251 (2000) 141–160
www.elsevier.nl / locate / jembe
Chemical warfare among scleractinians: bioactive natural
products from Tubastraea faulkneri Wells kill larvae of
potential competitors
a,b ,* b
Esther G.L. Koh , Hugh Sweatman
a
Department of Marine Biology, James Cook University, Townsville, Queensland 4811, Australia
b
Australian Institute of Marine Science, PMB No. 3, Townsville M.C., Queensland 4810, Australia Received 5 August 1999; accepted 18 February 2000
Abstract
Competition for space among scleractinians by overgrowth, overtopping, extracoelenteric digestion and the use of sweeper tentacles is well recognized, but another potential mode of competitive interaction, allelopathy, is largely uninvestigated. In this study, chemical extracts from
Tubastraea faulkneri Wells were tested for deleterious effects on competent larvae of 11 other
species of coral belonging to seven genera of four scleractinian families. Larvae exposed to extract
21
concentrations from 10 to 500 mg ml consistently suffered higher mortality than larvae in solvent controls. Larvae of Platygyra daedalea (Ellis and Solander) and Oxypora lacera (Verrill) were the most sensitive, experiencing high mortality even at the lowest extract concentration. The toxic compounds from T. faulkneri did not kill any conspecific larvae. The estimated con-centrations of active compounds within T. faulkneri tissues were 100–5000 times higher than the experimental concentrations. Pure compounds isolated from bioactive fractions of the extract were indole alkaloids identified as aplysinopsin, 6-bromoaplysinopsin, 6-bromo-29 -de-N-methyla-plysinopsin and its dimer. The first three occur in other non-zooxanthellate corals in the same family as T. faulkneri, whereas the dimer is novel. These compounds could act as allelochemicals that prevent potential competitors from recruiting in the vicinity of T. faulkneri colonies and help to pre-empt interactions with competitively dominant species. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Allelopathy; Antimicrobial; Aplysinopsin; Bioactive metabolites; Scleractinian coral larvae; Tubastraea faulkneri
*Corresponding author. Present address: Institute of Molecular and Cell Biology, 30 Medical Drive, Singapore 117609, Singapore. Tel.: 165-874-7820 / 6872; fax: 65-7791-117.
E-mail address: esther-g-l [email protected] (E.G.L. Koh).]
1. Introduction
Competition among sessile organisms for space on coral reefs can be intense. Competitive interactions are frequently manifested as overgrowth of adjacent individuals and epibiosis. Apart from the fundamental requirement for space in which to grow, sessile marine invertebrates depend on unobstructed water flow to obtain food. In addition to being filter-feeders, some also possess symbiotic algae that require access to sunlight. Hence it is necessary for these organisms to avoid encroachment and overgrowth by neighbors. Organisms that have slow growth rates will be overgrown unless they have some mechanisms for resisting invasion by faster growing organisms. These alternative means of competing for space include the use of spines (Stebbing, 1973; Harvell, 1998), sweeper tentacles, mesenterial filaments (Lang and Chornesky, 1990) and allelopathy (Sullivan et al., 1983). Allelopathy has been studied in sponges (Thompson, 1985; Thompson et al., 1985; Nishiyama and Bakus, 1999) and soft corals (Coll, 1992; Maida et al., 1995a,b) but has received little attention in scleractinian corals.
Compounds involved in allelopathic interactions range in their action from repellents to toxins and vary in the distances over which they act. Some allelochemicals are soluble or volatile so as to diffuse away from the source (Hadfield and Scheuer, 1985; Slattery et al., 1997). Others may be deposited on the surface of the organism and act by direct contact with potential competitors (Schmitt et al., 1995). Dispersion of allelochemicals provides the obvious advantages of increasing the range of influence but these advantages may be limited by additional costs involved in maintaining active con-centrations of molecules. If the molecules can bind to surrounding substrates and not be easily advected away, less would need to be secreted over time. In the marine environment water-soluble compounds diffuse readily whereas compounds which are insoluble in water would have a longer residence time (Woodin, 1993). Allelochemicals allow the producer to expand or prevent neighboring organisms from encroaching on the space it occupies. For example, tropical soft corals and sponges have been shown to release allelochemicals that kill tissue of adjacent scleractinian corals (Coll et al., 1982; Sammarco et al., 1983; Sullivan et al., 1983; Porter and Targett, 1988; Nishiyama and Bakus, 1999).
ability of scleractinian corals to inhibit the settlement of invertebrate larvae is only starting to be investigated.
The soft corals Sinularia flexibilis and Sarcophyton glaucum secrete toxic secondary metabolites into the water (Coll et al., 1982). Settlement of scleractinian larvae around these two species were inhibited, suggesting an allelopathic function for the secreted compounds (Maida et al., 1995a,b). Bioactive compounds have been detected in scleractinians (Fusetani et al., 1986; Rashid et al., 1995; Koh, 1997), some of which could function as allelochemicals. In a study of the antimicrobial activity of extracts from 100 scleractinian species, Tubastraea faulkneri inhibited the most species of microbes (Koh, 1997). Tubastraea faulkneri may also produce compounds that are toxic to coral competitors. T. faulkneri is a non-zooxanthellate coral of the family De-ndrophylliidae and it inhabits overhangs and vertical surfaces. It has a reported depth range of 3–7 m (Wells, 1982), but it occurs from shallow reef flats down to at least 15 m (personal observations). This slow-growing species shares the shallower parts of its range with other faster growing zooxanthellate species of corals, and therefore is likely to experience interspecific competition. The use of allelochemicals that inhibit settlement and growth of coral competitors would aid T. faulkneri in persisting among the faster growing species.
The present study investigated the hypothesis that a coral can produce chemicals that are toxic to the larvae of other coral species that are potential competitors. We examined the effects of natural products from Tubastraea faulkneri on its own larvae and the larvae of 11 other sympatric coral species. The specific aims of this study were: (a) to determine if natural metabolites produced by T. faulkneri are deleterious to larvae of other scleractinian coral species, (b) to evaluate the likelihood of larvae encountering these compounds at relevant concentrations in their natural environment, and (c) to characterize the bioactive compounds in T. faulkneri.
2. Materials and methods
2.1. Extract preparation
Tubastraea faulkneri colonies were collected from Davies Reef (188509S, 1478409E), Great Barrier Reef, Australia. Specimens were frozen immediately for storage and transportation. In a pilot study, crude extracts of T. faulkneri were prepared from five samples taken from separate coral colonies. Absolute ethanol was used for extraction because it is a good solvent and is miscible with water that was present in the frozen specimens. The extracts were tested on the planulae of Acropora formosa (Dana) and Platygyra sinensis (Edwards and Haime). This showed that natural products of T. faulkneri were toxic to these two species of coral larvae, so more specimens of T. faulkneri were collected to prepare a batch extract for testing on coral larvae from a wider range of species.
faulkneri into three coarse fractions according to their polarities, from the non-polar dichloromethane extract to the highly polar distilled water extract. Each extract was dried under vacuum and weighed. To determine the weight of coral tissue from which the metabolites were extracted, the remaining coral was bleached to remove the tissue, rinsed carefully with distilled water and freeze-dried completely before weighing again to obtain the skeleton weight. The weight of coral tissue (calculated by subtracting skeleton weight from weight of whole coral) was then used to determine natural extract concentrations within the coral tissue.
Antimicrobial activity was used as a representative measure of bioactivity in the dichloromethane, methanol and distilled water extracts of Tubastraea faulkneri. The three extracts and subsequent fractions were screened for activity against a broad range of microbes used in a previous large-scale survey of scleractinian bioactivity (Koh, 1997). These microbes were Vibrio alginolyticus (Miyamoto et al.), V. harveyi (Johnson and Shunk), V. parahaemolyticus (Fujino et al.), Photobacterium damsela (Love et al.), Alteromonas rubra Gauthier, Synechococcus sp. and Staphylococcus aureus Rosenbach. The first six species were chosen because they were isolated from Australian waters and are therefore likely to be encountered by T. faulkneri. S. aureus is terrestrial and served as an indicator for broader spectrum bioactivity. A. rubra was isolated from a coral (Streiner, 1990); P. damsela and V. harveyi are pathogenic to some marine organisms (Fouz et al., 1992; Sutton and Garrick, 1993). V. alginolyticus and V. parahaemolyticus occur in bacterial films on marine substrates (Kaneko and Colwell, 1975; Belas and Colwell, 1982). Synechococcus sp. is a cyanobacterium common in seawater and marine sediments (Glover, 1985). The tests indicated that activity was present almost exclusive-ly in the methanol extract (Table 1). Because toxicity to coral larvae can onexclusive-ly be tested during a brief period each year when planulae are available, priority was given to testing the methanol extract.
2.2. Rearing competent coral planulae
Eleven species of scleractinians were selected, representing a range of habitat preferences and competitive abilities. The species belong to four families: Acroporidae: Montipora digitata (Dana), Acropora formosa, A. millepora (Ehrenberg), A. pulchra
Table 1
a
Antimicrobial bioassay of crude extracts from Tubastraea faulkneri tested at 500mg per disc Bioassay organism Inhibition zone of extracts (mm)
Dichloromethane Methanol Distilled water
Vibrio alginolyticus 0 3.0 0
Vibrio harveyi 0 2.8 0
Vibrio parahaemolyticus 0 0 0
Photobacterium damsela 0 3.8 0
Alteromonas rubra 0 5.0 0
Staphylococcus aureus 0 2.8 0
Synechococcus sp. 1.5 13.0 0
a
(Brook), A. tenuis (Dana), Faviidae: Favia pallida (Dana), Goniastrea aspera Verrill, Platygyra daedalea, P. sinensis, Fungiidae: Fungia fungites (Linnaeus) and Pectiniidae: Oxypora lacera. Only competent larvae were used because they are more likely to encounter active concentrations of allelochemicals from established colonies during the process of settling on a reef. Pre-competent larvae or embryos are assumed to inhabit the water column away from adult corals.
The larvae were obtained from corals at Magnetic Island (19889S, 1468499E) and Orpheus Island (188329S, 1468289E) during three mass spawnings using methods developed by Babcock and Heyward (1986). Gravid colonies of broadcast spawning species were collected during the day and kept in large containers of seawater. When gametes were released at night, they were collected and mixed with those from other colonies of the same species. The resulting larvae were incubated at their natal reefs in floating plastic containers fitted with plankton mesh lids to ensure close approximation to natural growth conditions. Larvae of most broadcast spawning corals of the Great Barrier Reef reach competence when they are 4–7 days old (Babcock and Heyward, 1986). The larvae used in the toxicity assays were 4–5 days old.
To determine the effects of Tubastraea faulkneri extracts on conspecific larvae, adult colonies were collected 1 week after the main spawning event and maintained in flow-through aquaria. The brooded larvae were released about 2 weeks after the main spawning event and were competent within a day, settling readily in the aquaria. Day-old larvae were used for the assays.
2.3. Toxicity bioassay
The toxicity of Tubastraea faulkneri extract to scleractinian planulae was determined for a range of concentrations that were 100 or more times below the concentration in T. faulkneri tissues. The highest test concentration was made up by dissolving a known weight of the T. faulkneri extract in a small volume of 80% ethanol and adding the appropriate volume of sea water. Ethanol formed only 1.6% of this solution. Lower extract concentrations were obtained by serial dilution with seawater, minimizing the error that could arise when pipetting small aliquots of a volatile liquid.
In the pilot experiment, ethanolic extracts from five separate Tubastraea faulkneri colonies were tested on Platygyra sinensis and Acropora formosa planulae at five
21
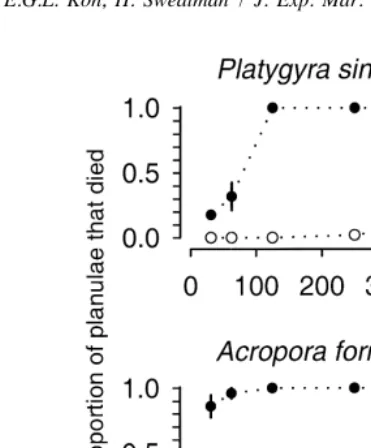
Fig. 1. Toxic effects of Tubastraea faulkneri extracts from five separate colonies on the planulae of Platygyra sinensis and Acropora formosa. Error bars are61 sample standard error [solid symbols5proportion planulae dead after 4 h in T. faulkneri extract; open symbols5proportion planulae dead after 4 h in solvent controls].
only 5%. The pilot study showed that coral planulae were adversely affected by the extract in comparison to the solvent (Fig. 1, Table 2).
A bulk batch of extract was prepared for further tests using a method that allowed the
Table 2
Pilot experiment: toxicity of Tubastraea faulkneri extract to planulae of Platygyra sinensis and Acropora
a
formosa
Planula species Source df S.S. M.S. F P
Platygyra sinensis Treatment 1 5.4 5.4 205.7 ,0.001*
Concentration 4 2.1 0.5 20.4 ,0.001*
Treat.3Conc. 4 1.5 0.4 14.1 ,0.001*
Trend: Linear 1 0.4 0.4 17.1 0.000*
Quadratic 1 0.8 0.8 30.0 0.000*
Other 2 0.2 0.1 4.7 0.015*
Error 40 1.0 0.0
Acropora formosa Treatment 1 10.3 10.3 1422.1 ,0.001*
Concentration 4 0.1 0.0 1.6 0.201
Treat.3Conc. 4 0.0 0.0 1.4 0.261
Error 40 0.3 0.0
a
natural concentration to be calculated (see Section 2.1). The number of experimental extract concentrations was reduced to allow more species of coral larvae to be tested. Based on the pilot study, the other ten species of planulae were tested at three extract
21
concentrations of 50, 100 and 200 mg ml seawater. In addition, the bioassays using Oxypora lacera and Platygyra daedalea were repeated at a later date with the addition
21
of four lower extract concentrations at 10, 20, 30 and 40mg ml seawater.
The results for planula of each species were analyzed using two-way analysis of variance. The two main effects were ‘Treatment’ (ethanol with or without Tubastraea faulkneri extract) and ‘Concentration’. The equality of variances associated with main effects and interactions was tested using the Brown–Forsythe test (Brown and Forsythe, 1974; Keppel, 1991). If variances are heterogeneous, the probability of type I error is increased (Keppel, 1991; Underwood, 1997) so marginally significant differences are suspect. We guarded against this by reanalyzing such cases using a randomization procedure. This approach is based on the planulae being assigned to treatments at random (Manly, 1991). If the null hypothesis is true, then the treatments should have no effect. The pattern of mortality we observed would then represent one random arrangement of the groups of planulae (showing inherently variable mortality rates) among the treatment combinations. The probability of obtaining this result by chance can be estimated as follows. The observed mortalities can be randomly reassigned to treatment groups and a relevant statistic reflecting the difference in mortality among groups is calculated for each rearrangement. Multiple randomizations are used to generate a frequency distribution for the statistic. The likelihood of obtaining the observed value of the statistic (or a more extreme one) by chance can then be estimated. We used the software ‘RT’ (Centre for Applications of Statistics and Mathematics, University of Otago, New Zealand); the test statistic was the percentage of the total sum of squares associated with each factor. In each case, the test distribution was conservatively based on 4999 randomizations plus the original sample (Manly, 1991).
A significant interaction term would indicate that the mortality rates for planulae with and without extract diverged with changes in concentration. All interaction terms were tested for linear and higher order trends with polynomial interaction contrasts for unequally spaced concentrations (Keppel, 1991). If there was no significant interaction, then a significant effect of ‘Treatment’ would indicate that mortality rates for planulae with and without extract were consistently different. In the absence of an interaction, a significant effect of ‘Concentration’ would indicate that the solvent had an effect on larval mortality. The main effect of ‘Concentration’ was also tested for linear and quadratic trends using polynomial contrasts for unequally spaced concentrations.
2.4. Isolation of pure compounds
The active components of the methanol extract were isolated and purified by a combination of chromatographic techniques directed by results from antimicrobial assays. The antimicrobial assays were described in detail in an earlier study (Koh, 1997). Antimicrobial activity was used as a general indicator of bioactivity in fractions obtained at each step of the isolation process.
chromatography on silica gel (see Coll and Bowden, 1986). Briefly, the extract was dissolved in a 1:1 mixture of methanol and dichloromethane. A small amount of silica gel was added to the solution and the solvents were removed using a rotary-evaporator. The dry, even mixture of the extract and silica gel was then loaded onto a silica gel column for fractionation. Solvent mixtures were used to elute six fractions under vacuum, starting at 5% and finishing with 25% methanol in dichloromethane. The flash column fraction with the highest antimicrobial activity was prepared for HPLC by loading it onto a C18 Sep Pak (Waters Millipore Corporation) and eluting it with solvents of decreasing polarity. The first fraction was eluted with 4:6 methanol / distilled water, the second with methanol, the third with 1:1 methanol / ethyl acetate, the fourth with ethyl acetate and the last with hexane. The first two fractions contained antimicrobial activity and were combined for reversed-phase HPLC (no antimicrobial activity was detected in the third and fourth fractions and nothing was eluted with hexane).
Reverse-phase HPLC was carried out on an Alltech Econosil C18 column (10 mm; 250 mm long and 10 mm internal diameter) with an isocratic solvent regime (nine parts methanol and one part 5% ammonium acetate in distilled water) at a flow-rate of 1.5 ml
21
min . Four major HPLC fractions were collected at the following retention times: 8, 9.5, 10.5 and 13 min. Ammonium acetate was removed from the fractions before they were tested for antimicrobial activity. The fractions were further purified under the same HPLC conditions before they were characterized by melting point analysis, NMR, UV and mass spectrometry.
3. Results
3.1. Natural concentration of Tubastraea faulkneri extracts
Tubastraea faulkneri specimens (15–20 colonies of varying sizes) with a total wet weight of 247.47 g weighed 152.94 g after freeze-drying. Freeze-dried dichloromethane, methanol and distilled water extracts weighed 0.51, 5.40, and 2.23 g, respectively. Subtracting the weight of the dried skeleton alone gave a wet weight for tissue of 108.63 g (14.1 g dry weight). Therefore the natural concentration of the methanol extract was
21 21
49.7 mg g wet tissue or 383.0 mg g dry tissue. The natural concentration of the
21 21
distilled water extract was 20.5 mg g wet tissue or 158.2 mg g dry tissue and that of
21 21
the dichloromethane extract was 4.7 mg g wet tissue or 36.2 mg g dry tissue.
3.2. Bioactivity of extracts isolated from Tubastraea faulkneri
Synechococcus sp., but much less intensely than the methanol extract. No inhibitory activity was recorded for the distilled water extract.
3.3. Toxicity of Tubastraea faulkneri extract to scleractinian larvae
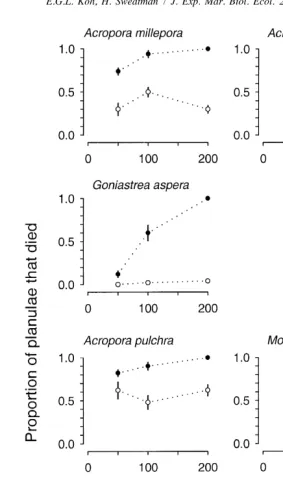
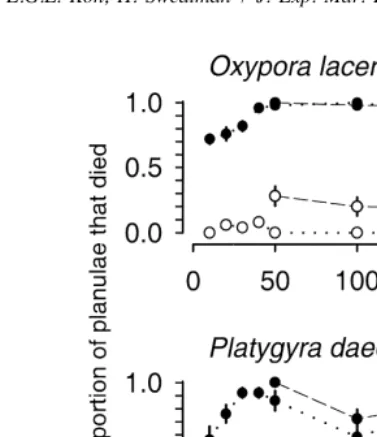
Tubastraea faulkneri larvae alone among the 12 test species showed no visible toxic effects from exposure to the extract of T. faulkneri. No conspecific larvae died in any of the bioassays. In contrast, the larvae of all 11 other species suffered higher mortality at all concentrations of T. faulkneri extract when compared with solvent controls (Figs. 1–3). Mortality of planulae after a 4-h exposure to solvent controls ranged from 0% in four species (Platygyra sinensis, P. daedalea, Goniastrea aspera, Oxypora lacera) to 72% in Montipora digitata.
There was significant interaction between treatments (presence or absence of extract) and concentrations for six planula species (Acropora millepora, A. tenuis, Goniastrea aspera, Platygyra sinensis, P. daedalea, Oxypora lacera; Tables 2–4), meaning that the effect of increasing concentration differed between larvae treated with solvent plus Tubastraea faulkneri extract and those treated with solvent alone. Acropora millepora and A. tenuis showed only linear interaction trends implying that response curves for planulae treated with extract diverged in a consistent manner from response curves for those treated only with solvent (Fig. 2, Table 3). For P. sinensis, G. aspera and O. lacera, the response curves for the treatments diverged, but not in a linear fashion (Figs. 1–3, Tables 2–4). In the case of P. sinensis, the nonlinearity was due to total mortality occurring at relatively low concentrations of extract, so while the mortality increased with increasing concentration of solvent alone, the mortality of larvae treated with T. faulkneri extract could not increase above 100%. For G. aspera and O. lacera, the nonlinearity resulted from the sharp rise in mortality with increasing concentrations of extract while mortality remained fairly low and constant as the concentration of solvent alone increased. Platygyra daedalea shows a cubic trend though the consistent decline in
21
mortality at 100 mg ml has no simple explanation (Fig. 3, Table 4). There were no significant trends in interactions in any of the species with non-significant overall interaction terms.
Acropora formosa, A. pulchra, Montipora digitata, Fungia fungites, Favia pallida and Oxypora lacera (at high concentrations) all had significantly higher mortality in the presence of Tubastraea faulkneri extract (Tables 2–4, Figs. 1–3). Included in this group are the only two species, M. digitata and O. lacera (tested at high concentrations) showing unequal variances among treatment groups; these are associated with main effect ‘Treatment’. The inequality comes about because all the larvae died when exposed to extract of T. faulkneri at experimental concentrations (Figs. 2, 3). As a result, these treatment groups had zero variance in mortality while control groups showed less mortality and some variance. The differences in mortality are highly significant by both ANOVA and the randomization tests (Tables 3, 4).
Fig. 3. Toxic effects of Tubastraea faulkneri extract on planulae of Oxypora lacera and Platygyra daedalea. The bioassays were initially conducted at three extract concentrations and repeated at a later date at seven extract concentrations (see Materials and methods). Error bars are61 sample standard error [solid symbols5 proportion planulae dead after 4 h in T. faulkneri extract; open symbols5proportion planulae dead after 4 h in solvent controls].
experimental concentrations of the solvent were toxic to planulae of these two species, but in each case the presence of Tubastraea faulkneri extract caused significant additional mortality (Fig. 2, Table 3).
3.4. Isolation and purification of active components
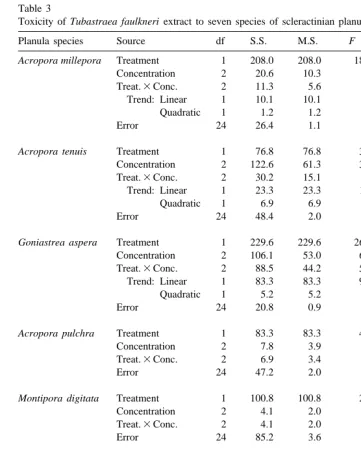
Table 3
a
Toxicity of Tubastraea faulkneri extract to seven species of scleractinian planulae
Planula species Source df S.S. M.S. F P RP
Acropora millepora Treatment 1 208.0 208.0 189.1 ,0.001* n / a
Concentration 2 20.6 10.3 9.4 0.001*
Treat.3Conc. 2 11.3 5.6 5.1 0.014*
Trend: Linear 1 10.1 10.1 9.1 0.006*
Quadratic 1 1.2 1.2 1.1 0.305
Error 24 26.4 1.1
Acropora tenuis Treatment 1 76.8 76.8 38.1 ,0.001* n / a
Concentration 2 122.6 61.3 30.4 ,0.001*
Treat.3Conc. 2 30.2 15.1 7.5 0.003*
Trend: Linear 1 23.3 23.3 11.6 0.002*
Quadratic 1 6.9 6.9 3.4 0.078
Error 24 48.4 2.0
Goniastrea aspera Treatment 1 229.6 229.6 265.0 ,0.001* n / a Concentration 2 106.1 53.0 61.2 ,0.001* Treat.3Conc. 2 88.5 44.2 51.0 ,0.001* Trend: Linear 1 83.3 83.3 95.7 0.000*
Quadratic 1 5.2 5.2 6.0 0.022*
Error 24 20.8 0.9
Acropora pulchra Treatment 1 83.3 83.3 42.4 ,0.001* n / a
Concentration 2 7.8 3.9 2.0 0.160
Treat.3Conc. 2 6.9 3.4 1.8 0.196
Error 24 47.2 2.0
Montipora digitata Treatment 1 100.8 100.8 28.4 ,0.001* ,0.001*
Concentration 2 4.1 2.0 0.6 0.572 0.752
Treat.3Conc. 2 4.1 2.0 0.6 0.572 0.776
Error 24 85.2 3.6
Fungia fungites Treatment 1 240.8 240.8 62.3 ,0.001* n / a
Concentration 2 51.7 25.8 6.7 0.005*
Trend: Linear 1 50.1 50.1 12.9 0.001*
Quadratic 1 1.6 1.6 0.4 0.525
Treat.3Conc. 2 6.1 3.0 0.8 0.468
Error 24 92.8 3.9
Favia pallida Treatment 1 563.3 563.3 1024.2 ,0.001* n / a
Concentration 2 5.1 2.5 4.6 0.020*
Trend: Linear 1 4.6 4.6 8.4 0.008*
Quadratic 1 0.5 0.5 0.8 0.371
Treat.3Conc. 2 1.9 0.9 1.7 0.205
Error 24 13.2 0.6
a
Table 4
a
Toxicity of Tubastraea faulkneri extract to planulae of Oxypora lacera and Platygyra daedalea
Planula species Source df S.S. M.S. F P RP
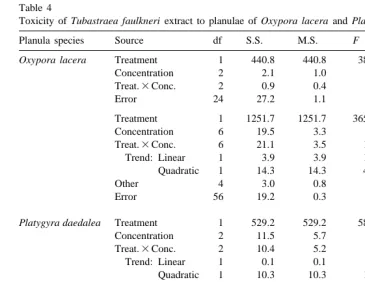
Oxypora lacera Treatment 1 440.8 440.8 389.0 ,0.001* ,0.001*
Concentration 2 2.1 1.0 0.9 0.415 0.933
Treat.3Conc. 2 0.9 0.4 0.4 0.686 0.973
Error 24 27.2 1.1
Treatment 1 1251.7 1251.7 3650.7 ,0.001* n / a
Concentration 6 19.5 3.3 9.5 ,0.001*
Treat.3Conc. 6 21.1 3.5 10.3 ,0.001*
Trend: Linear 1 3.9 3.9 11.3 0.001*
Quadratic 1 14.3 14.3 42.0 ,0.001*
Other 4 3.0 0.8 2.2 0.079
Error 56 19.2 0.3
Platygyra daedalea Treatment 1 529.2 529.2 588.0 ,0.001* n / a
Concentration 2 11.5 5.7 6.4 0.006*
Treat.3Conc. 2 10.4 5.2 5.8 0.009*
Trend: Linear 1 0.1 0.1 0.1 0.760
Quadratic 1 10.3 10.3 11.5 0.002*
Error 24 21.6 0.9
Treatment 1 1010.8 1010.8 879.0 ,0.001* n / a
Concentration 6 51.1 8.5 7.4 ,0.001*
Treat.3Conc. 6 41.6 6.9 6.0 ,0.001*
Trend: Linear 1 0.3 0.3 0.3 0.614
Quadratic 1 2.0 2.0 1.7 0.195
Other 4 39.3 9.8 8.5 0.000*
Error 56 64.4 1.2
a
Factor ‘Treatment’ refers to presence or absence of extract in test solution. Factor ‘Concentration’ refers to concentrations of extract and corresponding solvent controls. The bioassays were repeated at lower extract concentrations (see Materials and methods). * Shows significant P values (,0.05). Variances were homoge-neous among treatment groups for interaction and main effects, except for Oxypora lacera when tested at higher concentrations. RP values are probabilities from randomization tests; these supported the ANOVA results. Trends refer to partitioning of the significant interaction term using polynomial contrasts.
Table 5
Antimicrobial bioassay of HPLC fractions of the methanol-soluble extract from Tubastraea faulkneri, tested at
a
concentrations proportionate to levels found in 500mg of extract Bioassay organism Inhibition zone of fractions (mm)
HPLC 3 HPLC 4 HPLC 5 HPLC 6
21 21 21 21
[96mg disc ] [107mg disc ] [146mg disc ] [11mg disc ]
Vibrio alginolyticus 0 0 0 0
Vibrio harveyi Not tested 0 0 0
Photobacterium damsela 0 0 0 0
Alteromonas rubra 0 0 0 0
Staphylococcus aureus 1.0 0 1.5 0
Synechococcus sp. 4.5 15.0 8.5 6.0
a
Table 6
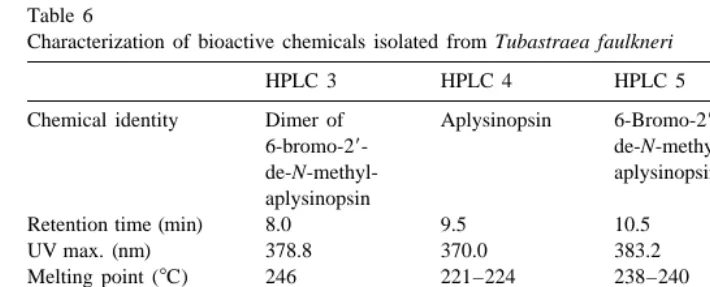
Characterization of bioactive chemicals isolated from Tubastraea faulkneri
HPLC 3 HPLC 4 HPLC 5 HPLC 6
Chemical identity Dimer of Aplysinopsin 6-Bromo-29- 6-Bromo-6-bromo-29- de-N-methyl- aplysinopsin
de-N-methyl- aplysinopsin
aplysinopsin
Retention time (min) 8.0 9.5 10.5 13.0
UV max. (nm) 378.8 370.0 383.2 379.2
Melting point (8C) 246 221–224 238–240 254
Molecular weight 636 254 318 332
Concentration in wet 9.5 10.6 14.5 1.1
21
tissue (mg g )
Concentration in dry 73.5 82.0 111.8 8.4
21
tissue (mg g )
4. Discussion
4.1. Toxicity to larvae of other coral species
This is the first record of extracts from an azooxanthellate scleractinian having toxic effects on the larvae of zooxanthellate scleractinian species. Extracts from Tubastraea faulkneri killed competent larvae of 11 species of corals from seven genera and four families. For the two most sensitive planulae (Oxypora lacera and Platygyra daedalea)
21
the extract was toxic at 10 mg ml (Fig. 3). To compare the experimental con-centrations with the natural tissue concon-centrations, the former can be expressed as mg
21 21
extract g seawater. Since the density of seawater is 1.02 g ml under the experimental conditions of 258C and 1 Atm, 1 ml of seawater would weigh
approximate-21 21
ly 1 g. Therefore 10 mg ml is approximately equivalent to 10mg g seawater. The
21
concentration of extract in T. faulkneri tissue (49.7 mg g wet tissue) was 5000 times
21
higher. Compared to the highest experimental dose of 500 mg ml , the tissue concentration was still about 100 times higher. This suggests that the toxicity we observed was not a result of unrealistically high extract concentrations in the bioassays. Nor was lack of oxygen a problem, for larvae of P. sinensis suffered little mortality when held for a week in solvent control vials. Experimental larvae were held at the same densities for only 4 h.
21
100mg ml is puzzling; there is currently no explanation for the effect. The ability of T. faulkneri to inhibit settlement of coral larvae could be tested in the field by deploying settlement plates around T. faulkneri colonies and in control sites, following Maida et al. (1995a,b). Settlement rates should be reduced in the vicinity of T. faulkneri colonies compared to control plates.
Toxicity to larvae of potential competitors raises the question of whether these larvae actually encounter the compounds in nature at concentrations that are high enough to have an effect. In a similar study on the soft coral Sinularia flexibilis, two bioactive
21
compounds were detected at 1–5 mg ml in the water surrounding a colony of S. flexibilis in the field (Coll et al., 1982). The compounds, flexibilide and
dihydroflex-21
ibilide, were present at approximately 3.5 and 1 mg g dry tissue, respectively (Kazlauskas et al., 1978). Tissue concentrations of the compounds we isolated from Tubastraea faulkneri were up to two orders of magnitude higher than those of flexibilide and dihydroflexibilide in S. flexibilis (see Table 6). If T. faulkneri secretes its bioactive compounds at a similar rate to S. flexibilis, the concentrations in T. faulkneri tissues
21 suggest that those in the surrounding water would be about 8–100 mg ml . This
21
corresponds to about 400mg methanol extract ml water, a concentration which is well above the toxic levels demonstrated in our study (Figs. 1–3). Although we have not shown that larvae are exposed to these levels in nature, our results suggest that secretion of allelochemicals by T. faulkneri is a possibility. This hypothesis could be tested in the field.
The effectiveness of allelopathy will depend in part on the rate of dissipation of allelochemicals from the colony. This loss could be minimized if the compounds were able to bind to the surrounding substrate. The polychaete Thelepus crispus releases halogenated aromatic compounds that inhibit the settlement of another polychaete Nereis vexillosa, and the compounds may have long residence times in the substrate because of their low solubility in water (Woodin, 1993). Three of the pure compounds we isolated from the methanol extract of Tubastraea faulkneri are brominated aromatic compounds that are not soluble in water. The fourth is not halogenated but belongs to the same class of aromatic compounds, and is also not water-soluble. If secreted by the coral, these putative allelochemicals could therefore bind selectively to organic films on surfaces in the vicinity of the coral and not be wafted away. The immobilization of the compounds to the substrate would also localize them in the most relevant place for inhibiting the settlement of competitors.
4.2. Non-toxicity to conspecific larvae
Although the methanol extract of Tubastraea faulkneri killed larvae of all the other coral species, it was not toxic to conspecific larvae. There was no mortality of T.
21
externally). Scleractinian larvae are not universally immune to toxic compounds produced by adults of the same species: extracts from two broadcast spawning species, P. daedalea and Fungia fungites, are toxic to conspecific larvae (Fearon and Cameron, 1997).
The ability of Tubastraea faulkneri larvae to withstand high concentrations of the larvotoxic compounds produced by T. faulkneri adults could provide additional advantages if the compounds indeed serve as allelochemicals. Tubastraea faulkneri larvae would be able to settle near adults without harm and face no competition from other coral recruits. Survival of recruits may also be increased because the presence of adults indicates that conditions have allowed survival for some time (Crisp, 1974; Pawlik, 1992). Larvae of T. faulkneri reportedly settle preferentially in the presence of adult colonies (Wolstenholme, 1991). From the perspective of adult colonies of T. faulkneri, juvenile conspecifics that settle nearby pose no threat since slow growing juveniles are unlikely to become serious competitors. In the longer term, the proximity of conspecifics may increase fertilization success rates.
4.3. Bioactive compounds in scleractinian corals
The number of metabolites identified in scleractinian corals has been increasing slowly but steadily (see reviews by D.J. Faulkner in Natural Product Reports from 1984 to 1999). Three of the compounds that were identified in this study (aplysinopsin, 6-bromoaplysinopsin and 6-bromo-29-de-N-methylaplysinopsin) have previously been isolated from other dendrophylliid corals (Okuda et al., 1982; Fattorusso et al., 1985; Fusetani et al., 1986; Guella et al., 1988, 1989). They are also present in a number of sponges (Kazlauskas et al., 1977; Hollenbeak and Schmitz, 1977; Djura and Faulkner, 1980). The fourth, a dimer of 6-bromo-29-de-N-methylaplysinopsin, has never been reported before. Among the dendrophylliid corals, this group of indole alkaloid compounds has only been found in the azooxanthellate members of the family. Tubastraea faulkneri also contains two cytotoxic compounds that are not related to aplysinopsins–mycalolides C and D (Rashid et al., 1995). These compounds are distinct from the aplysinopsin-like compounds in their polarity (having been extracted in the less polar solvents, chloroform and carbon tetrachloride) and therefore could not have been responsible for the antimicrobial activity of the methanol-soluble compounds reported here.
4.4. Summary
suggest that allelopathy against larvae of competitors may occur in scleractinian species, as has been reported for soft corals and scleractinian larvae (Maida et al., 1995a, b). We postulate that the toxic compounds may enable the slow growing T. faulkneri to persist among other faster growing corals by inhibiting the settlement of their larvae. Tubastraea faulkneri is not known to possess sweeper tentacles and does not appear to use extracoelenteric digestion to prevent encroachment of other corals (for list of corals that employ these defenses see Lang and Chornesky, 1990). Its growth rate (linear extension) is not known but other azooxanthellate members of the genus have growth rates similar to slow growing zooxanthellate corals (Alcala et al., 1981; Gomez et al., 1985). A slower growth rate would mean that it could have difficulty competing against faster growing corals for space to grow and also to prevent itself from being smothered. However, an ability to pre-empt competition by killing larvae of potential competitors could protect T. faulkneri from being out-competed by faster growing zooxanthellate corals in the shallower reef habitats.
Acknowledgements
This work was supported by internal grants to E.G.L. Koh from the Department of Marine Biology, James Cook University and the Australian Institute of Marine Science (AIMS). We thank P. Harrison, C. Mundy and B. Willis for providing planulae of broadcast spawning corals. R. Cocciolone, D. Sutton and H. Streiner kindly provided pure cultures of the microbes used in this study. We thank W. Shipton and R. Hirst for the use of the microbiology facilities at the Department of Biomedical and Tropical Veterinary Science, JCU. Mass spectrometry analyses were performed by J. Macleod, Research School of Chemistry, Australian National University and D. Bourne, AIMS. We are grateful to members of the Marine Bioproducts Unit at AIMS for access to laboratory facilities and for their assistance. E.G.L. Koh is especially grateful to P. Murphy for his invaluable advice in marine natural products chemistry and his unstinting support. We also thank C. Alexander, P. Murphy and J. Feingold for helpful comments on an earlier version of the manuscript. This is contribution number 1018 from AIMS [SS].
References
Aceret, T.L., Sammarco, P.W., Coll, J.C., 1995. Effects of diterpenes derived from the soft coral Sinularia flexibilis on the eggs, sperm and embryos of the scleractinian corals Montipora digitata and Acropora tenuis. Mar. Biol. 122, 317–323.
Alcala, A.C., Alcala, L.C., Gomez, E.D., Cowan, M.E., Yap, H.T., 1981. Growth of certain corals, molluscs and fish in artificial reefs in the Philippines. Proc. 4th Int. Coral Reef Symp. 2, 215–220.
Babcock, R.C., Heyward, A.J., 1986. Larval development of certain gamete-spawning scleractinian corals. Coral Reefs 5, 111–116.
Belas, M.R., Colwell, R.R., 1982. Adsorption kinetics of laterally and polarly flagellated Vibrio. J. Bacteriol. 151, 1568–1580.
Butler, A.J., Van Altena, I.A., Dunne, S.J., 1996. Antifouling activity of lyso-platelet-activating factor extracted from Australian sponge Crella incrustans. J. Chem. Ecol. 22, 2041–2061.
Coll, J.C., 1992. The chemistry and chemical ecology of octocorals (Coelenterata, Anthozoa, Octocorallia). Chem. Rev. 92, 613–631.
Coll, J.C., Bowden, B.F., 1986. The application of vacuum liquid chromatography to the separation of terpene mixtures. J. Nat. Prod. (Lloydia) 49, 934–936.
Coll, J.C., Bowden, B.F., Tapiolas, D.M., Dunlap, W.C., 1982. In situ isolation of allelochemicals released from soft corals (Coelenterata: Octocorallia): a totally submersible sampling apparatus. J. Exp. Mar. Biol. Ecol. 60, 293–299.
Crisp, D.J., 1974. Factors influencing the settlement of marine invertebrate larvae. In: Grant, P.T., Mackie, A.N. (Eds.), Chemoreception in Marine Organisms. Academic Press, New York, pp. 177–265.
Davis, A.R., Wright, A.E., 1990. Inhibition of larval settlement by natural products from the ascidian, Eudistoma olivaceum (Van Name). J. Chem. Ecol. 16, 1349–1357.
Djura, P., Faulkner, D.J., 1980. Metabolites of the marine sponge the Dercitus sp. J. Org. Chem. 45, 737–737. Fattorusso, E., Lanzotti, V., Magno, S., Novellino, E., 1985. Tryptophan derivatives from a Mediterranean
anthozoan, Astroides calycularis. J. Nat. Prod. 48, 924–927.
Faulkner, D.J., 1984. Marine natural products: metabolites of marine invertebrates. Nat. Prod. Rep. 1, 551–598.
Faulkner, D.J., 1986. Marine natural products. Nat. Prod. Rep. 3, 1–33. Faulkner, D.J., 1987. Marine natural products. Nat. Prod. Rep. 4, 539–576. Faulkner, D.J., 1988. Marine natural products. Nat. Prod. Rep. 5, 613–663. Faulkner, D.J., 1990. Marine natural products. Nat. Prod. Rep. 7, 269–309. Faulkner, D.J., 1991. Marine natural products. Nat. Prod. Rep. 8, 97–147. Faulkner, D.J., 1992. Marine natural products. Nat. Prod. Rep. 9, 323–364. Faulkner, D.J., 1993. Marine natural products. Nat. Prod. Rep. 10, 497–539. Faulkner, D.J., 1994. Marine natural products. Nat. Prod. Rep. 11, 355–394. Faulkner, D.J., 1995. Marine natural products. Nat. Prod. Rep. 12, 223–269. Faulkner, D.J., 1996. Marine natural products. Nat. Prod. Rep. 13, 75–125. Faulkner, D.J., 1997. Marine natural products. Nat. Prod. Rep. 14, 259–302. Faulkner, D.J., 1998. Marine natural products. Nat. Prod. Rep. 15, 113–158. Faulkner, D.J., 1999. Marine natural products. Nat. Prod. Rep. 16, 155–198.
Fearon, R.J., Cameron, A.M., 1996. Larvotoxic extracts of the hard coral Goniopora tenuidens: allelochemi-cals that limit settlement of potential competitors? Toxicon 34, 361–367.
Fearon, R.J., Cameron, A.M., 1997. Preliminary evidence supporting the ability of hermatypic corals to affect adversely larvae and early settlement stages of hard coral competitors. J. Chem. Ecol. 23, 1769–1780. Fleck, J., Fitt, W.K., 1999. Degrading mangrove leaves of Rhizophora mangle Linne provide a natural cue for
settlement and metamorphosis of the upside down jellyfish Cassiopea xamachana Bigelow. J. Exp. Mar. Biol. Ecol. 234, 83–94.
Fouz, B., Larsen, J.L., Nielsen, B., Barga, J.L., Toranzo, A.E., 1992. Characterization of Vibrio damsela strains isolated from turbot Scophthalmus maximus in Spain. Dis. Aquat. Org. 12, 155–166.
Fusetani, N., Asano, M., Matsunaga, S., Hashimoto, K., 1986. Bioactive marine metabolites. XV. Isolation of aplysinopsin from the scleractinian coral Tubastrea aurea as an inhibitor of development of fertilized sea urchin eggs. Comp. Biochem. Physiol. 85B, 845–846.
Glover, H.E., 1985. The physiology and ecology of the marine cyanobacterial genus Synechococcus. In: Jannasch, H.W., Williams, P.J.le B. (Eds.). Advances in Aquatic Microbiology, Vol. 3. Academic Press, London, pp. 49–107.
˜
Gomez, E.D., Alcala, A.C., Yap, H.T., Alcala, L.C., Alino, P.M., 1985. Growth studies of commercially important scleractinians. Proc. 5th Int. Coral Reef Symp. 6, 199–204.
Guella, G., Mancini, I., Zibrowius, Z., Pietra, F., 1988. Novel aplysinopsin-type alkaloids from scleractinian corals of the family Dendrophylliidae of the Mediterranean and the Philippines. Configurational-assignment criteria, stereospecific synthesis, and photoisomerization. Helv. Chim. Acta 71, 773–782.
Hadfield, M.G., 1977. Chemical interactions in larval settling of a marine gastropod. In: Faulkner, D.J., Fenical, W.H. (Eds.), Marine Natural Products Chemistry. Plenum Press, New York, pp. 403–413. Hadfield, M.G., 1978. Metamorphosis in marine molluscan larvae: an analysis of stimulus and response. In:
Chia, F.-S., Rice, M.E. (Eds.), Settlement and Metamorphosis of Marine Invertebrate Larvae. Elsevier, New York, pp. 165–175.
Hadfield, M.G., 1984. Settlement requirements of molluscan larvae: new data on chemical and genetic roles. Aquaculture 39, 283–298.
Hadfield, M.G., Pennington, J.T., 1990. Nature of the metamorphic signal and its internal transduction in larvae of the nudibranch Phestilla sibogae. Bull. Mar. Sci. 46, 455–464.
Hadfield, M.G., Scheuer, D., 1985. Evidence for a soluble metamorphic inducer in Phestilla sibogae: ecological, chemical and biological data. Bull. Mar. Sci. 37, 556–566.
Harvell, C.D., 1998. Genetic variation and polymorphism in the inducible spines of a marine bryozoan. Evolution 52, 80–86.
Hollenbeak, K.H., Schmitz, F.J., 1977. Aplysinopsin: antineoplastic tryptophan derivative from the marine sponge Verongia spengelii. Lloydia 40, 479–481.
Kaneko, T., Colwell, R.R., 1975. Adsorption of Vibrio parahaemolyticus onto chitin and copepods. Appl. Microbiol. 29, 269–274.
Kato, T., Kumanireng, A.S., Ichinose, I., Kitihara, Y., Kakinuma, Y., Nishihira, M., Kato, M., 1975. Active components of Sargassum tortile effecting settlement of swimming larvae of Coryne uchidai. Experientia 31, 433–434.
Kazlauskas, R., Murphy, P.T., Quinn, R.J., Wells, R.J., 1977. Aplysinopsin, a new tryptophan derivative from a sponge. Tetrahedron Lett. 1, 61–64.
¨
Kazlauskas, R., Murphy, P.T., Wells, R.J., Schonholzer, P., Coll, J.C., 1978. Cembranoid constituents from an Australian collection of the soft coral Sinularia flexibilis. Aust. J. Chem. 31, 1817–1824.
Keppel, G., 1991. Design and Analysis: A Researcher’s Handbook, 3rd Edition. Prentice-Hall, Englewood Cliffs, NJ.
Koh, E.G.L., 1997. Do scleractinian corals engage in chemical warfare against marine microbes? J. Chem. Ecol. 23, 379–398.
Lang, J.C., Chornesky, E.A., 1990. Competition between scleractinian reef corals – a review of mechanisms and effects. In: Dubinsky, Z. (Ed.). Ecosystems of the World, Vol. 25. Elsevier, Amsterdam, pp. 209–252. Maida, M., Sammarco, P.W., Coll, J.C., 1995a. Preliminary evidence for directional allelopathic effects of the soft coral Sinularia flexibilis (Alcyonacea: Octocorallia) on scleractinian coral recruitment. Bull. Mar. Sci. 56, 303–311.
Maida, M., Sammarco, P.W., Coll, J.C., 1995b. Effects of soft corals on scleractinian coral recruitment. I. Directional allelopathy and inhibition of settlement. Mar. Ecol. Prog. Ser. 121, 191–202.
Manly, B.F.J., 1991. Randomization and Monte Carlo Methods in Biology. Chapman and Hall, London. Nishiyama, G.K., Bakus, G.J., 1999. Release of allelochemicals by three tropical sponges (Demospongiae) and
their toxic effects on coral substrate competitors. Mem. Queensland Museum 44, 411–417.
Okuda, K.K., Klein, D., Kinnel, R.B., Li, M., Scheuer, P.J., 1982. Marine natural products: the past twenty years and beyond. Pure Appl. Chem. 54, 1907–1914.
Pawlik, J.R., 1992. Chemical ecology of the settlement of benthic marine invertebrates. Oceanogr. Mar. Biol. Annu. Rev. 30, 273–335.
Porter, J.W., Targett, N.M., 1988. Allelochemical interactions between sponges and corals. Biol. Bull. 175, 230–239.
Rashid, M.A., Gustafson, K.R., Cardellina, II J.H., Boyd, M.R., 1995. Mycalolides D and E, new cytotoxic macrolides from a collection of the stony coral Tubastrea faulkneri. J. Nat. Prod. 58, 1120–1125. Rodriguez, S.R., Ojeda, F.P., Inestrosa, N.C., 1993. Settlement of benthic marine invertebrates. Mar. Ecol.
Prog. Ser. 97, 193–207.
Sammarco, P.W., Coll, J.C., La Barre, S., Willis, B., 1983. Competitive strategies of soft corals (Coelenterata: Octocorallia): allelopathic effects on selected scleractinian corals. Coral Reefs 1, 173–178.
Schmitt, T.M., Hay, M.E., Lindquist, N., 1995. Constraints on chemically mediated coevolution: multiple functions for seaweed secondary metabolites. Ecology 76, 107–123.
Stebbing, A.R.D., 1973. Observations on colony overgrowth and spatial competition. In: Larwood, G.P. (Ed.), Living and Fossil Bryozoa. Academic Press, London, pp. 173–183.
Streiner, H.M. (Ed.), 1990. The Australian collection of marine microorganisms. Catalogue of strains. Sir George Fisher Centre for Tropical Marine Studies, James Cook University of North Queensland, Australia. Sullivan, B., Faulkner, D.J., Webb, L., 1983. Siphonodictidine, a metabolite of the burrowing sponge
Siphonodictyon sp. that inhibits coral growth. Science 221, 1175–1176.
Sutton, D.C., Garrick, R., 1993. Bacterial disease of cultured giant clam Tridacna gigas larvae. Dis. Aquat. Org. 16, 47–53.
Thompson, J.E., 1985. Exudation of biologically active metabolites in the sponge Aplysina fistularis. I. Biological evidence. Mar. Biol. 88, 23–26.
Thompson, J.E., Walker, R.P., Faulkner, D.J., 1985. Screening and bioassays for biologically-active substances from 40 marine sponge species from San Diego, California, USA. Mar. Biol. 88, 11–21.
Underwood, A.J., 1997. Experiments in Ecology. Their Logical Design and Interpretation Using Analysis of Variance. Cambridge University Press, London.
Walters, L.J., Hadfield, M.G., Smith, C.M., 1996. Waterborne chemical compounds in tropical macroalgae: positive and negative cues for larval settlement. Mar. Biol. 126, 383–393.
´
Wells, J.W., 1982. Notes on Indo-Pacific scleractinian corals. Part 9. New corals from the Galapagos Islands. Pac. Sci. 36, 211–219.
Wolstenholme, J.K., 1991. An investigation of the reproductive biology and early life history of the scleractinian coral Tubastraea faulkneri (Wells 1982). Honors thesis, James Cook University of North Queensland, Townsville, Australia.
Woodin, S.A., 1993. Allelochemical inhibition of recruitment in a sedimentary assemblage. J. Chem. Ecol. 19, 517–530.