Summary Two hybrid poplar (Populus) clones (the fast-growing clone Beaupré (P. trichocarpa Torr. and Gray ×P. del-toides Bartr. ex Marsh.) and the slow-growing clone Robusta (P. deltoides Bartr. ex Marsh. ×P. nigra L.)) were grown from hardwood cuttings for one or two growing seasons (1993--1994) in either ambient or elevated (= ambient + 350 µmol mol−1) CO2 in open-top chambers at the University of
Antwer-pen. Both clones responded positively to the elevated CO2
treatment with increased stem volume and aboveground biomass production; however, the clones exhibited different response strategies to the elevated CO2 treatment, and the
responses varied with cutting age and duration of exposure. Clone Beaupré responded to the elevated CO2 treatment with
increases in leaf area and leaf area index during both the first and second growing seasons, but little increase in height growth. Clone Robusta exhibited increased height growth, leaf biomass and total leaf nitrogen content in response to elevated CO2, but no increase in leaf area index. The elevated CO2
treatment increased the total number of branches and total branch biomass in both clones during both growing seasons. At the end of the first growing season, woody stem biomass of the fast- and slow-growing clones was increased by 38 and 55%, respectively. At the end of the second growing season, stem volume was increased by 43% in clone Beaupré and by 58% in clone Robusta. The increase in stem volume was a result of the stimulation of both height and diameter growth in the slow-growing clone, whereas only height growth was stimu-lated in the fast-growing clone. In the fall of the first growing season, the average date of bud set in clone Robusta was advanced by 4 days in the elevated CO2 treatment; there were
no other significant effects of the elevated CO2 treatment on
bud set. The elevated CO2 treatment enhanced leaf C/N ratios
in both clones in both years.
Keywords: branches, leaf area index, nitrogen content, poplar, volume index.
Introduction
The growth of deciduous trees often increases by about 30% in response to elevated concentrations of atmospheric CO2,
whereas the overall mean effect of elevated CO2 on total
aboveground biomass production (or total gross photosynthe-sis) ranges from 0.5 to 15% (Strain 1987, Norby 1989, Eamus and Jarvis 1989, Mousseau and Saugier 1992, Ceulemans and Mousseau 1994). However, among different tree genera as well as among different species and genotypes within the same genus, responses to elevated CO2 vary greatly (Norby et al.
1986, O’Neill et al. 1987, Radoglou and Jarvis 1990, Rogers et al. 1994, Ceulemans and Mousseau 1994). For example, El Kohen et al. (1993) observed that elevated CO2 had no effect
on leaf area of sweet chestnut, whereas it increased total leaf area of beech plants. Duff et al. (1994) found significant differences in the growth rates of two closely related eucalypt species of northern Australia in response to CO2 enrichment.
Because tree responses to increasing atmospheric CO2
con-centrations appear to be species- and genotype-specific, there is an urgent need to obtain more detailed information on the likely consequences of the predicted global CO2 change on
growth, development and productivity of forest trees. Accord-ingly, the ECOCRAFT research network, consisting of more than 12 European university laboratories and research insti-tutes, was established in 1990 with the overall objective of examining the impacts of increasing atmospheric CO2
concen-tration and temperature on European forests. Within the frame-work of this netframe-work, we have studied the effects of elevated atmospheric CO2 concentrations on the growth, physiology
and development of two genotypes of poplar (Populus). Poplar species and hybrids were used because they are planted as high productivity, short-rotation tree crops (e.g., Ceulemans et al. 1987, 1993) and generally show a large positive response to short-term CO2 enrichment (Gaudillère and Mousseau 1989,
Radoglou and Jarvis 1990, Brown 1991) under controlled environment conditions.
To examine the long-term effects of an increase in atmos-pheric CO2 concentration on two Populus clones, Beaupré
First- and second-year aboveground growth and productivity of two
Populus
hybrids grown at ambient and elevated CO
2
R. CEULEMANS,
1B. Y. SHAO,
1X. N. JIANG
2and J. KALINA
31 Department of Biology, University of Antwerpen (UIA), Universiteitsplein 1, B-2610 Wilrijk, Belgium
2 Experimental Center of Forest Biology, Beijing Forestry University, 100083 Beijing, Peoples Republic of China
3 Academy of Sciences of the Czech Republic, Institute of Landscape Ecology, Department of Ecological Physiology of Forest Trees, CS-60300 Brno,
Czech Republic
Received March 2, 1995
(P. trichocarpa Torr. and Gray ×P. deltoides Bartr. ex Marsh.) and Robusta (P. deltoides Bartr. ex Marsh. × P. nigra L.), differing in growth performance, we exposed clonal cuttings to elevated atmospheric CO2 concentrations for two growing
seasons in open-top chambers. Clone Beaupré is a fast-grow-ing clone (Ceulemans et al. 1987, Radoglou and Jarvis 1990, Barigah et al. 1994) and clone Robusta is a slow-growing clone (Radoglou and Jarvis 1990, Barigah et al. 1994). The clones exhibited different responses to elevated CO2 treatment, which
varied with cutting age and duration of exposure.
Materials and methods
Plant materials and planting pattern
Two hybrid poplar clones, the fast-growing interamerican clone Beaupré and the slow-growing euramerican clone Robusta, were raised from clonal hardwood cuttings (25 cm long) in four open-top chambers (OTCs) at the University of Antwerpen, Belgium. On April 22, 1993, the cuttings, 15 per OTC (i.e., 7 to 8 plants of each clone), were planted in a circle at a spacing of 0.6 m. The distance between adjacent OTCs ranged from 8 to 10 m, and to reduce the boundary effect, each OTC was surrounded by 19 plants. Before planting, the heavy clay soil was excavated from each OTC to a depth of 50 cm and replaced with fertile, black horticultural soil. Each OTC was manually weeded, and all plants were automatically drip irri-gated throughout the two growing seasons. Because nutrient conditions were near optimal at the start of the experiment, fertilizer was only applied once during the first growing season and three times during the second growing season.
Open-top chamber design
Each decagonal OTC (3 m in diameter and 6 m in height) was made of 1-m wide Plexiglas sheets and had a usable ground area of 7 m2. Incoming chamber air was supplied by large
ventilators (S and P, CBM 320-65, Spain) at a rate of about 6500 m3 h−1, i.e., about two air volume changes per minute. Air distribution within the OTC was effected through a flexible spiroid pipe attached to a perforated polythene annulus 1 m above the ground. Two OTCs provided a control treatment at ambient atmospheric CO2 (about 350 µmol mol−1), and the
other two OTCs provided an elevated CO2 treatment at ambient
+ 350 µmol mol−1 (about 700 µmol mol−1). Input of CO2 was
controlled by a gas mass flowmeter (Model 5851E, Brooks Instruments, Rosemount, The Netherlands) and was supplied 24 h per day from April 1993 until early November 1993 and from mid-March 1994 until early November 1994. No CO2
enrichment was supplied from November 1993 to March 17, 1994.
Nondestructive height and leaf area measurements
Total plant height was monitored throughout each growing season until November. Leaf area index (LAI) was determined nondestructively for each clone and each treatment. In the first year, individual leaf area was estimated by means of allometric relations between leaf dimensions and leaf area (Ceulemans et
al. 1993), and then the individual leaf areas were summed to obtain the total leaf area per plant. In the second year, only dimensions of individual leaves on the current terminal leader were measured, and branch dimensions and allometric rela-tionships of branch diameter versus leaf area (Ceulemans et al. 1993) were used to determine total leaf area per plant. Leaf area indices of each clone and each treatment were obtained by dividing the total leaf area of 15 plants (first year) or eight plants (second year) by the ground area (7 m2) of each OTC, assuming no interclonal competition within each OTC.
Phenological observations
From September onward, observations on bud set and leaf fall were made per clone and per treatment. Dates of bud set were determined for terminal shoots as well as for branches (second year). For all phenological observations, mean values (± SE) of all plants (15 and eight plants per clone and per treatment in the first and second year, respectively) were calculated.
Stem volume determinations
At the end of the first growing season, stem diameters at the top and bottom of each plant were measured (in two perpen-dicular directions) with a caliper. Stem volume index was estimated for each plant as (π/3)H(R12 + R1R2 + R22), where R1
and R2 are the radii at the bottom and the top of the stem,
respectively, and H is total plant height. At the end of the second year, stem diameters of all harvested plants were meas-ured at 1-m intervals. For each 1-m stem segment, the volume was calculated based on the formula for a truncated cone as above, but where R1 and R2 are the radii at the base and the top
of each stem segment, respectively, and H is the length of the segment. Total stem volume per plant was obtained by sum-ming the volumes of all individual stem segments (Causton 1985).
Destructive biomass measurements
At the end of November 1993, half of the plants (i.e., three to four plants from each clone per OTC) were harvested for analysis of aboveground biomass. The remaining plants in each OTC were measured during the second growing season and harvested on December 8--12, 1994. For both 1993 and 1994 harvests, leaves, branches and stems were oven dried at 80 °C for 3--7 days and weighed. All fallen leaves per OTC were collected, dried and used for determination of total leaf biomass. Because biomass data for 2-year-old stems are not yet available, average biomass per clone per treatment was esti-mated at the end of the first year using the average stem volume index (calculated from height and diameter measurements) of all plants. Total leaf biomass per clone and per treatment at the end of the second growing season was estimated from total stem volumes. Total number of branches per plant was deter-mined at the times of the 1993 and 1994 harvests.
Leaf nitrogen analysis and C/N ratio
technique. Five replicative measurements were made per sam-ple.
The carbon to nitrogen ratio (C/N) was determined for leaves of both treatments from the corresponding data on biomass and nitrogen content assuming that 1 g dry weight = 0.45 g C (Larcher 1980).
Statistical treatment of data
A one-way or two-way analysis of variance (ANOVA) was used to test significance of CO2 treatment and clone, as well as
interactions between these factors. By including chamber within CO2 treatment as the experimental unit, between
cham-ber variation could be accounted for in the analysis. Tukey’s multiple range test or confidence interval test at P = 0.05 or 0.01 followed each ANOVA. A two-sample Student’s t-test was performed on phenological observations (at P = 0.05).
Results
Seasonal evolution of height and leaf area index
Treatment differences in average plant height were small in the middle of the first season, but by the end of the first growing season, heights of clones Beaupré and Robusta were 14 and 17% greater, respectively, in the elevated CO2 treatment than
in the ambient CO2 treatment (P < 0.05) (Figure 1, top). At the
end of the first growing season, clone Beaupré was signifi-cantly taller (P < 0.01) than clone Robusta in both CO2
treat-ments. By the end of the second growing season, however, the only significant clonal difference in plant height was in the ambient CO2 treatment. At this time, clones Beaupré and
Robusta were 4.85 and 4.30 m in height, respectively, in the ambient CO2 treatment (Figure 1, bottom). Although there
were no clonal differences in plant height in the elevated CO2
treatment at the end of the second growing season, Beaupré and Robusta plants were 7 and 18% taller, respectively, in the elevated CO2 treatment than in the ambient CO2 treatment (P
< 0.05). For clone Beaupré, the relative stimulation of plant height growth by elevated CO2 was less in the second year than
in the first year, whereas it was nearly 17% in both years for clone Robusta. For both clones and both treatments, plant height more than doubled in the second year compared with the first year. The difference in average height values at the onset of the second year compared with the end of the first year (Figure 1) was caused by the harvest of half of the plants in each OTC at the end of the first year.
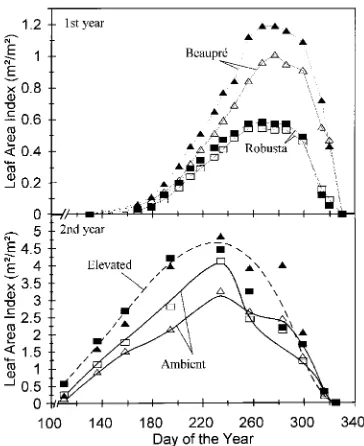
Leaf area index development generally showed the same trend for both clones during both years, except for the later start in the establishment year (Figure 2). In the first year, the LAI of clone Beaupré was more than twice that of clone Robusta in both CO2 treatments (Figure 2, top). The elevated
CO2 treatment significantly increased the LAI of clone
Beau-pré (P < 0.05) in both years. The larger CO2-induced increase
in LAI of clone Beaupré in the second year than in the first year was the result of a low LAI in the ambient treatment in the second year (Figure 2, bottom). The elevated CO2 treatment
had no significant effect on the LAI of clone Robusta at the end of the first or second year (P < 0.05) (Figure 2, bottom).
Figure 1. Time course of average plant height with time during the first (top panel) and second (bottom panel) growing season of poplar clones Beaupré (triangles) and Robusta (squares) grown in open-top cham-bers in ambient or elevated atmospheric CO2. Solid lines and open symbols represent the ambient treatment; dotted lines and filled sym-bols represent the elevated treatment. Fine vertical lines represent the standard error of the mean (15 replications in the first year versus eight replications in the second year).
Figure 2. Time course of leaf area index with time during the first (top panel) and second (bottom panel) growing season of poplar clones Beaupré (triangles) and Robusta (squares) grown in open-top cham-bers in ambient or elevated atmospheric CO2. Solid lines and open
By the end of the second growing season, we had collected over 55% more total leaf biomass in the OTCs providing the elevated CO2 treatment than in the OTCs providing the
ambi-ent CO2 treatment. We calculated that the elevated CO2
treat-ment resulted in increases in total leaf biomass of 36 and 49% for clones Beaupré and Robusta, respectively (Table 1). Thus, although the LAI of clone Robusta was not significantly in-creased in the elevated CO2 treatment, its total leaf biomass
increased by nearly 50%, suggesting a large decrease in spe-cific leaf area.
Bud phenology
The elevated CO2 treatment had no significant effects on
aver-age bud set dates, except at the end of the first year when bud set in clone Robusta was significantly (P < 0.01) advanced by 4 days in the elevated CO2 treatment (Table 2). Although there
was no significant treatment effect on terminal bud set date in clone Beaupré (Table 2), we observed considerable variation among individual plants. For example, at the end of the first year, the first Beaupré and Robusta plants in the elevated CO2
treatment set bud 20 and 9 days earlier, respectively, than in the ambient CO2 treatment. Lateral branches set bud more than
1 month before the terminal shoot, and bud set of lateral branches occurred 10 days earlier in clone Beaupré than in clone Robusta (Table 2). Although a difference in bud set dates of nearly 20 days was observed between the two clones at the end of the first growing season (September 10 for clone Beau-pré and October 1 for clone Robusta), no significant (P < 0.05) clonal differences were observed at the end of the second growing season.
Number and biomass of branches
Because of the small number of sylleptic side branches pro-duced during the first year and the error involved in counting the small branches of clone Robusta, we likely overestimated the effect of elevated CO2 on small branch production
(Ta-ble 3). After two growing seasons, the elevated CO2 treatment
caused a significant (P < 0.01) increase (21--26%) in total number of branches produced by each clone. The elevated CO2
treatment enhanced the production of major proleptic branches more than the production of smaller, sylleptic side shoots (data not shown). The production of these major proleptic branches, which only appear after the first growing season (Ceulemans et al. 1990), accounts for the increase in total branch biomass
Table 1. Nitrogen concentration (% dry weight), total biomass (gDW plant−1), total nitrogen content (g plant−1) and C/N ratio of leaves of poplar
clones Beaupré and Robusta grown in open-top chambers in ambient or elevated CO2. Leaf nitrogen concentration data from two different sampling
dates in the second year (i.e., August and September 1994) are shown, as well as overall mean values for the first and second year. Data on total leaf biomass and leaf nitrogen content are for the end of the second year and are expressed per plant. Values within parenthesis represent standard error of the mean.
Beaupré Robusta
Ambient Elevated Ambient Elevated
Leaf nitrogen concentration (% dry weight)
Overall mean, first year 1.53 (0.12) 1.19 (0.12) 1.79 (0.06) 1.23 (0.17) August, second year 1.56 (0.21) 1.07 (0.07) 1.62 (0.21) 1.25 (0.07) September, second year 2.12 (0.11) 1.30 (0.05) 2.11 (0.05) 1.81 (0.05) Overall mean, second year 1.84 (0.23) 1.19 (0.06) 1.86 (0.16) 1.53 (0.14)
Leaf biomass and leaf nitrogen content
Total leaf DW per plant (g) 235 (84) 319 (39) 169 (60) 250 (41) Total leaf N content per plant (g) 4.32 (1.64) 3.79 (0.50) 3.14 (1.15) 3.83 (0.72) C/N ratio of leaves 24.5 (9.9) 37.8 (5.4) 24.2 (9.7) 29.4 (5.9)
Table 2. Average bud set dates for poplar clones Beaupré and Robusta grown in open-top chambers in ambient or elevated CO2. Data from the ends
of the first and second growing season are shown; all numbers are days of the year. Mean values of 15 individual plants per clone per treatment in the first year versus eight individual plants during the second year are presented. Values within parenthesis indicate standard error of the mean.
Beaupré Robusta
Ambient Elevated Ambient Elevated
First year
Terminal bud set 274 (4) 274 (4) 253 (1) 249 (2) October 1 October 1 September 10 September 6
Second year
Bud set of lateral branches 250 (2) 249 (4) 260 (4) 260 (4) September 7 September 6 September 17 September 17 Terminal bud set 296 (9) 293 (5) 295 (8) 292 (8)
during the second year in the elevated CO2 treatment
(Fig-ure 3).
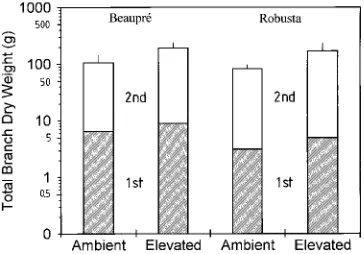
Figure 3 depicts the total branch dry weight per plant pro-duced during the first and second growing seasons in the ambient and elevated CO2 treatments. In both clones and both
years, there was a significant (P < 0.01) increase in branch biomass per plant. The increase in total branch biomass caused by the elevated CO2 treatment was proportionally larger for
clone Robusta (108%), which had fewer branches and smaller total branch biomass, than for clone Beaupré (Table 3, Fig-ure 3). In both clones, the stimulation of branch biomass by elevated CO2 was greater in the second year than in the first
year. Absolute values of total branch biomass ranged from 3 to 9 g after 1 year and amounted to 80 to 190 g after 2 years of growth.
Stem volume and biomass
We used total stem volume production per plant (or per OTC) to estimate overall plant productivity. After 2 years of growth in the presence of elevated CO2, we observed significant
in-creases (P < 0.01) in stem volume per plant of 43 and 58% in clones Beaupré and Robusta, respectively (Figure 4). The increased stem volume production in the elevated CO2
treat-ment was explained not only by stimulated height growth but also by increased stem diameters. We also observed significant (P < 0.01) clonal differences in stem volume production per
plant. During both growing seasons and in both CO2
treat-ments, clone Beaupré had significantly (P < 0.01) greater stem volume production than clone Robusta (2.5 versus 2.0 dm3 per plant).
At the end of the first year, there was a significant clonal difference (P < 0.01) in total biomass production (stem + branches + leaves). Total biomass production of clone Beaupré was 103 g in the ambient CO2 treatment versus 128 g in the
elevated CO2 treatment. The corresponding values for clone
Robusta were 48 and 66 g.
Nitrogen content of leaves
The elevated CO2 treatment significantly decreased leaf
nitro-gen concentration (1.2 versus 1.8% of dry weight) in both clones (Table 1) (cf. Ceulemans et al. 1995). Differences in mean percent nitrogen concentration were not significant (P < 0.05) between growing seasons, except for clone Robusta in the elevated CO2 treatment. During both years, total nitrogen
content of the leaves (i.e., total amount of aboveground nitro-gen in leaf material) was not significantly decreased by the elevated CO2 treatment in clone Beaupré (Table 1). In clone
Robusta, the total leaf nitrogen content decreased in the ele-vated CO2 treatment during the first year, but increased by 22%
in the second year. The foliar C/N ratio significantly increased in both clones and both years (Table 1). The elevated CO2
treatment had little effect on the total amount of nitrogen in the plant (leaves + stems + branches) in the first year (−7% for clone Beaupré and +8% for clone Robusta).
Table 3. Total number of branches (per plant) of poplar clones Beaupré and Robusta during two growing seasons in open-top chambers in ambient or elevated CO2. The number of branches was determined at the end of each growing season and includes small (sylleptic) as well as larger
(proleptic) branches. Mean values of 15 replications (first year) versus eight replications (second year) are presented. Values within parenthesis represent standard error of the mean.
Beaupré Robusta
Ambient Elevated Ambient Elevated
First year 9 19 1 5
Second year 18.3 (1.8) 23.0 (2.6) 19.3 (2.4) 23.3 (2.6)
Figure 3. Total dry weight of branches (per plant) produced during the first (shaded bar) and second (open bars) growing season by poplar clones Beaupré (left) and Robusta (right) grown in ambient or elevated atmospheric CO2. Total branch dry weight is presented on a
logarith-mic scale and expressed per plant. Fine vertical lines represent stand-ard error of the mean (15 replications in the first year versus eight replications in the second year).
Figure 4. Total stem volume (per plant) of poplar clones Beaupré (left) and Robusta (right) at the end of the second growing season in open-top chambers supplied with ambient (open bars) or elevated (shaded bars) CO2. Fine vertical lines represent standard error of the mean
Discussion
The two poplar clones exhibited different responses to elevated atmospheric CO2, and there was an interaction between CO2
response and growing season, as has also been found in sour orange trees (Idso et al. 1991). Thus, the concept of response specificity among tree genera to an increase in atmospheric CO2, as suggested by Mousseau and Enoch (1989) and El
Kohen et al. (1993), might also hold true within genera. Both clones responded positively to elevated CO2 with an
increase in stem volume. The increase was larger in the second year than in the first year indicating that the stimulation by elevated CO2 is affected by plant age or the duration of
expo-sure, or both. The slow-growing clone Robusta was more responsive to the elevated CO2 treatment than the fast-growing
clone Beaupré as indicated by the larger increases in total branch biomass, total leaf biomass and stem volume (Table 4). The increase in stem volume in clone Beaupré was primarily associated with increases in stem diameter, whereas the in-crease in stem volume in clone Robusta was primarily associ-ated with increases in stem height (18%). Stimulassoci-ated height growth in response to elevated CO2 is in agreement with
previous observations on seedlings of several woody plants (Eamus and Jarvis 1989, Radoglou and Jarvis 1990, Brown 1991, Norby et al. 1992, Duff et al. 1994). We have previously shown that, in both clones, the stimulatory effect of elevated CO2 on first-year height growth was caused by an increase in
internode length (i.e., enhanced stem elongation) and not by an increase in the number of internodes produced (Ceulemans et al. 1995).
Although net photosynthesis per unit leaf area was signifi-cantly increased in both clones in the elevated CO2 treatment
(Ceulemans et al. 1995), there was a large clonal difference with respect to the effects of CO2 enrichment on total leaf area.
In response to the elevated CO2 treatment, clone Beaupré
displayed a significantly higher LAI than clone Robusta. In the first and second years, the elevated CO2 treatment increased
LAI of clone Beaupré by 18 and 48%, respectively, whereas the increases in total leaf biomass were only 4 and 36%, respectively, suggesting a large decrease in specific leaf area in the elevated CO2 treatment. The decrease in specific leaf area
was reflected in a decreased total N content of the leaves of clone Beaupré in the elevated CO2 treatment.
Clone Robusta exhibited a small but significant increase (8%) in LAI in response to elevated CO2 that remained
con-stant during both growing seasons. Similar findings have been reported for sweet chestnut (El Kohen et al. 1992, 1993) and sour orange (Idso et al. 1991). The 8% increase in LAI ob-served in the middle of the first growing season in the elevated CO2 treatment was reflected in a similar increase in total leaf
biomass by the end of the growing season (Table 4). However, in the second year, clone Robusta produced much more leaf biomass (reflected in an increase in total leaf N content per plant) without an accompanying increase in leaf area, which led to a significant reduction in overall specific leaf area (cf. Oberbauer et al. 1985, Reekie and Bazzaz 1989). Reductions in specific leaf area are generally ascribed to increased leaf thickness (Mousseau and Enoch 1989) or foliar carbohydrate accumulation (Wong 1990), or both (Jolliffe and Ehret 1985). For both clones, the woody biomass produced per unit of leaf biomass (Table 2) or per unit of leaf area (data not shown) significantly increased. This increase was also reflected in the leaf weight ratio, which decreased by 14 and 20% in clones Beaupré and Robusta, respectively (Ceulemans et al. 1995). Thus, in the elevated CO2 treatment, the slow-growing clone
became relatively more efficient in terms of the amount of biomass produced per unit leaf than the fast-growing clone; however, the fast-growing clone showed a more pronounced decrease in specific leaf area in response to CO2 enrichment
than the slow-growing clone.
The elevated CO2 treatment induced large increases in the
biomass and number of branches (Table 4) (cf. Ceulemans et al. 1990, Ceulemans et al. 1995). Large increases in branch biomass in response to elevated CO2 have been reported for
sour orange trees (Idso et al. 1991). Idso et al. (1991) reported that orange trees produced fourth-, fifth- and sixth-order branches 2.5 years after exposure to elevated atmospheric CO2
conditions, whereas trees in the ambient treatment had no branches above the third order. In contrast, in eucalypts, ele-vated CO2 treatment did not affect branch biomass but caused
large increases in stem and root biomass (Duff et al. 1994). We have previously shown that elevated CO2 delays bud
Table 4. Relative difference (percent) between the elevated and ambient CO2 treatments for a number of aboveground growth and developmental
characteristics of poplar clones Beaupré and Robusta after 1 and 2 years of growth in open-top chambers. All differences are significant (per clone per year), unless followed by ns.
Beaupré Robusta
burst by 2 to 5 days in clone Beaupré, whereas the treatment has no significant effect on bud phenology in clone Robusta (Ceulemans et al. 1995). Although no temperature differences were observed between OTCs with different CO2 treatments,
Beuker (1994) suggested that global warming would result primarily in important differences in leaf phenology and the timing of bud burst, at least in coniferous plant species. At the end of the first year, bud set of clone Robusta was advanced by 4 days in the elevated CO2 treatment. Similar findings have
been reported for sweet chestnut (Mousseau and Enoch 1989) and birch (Evans and Lee 1993). We did not observe any significant treatment differences in the timing of bud set for branches or the current terminal leader. Gunderson et al. (1993) has suggested that elevated CO2 effects on bud
phenol-ogy are dependent on species or interactions of CO2 with
temperature, or both.
We conclude that both clones respond positively to elevated CO2 concentrations. The CO2-induced responses of clone
Beaupré included increased investment in branch and leaf biomass that resulted in a significantly increased leaf area index. The CO2-induced responses of the slow-growing clone
Robusta included an increase in height growth and increased investment in branch biomass and total leaf N content; how-ever, these increases were not reflected in a higher LAI.
Acknowledgments
This research forms part of the European ECOCRAFT network, sup-ported by the EC R and D Environment Program (Contract No. EV5V-CT92-0127) and coordinated by the University of Edinburgh, Scotland. Further support was provided by the Belgian National Fund for Scientific Research (Brussels) and the Special Research Fund of the University of Antwerpen (UIA). We gratefully acknowledge K. Brouwers and N. Calluy for nitrogen analysis, F. Kockelbergh and G. Heiler for technical assistance, and K. Bortier, L. Van Praet and B. Van Staeyen for help with biomass determinations. RC is a Senior Research Associate of the Belgian NFWO.
References
Barigah, T.S., B. Saugier, M. Mousseau, J. Guittet and R. Ceulemans. 1994. Photosynthesis, leaf area and productivity of 5 poplar clones during their establishment year. Ann. Sci. For. 51:613--625. Beuker, E. 1994. Adaptation to climatic changes of the timing of bud
burst in populations of Pinus sylvestris L. and Picea abies (L.) Karst. Tree Physiol. 14:961--970.
Brown, K.R. 1991. Carbon dioxide enrichment accelerates the decline in nutrient status and relative growth rate of Populus tremuloides
Michx. seedlings. Tree Physiol. 8:161--173.
Causton, D.R. 1985. Biometrical, structural and physiological rela-tionships among tree parts. In Trees as Crop Plants. Eds. M.G.R. Cannell and J.E. Jackson. Institute of Terrestrial Ecology, Hunting-don, U.K., pp 137--159.
Ceulemans, R. and M. Mousseau. 1994. Effects of elevated atmos-pheric CO2 on woody plants. Tansley Review No. 71. New Phytol.
127:425--446.
Ceulemans, R., I. Impens and V. Steenackers. 1987. Variations in photosynthetic, anatomical, and enzymatic leaf traits and correla-tions with growth in recently selected Populus hybrids. Can. J. For. Res. 17:273--283.
Ceulemans, R., X.N. Jiang and B.Y. Shao. 1995. Growth and physiol-ogy of one-year-old poplar (Populus) under elevated atmospheric CO2 levels. Ann. Bot. 75:609--617.
Ceulemans, R., J.Y. Pontailler, F. Mau and J. Guittet. 1993. Leaf allometry in young poplar stands: reliability of leaf area index estimation, site and clone effects. Biomass Bioenergy 4:315--321. Ceulemans, R., R.F. Stettler, T.M. Hinckley, J.G. Isebrands and P.E.
Heilman. 1990. Crown architecture of Populus clones as deter-mined by branch orientation and branch characteristics. Tree Physiol. 7:157--167.
Duff, G.A., C.A. Berryman and D. Eamus. 1994. Growth, biomass allocation and foliar nutrient contents of two Eucalyptus species of the wet-dry tropics of Australia grown under CO2 enrichment.
Funct. Ecol. 8:502--508.
Eamus, D. and P.G. Jarvis. 1989. The direct effects of increase in the global atmospheric CO2 concentration on natural and commercial
temperate trees and forests. Adv. Ecol. Res. 19:1--55.
El Kohen, A., H. Rouhier and M. Mousseau. 1992. Changes in dry weight and nitrogen partitioning induced by elevated CO2 depend
on soil nutrient availability in sweet chestnut (Castanea sativa
Mill.). Ann. Sci. For. 49:83--90.
El Kohen, A., L. Venet and M. Mousseau. 1993. Growth and photosyn-thesis of two deciduous forest species at elevated carbon dioxide. Funct. Ecol. 7:480--486.
Evans, L. and H.S.J. Lee. 1993. The effect of elevated CO2 on the long
term growth of birch. J. Exp. Bot. 44(suppl.):12
Gaudillère, J.P. and M. Mousseau. 1989. Short term effect of CO2
enrichment on leaf development and gas exchange of young poplars (Populus euramericana cv. I-214). Oecol. Plant. 10:95--105. Gunderson, C.A., R.J. Norby and S.D. Wullschleger. 1993. Foliar gas
exchange responses of two deciduous hardwoods during 3 years of growth in elevated CO2: no loss of photosynthetic enhancement. Plant Cell Environ. 16:797--807.
Idso, S.B., B.A. Kimball and S.G. Allen. 1991. CO2 enrichment of sour orange trees: 2.5 years into a long-term experiment. Plant Cell Environ. 14:351--353.
Jolliffe, P.A. and D.L. Ehret. 1985. Growth of bean plants at elevated CO2 concentrations. Can. J. Bot. 63:2021--2025.
Larcher, W. 1980. Physiological plant ecology. 2nd Edn. Springer-Ver-lag, Berlin, 303 p.
Mousseau, M. and Z.H. Enoch. 1989. Carbon dioxide enrichment reduces shoot growth in sweet chestnut seedlings (Castanea sativa
Mill.). Plant Cell Environ. 12:927--934.
Mousseau, M. and B. Saugier. 1992. The direct effect of increased CO2
on gas exchange and growth of forest tree species. J. Exp. Bot. 43:1121--1130.
Norby, R.J. 1989. Direct responses of forest trees to rising atmospheric carbon dioxide. In Air Pollution Effects on Vegetation. Eds. R.D. Noble, J.L. Martin and K.F. Jensen. USDA Forest Service Publica-tion, Minneapolis-St. Paul, MN, pp 243--249.
Norby, R.J., E.G. O’Neill and R.J. Luxmoore. 1986. Effects of atmos-pheric CO2 enrichment on the growth and nutrition of Quercus alba
seedlings in nutrient poor soil. Plant Physiol. 82:83--89.
Norby, R.J., C.A. Gunderson, S.D. Wullschleger, E.G. O’Neill and M.K. McCracken. 1992. Productivity and compensatory response of yellow poplar trees in elevated CO2. Nature 357:322--324.
Oberbauer, S.F., B.R. Strain and N. Fletcher. 1985. Effect of CO2
enrichment on seedling physiology and growth of two tropical species. Physiol. Plant. 65:353--356.
O’Neill, E.G., R.J. Luxmoore and R.J. Norby. 1987. Elevated atmos-pheric CO2 effects on seedling growth, nutrient uptake, and
Radoglou, K.M. and P.G. Jarvis. 1990. Effects of CO2 enrichment on
four poplar clones. I. Growth and leaf anatomy. Ann. Bot. 65:617--626.
Reekie, E.G. and F.A. Bazzaz. 1989. Competition patterns of resource use among seedlings of five tropical trees grown at ambient and elevated CO2. Oecologia 79:212--222.
Rogers, H.H., G.B. Runion and S.V. Krupa. 1994. Plant responses to atmospheric CO2 enrichment with emphasis on roots and the
rhi-zosphere. Environ. Pollut. 83:155--189.
Strain, B.R. 1987. Direct effects of increasing atmospheric CO2 on
plants and ecosystems. Trends Ecol. & Evol. 2:18--21.
Wong, S.C. 1990. Elevated atmospheric partial pressure of CO2 and