pencema hemiselulosa (HUNGATE, 1966) B. fibrisolvens bahkan lebih dikenal sebagai pencema hemiselulosa (STEWART, 1988). Pencema hemiselulosa lainnya adalah Bacteroides ruminicola, B. amylogenes, dan Eubacterium (HUNGATE, 1966). Secara morfologis, bakteri selulolitik terpenting dan lazim dijumpai dalam rumen, terbagi kedalam bentuk coccus (R.albus dan R. flavefaciens) dan bentuk batang (B. succinogenes, B. fibrisolvens, dan C. locheadii), dimana tipe struktur dinding sel dari masing-masing kelompok
Seminar Nasional Peternakan dan Veteriner 2000
morfologis ini berturut-turut adalah Gram-positip dan Gram-negatip(HUNGATE, 1966; OGIMOTOdap IMAI, 1981).
Protozoa rumen terbagi kedalam kelompok flagellata dan ciliata, tapi hampir semua protozoa rumen adalah protozoa ciliata(OGIMOTOdanIMAI, 1981).
Tidak semua jenis protozoa mempunyai aktivitas selulolitik atau berpengaruh positip terhadap kecernaan serat, bahkan ada protozoa yang berpengaruh negatip dalam sistem pencernaan rumm Protozoa dari sub-kelas holotrich dan Isotricha spp. menyebabkan penurunan pH rumen dan mengganggu pencernaan pakan berserat (WIDYASTUTI, 1994). Protozoa entodinium berpengaruh positip terhadap pencernaan pakan berserat. Namun beberapa entodinium berukuran kecil dilakukan pengaruhnya terhadap pencernaan serat. Protozoa rumen berukuran besar dapat berperan sebagai predator bagi bakteri baik secara langsung maupun tidak langsung.
Nilai kualitatif dan kuantitatif/komposisi mikroba rumen dari hewan ruminansia yang berbeda diasumsikanjuga berbeda, yang tentu juga memberikan aktivitas yang berbeda dalam mencerna serat pakan. Untuk itu dilaporkan dari percobaan ini mengenai aspek kualitatif dan kuantitatif mikroba (bakteri dan protozoa) yang bersumber dari hewan-hewan ruminansia.
MATERI DAN METODE
Substrat yang digunakan adalah holoselulosa, selulosa dan hemiselulosa yang diisolasi dari dinding seljerami padi menurut prinsip prosedurBROWNING (1963).
Jerami padi yang masih segar dipotong-potong (± 5 cm), dikeringkan pada suhu60°C selama 3 hari, kemudian digiling halus.
Isolasi dinding sel
Dinding sel diperoleh dengan menghilangkan fraksi ekstraktifrlya. 200 g serbuk jerami (40 mesh) diekstraksi dengan 500 ml pelarut etanol-benzen (1 :2) secara sokletasi. Ekstraksi dilakukan sampai pelarut terlihat jernih, kemudian ekstraksi dilanjutkan dengan larutan bufer fosfat (pH 7). Residu hasil penyaringan dicuci dengan aquades, dibilas dengan etanol dan eter, kemudian dikeringkan dalam oven 105°C.
Isolasi holoselulosa
Holoselulosa diperoleh dengan cara penghilangan lignin dari fraksi yang telah bebas dari fraksi ekstraktif. Residu sebagai fraksi dinding sel dipanaskan dalam 3 L aquades pada suhu 70-80°C, dengan sambil diaduk kuat ditambahkan 22 ml asam asetat glasial,66 g natrium klorit, dan 5 ml 1 oktanol. Setelah residu terlihat putih, pengadukan dihentikan, kemudian disaring dan dicuci dengan aquades. Residu selanjutnya dikeringkan dalam oven 105°C.
Isolasi hemiselulosa dan selulosa
Hemiselulosa dan selulosa diperoleh dari proses ekstraksi fraksi holoselulosa dengan basa kuat. Bagian yang terlarut hemiselulosa dan bagian yang tidar larut adalah selulosa. 100 g holoselulosa 342
Adaptasi mikroba pada substrat
Uji aktifitas mikroba
ditempatkan dalam erlenmeyer 5 L, sambil dialiri gas nitrogen ditambahkan 1 L kalium hidroksida 5% pada suhu 20°C dan diaduk. Setelah didiamkan beberapa saat campuran disaring clan residu dibilas dengan 200 ml kalium hidroksida 5%, lalu diikuti dengan 600 ml aquades.
Filtrat dan hasil bilasan ditambah 3 L etanol 95% sambil diaduk dan ditambahkan secara perlahan asam asetat sampai campuran menjadi netral. Endapan yang terbentuk adalah hemiselulosa, lalu dikeringkan dengan freeze drier. Residu hasil penyaringan selanjutnya diperlukan dengan cara yang sama kalium hidroksida 24% untuk mengambil fraksi hemiselulosa yang tersisa. Residu terakhir adalah selulosa dan dikeringkan dalam oven 105°C.
Mikroba yang bersumber dari rumen kerbau, sapi, kambing, domba dan rusa masing-masing diadaptasikan pada substrat holoselulosa, selulosa dan hemiselolosa secara anaerobik pada kondisi suhu 390C dan pH 6,9 di dalam media fermentasi. Selama masa adaptasi masing-masing mikroba pada setiap substrat dilakukan pengukuran produksi gas. Media fermentasi clan pengukuran produksi gas didasarkan pada prosedurTHEODORou danBROOKS (l990). Masa adaptasi bagi setiap mikroba pada setiap substrat berlangsung hingga diperoleh kurva (produksi gas versus waktu) plateu dengan nilai slope yang relatif konstan. Selanjutnya setalah masing-masing adaptasi selesai dilakukan penghitungan populasi protozoa dengan metode whitlock universal dan populasi bakteri dengan metode roll tube menurut prosedur OGIMOTO dan ImAl (l981). Secara kualitatif masing-masing mikroba yang teradaptasi pada setiap substrat dilakukan menurut prosedur OGIMOTO clan ImAi (1981) untuk protozoa clan bakteri.
Akrifitas masing-masing mikroba yang telah beradaptasi pada holoselulosa, clan untuk mencerna jerami padi clan rumput Gajah diuji secara in vitro menurut prosedur THEODOROu dan BROOKS (1990) dengan waktu inkubasi 48 jam. Prosedur mencakup inkubasi substrat dengan penambahan inokulum (mikroba yang telah teradaptasi pada holoselulosa) sebanyak 10 ml kedalam medium fermentasi pada suhu 39°C clan pH 6,9. Komposisi media terdiri dari 86 bagian volume larutan basal (mengandung bufer, makromineral dan makromineral), dan 4 bagian volume larutan pereduksi. Nilai in vitro keecemaan bahan kering (in vitro DMD) substrat ditetapkan berdasarkan banyaknya residu hasil fermentasi sebstrat melalui tahap penyaringan clan pengeringan suhu 105°C selama 1 malam dan dilanjutkan dengan penetapan OMD.
HASIL DAN PEMBAHASAN Perolehan serbuk holoselulosa, selulosa clan hemiselulosa
Pemisahan holoselulosa, selulosa clan hemiselulosa dawali dengan pemisahan dinding sel dari bahan serbuk jerami padi. Serbuk holoselulosa kering diperoleh 86,54% dari sernuk dinding sel
Seminar Nasional Peternakan dan Veteriner 2000
Adaptabilitas mikroba rumen pada substrat holoselulosa, selulosa, dan hemiselulosa
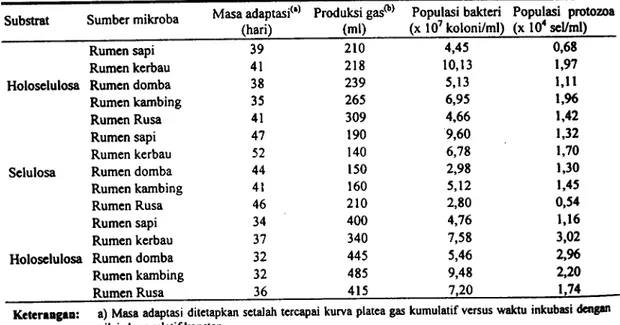
Adaptabilitas mikroba rumen dari sapi, kerbau, domba, kambing, dan rusa pada substrat holoselulosa, selulosa, dan hemiselulosa diperlihatkan pada Tabel 1 . Waktu yang diperlulcm oldt masing-masing mikroba untuk beradaptasi pada substrat ditetapkan berdasarkan pada pencapaian kwva plateau (produksi gas kumulatif versus waktu inkubasi) yang relatiftidak lagi mengalarni perubahan nilai slope dengan kura plateau sebelumnya.
Waktu yang diperlukan mikroba untuk beradaptasi pada substrat (holoselulosa, selulosa dan hemiselulosa memperlihatkan kecendrungan dengan urutan: selulosa > holoselulosa > hemiselulosa). Hal ini mentmjukkan bahwa hemiselulosa adalah yang termudah didegradasi oleh mikroba dibandingkan dengan 2 substrst lainnya; dan kencendrungan ini didukung oleh produksi gas dimana produksi gas tertinggi secara umum diperlihatkan substrat hemiselulolitik yang diikuti berikutnya oleh holoselulosa dan selulosa.
Jumlah bakteri dan protozoa pada saat masa adaptasi telah tercapai memperlihatkan suatu angka yang jauh dibawah jumlah normal bakteri maupun protozoa di dalam rumen. Hitungan total bakteri di dalam rumen sapi berkisar 109-10 10 koloni/ml cairan rumen(THALIB et al., 2000).
Jtunlah normal protozoa di dalam rumen berkisar 105-106 sel per ml cairan rumen (OGtmoTo danIMAi, 1981). Jumlah bakteri dan protozoa untuk semua hasil adaptasi masing-masing berkisar
107 koloni/ml dan 104 sel/ml (Tabel 1). Jumlah bakteri dan protozoa yang terhitung untuk semua hasil adaptasi di duga merupakan suatu komposisi protozoa selulolitik clan hemiselulolitik. MiNATo et al. (1990) melaporkan dari hasil percobaannya bahwa jumlah bakteri total dalam rumen berkisar >109 koloni/ml cairan rumen sapi clan jumlah bakteri selulotik berkisar 107-108 koloni/ml cairan rumen.
Tabel 1 . Masa ddaptasi mikroba pada substrat holoselulosa, selulosa clan hemiselulosa serta populasi bakteri clan protozoa pada akhir masa adaptasi
Substrat Sumber mikroba Masa adaptasit`l Produksi gas(b) Populasi bakteri Populasi protozoa(hari) (ml) (x 107 koloni/ml) (x 104 sel/ml)
Rumen sapi 39 210 4,45 0,68
Rumen kerbau 41 218 10,13 1,97
Holoselulosa Rumen domba 38 239 5,13 1,11
Rumen kambing 35 265 6,95 1,96
Rumen Rusa 41 309 4,66 1,42
Rumen sapi 47 190 9,60 1,32
Rumen kerbau 52 140 6,78 1,70
Selulosa Rumen domba 44 150 2,98 1,30
Rumen kambing 41 160 5,12 1,45
Rumen Rusa 46 210 2,80 0,54
Rumen sapi 34 400 4,76 1,16
Rumen kerbau 37 340 7,58 3,02
Holoselulosa Rumen domba 32 445 5,46 2,96
Rumen kambing 32 485 9,48 2,20
Rumen Rusa 36 415 7,20 1,74
Keteraagaa: a) Mssaadaptasi ditetapkan setalah tercapai kurva plateagaskumulatif versus waktu inkubasi dengan
nilai slope relatifkonstan.
344
ldentifikasi bakteri dan protozoa
Bakteri dan protozoa yang terdapat dalam inokulum hasil adaptasi pada substrat holoselulolitik, selulosa dan hemiselulosa diduga tergolong selulolitik maupun hemiselulolitik, dan identifikasi morfologisnya seperti yang terlihat dalam Tabel 2 dan Tabel 3.
Semua mikroba teradaptasi memperlihatkan keragaman spesies bakteri yang hampir sama, namtun komposisi populasinya diduga berbeda satu sarna lain. Bentuk Rosette hanya dijumpai pada sumber mikroba dari rumen domba (Tabel 2). Kecuali sumber mikroba dari rumen rusa, bakteri yang teridentifikasi pada semua sumber mikroba memperlihatkan dominasi bakteri dengan struktur dinding sel tipe Gram negatif. Hal ini sesuai dengan yang dilaporkan LING (1990) bahwa bakteri Gram negatif biasanya yang terbanyak dalam komposisi populasi bakteri, khususnya bila ternak diberi pakan berserat. Suatu hal yang menarik pada sumber mikroba dari rumen rusa dimana tipe struktur dinding sel yang terindentifikasi memperlihatkan keadaan yang seimbang antara Gram positip dan Gram negatif. Suatu indikasi yang mungkin dapat diperkirakan dari hasil identifikasi ini bahwa bakteri tipe Gram positif (dari rumen rusa) dapat beradaptasi dengan serat lebih baik dari pada bakteri tipe Gram positifdari rumen hewan-hewan ruminansia lainnya.
Tabel2. Identifikasi bakteri dari rumen yang telah beradaptasi pada substmt holoselulosa, selulosa dan hemiselulosa
Mikroskrop
3 = cocci in tetrads 8 = coccobacilli 13 =rosette
Sumber mikroba Substrat
Morfologi Gram Keterangan
Rumen sapi Holoselulosa 1-4-5-6
Selulosa 1-6-7-14
Hemiselulosa 1-2-4-5-6-7-8-10-14 D (-)
Rumen kerbau Holoselulosa 1-2-4-5-6-8-10
Selulosa 1-2-4-5-{r-7-8-10-14 + D (-)
Hemiselulosa 1-4-5-6-7-8-10-14
Rumen domba Holoselulosa 1-4-5-6-8-10-14 - + D (-)
Selulosa 1-4-5-6-8 + - D (+)
Hemiselulosa 1-2-6-7-8-10-14 - + D (-)
Rumen kambing Holoselulosa 1-4-6-7-8-10 - + D (-)
Selulosa 1-4-Cr7-8-10-14 - + D (-)
Hemiselulosa 1-5-6-7-8-10 - + D (-)
Rumen rusa Holoselulosa 1-2-5--6-7-8-14 +
Selulosa 1-6-7-8-10 +
Hemiselulosa 1-2-5-fi--7-8
-Keterangan: D = Dominan
I = single cocci 6 = rods with square ends I 1 =treponema 2 = cocci in pairs 7 =rods irith round ends 12 =borrelia
Spesies protozoa yang mampu beradaptasi bersama-sama bakteri pada semua sumber rnikroba memperlihatkan dominasi protozoa entodiniomorphid. Hampir semua protozoa yang teridentifikasi dari semua sumber milaoba termasuk order entodinomorphid, kecuali Oligoisotricha (pada sumber
mikroba domba) dan Dasytricha(pada sumber mikroba kambing dan rusa), dua-duanya termasuk
order Trichostomatida. Protozoa entodinomorphidtergolong protozoa ciliata. Hampir semua pmtozoa
yang terdapat dalam rumen tergolong ciliata (OGwtOTO danIMAi, 1981).Keuntungan dan kerugian dari akfvitas protozoa dalam sistem pencemaan rtunen telah dibahas secara luas oleh JOUANYdanUSHDA (1990). Hampir semua protozoa entodinomorphid memiliki potensi enzunatik untuk mencema selulosa secara efisien, dan secara simultan juga bersifat hemiselulolitik(JOUANYdanUSDA ,1990).Namun sifM predator protozoa terhadap bakten menrpakan kerugian dan kehadiran protozoa dalam sistem pencemmn rumen. Aksi predator protozoa dalam rumen menyebabkan penurunan bakten amilolib& (J(xUANYdan USRDA,1990).
Tabel3. Identifikasi Protozoa dari rumen yang telah beradaptasi pads substrat holoselulosa, selulosa dan hemiselulosa
Aktivitas mikroba beradaptasi
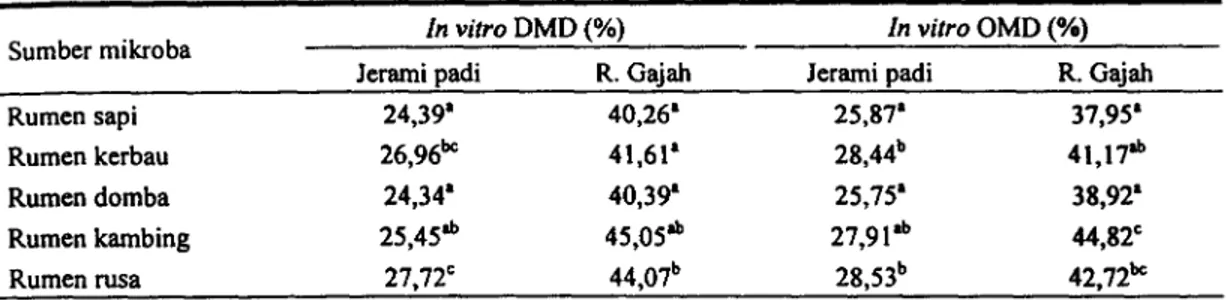
Mikroba dari semua sumber yang digunakan diuji aktivitasnya mencerna substrat jerami padi dan rumput Gajah, dan hasilnya seperti yang terlihat pada Tabel 4.
Nilai kecernaan jerami padi yang tertinggi diberikan oleh sumber mikroba rusa, kerbau dan kambing, sedangkan nilai kecernaan rurnput Gajah tertinggi diberikan oleh sumber mikroba rusa dan kambing. Efektifitas mikroba sebagai pencerna substrat/bahan pakan tidak saja ditentukan oleh
346
Seminar Nasional Peternakan dan Veteriner 2000
Sumber mikroba Subs= _Spesi es
Rumen sapi Holoselulosa Entodinium parvum
Selulosa Entodinium caudatum
Entodinium ogimotoi
Hemiselulosa Entodinium bimastm
Rumen kerbau Holoselulosa Entodinium costatum
Entodiniumparvum
Pol)plastron muldvesiculatum
Selulosa Entodinium aculeatum
Hemiselulosa Entodiniumparvum
Rumen domba Holoselulosa Oligoisotricha bubali
Selulosa Entodinium simplex
Hemiselulosa Eremoplastron bubalus
Rumen kambing Holoselulosa Entodinium parvum
Selulosa Entodinium caudatum
Hemiselulosa Dasytricha ruminandum
Rumen ruse Holoselulosa Dasytricha ruminandum
Selulosa Entodiniumfujitai
Hemiselulosa Entodinium longinucleatum Entodinium parum
komposisi spesies tapi juga oleh komposisi populasi. Dengan demikian diperlukan kombinasi berimbang antara spesies dan populasi, dan setiap jenis substrat/bahan pakan memerlukan kombinasi yang berbeda.
Tabel 4. In vitro DMD dan OMD substrat jerami padi dan rumput Gajah dengan inokulum mikroba yang teradaptasi pada holoselulosa
Keterangan: Perbedaan tanda huruf pada kolom yang sama menunjukkan perbedaan yang nyata (P<0,05) KESIMPULAN
Disimpulkan dari percobaan ini bahwa bakteri dan protozoa dari rumen hewan-hewan ruminansia yang dapat beradaptasi pada substrat selulosa dan hemiselulosa, menunjukkan keragaman spesies yang hampir sama. Sedangkan komposisinya diduga berbeda antar masing-masing jenis rumen hewan ruminansia. Untuk itu perlu dilakukan lebih jauh lagi dari aspek kuantitatifnya untuk dapat mengetahui komposisi populasi dari spesies (bakteri maupun protozoa) yang ada di dalam rumen setiap hewan ruminansia.
DAFTAR PUSTAKA
BROWNING, B.L. 1963. The Chemistry of Wood. Institute of Paper Chemistry Appleton, Wilscounsin. Intersciences Publisher. New York.
DEMEYER, D.I. 1981 . Rumen Microbes and Digestion of Plant Cell Wall. Agricultural and Environment. Elseveir Scientific Publ. Co. Amsterdam. pp. 6-37.
HUNGATE, R.E. 1966. The Rumen and Its Microbes. A.P. Inc. N. Y.
JOUANY, J.P. and K. USHIDA. 1990. Protozoa and fibre digestion in the rumen. In : The Rumen Ecosystem, (Eds HOSwNO et al). Japan Sci. Soc. Press, Tokyo. pp. 139-150.
JuvANY, J .P. 1991 . The Microbial Metabolism and Ruminant Digestion. INRA Editions, Paris.
LING, J.R. 1990. Digestion of bacterial cell walls in the rumen. In : The Rumen Ecosystem (Eds : S. Hoshino, R.Onodera, H. Minato and H. Itabashi). Jap. Sci. Soc. Press. Tokyo. pp. 83-90.
MINATo, H., E. MIYAGAWA, and T. SUTo. 1990. Techniques for analysis of rumen microbial ecosystems . In The Rumen Ecosystem, (Eds: S. Hoshino, R. Onedera, H. Minato, and Itabashi). Japan Scienties Press, Tokyo. pp. 3-12.
OGIMOTO,K. and S. ImAI. 1981 . Atlas ofRumen Microbiology. Jap. Sci. Press, Tokyo.
STEWART, C.S. 1988. The rumen bacteria. In: The Rumen Microbial Ecosystem. (Ed. P.N. Hobson). Elsevier Sci . Publ. Ltd. England. pp. 21-75.
Sumber mikroba In vitro Jerami padi DMD (%) R. Gajah In vitro Jerami padi OMD(%) R. Gajah Rumen sapi 24,39' 40,26' 25,87' 37,95' Rumen kerbau 26,96` 41,61' 28,44° 41,17'' Rumen domba 24,34' 40,39' 25,75' 38,92' Rumen kambing 25,45'' 45,05'' 27,91'' 44,82` Rumen rusa 27,72` 44,07' 28,53' 42,72'`
Seminar Nasional Peternakan dan Veteriner 2000
TI-IALIB, A. J. BESTARI, Y. WIDIAWATI, H. HAMID dan D. SUHERMAN. 2000 . Pengaruh perlakuan silase jerami padi dengan mikroba rumen kerbau terhadap daya cerna dan ekosistem rumen sapi . J./lmu Ternak Vet. 5(1):1-6.
T HEODOROU, M.K. and BROOKS. 1990. Evaluation of a. new laboratory procedure for estimating the fermentation kinetics of tropical feeds. Annual Report. AFRC . Inst. Hurley, Maidenhead, U.K.
WmyAsTtrrt, Y. 1994. Pengaruh adaptasi dan transfer terhadap aktivitas kecernaanRuminococcus favefaciens 17 . Pros . Seminar Hasil Penelitian dan Pengembangan Bioteknologi II . Cibinong. pp. 313-319.