Summary Dependence of shoot morphology on relative ir-radiance (diffuse site factor, ad) and total tree height (TTH) was studied in a natural canopy of Picea abies (L.) Karst. Surface area of foliar elements was characterized by total needle area (TLA, all-sided needle surface), projected needle area (PLA, area of detached needles laid flat on a horizontal surface), and vertical and horizontal silhouette shoot areas (SSA0 and SSA90, defined as shadows cast by shoots with fixed horizontal incli-nations of 0° and rotation angles of 0° and 90°, respectively).
With increasing irradiance and tree size, packing of needles (needle number per unit shoot length, Ls) and packing of TLA (TLA/Ls) increased, but packing of SSA0 (SSA0/Ls) was invari-ant. Accordingly, the SSA0 to TLA ratio (STAR0) decreased, signifying that the proportion of TLA exposed to direct solar radiation declined, and thus the cost of SSA0 in terms of TLA increased with increasing ad and TTH. Factors causing costlier SSA0 at high irradiances and in tall trees included increases in needle packing, changes in needle morphology (increases in needle dry weight per TLA and decreases in TLA/PLA), and changes in needle position and angle of needle inclination.
The weight of the shoot axis (Ma) was used as an estimate of biomass costs for mechanical support of the needles and water supply to the needles. Foliage-area-based needle-support costs (NSCa), defined as Ma per unit foliage surface (TLA, PLA or SSA0) increased with ad and were not related to TTH, whereas foliage-weight-based needle-support costs (Ma/Mn) decreased with increasing TTH and were independent of ad.
Keywords: conifers, foliage area, LAI, leaf morphology, needle packing, silhouette area, STAR.
Introduction
Because annual production of a stand quasi-linearly depends on intercepted shortwave solar radiation (Jarvis and Leverenz 1983, Linder 1985), canopy attributes, such as leaf inclination and clumping, which influence light absorption at constant values of leaf area index (LAI, foliage area per ground area) and thus determine the effectiveness of unit foliage area for light interception, are important stand parameters.
If canopy foliage elements are distributed randomly, trans-mitted radiation (I) decreases exponentially with increasing stand LAI according to Lambert-Beer’s law (e.g., Monsi and Saeki 1953):
I = I0e−k LAI /cosα , (1)
(where I0 is the irradiance above the stand, k is the extinction coefficient, and α is the solar zenith angle), hence permitting simultaneous estimation of canopy transmittance and LAI on the basis of light measurements alone, provided that the mean leaf inclination determining k is known. In coniferous trees, however, needles are grouped on shoots, and canopy models that assume random foliage distribution significantly underes-timate stand LAI (Gower and Norman 1991, Chen and Black 1992). To account for needle clumping, a conifer shoot is considered as the foliage element corresponding to a leaf of a broad-leaved species (Oker-Blom 1986, Leverenz and Hinck-ley 1990). Accordingly, a detailed knowledge of shoot archi-tecture and identification of patterns of variation are needed to estimate foliage area and describe the light environment in a coniferous canopy (Oker-Blom and Smolander 1988, Gower and Norman 1991). Furthermore, dry matter production may be directly related to shoot architecture, because the ability to change shoot structure with respect to shading within the canopy correlates closely with the efficiency of light-intercept-ing foliage display and the upper limit of LAI the species may attain (Leverenz and Hinckley 1990, Leverenz 1992).
Several qualitative modifications in leaf morphology and functioning along an irradiance gradient across the canopy have been reported for leaves (e.g., Björkman 1981), but much less is known about morphological acclimation in conifer shoots with respect to irradiance. The ratio of shoot silhouette to total needle surface area (STAR), which depends on several shoot parameters, including needle orientation and inclination (Carter and Smith 1985) and needle overlapping (Carter and Smith 1985, Oker-Blom and Smolander 1988), has been used to characterize shoot morphology and needle clumping (e.g., Oker-Blom and Smolander 1988). The STAR is equivalent to the extinction coefficient in Equation 1 (Oker-Blom and
Smo-Effects of light availability and tree size on the architecture of
assimilative surface in the canopy of Picea abies: variation in shoot
structure
ÜLO NIINEMETS
1,2and OLEVI KULL
11 Institute of Ecology, Estonian Academy of Sciences, 40 Lai Street, EE2400 Tartu, Estonia
2
Present address: LS Pflanzenökologie, BITÖK, Universität Bayreuth, Postfach 10 12 51, 95448 Bayreuth, Germany
Received February 2, 1995
lander 1988). In Picea abies (L.) Karst., consideration of only changes in needle structure and needle number per shoot length predicted a linear decrease in STAR with increasing irradiance (Niinemets and Kull 1995). We conclude, therefore, that relative irradiance is a relevant determinant of shoot struc-ture, and have undertaken to identify explicitly the patterns of variation in shoot traits along a light gradient in the canopy.
In conifers, needle morphology is also affected by tree age (Greenwood 1984, Steele et al. 1989) and dimensions (Niine-mets and Kull 1995). Inasmuch as large environmental gradi-ents and gradigradi-ents of water potential exist in the crowns of tall trees (Hellkvist et al. 1974) and the fraction of annual produc-tion allocated for supporting tissues increases with total tree height (Givnish 1988), it is likely that shoot morphology as well as partitioning of assimilative and supporting structures within the shoot also change with changing tree size. Accord-ingly, we have examined variations in shoot structure and partitioning of shoot biomass with respect to a natural gradient of relative irradiance and to total tree height in a stand of P. abies.
Material and methods
The study was conducted in a nemoral spruce forest, located on a plateau-like crest of a drumlin with brown pseudopodzolic soil, at Voore Ecological Station, Estonia (58°44′ N, 26°45′ E, elevation 90 m above sea level), at the beginning of August 1989. Frey (1977) gives a thorough description of the study site.
Current-year shoots at terminal branch positions were col-lected from the southern aspect of six variously exposed P. abies trees. At two to five canopy heights per tree, four to seven shoots per canopy height were taken, and means of all shoot parameters for a given canopy height were determined. Total tree height ranged from 1.6 to 35.5 m, and relative sampling height (Rh), i.e., sampling height per total tree height, ranged from 0.07 to 0.96, but the maximum Rh per tree was never less than 0.78.
The hemispherical photographic technique (Anderson 1964, Nilson and Ross 1979) was used to estimate relative irradiance at the sample locations. The proportion of canopy gaps was measured with respect to zenith angle, and the diffuse site factor (ad, proportion of penetrating diffuse solar radiation) was calculated. The value of ad is equal to unity above the canopy and to zero in complete shade without any canopy gaps. Further details about shoot sampling and estimation of ad are given in Niinemets and Kull (1995).
Changing patterns of water relations and increasing gradi-ents of environmental parameters with increasing tree size were assumed to cause changes in shoot structure. Therefore, total tree height (TTH), which was not significantly related to ad (P > 0.2), was included in the statistical models as a second independent variable.
Shoot silhouette area (SSA, shadow cast by a shoot) was measured with an optical planimeter. The shoot axis was kept perpendicular to the light source, and vertical (SSA0) and horizontal (SSA90) silhouette areas (Boyce 1993, Figure 1)
were measured. There are an infinite number of silhouette areas with respect to different sun elevations and shoot incli-nations; however, we used a fixed angle of 90° between view direction and shoot axis and only studied the variation in SSA with rotation angle. A shoot with a horizontal inclination of 0° has a silhouette area that is equal to SSA0 if the sun is in the zenith (0°), and equal to SSA90 if the sun is at the horizon (90°) and the azimuth angles of the shoot and sun are normal to each other (Figure 1). Although these extreme conditions rarely occur, various combinations of shoot and sun angles would yield the situations depicted in Figure 1. In experimental stud-ies, only SSA0 is usually measured (e.g., Carter and Smith 1985). Because measures of SSA also included the area of the shoot axis, SSA was somewhat overestimated compared to the silhouette area of needles only.
Ten needles were randomly selected from the central part of a shoot, and mean total (TLAn) and projected (PLAn) needle surface areas for a single needle calculated by means of a geometric model that describes needle shape as a rhomboid (Niinemets and Kull 1995). Total (TLA) and projected (PLA) needle surface areas for a shoot were calculated from the estimations of TLAn, PLAn and needle number per shoot (N). The ratios of silhouette area to both total needle surface area (STAR0) and projected surface area (SPAR0) were calculated as the ratios of SSA0 to TLA and PLA, respectively. Needle packing (NP) was calculated as the ratio of N to shoot length (Ls, length of shoot axis) (Carter and Smith 1985, Boyce 1993). Packing of TLA (TLAP), PLA (PLAP) and SSA (SSAP) was defined as TLA, PLA and SSA0, respectively, per unit shoot length. Shoots were oven-dried at 85 °C for 48 h to determine dry weights of needles (Mn) and shoot axis (Ma). Needle weight per TLA (LWA) and per SSA0 (SWA), and packing of needle dry weight (Mn/Ls) were calculated.
Because a balance exists between needle demand for water and the capacity of conductive tissues to provide water, al-lometric relationships between foliar and sapwood area (Kauf-mann and Troendle 1981), foliar biomass and sapwood area (Grier and Waring 1974, Kaufmann and Troendle 1981) or between foliar area and sapwood volume (Ryan 1989) are often found. Thus the ability of the shoot axis to supply the needles with water must scale with the amount of foliage on the shoot, and hence Ma may serve as an estimate of biomass
costs for needle support (NSC). Moreover, the shoot axis also provides needles with mechanical support, thus adding another dimension to the term NSC. By using Ma as an estimate of NSC, we assume that the costs for construction and mainte-nance (e.g., carbon costs for respiration) of the shoot axis (Chazdon 1986) and for structures of the shoot axis that are not directly related to water conduction and mechanical strength (bark, pith, buds) are proportional to the weight of the shoot axis. For comparative purposes, relative measures of NSC were found: NSC per unit foliage weight (NSCm) was defined as Ma/Mn, and NSC per unit foliage area (NSCa) as Ma/TLA (NSCTLA ), Ma/PLA (NSCPLA ) and Ma/SSA0 (NSCSSA0 ).
Linear regression and correlation techniques were used, and all relationships were considered significant at P < 0.05. Defi-nitions and mean values (± SE) of all needle and shoot parame-ters are listed in Table 1.
Results
Packing of needles, needle surface and weight
Needle number per unit shoot length (NP) and total needle area packing (TLAP) increased significantly with increasing total tree height (TTH) (r2 = 0.362, P < 0.01 and r2 = 0.322, P < 0.02, respectively) and diffusive site factor (ad) (r2 = 0.708, P < 0.001 and r2 = 0.497, P < 0.002, respectively) (Figure 2A and 2B). Because TLAP = NP × TLAn, and mean TLA per needle (TLAn) was not correlated with either ad or TTH (P < 0.05, Niinemets and Kull 1995), the dependence of TLAP on ad and TTH is mainly attributable to variation in NP.
Packing of PLA (PLAP) increased with TTH (r2 = 0.293, P < 0.05) and ad (r2 = 0.238, P < 0.05). Inasmuch as PLAP is equal to:
PLAP = TLAP TLA/PLA ,
(2)
and the ratio TLA/PLA increased linearly with ad (Niinemets and Kull 1995), there are two opposing tendencies with in-creasing ad: (1) decreasing PLA at constant TLA, and (2) increasing needle packing (Figure 2A) and consequently TLAP (Figure 2B). As a result of (1) and (2), there was only a poor correlation between PLAP and ad.
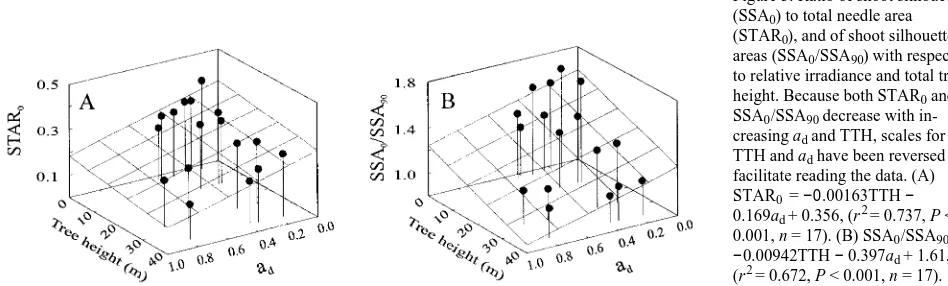
Packing of SSA (SSAP) was not significantly correlated with either ad or TTH (P > 0.05); therefore, SSA0/TLA (STAR0), which can be expressed as:
STAR0=
SSAP
TLAP , (3)
decreased significantly with increasing TTH (r2 = 0.393, P < 0.01) and irradiance (r2 = 0.586, P < 0.001, Figure 3A), as did SPAR0 (r2 = 0.430, P < 0.01 and r2 = 0.366, P < 0.01 for TTH and ad, respectively). The ratio SSA0/SSA90, characterizing the Table. 1. Definitions and means (n = 86) for all studied shoot parame-ters.
Abbreviation Definition Mean ± SE
Ls Length of shoot axis 5.85 ± 0.37 cm LWA Needle dry weight per TLA 62.4 ± 2.5 g m−2
Ma Weight of shoot axis 0.072 ± 0.011 g
Mn Needle dry weight per shoot 0.226 ± 0.021 g
N Needle number per shoot 90.6 ± 6.1 NP Needle packing (N/Ls) 15.40 ± 0.44 cm−1 NSC Needle-supporting costs
NSCa Area-based NSC
NSCm Weight-based NSC (Ma/Mn) 0.269 ± 0.011 g g−1 NSCTLA TLA-based NSCa (Ma/TLA) 16.7 ± 0.9 g m−2 NSCPLA PLA-based NSCa (Ma/PLA) 48.8 ± 3.1 g m−2 NSCSSA0 SSA0-based NSCa (Ma/SSA0) 66.2 ± 4.2 g m−2 PLA PLAn× N 12.17 ± 0.96 cm2 PLAn Projected needle area per needle 12.95 ± 0.39 mm2 PLAP PLA packing (PLA/Ls) 16.6 ± 0.8 cm2 cm−1 SSA Shoot silhouette area
SSA0 SSA for sun in zenith 9.14 ± 0.74 cm2 SSA90 SSA for sun at horizon 7.21 ± 0.54 cm2 SSAP SSA0 packing (SSA0/Ls) 23.1 ± 1.5 cm2 cm−1 SSA0/SSA90 Ratio of SSA0 to SSA90 1.29 ± 0.03 cm2 cm−2 SPAR0 SSA0 to PLA ratio 0.76 ± 0.02 cm2 cm−2 STAR0 SSA0 to TLA ratio 0.27 ± 0.01 cm2 cm−2 SWA Needle weight per SSA0 247 ± 12 g m−2 TLA TLAn× N 35.2 ± 3.1 cm2 TLAn Total needle area per needle 36.5 ± 1.1 mm2 TLA/PLA Ratio of TLA to PLA 2.84 ± 0.04 cm2 cm−2 TLAP TLA packing (TLA/Ls) 5.65 ± 0.23 cm2 cm−1
radial symmetry of a shoot, declined with increasing TTH (r2 = 0.548, P < 0.001) and ad (r2 = 0.342, P < 0.02, Figure 3B), signifying that both the fraction of needles lying outside the SSA0 plane and the symmetry of the shoot about its axis increase with increasing ad and TTH. Both STAR0 (Figure 4) and SPAR0 (data not shown) were dependent on changes in needle morphology (TLA/PLA, Figure 4A) and shoot struc-ture (SSA0/SSA90 and NP, Figure 4B and 4C).
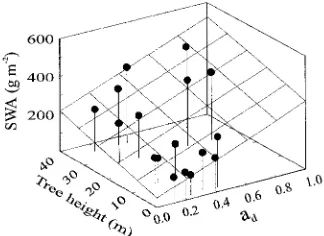
Needle weight per SSA0 (SWA) increased with increasing TTH (r2 = 0.416, P < 0.01) and ad (r2 = 0.674, P < 0.001, Figure 5). Because SWA is equal to:
SWA = LWA STAR0
, (4)
(where LWA is needle dry weight per TLA), changes in both needle morphology, i.e., increasing LWA with ad and TTH (Niinemets and Kull 1995), and shoot structure (STAR0, Fig-ure 3A) are responsible for the variation in SWA. Similarly, an increase in packing of needle dry weight (Mn/Ls) with increas-ing TTH (r2 = 0.501, P < 0.002) and ad (r2 = 0.559, P < 0.001) can be ascribed to both increasing needle packing and increas-ing LWA (Mn/Ls = LWA × NP × TLAn).
Needle support
Needle-support costs per foliage area (NSCa, equal to NSCTLA , NSCPLA and NSCSSA0 ) increased with increasing ad (P < 0.01, Figure 6A), whereas NSCa was not significantly correlated
with TTH. Similarly, Ma/Ls increased with increasing ad (r2 = 0.497, P < 0.01) but was independent of TTH. Because NSC per unit foliage weight (NSCm) equals:
NSCm=
NSCTLA LWA ,
(5)
and LWA increases with ad and TTH (Niinemets and Kull 1995), NSCm should be inversely related to TTH. Indeed, NSCm decreased with increasing TTH (r2 = 0.347, P < 0.02, Figure 6B) but was not related to ad. Furthermore, NSCm (r2 = 0.599, P < 0.001) and NSCa (P < 0.001, r2 = 0.197, 0.172 and 0.131 for NSCTLA , NSCPLA and NSCSSA0 , respectively) in-creased with increasing Ls (n = 86), but Ls was not related to TTH, ad or to most indices of shoot structure (P > 0.05), except for weak correlations with TLAP (r = 0.293, P < 0.01) and PLAP (r = 0.266, P < 0.02).
Discussion
Packing of needles, needle surface and weight
Needle packing (NP) increases similarly (Figure 2A) with increasing irradiance in Abies lasiocarpa (Hook.) Nutt., Picea engelmannii Parry ex Engelm., Pinus contorta Dougl. ex Loud. (Carter and Smith 1985, Smith and Carter 1988), Picea sitchensis (Bong.) Carr. (Leverenz and Jarvis 1980b), Pinus taeda L. (recalculation of data of Cregg et al. 1993) and
Figure 3. Ratio of shoot silhouette (SSA0) to total needle area (STAR0), and of shoot silhouette areas (SSA0/SSA90) with respect to relative irradiance and total tree height. Because both STAR0 and SSA0/SSA90 decrease with in-creasing ad and TTH, scales for TTH and ad have been reversed to facilitate reading the data. (A) STAR0 = −0.00163TTH − 0.169ad + 0.356, (r2 = 0.737, P < 0.001, n = 17). (B) SSA0/SSA90 = −0.00942TTH − 0.397ad + 1.61, (r2 = 0.672, P < 0.001, n = 17).
Figure 4. Dependencies of STAR0 on needle morphology
(TLA/PLA) and shoot structure (NP, SSA0/SSA90). (A) STAR0 versus TLA/PLA; STAR0 = 0.690 − 0.152TLA/PLA (r2 = 0.546, P < 0.001, n = 17). (B) STAR0 versus NP; STAR0 = 0.486 − 0.0141NP (r2 = 0.777, P < 0.001, n = 17). (C) STAR0 versus SSA0/SSA90; STAR0 = 0.024 +
Pseudotsuga menziesii (Mirb.) Franco (Drew and Ferrell 1977), and with increasing canopy height in Abies balsamea (L.) Mill. and Picea rubens Sarg. (Boyce 1993). However, in Pinus sylvestris L., NP decreased with increasing canopy height (Kellomäki and Oker-Blom 1983). High NP in tall trees might be associated with increased water limitations in the crowns of large trees, resulting in reduced water potentials and retarded extension growth of shoots. For example, decreasing values of NP with increasing Ls in Abies fraseri (Pursh) Poir. (Brewer et al. 1992) and Tsuga canadensis (L.) Carr. (Powell 1992) were attributable to a decrease in overall plant vigor, resulting in declining rates of shoot extension growth. How-ever, in P. taeda (recalculation of data of Cregg et al. 1993) NP increased with increasing Ls, and in our study, the decrease in
Ls with increasing TTH was not significant and there was no relationship between Ls and NP, suggesting that other mecha-nisms may govern TTH-related variation in NP.
The amount of TLA that can be accommodated per unit Ls (TLAP) is influenced by relative irradiance and TTH because NP increases with increasing TTH and ad (Figure 2A and 2B). However, as the complexity of foliage organization (TLA < PLA < SSA0) increases, the effect of NP on surface area packing gradually decreases (Equations 2 and 3), and the shading effect of a unit shoot length (SSAP) becomes inde-pendent of ad and TTH. Also, increased needle packing is counterbalanced by decreasing exposure of needle surface area at a constant total needle area (TLA/PLA) (Niinemets and Kull 1995, Figure 4A) and increasing needle overlapping, needle deviation from the SSA0 plane (Figure 3B and 4C), and angle
of needle inclination (Greis and Kellomäki 1981). Thus costs for interception of direct radiation by a unit SSA0 are higher in the upper crown and in taller trees than in the lower crown and in smaller trees as illustrated by constant values of SSAP with increasing NP, or decreasing values of STAR0 and SPAR0 (the construction efficiencies of silhouette area in units of TLA and PLA, respectively) with increasing ad and TTH (Figure 3A), and NP (Figure 4B). Similar patterns have also been found for other conifers. For example, STAR0 decreased with increasing canopy height in P. rubens and A. balsamea (Boyce 1993), and it was less in sun than in shade in P. engelmanni and A. lasio-carpa (Carter and Smith 1985). Tucker et al. (1987) reported that SPAR0 decreased with increasing irradiance in Abies
amabilis Dougl. ex Forbes. However, because shoot asymme-try increases with decreasing ad and TTH, e.g., SSA0/SSA90 (Figure 3B), the efficacy of various shoots depends on solar elevation. Thus sun shoots and shoots of tall trees exhibiting low efficiency in intercepting direct radiation at high solar elevations, as indicated by low values of STAR0 and SPAR0, may be more efficient at low solar elevations than shade shoots and shoots of small trees. For broad-leaved species, canopy photosynthesis is maximized when LWA follows the light gradient (Gutschick and Wiegel 1988). Because needles at various positions within a shoot have about the same photosyn-thetic capacity (Leverenz and Jarvis 1980a, Carter and Smith 1985), it can be advantageous to invest more photosynthetic weight and area where ad is high, which would lead to a shoot architecture characterized by high values of NP and SWA and low values of STAR0 and SPAR0 (Figures 2--5).
In several species, it has been found that angles of leaf inclination increase with increasing light availability (McMil-len and McC(McMil-lendon 1979), allowing light to penetrate deeper foliage layers. The extinction coefficient (Equation 1), which determines the light absorption efficiency of foliage area, de-pends on foliage grouping and inclination angle (Anderson 1966), and values of STAR0 (if LAI is for TLA) and SPAR0 (if LAI is for PLA) are close to the extinction coefficient if sun is in the zenith (Oker-Blom 1986, Oker-Blom and Smolander 1988). Accordingly, increasing STAR0 and SPAR0 with de-creasing ad may result in a situation where the extinction coefficient is dependent on irradiance, and accordingly the transmitted irradiance at each canopy height (Equation 1) is influenced by both cumulative foliar area and the adaptation of foliar architecture to incident irradiance. Though a mean
ex-Figure 6. Dependence of needle support costs (NSC) on relative irradiance and to-tal tree height. (A) NSC per unit foliage area (NSCa, g m−2) versus ad; s = NSC per unit TLA, NSCTLA = 18.7ad + 9.26, (r2 = 0.370, P < 0.01, n = 17); d = NSC per unit PLA, NSCPLA = 71.0ad + 19.9 (r2 = 0.496, P < 0.01, n =17); n = NSC per
unit SSA0, NSCSSA0 = 127.4ad + 16.5 (r2 = 0.730, P < 0.001, n = 17). (B) NSC per unit foliage weight (NSCm, g g−1) versus TTH; NSCm = 0.351 − 0.00408TTH, (r2 = 0.347, P < 0.02, n = 17).
tinction coefficient for the whole canopy may be derived from the light measurements from above and below the canopy, canopy models can be improved by the use of several extinc-tion coefficients that take account of canopy structure (Baldoc-chi et al. 1984, Koppel and Oja 1984).
Variation in shoot structure also influences canopy energy balance by affecting the convective energy exchange of shoots. Overstory leaves might be expected to favor adaptations con-tributing to convective cooling because they are exposed to high radiation loads (Vogel 1968). However, increased needle packing at higher ad results in higher needle temperatures and less effective convective cooling (Koppel 1982, Smith and Carter 1988, Figure 2A). Furthermore, in Picea pungens Engelm., up to 20% of the variation in needle temperature is explained by differences in needle position with respect to wind (Tibbals et al. 1964). Thus, increasing needle width with increasing ad in P. abies (Niinemets and Kull 1995) is also likely to cause significantly higher needle temperatures for needles grown at high ad at the same radiation loads. Elevated needle temperature may be advantageous if the air temperature is lower than optimum for photosynthesis (Givnish 1987, Smith and Carter 1988, Sprugel 1989, Smith and Brewer 1994). Because light reactions of photosynthesis are not very temperature sensitive, overall shoot photosynthesis only de-pends on temperature when leaves are light-saturated (Sprugel 1989), and inasmuch as needles located deeper in the canopy seldom reach light saturation, high needle temperature would preferentially enhance photosynthetic rates at high irradiances. Therefore, unlike many deciduous species, variation in ever-green shoot structure with ad may favor less efficient convec-tive cooling at high solar radiation loads.
Increased needle packing at high relative irradiances may also serve a protective function. After overstory removal, seri-ous damage to photosynthetic apparatus of understory-grown individuals of A. amabilis was related to elevated irradiance (Tucker et al. 1987). In Macroptilium atropurpureum DC., the damage was related to increased angle of leaf inclination, diminished light interception, and decreased leaf photoinhibi-tion (Ludlow and Björkman 1984). Accordingly, increasing needle overlapping and inclination from the horizontal (i.e., higher cost of unit SSA0 in terms of TLA (Figure 3A)) with increasing ad, or increasing shoot radial symmetry (i.e., lower SSA0/SSA90 (Figure 3B)) with increasing ad are likely to reduce the probability of photoinhibition at high ad, because all of these modifications work in concert to decrease mean irra-diance at the needle surface.
Needle support
In several understory palm species, NSCa increases with in-creasing plant age (Chazdon 1986). However, TTH did not influence NSCa in P. abies, and NSCm decreased with increas-ing TTH (Figure 6B). Costs for needle support at different relative irradiances scale with foliage weight rather than with area, as indicated by the constant values of NSCm with increas-ing ad, and the finding that, with increasing ad, NSCa increased in direct proportion with increasing foliage weight per area (Figure 5, NSCSSA0 = NSCm× SWA).
As in P. abies, NSCm in P. taeda tended to decrease with age of grafted scion (recalculation of data of Greenwood 1984). Increasing NSCm with increasing Ls and the high supporting efficiency (1/NSCm) of large trees (Figure 6B) may indicate that young and small trees with relatively long and sparsely needled shoots (Figure 2A) preferentially allocate resources to extension growth rather than to diameter growth of stem and branches as in large trees. Small trees may also have more plagiotropic shoots than large trees, because a horizontal shoot axis can bear less needle weight per unit twig weight than a vertical shoot axis (Horn 1971, Givnish 1988), resulting in higher NSCm. Furthermore, small trees must sustain higher snow loads during the winter than large trees, because crown weight increases more rapidly with TTH than does crown area, resulting in relatively thicker stems in saplings than in large trees (King 1991).
However, expenditures for mechanical support in terms of biomass are usually much higher than actual requirements (Chazdon 1986, King 1991) and lower NSCm in large trees may be inadequate to meet the needs for water transport, resulting in higher water stress in large than in small trees. Decreasing photosynthetic rates with increasing plant age in Chrysothamnus nauseosus (Pall.) Britt. (Donovan and Ehler-inger 1992), P. contorta and Pinus ponderosa Dougl. (Yoder et al. 1994) were not related to declining biochemical capacities of photosynthetic apparatus but to increasing water limitation in crowns of large trees.
Implications for canopy dynamics
Biomass investment for production of unit leaf area (LWA) increased with tree age in Eucalyptus grandis W. Hill ex Maiden. (Leuning et al. 1991) and P. sitchensis (Steele et al. 1989), and with TTH in P. abies (Niinemets and Kull 1995). Moreover, because of increasing needle packing (Figure 2A) and shoot radial symmetry (Figure 3B) with increasing TTH, biomass requirements for construction of unit shoot area (SWA) increased with increasing TTH in P. abies more rapidly than the costs for the production of a unit TLA (Figure 5, Equation 4). Because the ability of a species to produce unit SSA0 with minimum structural requirements ultimately deter-mines the LAI the species can attain (Leverenz and Hinckley 1990, Leverenz 1992), decreasing values of STAR0 and SPAR0 with increasing TTH (Figure 3A) may result in decreased values of stand LAI. Furthermore, because total biomass pro-duction (Pta , annual biomass production per ground area, g m−2) of a stand decreases after maximum LAI is reached (Linder 1985), and relative allocation of annual production for foliage construction (Pf/Pta, where Pf is annual foliage produc-tion per ground area, g m−2) decreases with TTH (Whittaker and Woodwell 1968, Givnish 1988), it follows that stand LAI should decline with increasing TTH after a threshold TTH has been reached:
LAI =Pta(Pf /Pta) LWA .
(6)
area), not LAI per se, scales with light interception, and there-fore, the effect of TTH on light transmission by the canopy is likely to be greater than indicated by the decrease in stand LAI (Equations 4 and 6, Figures 3--5).
The amount of irradiance transmitted by the overstory is an important ecological factor determining long-term dynamics and productivity of the understory as well as self-regeneration of canopy dominants. Thus there is a trade-off between high stand LAI, resulting in enhanced light interception that is advantageous in shading out possible competitors (Gutschick and Wiegel 1988), and natural regeneration, benefiting from high irradiances in the understory. Dispersal and habitat occu-pation by young trees may also be facilitated by the ability of young trees to form extensive foliar displays that enable effi-cient light capture in the understory with minimum investment in biomass (Figure 5).
Acknowledgments
We thank Kalle Sôber for skillful technical assistance.
References
Anderson, M.C. 1964. Studies of the woodland light climate. I. The photographic computation of light conditions. J. Ecol. 52:27--41. Anderson, M.C. 1966. Stand structure and light penetration. II. A
theoretical analysis. J. Appl. Ecol. 3:41--54.
Baldocchi, D.D., D.R. Matt, B.A. Hutchinson and R.T. McMillen. 1984. Solar radiation within an oak-hickory forest: an evaluation of the extinction coefficients for several radiation components during fully-leafed and leafless periods. Agric. For. Meteorol. 32:307--322. Björkman, O. 1981. Responses to different quantum flux densities. In Physiological Plant Ecology. Vol. I. Eds. O.L. Lange, P.S. Nobel, C.B. Osmond and H. Ziegler. Encyclopedia of Plant Physiology 12A (New Series). Springer-Verlag, Berlin, Heidelberg, New York, pp 57--107.
Boyce, R.L. 1993. A comparison of red spruce and balsam fir shoot structures. Tree Physiol. 12:217--230.
Brewer, J.F., L.E. Hinesley and L.K. Snelling. 1992. Foliage attributes for current-year shoots of Fraser fir. HortScience 27:920--925. Carter, G.A. and W.K. Smith. 1985. Influence of shoot structure on
light interception and photosynthesis in conifers. Plant Physiol. 79:1038--1043.
Chazdon, R.L. 1986. The costs of leaf support in understory palms: economy versus safety. Am. Nat. 127:9--30.
Chen, J.M. and T.A. Black. 1992. Foliage area and architecture of plant canopies from sunfleck size distributions. Agric. For. Me-teorol. 60:249--266.
Cregg, B.M., O. Teskey and P.M. Dougherty. 1993. Effect of shade stress on growth, morphology, and carbon dynamics of loblolly pine branches. Trees 7:208--213.
Donovan, L.A. and J.R. Ehleringer. 1992. Contrasting water-use pat-terns among size and life-history classes of a semi-arid shrub. Funct. Ecol. 6:482--488.
Drew, A.P. and W.K. Ferrell. 1977. Morphological acclimation to light intensity in Douglas-fir seedlings. Can. J. Bot. 55:2033--2042. Frey, T. 1977. IBP research at the Vooremaa forest ecology station. In
Spruce Forest Ecosystem Structure and Ecology. Vol. 1. Introduc-tory Data on the Estonian Vooremaa Project. Ed. T. Frey. Estonian IBP Report 11, Acad. Sci. Estonian S.S.R., Estonian Republican Committee for IBP, Tartu, pp 21--36.
Givnish, T.J. 1987. Comparative studies of leaf form: assessing the relative roles of selective pressures and phylogenetic constraints. New Phytol. 106:131--160.
Givnish, T.J. 1988. Adaptation to sun and shade: a whole-plant per-spective. Aust. J. Plant Physiol. 15:63--92.
Gower, S.T. and J.M. Norman. 1991. Rapid estimation of leaf area index in conifer and broad-leaf plantations. Ecology 72:1896--1900. Greenwood, M.S. 1984. Phase change in loblolly pine: shoot
develop-ment as a function of age. Physiol. Plant. 61:518--522.
Greis, I. and S. Kellomäki. 1981. Crown structure and stem growth of Norway spruce undergrowth under varying shading. Silva Fenn. 40:86--93.
Grier, C.C. and R.H. Waring. 1974. Conifer foliage mass related to sapwood area. For. Sci. 20:205--206.
Gutschick, V.P. and F.W. Wiegel. 1988. Optimizing the canopy photo-synthetic rate by patterns of investment in specific leaf mass. Am. Nat. 132:67--86.
Hellkvist, J., G.P. Richards and P.G. Jarvis. 1974. Vertical gradients of water potential and tissue water relations in Sitka spruce trees measured with the pressure chamber. J. Appl. Ecol. 11:637--667. Horn, H.S. 1971. The adaptive geometry of trees. Princeton University
Press, Princeton, New Jersey, 144 p.
Jarvis, P.G. and J.W. Leverenz. 1983. Productivity of temperate, de-ciduous and evergreen forests. In Physiological Plant Ecology. Vol. VI. Eds. O.L. Lange, P.S. Nobel, C.B. Osmond and H. Ziegler. Encyclopedia of Plant Physiology 12D (New Series). Springer-Ver-lag, Berlin, Heidelberg, New York, pp 233--280.
Kaufmann, M.R. and C.A. Troendle. 1981. The relationship of leaf area and foliage biomass to sapwood conducting area in four subal-pine forest tree species. For. Sci. 27:477--482.
Kellomäki, S. and P. Oker-Blom. 1983. Canopy structure and light climate in a young Scots pine stand. Silva Fenn. 17:1--21. King, D.A. 1991. Tree allometry, leaf size and adult tree size in
old-growth forests of western Oregon. Tree Physiol. 9:369--381. Koppel, A. 1982. Konvektivnaya teploperedatsha u pobegov yeli
yevropeiskoi. (On investigation of convective heat exchange in Norway spruce shoots). Proc. Acad. Sci. Estonian S.S.R. Biology 31:37--44 (In Russian).
Koppel, A. and T. Oja. 1984. Regime of diffuse solar radiation in an individual Norway spruce (Picea abies (L.) Karst.) crown. Photo-synthetica 18:529--535.
Leuning, R., R.N. Cromer and S. Rance. 1991. Spatial distributions of foliar nitrogen and phosphorus in crowns of Eucalyptus grandis. Oecologia 88:504--510.
Leverenz, J.W. 1992. Shade shoot structure and productivity of ever-green conifer stands. Scand. J. For. Res. 7:345--353.
Leverenz, J.W. and T.M. Hinckley. 1990. Shoot structure, leaf area index and productivity of evergreen conifer stands. Tree Physiol. 6:135--149.
Leverenz, J.W. and P.G. Jarvis. 1980a. Photosynthesis in Sitka spruce (Picea sitchensis (Bong.) Carr.). IX. The relative contribution made by needles at various positions on the shoot. J. Appl. Ecol. 17:59--68.
Leverenz, J.W. and P.G. Jarvis. 1980b. Photosynthesis in Sitka spruce (Picea sitchensis (Bong.) Carr.). X. Acclimation to quantum flux density within and between trees. J. Appl. Ecol. 17:697--708. Linder, S. 1985. Potential and actual production in Australian forest
stands. In Research for Forest Management. Eds. J.J. Landsberg and W. Parsons. CSIRO, Melbourne, pp 11--35.
McMillen, G.G. and J.H. McClendon. 1979. Leaf angle: an adaptive feature of sun and shade leaves. Bot. Gaz. 140:437--442.
Monsi, M. and T. Saeki. 1953. Über den Lichtfaktor in den Pflan-zengesellschaften und seine Bedeutung für die Stoffproduktion. Jpn. J. Bot. 14:22--52.
Niinemets, Ü. and O. Kull. 1995. Effects of light availability and tree size on the architecture of assimilative surface in the canopy of Picea abies: variation in needle morphology. Tree Physiol. 15:307--315.
Nilson, T. and V. Ross. 1979. Characterization of the transparency of a forest canopy by fish-eye photographs. In Spruce Forest Ecosys-tem Structure and Ecology. Vol. 2. Basic Data on the Estonian Vooremaa Project. Ed. T. Frey. Estonian IBP Report 12, Acad. Sci. Estonian S.S.R., Estonian Republican Committee for IBP, Tartu, pp 114--130.
Oker-Blom, P. 1986. Photosynthetic radiation regime and canopy structure in modeled forest stands. Acta For. Fenn. 197:1--44. Oker-Blom, P. and H. Smolander. 1988. The ratio of shoot silhouette
to total needle area in Scots pine. For. Sci. 34:894--906.
Powell, G.R. 1992. Patterns of leaf size and morphology in relation to shoot length in Tsuga canadensis. Trees 7:59--66.
Ryan, M.G. 1989. Sapwood volume for three subalpine conifers: predictive equations and ecological implications. Can. J. For. Res. 19:1397--1401.
Smith, W.K. and C.A. Brewer. 1994. The adaptive importance of shoot and crown architecture in conifer trees. Am. Nat. 143:528--532. Smith, W.K. and G.A. Carter. 1988. Shoot structural effects on needle
temperatures and photosynthesis in conifers. Am. J. Bot. 75:496--500.
Sprugel, D.G. 1989. The relationship of evergreenness, crown archi-tecture, and leaf size. Am. Nat. 133:465--479.
Steele, M.J., M.P. Coutts and M.M. Yeoman. 1989. Developmental changes in Sitka spruce as indices of physiological age. I. Changes in needle morphology. New Phytol. 113:367--375.
Tibbals, E.C., E.K. Carr, D.M. Gates and F. Kreith. 1964. Radiation and convection in conifers. Am. J. Bot. 51:529--538.
Tucker, G.F., T.M. Hinckley, J.W. Leverenz and S-M. Jiang. 1987. Adjustments of foliar morphology in the acclimation of understory Pacific silver fir following clearcutting. For. Ecol. Manage. 21:249--268.
Vogel, S. 1968. ‘Sun leaves’ and ‘shade leaves:’ differences in convec-tive heat dissipation. Ecology 49:1203--1204.
Whittaker, R.H. and G.M. Woodwell. 1968. Dimension and produc-tion relaproduc-tions of trees and shrubs in the Brookhaven Forest, New York. J. Ecol. 56:1--25.