www.elsevier.comrlocaterapplanim
Seasonal foraging behavioural compensation in
ž
reproductive wapiti hinds Cer
Õ
us elaphus
/
canadensis
J.V. Gedir
1, R.J. Hudson
)Department of Renewable Resources, 751 General SerÕices Building, UniÕersity of Alberta, Edmonton,
Alberta, Canada, T6G 2E1
Accepted 3 November 1999
Abstract
Captive wapiti hinds were observed during seven periods between June 1996 and November 1997. We investigated their activity and foraging behaviour on two pastures, heavily and lightly
Ž
grazed, during four phases of the reproductive cycle early gestation, late gestation, peak lactation,
.
and late lactation . Within season, differences in forage availability between pastures had little
Ž .
apparent effect on wapiti feeding behaviour p)0.05 . However, within each pasture, hinds tended to select locations of higher phytomass than the pasture average. Among seasons, effects of forage availability on feeding behaviour were pronounced. Hinds grazed longest in late lactation
Ž12.8 hrday , when they spent 94% of their active time foraging, whereas during early gestation.
Ž . Ž .
they foraged fewer hours 8.2 hrday and less intensively 66% of active time . The longest
Ž .
foraging bouts also occurred in late lactation 100 min and decreased linearly as the number of
Ž 2 . Ž . Ž
bouts per day increased R s0.88 . The annual peak bite rate BR was in late lactation 62
. Ž .
bitesrmin , whereas the annual nadir occurred in early gestation 37 bitesrmin . Smallest bite
Ž . Ž . Ž .
sizes BS mg were recorded in late gestation 127 mg , and increased linearly with forage
Ž 2 . Ž .
availability R s0.46 , with largest BS occurring during peak lactation 280 mg . This study demonstrated how seasonal modifications in activity and foraging behaviour enabled gestating and lactating wapiti hinds to satisfy their changing nutritional requirements on seasonal pastures.
)Corresponding author. Renewable Resources, Faculty of Agriculture, Forestry, and Home Economics,
2-10 Agriculture–Forestry Centre, Edmonton, Alberta, Canada T6G 2P5. Tel.: q1-780-492-2980; fax: q1-780-492-0097.
Ž . Ž .
E-mail addresses: [email protected] J.V. Gedir , [email protected] R.J. Hudson .
1
Tel.:q44-1865-281-520; fax:q44-1865-310-447.
0168-1591r00r$ - see front matterq2000 Elsevier Science B.V. All rights reserved.
Ž .
Knowledge of behavioural compensation in response to nutritional demand and pasture conditions will be useful in designing supplementation program for farmed wapiti.q2000 Elsevier Science B.V. All rights reserved.
Keywords: Wapiti; Elk; Grazing; Activity pattern; CerÕus elaphus
1. Introduction
Northern ungulates synchronize annual reproduction with seasonal cycles in range forage characteristics so energy requirements are matched with forage quality and quantity. However, within seasons, forage conditions vary, and herbivores respond proximally to these changes. Behavioural compensation is potentially the most important
Ž
tactic for meeting additional energy demands of reproduction Gittleman and Thompson,
.
1988 .
Daily feeding activity involves several feeding bouts organized at different hierarchal
Ž . Ž .
levels feeding patch, feeding station and bite Gates and Hudson, 1983 . Daily foraging may change with changes in pattern of feeding bouts and dynamic characteristics of
Ž .
each bout. Jiang and Hudson 1993 concluded that wapiti forage in a manner consistent
Ž .
with Charnov’s 1976 Marginal Value Theorem.
Ž .
Intake of grazed herbage can be described as the product of bite size BS , bite rate
ŽBR , and grazing time Spedding et al., 1966 . BS has the greatest influence on. Ž .
Ž .
short-term intake, BR and grazing time are compensatory variables Forbes, 1989 , and
Ž .
all are sensitive to sward conditions Hodgson, 1985 . Although energy balance may ultimately control intake, it is proximally governed by a combination of plant structural factors that influence ingestion rate, gut fill and the dynamics of gut repletionrdepletion, and social behaviour and environmental factors affecting the appetite–satiety complex
ŽForbes, 1989 ..
In this study, we explored the feeding behaviour of farmed wapiti hinds, specifically their response to seasonal variation in forage availability and nutritional requirements. The objectives were to investigate how they alter their activity budgets and foraging
Ž
dynamics on differing planes of nutrition, in order to meet the demands both time and
.
energy of reproduction. The findings are expected to facilitate development of optimal supplementation programs for farmed wapiti.
2. Material and methods
The study was conducted at Ministik Wildlife Research Station, located 48-km southeast of Edmonton, AB on the Cooking Lake glacial moraine. Vegetation is
Ž .
classified as lower boreal mixedwood Strong, 1992 , consisting of an overstory of
Ž . Ž .
balsam poplar Populus balsamifera and trembling aspen P. tremuloides , and
under-Ž . Ž .
storey dominated by beaked hazel Corylus cornuta and willow Salix spp. .
Grass-Ž .
ŽBromus inermis , white clover Trifolium repens , dandelion Taraxacum officinale ,. Ž . Ž .
Ž .
and Canada thistle Cirsium arÕense .
Between June 1996 and November 1997, wapiti hinds were observed over seven periods, each lasting 16 days. Four stages of the annual reproductive cycle were
Ž . Ž .
included: early gestationrautumn early November ; late gestationrspring early May ;
Ž .
peak lactationrearly summer late Junerearly July ; and late lactationrlate summer
Žlate August . Two enclosures with similar plant species composition were established as.
Ž . Ž .
heavily H and lightly L grazed pastures, with areas of 2.6 and 2.2 ha, respectively.
Ž
Commencing in early gestation 1996, smaller enclosures were employed H: 1.2 ha, L:
.
2.0 ha to enable more rapid defoliation in H. Prior to study commencement, H was stocked with at least ten mature wapiti for 2 weeks to provide sufficient defoliation, while L had been free of grazing for at least 2 months. Between experimental periods, H
Ž .
was always stocked except during snowcover periods to ensure continuous defoliation, while L remained empty. Except during late gestation, pasture phytomass between enclosures differed by at least 30%.
Ž . Ž
Five bred wapiti hinds mean weight: 289"18 kg of similar age distribution mean
.
age: 8"2.6 yr were randomly assigned to each pasture. All hinds raised calves which were present during lactation. Hinds were free-ranging and supplemented with
concen-Ž . Ž .
trate alfalfa-barley pellets year-round and hay winter , except during study sessions, when all supplemental feed was withdrawn. To monitor condition, animals were weighed on an electronic stock scale at the beginning and end of each of the seven 16-day trials. No attempt was made to hold animals without feed or water before weighing.
Ž .
Foraging behaviour was observed during morning 0800–1130 h and afternoon
Ž1400–1700 h intensive grazing sessions on several days each study period. Cropping.
Ž .
BRs bitesrmin were tabulated from 4–15-min sessions and corrected for nonforaging activities exceeding 30 s. BR observed during sessions less than 2 min duration were discarded.
Twice during each observational period, 10=20 cm plots paired with each grazing observation were hand-plucked to ground level to emulate maximum possible removal by grazing wapiti. Samples were freeze-dried for 72 h at y608C to determine feeding
Ž 2.
patch phytomass FPP, g DMrm . At the beginning, middle, and end of each 1997 session, five 0.25=0.25 m vegetation plots were sampled from randomly chosen grassland locations in each enclosure. These samples were oven-dried for 5 days at 608C
Ž .
to determine pasture phytomass kg DMrha . Phytomass measurements were collected
Ž
during a concurrent study at Ministik N. Donkor, unpublished data used with
permis-.
sion .
Also paired with grazing observations, mean BS were estimated by hand-plucking 20 to 40 emulated ‘‘bites’’, to duplicate the amount and species composition of bites
Ž .
ingested by wapiti Hudson and Nietfeld, 1985 . Variations in wapiti incisor bar width and observer bias were corrected with the following equation:
Corrected Bite Size BS
Ž
.
sBS=IrOŽ .
1Ž .
where I is wapiti incisor bar width mm and O is width on observer hand that prehends
Ž .
grazing greatly improves representation of BS, and as well, past cervid studies on
Ž
Ministik pastures have demonstrated reliable accuracy for this method Renecker and
.
Hudson, 1985; Jiang and Hudson, 1992 .
Activity budgets were determined using the predominant activity sampling method
ŽHutt and Hutt, 1970 , whereby a behaviour was assigned if it occurred more than half.
of the given interval, irrespective of its distribution within the interval. This sampling method is well-suited for captive free-ranging wapiti, as they are predictable and tend to sustain a behaviour for relatively long periods of time. For each group, behaviours were
Ž
recorded at 10-min intervals over a 24-h period diurnal period only during peakrlate
.
lactation 1996 , once per study period. Activities were categorized as foraging, bedded, standing, and other, representing the only behaviours routinely demonstrated for at least 5 consecutive minutes. Other activities exceeding 5 min were rare, and included
Ž . Ž .
movement running, walking , self-grooming, rubbing against fence post or tree ,
Ž . Ž
cow–calf interaction nursing, grooming , and cow–cow interaction agonistic,
groom-.
ing .
One-way analysis of variance was used to compare activity and foraging parameters,
Ž .
with reproductive season earlyrlate gestation, peakrlate lactation as fixed effects and
Ž
random effects for individual hinds. Bonferroni’s pairwise multiple comparison SPSS,
. Ž .
1998 and Tukey’s HSD test SPSS, 1998 were used to analyze activity and foraging
Table 1
Seasonal foraging parameters and forage availability for wapiti hinds on heavily and lightly grazed pasture
Ž1997 only at Ministik Research Station, AB LG. Ž slate gestation; PLspeak lactation; LLslate lactation;
. a,b,c Ž . Ž .
EGsearly gestation . Row means that do not share superscripts differ p-0.05 ANOVA
Spring Summer Autumn SE
Duration minrbout 67.9 83.3 114.2 54.5 6.0
a a b c
Feeding patch phytomass g DMrm 183 321 314 257 26
U U
ab b ab a
†
Ž .
Pasture phytomass kg DMrha 1625 2669 1680 1449 88
Lightly grazed
Duration minrbout 59.6 50.2 84.7 65.4 3.3
ab b c a
Feeding patch phytomass g DMrm 263 580 498 312 42
U U
a b b a
Ž .
Pasture phytomass kg DMrha † 1645 4875 3799 2152 243
U
Ž . Ž .
Heavily grazed differs from lightly grazed pasture p-0.01 within season t-test .
UU
Ž . Ž .
Heavily grazed differs from lightly grazed pasture p-0.05 within season t-test .
† Ž .
parameters, respectively, to identify group means that differed significantly. Analyses of covariance were performed to detect whether age, sex, or day were covariates, and no significant effects were revealed. If there was no significant difference between
respec-Ž .
tive seasons among years, data were pooled. Student’s t-test SPSS, 1998 was used to compare behaviour between pastures within seasons. Interrelationships among activity and foraging parameters were analyzed with non-linear regression. All statistical
compu-Ž .
tations were performed using SPSS Base 8.0 1998 , and p-0.05 were accepted as significant. Means are reported with standard errors.
3. Results
Ž
From nearly identical forage availability in both pastures in spring i.e.,
commence-.
ment of the growing season , there was an almost threefold increase in L in early
Ž .
summer, compared to approximately 60% in H Table 1 . Despite this seasonal difference, feeding and bedding patterns were similar between H and L in all seasons. There was no significant difference in diel, nocturnal, and diurnal foraging and bedding
Ž .
between pastures p)0.05 . Pasture differences in the organization of foraging bouts
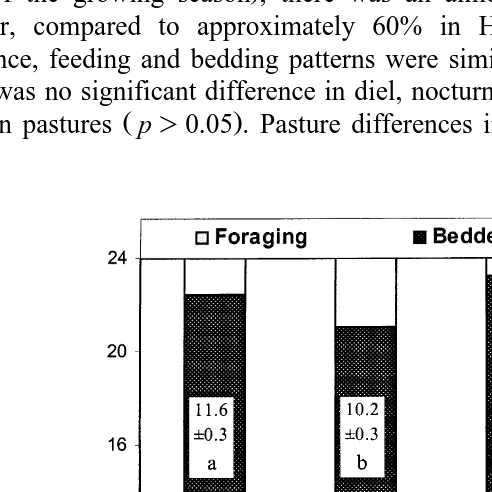
Fig. 1. Seasonal activity budgets of wapiti hinds at Ministik Research Station, AB. Other refers to any
Ž .
behaviours exhibited when not foraging or bedded e.g., standing, social interaction, nursing, movement .
Ž .
Values are mean"SE. Within foraging and bedding, seasons that do not share letters differ p-0.05 .
Ž .
were only significant during summer p-0.05 , when wapiti grazing L had more bouts of shorter duration. Differences in foraging intensity between pastures were only evident
Ž .
from early summer through autumn p-0.05 , with H being higher during summer, and lower in autumn. Finally, there was no significant difference in gains between H and
Ž .
L p)0.05 . Where seasonal differences were not significant, data were pooled. Wapiti showed polyphasic patterns of alternating foraging and bedding throughout the year, with foraging peaks at dawn and dusk. They usually bedded during the night in all seasons, particularly in the hours immediately after sunset and before sunrise. These peaks of activity and inactivity demonstrated seasonal and photoperiodic shifts.
Ž .
Diel grazing time was lowest in autumn 8.23"0.24 h and peaked in late summer
Ž12.77"0.43 h. Žp-0.01. ŽTable 1 . Peak bedding duration in autumn 12.64. Ž "0.25,
Ž . Ž . Ž . Ž .
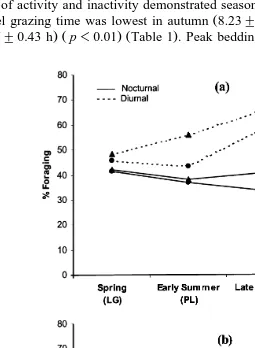
Fig. 2. Seasonal a foraging and b bedding of wapiti hinds on heavily H and lightly L grazed pasture, in diurnal and nocturnal periods, expressed as percentage of total hours of daylight and darkness, respectively.
Table 2
Ž . Ž .
Seasonal percentage of active time spent foraging for wapiti hinds on heavily H and lightly L grazed
Ž
pasture, at Ministik Research Station, AB LGslate gestation; PLspeak lactation; LLslate lactation;
. a,b,c Ž . Ž .
EGsearly gestation . Row means that do not share superscripts differ p-0.01 ANOVA
Spring Summer Autumn SE
ŽLG. Early PLŽ . Late LLŽ . ŽEG.
U UU U
a a b c
H 89.2 87.5 98.8 59.7 2.8
c a c a
L 85.3 70.2 89.2 73.1 1.7
ab a b c
Total 87.2 78.8 94.0 66.4 1.6
U
Ž . Ž .
H differs from L p-0.01 within season t-test .
UU
Ž . Ž .
H differs from L p-0.05 within season t-test .
. Ž .
p-0.001 coincided with the annual foraging nadir Fig. 1 . To accommodate seasonal variation in daylength, we expressed diurnal and nocturnal behaviours as percentage of total hours of daylight and darkness, respectively. Annual peak of nocturnal bedding in
Ž . Ž .
autumn 65.1"7.6% corresponded with nocturnal feeding minimum 27.8"6.5%
ŽFig. 2 . Late summer maximum of diurnal feeding 62.3. Ž "5.7% also paralleled the.
Ž .
annual low of diurnal bedding 35.2"5.0% .
Ž . Ž
Wapiti foraged most of their active time but this varied seasonally p-0.01 Table
. Ž .
2 . Foraging intensity increased in late summer 94.0"5.0% and declined in autumn
Ž66.4"8.2%. Žp-0.001 ..
Ž . Ž .
Foraging bouts were longest in late summer 99.5"20.3 min p-0.01 , followed
Ž . Ž . Ž .
by a sharp decline in autumn 59.9"11.1 min p-0.001 Table 1 . Number of bouts
Fig. 3. Seasonal duration of foraging bouts for wapiti on heavily and lightly grazed pasture in relation to
Ž
number of daily bouts, from spring to late summer at Ministik Research Station, AB ys10.8 xq173.6, 2 .
Ž . Ž .
per day were highest in spring 9.7"1.2 and early summer 10.2"2.2 , and fewest in
Ž . Ž . Ž . Ž .
late summer 7.0"0.8 and autumn 7.3"0.6 p-0.01 Table 1 .
Ž .
Foraging bout duration min increased with the number of foraging bouts from
Ž 2 . Ž .
spring through late summer ys10.8 xq173.6, p-0.01, R s0.88 Fig. 3 .
Forag-Ž .
ing bouts in autumn did not demonstrate this relationship p)0.05 .
Ž .
BRs were highest in late summer 61.9"1.6 bitesrmin and lowest in autumn
Ž37.2"1.5 bitesrmin. Žp-0.001. ŽTable 1 . There was no significant variation.
Ž .
between H and L in any season p)0.05 so data were pooled. BS and feeding patch
Ž .
phytomass showed similar seasonal patterns in both pastures Table 1 . Because
Ž . Ž .
Fig. 4. Seasonal foraging rates bitesrmin of wapiti on heavily and lightly grazed pasture in relation to a
Ž . Ž . Ž 2. ŽŽ .
bite size mg and b feeding patch phytomass g DMrm , at Ministik Research Station, AB a
y0 .0011 x 2 Ž . 2 .
differences were not significant, data from H and L pastures were pooled. Annual
Ž 2.
maxima occurred in early summer BS: 280"28 mg, FPP: 451"68 g DMrm , and
Ž 2. Ž .
minima in spring BS: 127"14 mg, FPP: 223"21 g DMrm p-0.01 .
Ž . Ž . Ž y0 .0011 x
BRs bitesrmin declined exponentially with BS mg ys59.5e , p-0.01,
2 . Ž 2. Ž
R s0.64 and inversely with increasing FPP g DMrm ys25.1q5548rx, p
-2 . Ž .
0.05, R s0.36 Fig. 4 . Regression of BS against FPP revealed a significant positive
Ž . Ž .
association p-0.001 . When regressed against forage availability kg DMrha , strong
Ž . Ž 2 .
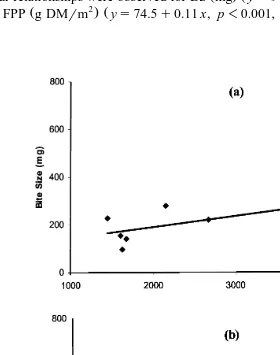
linear relationships were observed for BS mg ys97.7q0.05 x, p-0.05, R s0.46
Ž 2. Ž 2 . Ž .
and FPP g DMrm ys74.5q0.11 x, p-0.001, R s0.91 Fig. 5 .
Ž . Ž . Ž . Ž 2.
Fig. 5. Seasonal a bite size mg and b feeding patch phytomass g DMrm for wapiti on heavily and
Ž . ŽŽ .
lightly grazed pasture, in relation to forage availability kg DMrha , at Ministik Research Station, AB a
2 Ž . 2 .
Ž 0.75.
Hinds gained weight, except during spring y2.9"0.8 grkg . Gains peaked in
0.75 Ž .
early summer at 12.5"1.4 grkg p-0.001 , whereas hinds maintained weight
Ž 0.75. Ž 0.75. Ž .
stasis in late summer 0.3"0.8 grkg and autumn 0.6"1.2 grkg p)0.05 .
4. Discussion
Attempts to maintain marked difference in phytomass between H and L were successful. Unfortunately, between-session time constraints and concern about maintain-ing animal condition resulted in H forage availability bemaintain-ing well above that which would
Ž Ž
normally be regarded as limiting for wapiti e.g., 900 kg DMrha Gates and Hudson,
..
1983 . This may explain the general lack of contrast between pastures. For example,
Ž .
when Heydon et al. 1993 compared lactating red deer hinds on high and low
Ž
phytomass pastures, they found a considerable difference in BR high: 62 bitesrmin;
.
low: 82 bitesrmin . The higher phytomass pasture contained 1659 kg DMrha, while forage availability would likely be considered a limiting factor in the ‘low’ pasture, with only 466 kg DMrha. Further research similar to this study should be undertaken, incorporating pastures with a more limited forage supply, in order to better examine possible inter-pasture effects of seasonal availability during gestation and lactation.
Wild cervids are well known for their crepuscular habit, however, animals on game
Ž
farms or research stations often show feeding cycles throughout the day Gates and
.
Hudson, 1983; Jiang and Hudson, 1996 . The biphasic pattern displayed by Jiang and
Ž .
Hudson’s 1996 non-reproductive hinds in early summer, emphasizes the difference in lactating hinds that have higher energy demands and nurse their calves throughout the
Ž .
day. Gates and Hudson 1983 also observed biphasic patterns in spring and summer,
Ž
however, only three of nine animals were lactating i.e., there were also two stags and
.
four calves , which may account for the disparity.
Daily foraging time in the present study was comparable to those found in other
Ž
investigations Craighead et al., 1973; Collins et al., 1978; Hanley, 1982; Jiang, 1993;
.
Florkiewicz, 1994 . Some studies did not distinguish grazing from total active time, therefore, foraging was estimated by correcting with seasonal values of percent active
Ž .
time foraging found in this study Table 2 . Grazing time of reproductive hinds in the
Ž .
present study was similar to Jiang’s 1993 non-reproductive hinds at the same research station. This implies that greater nutrient requirements of gestating and lactating wapiti were not satisfied by increasing grazing time.
Ž .
Gates and Hudson 1983 suggest that fatigue and demand of alternate activities place an upper limit on daily foraging time, and this is commonly cited to be around 12 h.
Ž . Ž .
This presumption is supported by Clutton-Brock et al. 1982 and Heydon et al. 1992 , whereby, increases in daily grazing time between non-reproductive and reproductive red deer hinds were 9.8 to 11.8 h and 10.8 to 12.2 h, respectively. Perhaps, if lactating hinds had access to pastures with higher forage availability, they could better manipulate their BR and BS to maintain intake within these grazing time constraints.
When not foraging, wapiti usually bedded; however, maternal care disrupted this
Ž .
while hinds were lactating. For example, the reduction in nocturnal bedding during peak lactation was accompanied by a decrease in nighttime grazing. Clutton-Brock et al.
Ž1982 found the same proportion of diurnal to nocturnal foraging in lactating red deer.
hinds.
The slight increase in daylight foraging and reduced bedding in early summer may result from insect harassment. Higher prevalence of nocturnal foraging in spring and
Ž .
summer has been reported in other work on wapiti Gates and Hudson, 1983 and Dall’s
Ž . Ž .
sheep OÕis dalli Hoefs and McTaggart Cowan, 1979 .
The increased length and decreased frequency of foraging bouts in summer agree
Ž .
with other works on wapiti Hanley, 1982; Florkiewicz, 1994; Jiang and Hudson, 1996 .
Ž . Ž
Spring similarities to Jiang and Hudson’s 1996 non-reproductive hinds also at
. Ž
Ministik Jiang and Hudson: 70 minrbout, 9.2 boutsrday; this study: 64 minrbout, 9.7
. Ž
boutsrday and early summer differences Jiang and Hudson: 149 minrbout, 4.8
.
boutsrday; this study: 67 minrbout, 10.2 boutsrday , underscore the effects of lactation. In late gestation, reduced rumen capacity caused by the rapidly developing fetus, may force shorter feeding bouts, while increasing bout frequency. During lacta-tion, although diel grazing times are comparable, lactating hind foraging is regularly interrupted for nursing and other cow–calf interactions.
Adaptation of BR, BS, and grazing time help ruminants regulate and maintain intake
Ž
over a wide range of forage availability Allden and Whittaker, 1970; Arnold and
.
Dudzinski, 1978; Trudell and White, 1981 . For example, at Ministik on spring pasture
Ž . Ž .
of similar phytomass 1880 kg DMrha , Jiang’s 1993 wapiti hinds had lower mean
Ž . Ž .
BR 34 bitesrmin and nearly double the BS 229 mg of those in this study. To satisfy the increased energy demands of fetal growth, BR and grazing time were augmented; however, hinds were apparently unable to meet their nutritional requirements, as reflected in spring weight loss. As this is a natural element in their annual cycle of weight fluctuation, it is likely ineffectual to supplement hind nutrition during pre-parturi-tion. Moreover, calving problems can arise with overweight hinds.
In this study, peak BR occurred in late summer when forage availability was highest. This is unexpected, as other studies have demonstrated, that higher forage availability
Ž
was accompanied by lower BR e.g., Jamieson and Hodgson, 1979; Hudson and
.
Nietfeld, 1985; Heydon et al., 1993; Semiadi et al., 1993 . BR may be limited at both
Ž . Ž
very low through difficulty prehending short, stemmy vegetation and at very high due
.
to time involved masticating large bites phytomass levels. In this study, perhaps the range of higher phytomass values encountered occurred at levels where foraging rate
Ž .
declines with phytomass. Alternatively or additionally , a combination of greater intake requirements and foraging interruptions associated with lactation may demand
aug-Ž
mented BR from hinds in late summer, regardless of forage availability Trudell and
.
White, 1981; Forbes and Coleman, 1987 .
A positive correlation between BS and forage availability has commonly been
Ž
reported for wapiti Wickstrom et al., 1984; Hudson and Nietfeld, 1985; Hudson and
.
Watkins, 1986 . They also found a reciprocal response of BR with BS, and hence, phytomass. This may reflect differences in wapiti ingestion efficiencies. When grazing a well-cropped pasture, wapiti feed very close to the ground where inedible elements may
Ž .
occur, due to objects displacing prehended vegetation andror loss of plant material during manipulation and ejection of inedible portion. These problems are less likely to occur in the lightly grazed pasture, as long grass stems are not easily lost, and wapiti forage in a higher horizon. The possibility of greater ingestion efficiencies on superior pasture may incline managers to graze lactating hinds on the best available summer pasture.
Comparison of whole pasture and feeding patch phytomass indicates that wapiti tend to select foraging locations of higher forage availability. Criteria for selection of foraging location may be based more on vegetation density than plant species.
5. Conclusion
This study demonstrated how modifications in activity and foraging behaviour of gestating and lactating wapiti enable them to satisfy their seasonally changing nutritional requirements.
During late gestation, weight loss occurred despite augmented BRs and grazing time on exceptional spring pasture. Minimal provision of supplemental feed should be necessary, as reduced weight for parturition would be desirable to reduce the risk of calving difficulties. The necessity for frequent nursing sessions during peak lactation disrupts normal patterns of foraging and bedding. Despite high energy demands, hinds achieved compensatory growth.
As BR and grazing time remained constant, this demonstrates the efficacy of manipulating BS in maintaining intake. By late lactation, hinds are able to focus more on foraging and less on tending their older calves. At this time, they capitalize on the still ample high quality forage, to improve condition for breeding. Unlike peak lactation,
Ž
when BS was the compensatory variable, they did so by increasing BR annual
.
maximum , while BS and grazing time remained unchanged. This emphasizes the importance of providing premium pasture in late summer to facilitate condition recov-ery. Hinds grazed fewer hours and less intensively in the lightly grazed pasture, than in the heavily grazed, without compromising liveweight gain. This suggests that it may be advantageous during the post-partum period to move lactating hinds to pastures with higher forage availability, where their ingestion efficiency may be superior.
The information gathered in this study will contribute towards enhanced production of farmed deer, as well as, improved management of wild cervids.
Acknowledgements
Ž
Financial support was provided by the Alberta Agricultural Research Institute Direct
.
Funding Projecta960689 and the Natural Science and Engineering Research Council
References
Allden, W.G., Whittaker, I.A., 1970. The determination of herbage intake by grazing sheep: the interrelation-ships of factors influencing herbage intake and availability. Aust. J. Agric. Res. 21, 755–766.
Arnold, G.W., Dudzinski, M.L., 1978. In: Development in Animal and Veterinary Sciences: 2. Ethology of Free-Ranging Domestic Animals. Elsevier, New York, 198 pp.
Charnov, E.L., 1976. Optimal foraging, the marginal value theorem. Theor. Pop. Biol. 9, 129–136. Clutton-Brock, T.H., Iason, G.R., Albon, S.D., Guinness, G.E., 1982. Effects of lactation on feeding behaviour
and habitat use in wild red deer hinds. J. Zool., Lond. 198, 227–236.
Collins, W.B., Urness, P.J., Austin, D.D., 1978. Elk diets and activities on different lodgepole pine habitat segments. J. Wildl. Manage. 42, 799–810.
Craighead, J.J., Craighead, F.C. Jr., Ruff, R.L., O’Gara, B.W., 1973. Home ranges and activity patterns of nonmigratory elk of the Madison drainage herd as determined by biotelemetry. Wildl. Monogr. 33, 50 pp. Florkiewicz, R.F., 1994. Nutritional ecology of wapiti and carrying capacity of late winter and spring range in
the Yukon. MSc Thesis, University of Alberta, Edmonton, AB, 73 pp.
Forbes, T.D.A., 1989. Researching the plant-animals interface: the investigation of ingestive behaviour in grazing animals. J. Anim. Sci. 66, 2369–2379.
Forbes, T.D.A., Coleman, S.W., 1987. Herbage intake and ingestive behavior of grazing cattle as influenced
Ž .
by variation in sward characteristics. In: Horn, F.W., Hodgson, J., Mott, J.J., Brougham, R.R. Eds. , Proc. Special Session Grazing-Land Res. at the Plant–Animal Interface. Winrock International, Morrilton, AR, pp. 141–152.
Ž .
Gates, C.C., Hudson, R.J., 1983. Foraging behaviour of wapiti in a boreal forest enclosure. Nat. Can. Que. 110, 197–206.
Gittleman, J.L., Thompson, S.D., 1988. Energy allocation in mammalian reproduction. Am. Zool. 28, 863–875.
Hanley, T.A., 1982. Cervid activity patterns in relation to foraging constraints: western Washington. Northwest Sci. 56, 208–217.
Heydon, M.J., Loudon, A.S.I., Milne, J.A., Brinklow, B.R., 1992. Influence of plane of nutrition on seasonal changes in food intake and reproduction of grazing red deer. In: The Biology of Deer. Springer-Verlag, New York, NY, pp. 279–284.
Heydon, M.J., Sibbald, A.M., Milne, J.A., Brinklow, B.R., Loudon, A.S.I., 1993. The interaction of food
Ž
availability and endogenous physiological cycles on the grazing ecology of red deer hinds CerÕus
.
elaphus . Funct. Ecol. 7, 216–222.
Hodgson, J., 1985. The control of herbage intake in the grazing ruminant. Proc. Nutr. Soc. 44, 339–346.
Ž
Hoefs, M., McTaggart Cowan, I., 1979. Ecological investigation of a population of Dall’s sheep OÕis dalli
.
dalli Nelson . Syesis 12, 1–81.
Hudson, R.J., Nietfeld, M.T., 1985. Effect of forage depletion on the feeding rate of wapiti. J. Range Manage.
Ž .
38 1 , 80–82.
Hudson, R.J., Watkins, W.G., 1986. Foraging rates of wapiti on green and cured pastures. Can. J. Zool. 64, 1705–1708.
Hutt, S.J., Hutt, C., 1970. Direct Observation and Measurement of Behaviour. Thomas Publishing, Springfield, IL, 224 pp.
Jamieson, W.S., Hodgson, J., 1979. The effects of variation in sward characteristics upon the ingestive behaviour and herbage intake of calves and lambs under a continuous stocking management. Grass Forage Sci. 34, 273–282.
Ž .
Jiang, Z., 1993. Ecological and morphophysiological constraints on foraging by wapiti CerÕus elaphus . PhD
Thesis, University of Alberta, Edmonton, AB, 114 pp.
Jiang, Z., Hudson, R.J., 1992. Estimating forage intake and energy requirements of free-ranging wapiti
ŽCerÕus elaphus . Can. J. Zool. 70, 675–679..
Jiang, Z., Hudson, R.J., 1996. Ambient temperature, food and foraging activities of wapiti, CerÕus elaphus
Ž .
nelsoni, in aspen boreal forest. Acta Theriol. 16 4 , 250–258.
Semiadi, G., Muir, P.D., Barry, T.N., Veltman, C.J., Hodgson, J., 1993. Grazing patterns of sambar deer
ŽCerÕus unicolor and red deer Cer. Ž Õus elaphus in captivity. N.Z. J. Agric. Res. 36, 253–260..
Spedding, C.R.W., Large, R.V., Kydd, D.D., 1966. The evaluation of herbage species by grazing animals. Proc. 10th Intl. Grassland Cong., Helsinki, Finland, 479–483.
SPSS, 1998. Base 8.0 for Windows — User’s Guide. SPSS, Chicago, IL.
Strong, W.L., 1992. Ecoregions and Ecodistricts of Alberta: Vol. 1. Alberta Forestry, Lands, and Wildlife. Land Info. Services, Edmonton, AB, 77 pp.
Trudell, J., White, R.G., 1981. The effect of forage structure and availability on food intake, biting rate, bites size, and daily eating time of reindeer. J. Appl. Ecol. 18, 63–81.