250 (2000) 3–21
www.elsevier.nl / locate / jembe
Roles of experimental marine ecology in coastal management
and conservation
Juan Carlos Castilla
´ ´ ´
Departamento de Ecologıa, Facultad de Ciencias Biologicas, P. Universidad Catolica de Chile,
Casilla114-D, Santiago, Chile
Abstract
The paper reviews the main findings of rocky shore and subtidal nearshore experimental marine ecology (EME) in cold and temperate marine ecosystems during the past four decades. It analyzes the role of EME in coastal management and conservation. The historical development of strategies for managing single or multispecies fisheries are reviewed. The published results show over-exploitation and depletion of more than 60% of the fish stocks and a lack of connection between the management of fisheries and results derived from experimental marine ecology. This is mainly due to: (a) the different temporal and spatial scale at which most marine ecologists and fishery managers operate; (b) the lack of long-term fishery monitoring and adaptive techniques for management; and (c) limitations in the design of experiments on fisheries. Large-scale oceanic perturbations, due to combinations of excessive resource exploitation and environmental vari-ability coupled with present trends in management approaches are discussed. Modern approaches and tools for management of fisheries, such as Adaptive Management (AM), Territorial User Rights in Fisheries (TURFs), Individual Transferrable Quotas and Non-Transferrable Quotas (ITQs, INTQs) are discussed in the context of small-scale fisheries and EME. Published views on limits of applied ecological research with regards to management of fisheries are discussed. Linkages between EME, marine conservation and the establishment of Marine Protected Areas (MPAs) and experimental exclusions of humans are highlighted. Results derived from MPAs, such as: (a) species or community trophic cascades, and (b) the role of key-stone species and species interaction strengths, are discussed. It is concluded that the role of EME in conservation has been greater than has been the case in management of fisheries. The potential to link EME, conservation and the management of fisheries is exemplified through the proposed establishment in Chile of a connected network of Scientific Reserves, MPAs and TURFs sites. The final conclusion is that to cross-fertilize EME, conservation and management, there are three main challenges: (1) to end the traditional view of approaching the management of fisheries and marine conservation as contradictory / antagonizing issues; (2) to improve communications between experimental marine ecology and the management of fisheries through the implementation of experimentation and adaptive management; (3) to improve linkages between marine conservation, the management of fisheries and social sciences. 2000 Elsevier Science B.V. All rights reserved.
E-mail address: [email protected] (J.C. Castilla).
Keywords: Conservation; Experiment; Humans; Linkages; Marine ecology; Management; Nearshore; Rocky intertidal
1. Introduction
Ecology is the scientific study of the interactions among species and is concerned with the distributions and abundances of organisms (Andrewartha and Birch, 1954). Im-portant progress in answering ecological questions and testing alternative hypotheses has been achieved by the use of experimental work. Experimental marine ecology (EME) is a subdiscipline of ecology, which focuses on the marine realm and related environments. Within this realm, intertidal and inshore coastal ecosystems and their benthic assem-blages (communities) are readily accessible. It is less expensive to develop experimental approaches in such habitats than elsewhere. These ecosystems have served as the key environments where EME originated and produced significant scientific advances. Paine (1994) has reviewed the progress of ecological marine studies since the beginning of the last century and concluded that, during the first three decades, there was a notable development of quantitative methodologies, particularly in the case of marine soft sediments, but experimentation was absent. During the 1940s and 1950s, intertidal rocky shore ecological studies were numerous. They dealt mainly with the description of intertidal zonation patterns and demonstration of its universality (Stephenson and Stephenson, 1949; Guiler, 1955, 1959; Morgans, 1957). The patterns were convincing and causation strongly biased toward physical factors (see Lewis, 1964; Stephenson and Stephenson, 1972). Nevertheless, EME approaches were embryonic. In the 1960s, advanced technology was used to develop a series of descriptive deep-sea benthic studies (Sanders, 1968) and experimental manipulative studies were also initiated, mainly to demonstrate the role of biological interactions in intertidal rocky shores communities (i.e. Connell, 1961; Paine, 1966; Paine and Vadas, 1969). Throughout the 1970s and 1980s, experimental marine ecological studies flourished, particularly in cold and temperate rocky intertidal, subtidal nearshore reefs and kelp communities. The marine ecological literature was flooded with experimental studies (usually developed within short spatial and temporal scales), which nowadays represent the corner-stone of modern EME (i.e. Paine, 1971, 1974, 1977, 1980, 1984; Dayton, 1971, 1985; Connell, 1974; Estes and Palmisano, 1974; Branch, 1975, 1984; Menge, 1976, 1978; Menge and Sutherland, 1976, 1987; Lubchenco, 1978; Underwood, 1978, 1984, 1985, 1986; Estes et al., 1978; Sousa, 1979a,b; Santelices et al., 1980; Duggins, 1980; Castilla and Moreno, 1982; Fairweather et al., 1984; Moreno et al., 1984; Duggins and Dethier, 1985; Moran,
´ ´ ´
In the past 40 years, EME has peaked to the level of a solid scientific discipline and intertidal and nearshore hard substrata systems have provided the basis for classic experiments to test hypotheses about competition, predation, succession, niche, facilita-tion, perturbafacilita-tion, resilience, environmental heterogeneity and species richness. A large array of vertebrates, invertebrates and algae has been used in these endeavors and this represents one of the important contributions of modern EME. Furthermore, during the last 40 years, major ecological paradigms, such as the concept of the key-stone species, community resilience and / or ecologically alternative states and the influence of carnivores and hervibores on patterns of biomass in aquatic systems (species and community trophic cascades) and on species biodiversity have evolved, linked to marine systems (Paine, 1966; Sutherland, 1974; Bond, 1993; Mills et al., 1993; Castilla et al., 1994; Power et al., 1996; Polis, 1999). Nevertheless, during the past century, little scientific progress has been made using experimental approaches to study marine detrital webs and the role of pathogens.
An area that shows little progress, particularly in cold and marine temperate systems (tropical ecosystems are not addressed here), is the cross-linkage between EME and resource management. This has most probably been due to limitations in the design of fishery experiments (Underwood, 1990; McAllister and Peterman, 1992) and issues of large temporal and spatial scales. Nevertheless, there are stronger connections between EME and conservation issues (i.e. Mills et al., 1993; Heywood, 1999) than have been highlighted to date.
In this paper, I analyze EME studies to link them to conservation and management. I suggest that, EME, marine conservation and management should be approached through the use of long-term monitoring schemes, taking into account the relevant spatial and temporal scales, adaptive strategies and, whenever possible, the use of experimental protocols. Otherwise these activities will continue to be dissociated. Furthermore, I stress the need for the explicit incorporation of humans in EME protocols and urge scientists to bridge communication barriers between ecology, conservation, management and social sciences.
2. Management and conservation: definitions
Conservation is a term that has evolved in the past 150 years (reviewed by Meffe and Carroll, 1994; Heywood and Watson, 1995). In its widest sense it refers to the protection, use and sharing of benefits of the biosphere in a sustainable way (IUCN / UNEP/ WWF, 1980). In its narrowest sense, as used in the Convention of Biological Diversity, it embraces only ’preservation’ or the maintenance of some or all components of biological diversity. Furthermore, definitions of conservation vary between countries and agencies. For example, the Chilean legislation defines conservation as the ’adminis-tration (management) of the biosphere by the human being so that the greatest and sustainable benefits for the present and future generations are achieved’ (Castilla, 1996). On the other hand, conservation (i.e. wildlife rehabilitation; Estes, 1998) may also be perceived as ’a public demand’ and be interpreted as a major confrontation with the continued use of the resources.
3. Management and experimental marine ecology
3.1. Approaches to management of single and multispecies resources
(but see Ketchen (1956), Fukuda (1962) and the experimental options discussed by Parma and Deriso, 1990). The scheme for management in that example is based on a long-term monitoring program and it appears to contain some of the components of a research program (Underwood, 1990; Hurlbert, 1994; but see Parma and Deriso, 1990). According to McCaughran (1996), the IHC: (1) has control over short-term economic desires, (2) the Halibut fishermen tend to be conservative in their catches and (3) the fishermen share IHC’s goals for conservation of the stocks. Another example refers to the management of the Australian western rock lobster, Panulirus cygnus, where (1) an index of recruitment has been used for over 35 years, (2) there is on-going monitoring of the stocks, and (3) there exists an adaptive management program for the fishery. These have resulted in the long-term sustainable management of the fishery (Caputi et al., 1996). No obvious connection between EME findings and these two examples of successful fisheries can be seen. At the other extreme are managerial schemes for single species fisheries in which the assessments of stock may have been wrong, resulting in collapses of the fishery and in community cascading effects throughout the whole ecosystem (National Research Council, 1999).
Multispecies (Sissenwine and Dann, 1991) and ecosystem (Sherman et al., 1993) approaches to modelling fisheries appear to be ecologically more sound. In some cases (Murawski, 1991), a degree of success in the understanding of interactions among species and the consequences of alternative forms of management have been reported. Present trends in management of multispecies fisheries show the incorporation of modelling, including trophic interactions among the species. This allows inferences to be drawn about possible responses by the whole ecosystem (Pauly and Christensen, 1995; Christensen and Pauly, 1998; for a historical perspective, see Gulland, 1982). This opens up an opportunity for the future integration of findings about trophic interactions from EME, modelling and management.
3.2. The ecological approach to management of fisheries
can be overcome if: (1) experimentation is small-scale; (2) fishers are compensated when cuts in fishing effort are required; (3) fishers believe that experimentation is in their best interests and (4) control populations are held in reserve in case overharvesting occurs in a treated population. Controls are of course needed to evaluate the experi-ments. Controversies about such issues have been present in management of fisheries for the past 120 years (Huxley, 1884; Lankester, 1884).
In the future, according to Ludwig et al. (1993) and Hilborn and Ludwig (1993), the approach should be: (1) to continue fishing; (2) to monitor; (3) to get rid of frequentist statistical methods; and (4) to be adaptive. The sad fact is that in the almost total absence of experimentation and / or ecological advice (although poor) a large number of marine resources have been overexploited or depleted (Botsford et al., 1997). It is not known to what degree ecological inputs would have helped to avoid such a situation. Nevertheless, one thing is clear: such ecological inputs would have required testing hypotheses and, therefore, some degree of experimentation. A large number of collapses of fisheries have happened under the use of inappropriate management of single species (mainly using production or yield-per-recruit models). Hilborn and Ludwig’s (1993) prescription seems to be to continue fishing in the vacuum of knowledge about interacting processes. Accepting the limitations of experimental ecological research, and being myself much in favor of Adaptive Management, I do not completely agree with Hilborn and Ludwig’s (1993) suggestions. In fact, although ecological research may not be the most productive way to determine the limits of sustainability of any fishery, such limits cannot continue to be approached without better knowledge about ecosystems. Long-term experimentation in fisheries, despite its limitations, has to be done. There is a need to test hypotheses. Currently small-scale benthic invertebrate fisheries appear as the best candidates. Nevertheless, McAllister and Peterman (1992) reviewed several experiments on management of fin-fish (single species management of salmon and halibut and multispecies fisheries), where improvements on their designs, regarding controls over temporal and spatial variability, randomization, interspeciation, contrasting treatments and spatial replication, are needed, but possible. Furthermore, there are new managerial tools (see later, Section 3.5) that will enhance our future ability to do experiments on management of fisheries and to make bridges with findings from EME. Of course, to be able to develop experimentation in any fishery we will have to get rid of the ’ratchet effect’. This is the problem that managers under constant socio-political and economic pressure for greater harvesting of fish, to maximze short-term benefits, continue to allow catches to increase (Ludwig et al., 1993; Mace, 1996).
3.3. Small-scale fisheries
1999). What is being learned in small-scale fisheries, where small-scale and experimental ´
replication are becoming possible (Castilla and Fernandez, 1999), may be applied in the future to large-scale fisheries. This is a major challenge ahead and limitations are inevitable.
3.4. Large-scale perturbations
Modificiations of major oceanic ecosystem due to large-scale human perturbations, such as heavy exploitation and / or coupling with changes in the physical environment, have been reported from different areas of the world. For example, the Bering Sea and Newfoundland Sea (Myers et al., 1997; National Research Council, 1999) and the Barents Sea (Hamre, 1994) have suffered. Nevertheless, there are still too many scientific uncertainties to understand relatively ’simple’ modifications to ecosystems and to ascribe causality to the observed patterns (National Research Council, 1999). Large-scale modifications to fisheries, seem not to be fully used in a progressive adaptive management so as to gain insights into the dynamics and resilience of large ecosystems (but see Estes et al., 1998).
3.5. Present trends and the possibility of linking EME and fishery management
During the late 1980s and 1990s, new approaches to management of fisheries and new legal tools have been implemented. These are challenging the old paradigms and provide opportunities for more rational management and the application of knowledge generated through EME. I highlight three managerial tools which may facilitate the cross-linkage between EME and management of fisheries.
3.5.1. Adaptive management (AM)
3.5.2. Individual transferable quotas (ITQs)
In fisheries, good management is bound to be hampered by the lack of private (or communal) rights over the resource. This, under an open regime of access to the fishery, gives rise to the problem of common property or the ‘‘tragedy of the commons’’ (Hardin, 1960). ITQs consider the allocation of a share of the total allowable catches (under the management of single species) either to individual or to communal users of the resource. Quotas can be allocated for defined periods and constitute a quasi-right to property (Dewes, 1998). ITQs vary according to the resource, legislation, ethnic group, rental and the necessary mechanisms of adjusting and recovery by governments. They were among the first tools implemented to stop ’the race for the fish’. ITQs constrain the participants from seeking only to maximize profits (particularly in traditional fisheries). If the stake-holders in the resource have some degree of ownership in the stocks, they may still seek to maximize profits, but they will also be more willing to work with scientists in schemes of Adaptive Management. Further, the design of experimental fisheries implementation of ecosystem approaches and using ecological knowledge will be greatly enhanced. An alternative to ITQs, the Individual Non-Transferrable Quotas (INTQs), was experimentally used in Chile in the 1990s, with the management of the dive-fishery of the mollusc ’loco’, Concholepas concholepas (Castilla, 1994). They were implemented for the management of benthic resources jointly with the establishment of Territorial Use Rights in Fisheries (TURFs, see below) and under plans of co-management. These tools have enhanced the possibility to do local benthic resource re-stocking, rotational extractions and extractive experiments. They have also permitted the incorporation of knowledge from EME into the activities of small-scale fisheries
´ ´
(Fernandez and Castilla, 1997; Castilla and Fernandez, 1998; Castilla et al., 1998; ´
Castilla, 1999; Fernandez et al., 2000). Above all, INTQs have permitted the incorpora-tion of fishers as active role-players in the processes of management.
3.5.3. Territorial ‘‘use rights’’ in fisheries (TURFs)
and their application to pelagic fisheries is not easily foreseen, they have shown that critical constraints in the management of marine fisheries can be overcome. After all, experimentation, although limited, can be approached in the sea and TURFs present a potential for suitable replication and control in the design of such experiments (Castilla
´
and Fernandez, 1999). The challenges are to perfect and expand this direction, to include the resource users in management, to be more holistic in the approach (i.e. to use ecosystem approaches), to use schemes for managing multispecies fisheries and, thereby, to bridge fisheries management and EME.
4. Conservation and experimental marine ecology
4.1. Marine protected areas (MPAs)
MPAs have been established worldwide as a recognition of the conservation crisis, over-exploitation of marine resources and the misuse of coastal areas. They have provided an opportunity for studies of ecosystems and for understanding the long-term dynamics of marine systems. MPAs have been shown to be important tools to protect critical habitats, to provide protection from pollution, to increase the number and size of individuals of fished species, to serve as ’seeding grounds’ (i.e. to supply recruits to fished areas) and for ‘‘spillover’’ objectives (i.e. the export of biomass; Castilla, 1999). The increase or decrease, within MPAs, of predator, producer, competitor, engineer species, etc., may have different ecological consequences to the structure, dynamics or resilience of the community under conservation. The establishment of MPAs can result in different ecological results at the local or regional scale. In essence, the establishment of MPAs represents a human intervention into marine systems from which humans or some of their activities are totally or partially excluded. MPAs and other conservation tools (preserves, marine parks, marine sanctuaries, isolated islands, coastal areas protected by legal decrees such as shores around yales, military facilities) have provided unique observational and experimental opportunities for EME. An example of this is the test of the ecological role of predators and humans in cascades of species or community trophic interactions on rocky shores (i.e. Estes et al., 1998; Lindberg et al., 1998; Castilla, 1999). Nevertheless, in most cases, because of the MPAs’ locations, number or extension, the experiments have not been properly designed (e.g. there has been no proper replication). Alternative statistical approaches have been used instead. For instance, comparisons of means between two populations or ’before and after’ paired comparisons (MPAs versus outside localities; Underwood, 1991, 1997) or Bayesian inference concerning the difference between the two means (Box and Tiao, 1992). Confounding influences on the results exist and causality needs to be interpreted carefully. On the other hand, MPAs’ experimental approaches have notable advantages over small-scale experimental plots. They involve large components of an ecosystem and allow for direct ’exclusion of humans’. Results obtained from MPAs are more readily transferrable into societal issues, such as those of conservation and management (Castilla, 1999).
as tools for experimentation. In the future, they may play a critical role in the saga of the ´
resources of the oceans (Hockey and Branch, 1997; Castilla and Fernandez, 1999). Nevertheless, at present, most of the MPAs in the world are not under appropriate scientific scrutiny or regular monitoring. As ‘‘no-take’’ areas, most MPAs are fulfilling only one part of their main role. This is the conservation of a whole or a portion of an ecosystem. Concurrently, however, critical knowledge on the structure, dynamics and resilience of the preserved ecosystems has not been addressed, due to the lack of appropriate monitoring.
4.2. Human-exclusion experiments
In the marine literature, there are several partial or total manipulations of human activities (MPAs or other areas). These have had different objectives, methodologies and were at variable spatial and time-scales. Three cases involving predators, which are relevant examples in the EME literature and have had important consequences for conservation and / or managerial issues, are highlighted. First is the interactions among the sea otter, Enhydra lutris, sea urchins and kelp-beds in the Pacific North West and Alaska (Estes et al., 1978, 1998; Estes and Duggins, 1995). Second is the interactions between the rock lobsters Jasus lalandii, whelks and bivalves in South Africa (Barkai and Branch, 1988; Barkai and McQuaid, 1988; Castilla et al., 1994). The final example is that of the muricid snail Concholepas concholepas and its interactions with bivalves and barnacles in Chile (Castilla, 1999).
Two of these predators, Jasus and Concholepas, eat competitively dominant mussels, Choromytilus and Perumytilus. Enhydra eats herbivorous sea urchins. Jasus, Con-cholepas and Enhydra are, or have been, overfished or overhunted. The experimental reduction of fishing / hunting pressure, through legal decrees or the establishment of MPAs, has permitted the evaluation of their roles in the structure and dynamics of marine communities. It has also allowed evaluation of the roles of humans. For the sea otter, a recent change of biological interactions, probably triggered by large anthro-pogenic influences in oceanic ecosystems in the Northern Pacific (by offshore fisheries), has resulted in the increase of predation on otters by the top-level killer whales, Orcinus orca. This has cascaded into an increment of the populations of urchins and in macroalgal deforestation in Alaska (Estes et al., 1998; see Hairston et al., 1960 - HSS hypothesis, but see Polis (1999) for terrestrial ecosystems). This represents a classic example of a trophic cascade throughout a community. For C. concholepas, predation by humans and the cascading effects in the intertidal community where humans were excluded were studied at the Las Cruces MPA in central Chile. The knowledge acquired on ecosystem functioning and the rate of replenishment of resources (i.e. gastropods, sea urchins and crabs) inside the MPA have been used for conservation and management of fisheries. Based on these results, the government of Chile regulated access to benthic resources by divers through the establishment of TURFs, called Management and
´
in the country (Castilla, 1994, 1999). This case represents an example from which the results from a long-term EME protocol led to the creation of new tools for management, conservation and legislation.
4.3. Key-stone species and strength of interactions among species
In ecology, conservation and management, the concept of predatory key-stone species (Paine, 1966, 1969) has been widely used, cited, popularized, but also attacked. It has been misused when associated with that of a ’key-stone prey’, ’key-stone host’ and ’key-stone modifier’ species (Power et al., 1996). This has produced confusion in the ecological literature. Hurlbert (1997) considered that key-stone is not an ecological concept, but ’a casual metaphor’. It is a fact, however, that, in the past 30 years, few ecological concepts (metaphors) such as the key-stone species have been so frequently used in EME, exported to other systems and applied in conservation and management. It is known that dominant species (by abundance or biomass) are critical for the maintenance of the structure and dynamics of communities. They provide most of the energy-flow (as primary producers), or, as in the case of engineer species (Jones et al., 1994), habitat for other organisms. In contrast, a key-stone species is defined as one whose impact on its community or ecosystem is disproportionately large relative to its abundance (Power et al., 1996). Starfishes, sea otters, gastropods and humans can serve as examples (see review by Castilla, 1999). Mills et al. (1993) suggested that dropping of the key-stone species concept may ’be compensated by the development of management and policy guidelines that more explicitly account for the complexity of interactions in natural systems’. The conservation of natural ecosystems depends on our understanding of their structure, dynamics and resilience. I do not see how the dropping of a concept will enhance the development of wise conservation policies. Conversely, if an ecological concept, such as key-stone, is used in its proper context in conservation and management, it may produce useful results (Castilla et al., 1994; Estes et al., 1998; Castilla, 1999).
Recently, the issue of variation in the strengths of interactions among species, a theme related to key-stone species, has been reviewed (Berlow et al., 1999). According to these authors this is a key variable in most dynamic models of food-webs. Understanding it may guide management of natural resources and conservation. For example, it will help to predict the potential consequences in a community due to loss or severe reductions in density of species (i.e. in situations of over-fishing). This could enhance communication between empiricists and theoreticians, ecologists and fishery managers.
5. Bridging experimental marine ecology, conservation and management
Conservation and management are part of human goals, while experimental marine ecology is a scientific subject. Can they be bridged? Can they be linked? From the perspective of EME, it is important to notice that the future options for conservation and management are moving toward MPAs, TURfs, ITQs, INTQs, community-based and adaptive management. Cross-linkages will be easier and stronger. Further, the closer conservation / management schemes are to the logical components of a research program (Underwood, 1990, 1996, 1997; with the incorporation of resource-users), the cross-fertilization will be enhanced. The tools for conservation and management considered here focus on the use of the resources and habitat for the users’ direct benefit, whether users are individuals, a group or a whole community. This facilitates a more rational approach to the use of resources. It also promotes experimental / adaptive management and joint approaches to conservation (Castilla, 1996, 1999). Moreover, the implementa-tion of these tools has extra incentives. For instance, Castilla (1996) and Roberts (1997) highlighted the added value of having marine spatial connectivity with the schemes for conservation and / or management. If MPAs and TURFs are established with a view to ensuring spatial connections (i.e. source and sink areas and ocean currents), so as to enhance dispersal routes for pelagic larvae, spores or juveniles, then a network of MPAs or TURFs may result in greater yield in the fisheries and may also be used as replicate experimental units. Additionally, goals for management of ecosystems may be easily incorporated. To achieve this, much progress is needed in legislation for conservation and management and in the understanding of ecosystem inter-linkages, particularly for open populations.
In Chile, such connectivity, for the conservation and management of benthic resources along the coast, has been proposed. Progress has been made. The government has planned more than 160 MEAs along approximately 4000 km of coastline for the year 2000 (including TURFs, each of mean bottom surface area of approximately 150 ha;
´
Castilla and Fernandez, 1999), for the management of benthic resources. Furthermore, along the coast there are three small research MPAs (less than 10 ha each) which for the last 20 years have provided scientific ecological knowledge on the structure and dynamics of coastal systems (Castilla, 1996). What has been proposed is the further inclusion in this set-up of several large marine parks (Castilla, 1996; Castilla and
´
Fernandez, 1999; Fig. 1). These MPAs should have geographical and ecosystem representation and serve as units for conservation and science, but also will be recreational and educational entities. They should, however, also be for the enhancement of nearby populations, via ’overspilling and seeding grounds’. Field experience has shown that mechanisms operate on the coast of Chile to export resources from MPAs
´
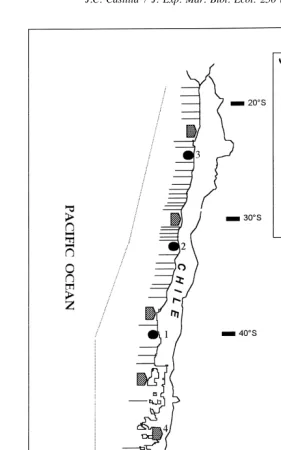
Fig. 1. Schematic representation (not at scale) of MPAs and TURFs areas for the coast of Chile. In 1999 the government designed 168 Management and Exploitation Areas (MEAs5TURFs) for the co-management of
benthic resources (only some of them are shown in the figure). Two scientific MPAs coastal Reserves ´
(15Mehuın; 25Las Cruces) and one MPA Reserve protected by Minera Escondida Ltd. at Antofagasta (3)
´
have over 10 year intertidal ecological monitoring schemes (Mehuın MPA was abandoned 2 years ago). Laguna San Rafael (4) is a terrestrial Park including portions of water. The Marine Parks shown in the figure do not exist. Their number, location and establishment need to be decided and approved by the government. The area surrounding the coast of Chile represents the 5-mile exclusive zone for artisanal fisheries (Chilean
´
These management procedures are in urgent need of implementation in the world, because present principles for managing or conserving resources recommend the adoption of ecosystem-based approaches (National Research Council, 1999). Advanced world legislation considers conservation as a tool for management (Castilla, 1996; Attwood et al., 1997; National Research Council, 1999). Recent models are evaluating fishery yields from MPAs comparing them with those produced through schemes of open-access fishery (Hastings and Botsford, 1999). The attractiveness of a spatially combined scheme is that it may serve conservation, biodiversity and the goals of fishery management. It may enhance connectivity and scientific, educational and recreational objectives (Ballantine, 1996; Castilla, 1996; Attwood et al., 1997; Roberts, 1997). To achieve this, two major challenges exist. First, it is crucial to end traditional views of conservation and management as contradictory issues. In the ocean, physical, chemical and biological processes are strongly linked (much stronger than in terrestrial systems). Goals for conservation and management should be co-ordinated to benefit humans, communities, associations, groups of people and not just an economic model. Then, it will be simpler to bridge conservation, management and experimental marine ecology. This, in the long-term, may lead to achieving the sustainability of marine resources and habitats and to ecosystem approaches.
The second challenge will be to improve communication between ecological findings, risks, management and conservation decisions, since they are associated with social values (Norton, 1998). Furthermore, a new code of conduct and, accordingly, the crafting of new local and regional institutions will be urgently needed (Botsford et al., 1997; Castilla, 1999).
Acknowledgements
I sincerely appreciate Tony Underwood’s invitation to participate in the celebration of volume 250 of JEMBE. The Presidential Chair in Science (Chile) and the Pew Charitable Fund Fellowship in Marine Conservation have significantly helped my research activities and paper writing during the past 4 years. I acknowledge their financial support. The paper reviews ecological work I have developed in Chile during the past 20 years, jointly with my students, assistants, Chilean and foreign marine colleagues. While working on the review, I faced the problem of selecting references to illustrate concepts and lessons derived from experimental marine ecology in the issues of management and conservation. I hope authors not included understand the constraints. I
´
sincerely appreciate the critical review of Miriam Fernandez, two anonymous reviewers and Tony Underwood’s tuning up of the manuscript. The paper is dedicate to the
´
memory of Professor Patricio Sanchez. [AU]
References
Attwood, C.G., Harris, J.M., Williams, A.J., 1997. International experience of marine protected areas and their relevance to South Africa. S. Afr. J. Mar. Sci. 18, 311–332.
Ballantine, W.J., 1996. No-take marine reserve networks support fisheries. In: Hancock, D.A., Smith, D.C., Grant, A., Beumer, J.P. (Eds.), Second World Fisheries Congress, CSIRO, Brisbane, Australia, pp. 702–706.
Barkai, A., Branch, G.M., 1988. Energy requirements for a dense population of rock lobster Jasus lalandii: novel importance of unorthodox food sources. Mar. Ecol. Prog. Ser. 50, 83–96.
Barkai, A., McQuaid, C.D., 1988. Growth and mortality of the mussels Choromytilus meridionalis (Kraus) and
Aulacomya ater (Molina) as indicators of biotic conditions. J. Molluscan Stud. 55, 239–342.
Berlow, E.L., Navarrete, S.A., Briggs, C.J., Power, M.E., Menge, B.A., 1999. Quantifying variation in the strengths of species interactions. Ecology 80, 2206–2224.
Beverton, R.J.H., Holt, S. (Eds.), 1957. On the Dynamics of Exploited Fish Populations. Her Majesty’s Stationery Office, London, p. 533.
Bond, W.J., 1993. Keystone species. In: Schulze, E.D., Mooney, H.A. (Eds.), Ecosystem Functioning and Biodiversity. Springer, Berlin, pp. 237–253.
Botsford, L.W., Castilla, J.C., Peterson, C.H., 1997. The management of fisheries and marine ecosystems. Science 277, 509–515.
Box, G.E.P., Tiao, G.C., 1992. Bayesian inference in statistical analysis. John Wiley and Son, Inc., N. York. 587 pp.
Branch, G.M., 1975. Intraspecific competition in Patella cochlear Born. J. Anim. Ecol. 44, 263–281. Branch, G.M., 1984. Competition between marine organisms: ecological and evolutionary implications.
Oceanogr. Mar. Biol. Rev. 22, 429–593.
Bustamante, R.H., Branch, G.M., 1996. The dependence of intertidal consumers on kelp-derived organic matter on the west coast of South Africa. J. Exp. Mar. Biol. Ecol. 196, 1–28.
Bustamante, R.H., Branch, G.M., Eekhout, S., 1995. Maintenance of an exceptional intertidal grazer biomass in South Africa: subsidy by subtidal kelps. Ecology 76, 2314–2329.
Caputi, N., Chubb, C., Hall, N., Pearse, A., 1996. Relationships between different life history stages of the western rock lobster, Panulirus cygnus, and their implications for management. In: Hancock, D.A., Smith, D.C., Grant, A., Beumer, J.P. (Eds.), Second World Fisheries Congress, CSIRO, Brisbane, Australia, pp. 579–585.
Casey, J.M., Myers, A., 1998. Near extinction of a large, widely distributed fish. Science 281, 690–692. Castilla, J.C., 1994. The Chilean small-scale benthic shellfisheries and the institutionalization of new
management practices. Ecol. Int. Bull. 21, 47–63.
´ Castilla, J.C., 1996. La futura red chilena de Parques y Reservas Marinas y los conceptos de conservacion,
´ ´
preservacion y manejo en la legislacion nacional. Rvta. Chilena Hist. Nat. 69, 253–270.
Castilla, J.C., 1999. Coastal marine communities: trends and perspectives from human-exclusion experiments. TREE 14, 280–283.
Castilla, J.C., Moreno, C.A., 1982. Sea urchins and Macrocystis pyrifera: experimental test of their ecological relations in southern Chile. In: Lawrence, J.M. (Ed.), International Echinoderms Conference, A.A. Balkema, Rotterdam, pp. 257–263.
´
Castilla, J.C., Duran, L.R., 1985. Human exclusion from the rocky intertidal zone of central Chile: the effects on Concholepas (Gastropoda). Oikos 45, 391–399.
´
Castilla, J.C., Fernandez, M., 1998. Small-scale benthic fisheries in Chile: on co-management and sustainable use of benthic invertebrates. Ecol. Appl. 8, S124–S132.
´
Castilla, J.C., Fernandez, M., 1999. Coastal marine community / ecosystem approaches in invertebrate multispecies management: ’Take’ and ’No-take’ areas network and Territorial Use Rights in Fisheries (TURFs). In: The Norway / UN Conference on Ecosystem Approach for Sustainable Use of Biological Diversity. September, Trondheim, Norway, p. 9.
Castilla, J.C., Branch, G.M., Barkai, B., 1994. Exploitation of two critical predators: the gastropod
Concholepas and the rock lobster Jasus lalandii. In: Siegfried, R.W. (Ed.), Rocky Shores: Exploitation in
Chile and South Africa. Springer, Berlin, pp. 101–130.
Chapman, M.G., 1986. Assessment of some controls in experimental transplants of intertidal gastropods. J. Exp. Mar. Biol. Ecol. 103, 181–201.
Christensen, V., Pauly, D., 1998. Changes in models of aquatic ecosystems approaching carrying capacity. Ecol. Appl. 6, 655–691.
Connell, J.H., 1961. Effects of competition, predation by Thais lapillus, and other factors on natural populations of the barnacle Balanus balanoides. Ecol. Monogr. 31, 61–104.
Connell, J.H., 1974. Ecology: field experiments in marine ecology. In: Mariscol, R.N. (Ed.), Experimental Marine Ecology. Academic Press, New York.
Dayton, P.K., 1971. Competition, disturbance and community organization: the provision and subsequent utilization of space in a rocky intertidal community. Ecol. Monogr. 41, 351–389.
Dayton, P.K., 1985. Ecology of kelp communities. Annu. Rev. Syst. Ecol. 16, 215–245.
Dewes, C.M., 1998. Effects of individual quota systems on New Zealand and British Columbia. Ecol. Appl. 8, S133–S138.
Duggins, D.O., 1980. Kelp beds and sea otters: an experimental approach. Ecology 61, 447–453.
Duggins, D.O., Dethier, M.N., 1985. Experimental studies of herbivory and algal competition in a low intertidal habitat. Oecologia 67, 183–191.
´
Duran, L.R., Castilla, J.C., 1989. Variation and persistence of the middle rocky intertidal community of central Chile, with and without human harvesting. Mar. Biol. 103, 555–562.
´
Duran, L.R., Castilla, J.C., Oliva, D., 1986. Intensity of human predation on rocky shores at Las Cruces, central Chile. Environ. Conserv. 14, 143–149.
Estes, J.A., 1998. Concerns about rehabilitation of oiled wildlife. Conserv. Biol. 12, 1156–1157.
Estes, J.A., Palmisano, J.F., 1974. Sea otters: their role in structuring nearshore communities. Science 185, 1058–1060.
Estes, J.A., Duggins, D., 1995. Sea otter and kelp forests in Alaska: generality and variation in a community ecological paradigm. Ecol. Monogr. 65, 75–100.
Estes, J.A., Smith, N.S., Palmisano, J.F., 1978. Sea otter predation and community organization in the western Aleutian Islands, Alaska. Ecology 59, 822–833.
Estes, J.A., Tinker, M.T., Williams, T.M., Doak, D.F., 1998. Killer whale predation on sea otter linking nearshore and oceanic ecosystems. Science 282, 473–476.
Fairweather, P.G., Underwood, A.J., Moran, M.J., 1984. Preliminary investigations of predation by the whelk
Morula marginalba. Mar. Ecol. Prog. Ser. 17, 143–156.
´
Fernandez, M., Castilla, J.C., 1997. The Chilean artisanal Stone crab (Homalaspis plana) fishery: catch trends in open access zones and the effect of Management Areas in central Chile. J. Shellfish Res. 16, 371–377.
´
Fernandez, M., Castilla, J.C., Alvarado, J., 2000. The Chilean small-scale shellfishery: what factors affect fishers behavior? Ocean Coast. Manag. (submitted).
Fukuda, Y., 1962. On the stocks of halibut and their fisheries in the northeastern Pacific. Bull. Int. North Pacific Fish. Comm. 7, 39–50.
Guiler, E.R., 1955. Australian intertidal belt-forming species in Tasmania. J. Ecol. 43, 138–148.
Guiler, E.R., 1959. The intertidal ecology of the Montemar area, Chile. Pap. Proc. R. Soc. Tasm. 93, 165–183. Gulland, J.A., 1982. Management policies and models of pelagic stocks: the Peruvian experience 1960-1972.
´
In: Castilla, J.C. (Ed.), Segundo Seminario Taller: bases biologicas para el uso y manejo de recursos
´ ´ ´
naturales renovables: recursos biologicos marinos. Monografıas Biologicas, Vol. 2, pp. 65–74. Gulland, J.A., 1983. Fish Stock Assessment. A Manual of Basic Methods. Wiley, Chichester.
Hairston, N.G., Smith, F.E., Slobodkin, L.B., 1960. Community structure, population control, and competition. Am. Nat. 94, 421–425.
Hamre, J., 1994. Biodiversity and exploitation of the main fish stocks in the Norwegian-Barents Sea ecosystem. Biodivers. Conserv. 3, 473–492.
Hardin, G., 1960. The tragedy of the commons. Science 162, 1243–1248.
Hastings, A., Botsford, L.W., 1999. Equivalence in yield from marine reserves and traditional fisheries management. Science 284, 1537–1538.
Heywood, V.H. (Ed.), 1999. The Global Biodiversirty Asessment. Press Syndicate of the University of Cambridge, Cambridge, UK, Published for the United Nations Environmental Programme.
Hilborn, R., Ludwig, D., 1993. The limits of applied ecological research. Ecol. Appl. 3, 550–552. Hjort, J., 1914. Fluctuations in the great fisheries of Northern Europe. Rapports Process-Verbaux Reunions
Cons. Int. Explor. Mer. 20, 1–228.
Hockey, P.A.R., Branch, G.M., 1997. Criteria, objectives and methodology for evaluating marine protected areas in South Africa. S. Afr. J. Mar. Sci. 18, 369–383.
Holling, C.S. (Ed.), 1978. Adaptive Environmental Assessment and Management. Wiley, New York. Hurlbert, S.H., 1994. Pseudoreplication and the design of ecological field experiments. Ecol. Monogr. 54,
184–211.
Hurlbert, S.H., 1997. Functional importance vs. keystoneness: reformulating some questions in theoretical biocenology. Aust. J. Ecol. 22, 369–382.
Huxley, T., 1884. Inaugural address. Fisheries Exhibition Literature 4, 1–22.
IUCN / UNEP/ WWF, 1980. World Conservation Strategy: Living Resource Conservation for Sustainable Development. Gland, Switzerland.
Jones, C.C., Lawton, J.H., Shachak, M., 1994. Organisms as ecosystem engineers. Oikos 69, 373–386. Ketchen, K.S., 1956. Climatic treds and fluctuations in yield of marine fishes in the northeast Pacific. J. Fish.
Res. Board Can. 13, 357–374.
Lankester, R., 1884. The scientific results of the exhibition. Fisheries Exhibition Literature 4, 405–445. Larkin, P.A., 1977. An epitaph for the concept of maximum sustainable yield. Trans. Am. Fish. Soc. 106,
1–11.
Lewis, J.R., 1964. The Ecology of Rocky Shores. English University Press, London.
Lindberg, D.R., Estes, J.A., Warheit, K.I., 1998. Human influences on trophic cascades along rocky shores. Ecol. Appl. 8, 880–890.
Lubchenco, J., 1978. Plant species diversity in a marine intertidal community: importance of herbivore food preferences and algal competitive abilities. Am. Nat. 112, 23–39.
Ludwig, D., Hilborn, R., Walters, C., 1993. Uncertainty, resource exploitation, and conservation: lessons from history. Science 260, 17–36.
Mace, P.M., 1996. Developing and sustaining world fisheries resources: the state of the science and management. In: Hancock, D.A., Smith, D.C., Grant, A., Beumer, J.P. (Eds.), Second World Fishery Congress, CSIRO, Brisbane, Australia, pp. 1–22.
Margalef, R. (Ed.), 1997. Our Biosphere. Excellence in Ecology, Vol. 10. Ecology Institute, Germany. McAllister, M.K., Peterman, R.M., 1992. Experimental design in the management of fisheries: A review. North
Am. J. Fish. Manag. 12, 1–18.
McCaughran, D.A., 1996. Seventy-five years of halibut management success. In: Hancock, D.A., Smith, D.C., Grant, A., Beumer, J.P. (Eds.), Second World Fishery Congress, CSIRO, Brisbane, Australia, pp. 680–686. Meffe, G.K., Carroll, R. (Eds.), 1994. Principles of Conservation Biology. Sinauer Associates, Sunderland,
MA, p. 600.
Menge, B.A., 1976. Organization of the New England rocky intertidal community: role of predation, competition and environment heterogeneity. Ecol. Monogr. 46, 355–393.
Menge, B.A., 1978. Predation intensity in a rocky intertidal community. Relation between predator foraging activity and environmental harshness. Oecologia 34, 1–16.
Menge, B.A., Sutherland, J.P., 1976. Species diversity gradients: synthesis of the role of predation, competition, and temporal heterogeneity. Am. Nat. 110, 351–369.
Menge, B.A., Sutherland, J.P., 1987. Community regulation: variation in disturbance, competition and predation in relation to environmental stress and recruitment. Am. Nat. 130, 730–757.
Mills, L.S., Soule, M.E., Doak, D.F., 1993. The keystone-species concept in ecology and conservation. BioScience 43, 219–224.
Moran, M.J., 1985. Effects of density, prey size and predator size on rates of feeding by an intertidal predatory gastropod Morula marginalba Blainville (Muricidae) on several species of prey. J. Exp. Mar. Biol. Ecol. 90, 97–105.
Moreno, C.A., Sutherland, J.P., Jara, F.H., 1984. Man as a predator in the intertidal zone of southern Chile. Oikos 42, 155–160.
Morgans, J.F.C., 1957. The benthic ecology of False Bay. Part I: the biology of infratidal rocks, observed by diving, related to that of intertidal rocks. Tran. Roy. Soc. S. Afr. 1, 387–443.
Myers, R.A., Hutchings, J.A., Barrowman, N.J., 1997. Why do fish collapse? The example of cod in Atlantic Canada. Ecol. Appl. 7, 91–106.
National Research Council, 1999. In: Sustaining Marine Fisheries. National Academy Press, Washington, DC, p. 164.
Norton, B.G., 1998. Improving ecological communication: the role of ecologists in environmental policy formation. Ecol. Appl. 8, 350–364.
Paine, R.T., 1966. Food web complexity and species diversity. Am. Nat. 100, 65–75.
Paine, R.T., 1969. The Pisaster-Tegula interaction: prey patches, predatory food preference and intertidal community structure. Ecology 52, 1096–1106.
Paine, R.T., 1971. A short term experimental investigation of resource partitioning in a New Zealand rocky intertidal habitat. Ecology 50, 950–961.
Paine, R.T., 1974. Intertidal community structure: experimental studies on the relationship between a dominant competitor and its principal predator. Oecologia 15, 93–120.
Paine, R.T., 1977. Controlled manipulations in the marine intertidal zone, and their contributions to ecological theory. Changing scenes in natural sciences, 1776-1976. Acad. Nat. Sci. Philos. Spec. Publ. 12, 245–270. Paine, R.T., 1980. Food webs: linkage, interaction strength and community infrastructure. J. Anim. Ecol. 49,
667–685.
Paine, R.T., 1984. Some approaches to modeling multispecies systems. In: May, R.M. (Ed.), Dahlen Konferenzen, Exploitation of Marine Communities. Springer, Berlin, pp. 191–207.
Paine, R.T., 1994. In: Marine Rocky Shores and Community Ecology: An Experimentalist’s Perspective. Excellence in Ecology, Vol. 4. Ecology Institute, Germany, p. 152.
Paine, R.T., Vadas, R.L., 1969. The effects of grazing by sea urchins, Strongylocentrotus spp. on benthic algal populations. Limnol. Oceanogr. 14, 710–719.
Parma, A.M., Deriso, R.B., 1990. Experimental harvesting of cyclic stocks in the face of alternative recruitment hypotheses. Can. J. Fish. Aquat. Sci. 47, 595–610.
Pauly, D., Christensen, V., 1995. Primary production required to sustain global fisheries. Nature 374, 255–257. Polis, G.A., 1999. Why are parts of the world green? Multiple factors control productivity and the distribution
of biomass. Oikos 86, 3–15.
Power, M.E., Tilman, D., Estes, J., Menge, B.A., Bond, W.J., Mills, L.S., Daily, G., Castilla, J.C., Lubchenco, J., Paine, R.T., 1996. Challenges in the quest for keystones. BioScience 46, 609–620.
Prince, J., Walters, C., Ruiz-Avila, R., Sluczanowski, P., 1998. Territorial user’s rights and the Australian abalone (Haliotis sp.) fishery. In: Jamieson, G.S., Campbell, A. (Eds.), Proceedings of the North Pacific Symposium on Invertebrate Stock Assessment and Management. Can. Spec. Publ. Fish. Aquat. Sci, Vol. 125, pp. 367–384.
Roberts, C.M., 1997. Connectivity and management of Caribbean coral reefs. Science 278, 35–38. Roberts, C.M., Hawkins, J.P., 1999. Extinction risk in the sea. TREE 14, 241–246.
Sanders, H.L., 1968. Marine benthic diversity: a comparative study. Am. Nat. 102, 243–282.
Santelices, B., Castilla, J.C., Cancino, J., Schmiede, P., 1980. Comparative ecology of Lessonia nigrescens and
Durvillaea antarctica (Pheophyta) in central Chile. Mar. Biol. 59, 119–132.
Sherman, K., Alexander, L.M., Gold, B.D., 1993. Large Marine Ecosystems: Stress, Mitigation and Sustainability. AAAS Press, Washington, DC.
Sissenwine, M.P., Dann, N., 1991. An overview of multispecies models relevant to the management of living marine resources. Int. Counc. Explor. Sea Mar. Sci. Symp. 193, 6–11.
Sousa, W.P., 1979a. Experimental investigations of disturbance and ecological succession in a rocky intertidal algal community. Ecol. Monogr. 49, 227–254.
Sousa, W.P., 1979b. Disturbance in marine intertidal boulder fields: the nonequilibrium maintenance of species diversity. Ecology 60, 1225–1239.
Sousa, W.P., 1984. The role of disturbance in natural communities. Rev. Ecol. Syst. 15, 353–391.
Stephenson, T.A., Stephenson, A., 1949. The universal features of zonation between tide marks on rocky coasts. J. Ecol. 37, 289–305.
Stephenson, T.A., Stephenson, A., 1972. Life Between Tidemarks On Rocky Shores. W.H. Freeman, San Francisco.
Underwood, A.J., 1978. An experimental evaluation of competition between three species of intertidal prosobranch gastropods. Oecologia 33, 191–201.
Underwood, A.J., 1984. Vertical and seasonal patterns in competition for microalgae between intertidal gastropods. Oecologia (Berl.) 64, 211–222.
Underwood, A.J., 1985. Physical factors and biological interactions: the necessity and nature of ecological experiments. In: Moore, P.G., Seed, R. (Eds.), The Ecology of Rocky Coasts. Hodder & Stoughton, London, pp. 371–390.
Underwood, A.J., 1986. What is a community. In: Raup, D.M., Jablonski, D. (Eds.), Patterns and Processes in the History of Life. Springer, Berlin, pp. 351–367.
Underwood, A.J., 1990. Experiments in ecology and management: their logics, functions and interpretations. Aust. J. Ecol. 15, 365–389.
Underwood, A.J., 1991. The logic of ecological experiments: a case history from studies of the distribution of macro-algae on rocky intertidal shores. J. Mar. Biol. Assoc. UK 71, 841–866.
Underwood, A.J. (Ed.), 1997. Experiments in Ecology. Cambridge University Press, Cambridge, p. 504. Walters, C.J., 1986. Adaptive Management of Renewable Resources. McMillan, New York.