TINJAUAN PUSTAKA
Limbah Udang
Limbah udang adalah hasil samping yang dibuang pada industri pengolahan udang beku (Arlius 1991). Hasil samping tersebut, berupa kepala, kulit keras (carapace), dan ekor (uropod) udang (Setyahadi 2006).
Kualitas limbah udang terutama ditinjau dari kandungan nutrien dan komposisi kimianya, cukup baik dan layak dijadikan sebagai sumber protein dalam ransum ternak. Hasil analisa di laboratorium Ilmu dan Teknologi Pakan IPB, memperlihatkan bahwa protein kasarnya cukup tinggi yaitu 41.58%, hampir sama dengan bungkil kedelai (45.6%). Begitu juga bahan keringnya (88.32% vs 88.0%). Akan tetapi ada perbedaan pada serat kasarnya yaitu 13.72% dalam limbah udang, sedangkan bungkil kedelai 4.58%, sehingga menjadi faktor pembatas karena kecernaannya yang rendah. Walaupun demikian, serat kasarnya (kitin) bisa berguna dalam menurunkan kolesterol daging. Oleh sebab itu, pemanfaatan limbah udang sebagai pakan ternak sebaiknya dilakukan proses pengolahan terlebih dahulu. Perbandingan komposisi kimia antara tepung limbah udang dengan tepung ikan dan bungkil kedelai dapat dilihat pada Tabel 1.
Tabel 1 Komposisi kimia tepung limbah udang, tepung ikan, dan bungkil kedelai
Nutrien Tepung limbah udang1) Tepung ikan2) Bungkil kedelai3) (% bahan kering)
---Protein kasar 41.58 52.6 45.60
Serat kasar 13.72 2.2 4.58
Lemak kasar 3.08 6.8 2.79
Abu 22.06 20.7 6.84
1)Hasil analisa di laboratorium Ilmu dan Teknologi Pakan IPB
2)Hartadi et al. (1997) 3)
Sutardi (2001)
Kualitas protein limbah udang sangat bagus karena mengandung semua asam amino esensial. Asam amino metionin yang sering menjadi faktor pembatas pada protein nabati, kandungannya lebih tinggi bila dibandingkan dengan bungkil kedelai dan hampir sama dengan tepung ikan bahkan mikroba rumen.
Perbandingan komposisi asam amino antara tepung limbah udang dengan tepung ikan, bungkil kedelai, dan mikroba rumen disajikan pada Tabel 2.
Tabel 2 Komposisi asam amino tepung limbah udang, tepung ikan, bungkil kedelai, dan mikroba rumen
Asam amino
Tepung limbah udang
Tepung ikan3) Bungkil kedelai4) Mikroba rumen5) Udang merah jambu
(Pandalus borealis)1)
Udang windu (Penaeus monodon)2)
--- (gram/100 gram protein)
---Alanin 5.25 ± 0.05 2.14 - 4.6 7.5 Arginin 6.13 ± 0.07 4.67 6.46 7.0 5.1 Asam Aspartat 11.17 ± 0.01 7.52 - 10.9 12.2 Asam Glutamat 12.85 ± 0.14 11.36 - 14.3 13.1 Fenilalanin 5.13 ± 0.07 5.52 4.64 3.9 5.1 Glisin 4.11 ± 0.03 17.76 7.70 3.5 5.8 Histidin 2.24 ± 0.09 2.35 2.78 1.8 2.0 Isoleusin 5.78 ± 0.13 4.16 4.30 2.1 5.7 Leusin 7.01 ± 0.02 8.65 7.20 7.3 8.1 Lisin 6.58 ± 0.07 4.58 7.55 5.9 7.9 Metionin 2.41 ± 0.08 - 2.47 0.7 2.6 Prolin 4.20 ± 0.10 - - - -Serin 4.11 ± 0.05 1.65 - 4.8 4.6 Sistein 0.91 ± 0.01 - 1.01 0.7 -Tirosin 4.53 ± 0.01 12.13 3.46 3.0 4.9 Treonin 4.14 ± 0.20 - 4.28 3.5 5.8 Triptophan 1.19 ± 0.07 - 0.85 - -Valin 5.95 ± 0.06 - 5.29 4.6 6.2
1)Shahidi dan Synowiecki (1992)
2)Purwantiningsih (1992)
3)Hartadi et al. (1997)
4)Thomas dan Beeson (1977)
5)Clark et al. (1992)
Kandungan mineral tepung limbah udang terutama kalsium (Ca) lebih tinggi dari tepung ikan, perbandingannya lebih dari 3 : 1. Akan tetapi phosfornya (P) lebih rendah, perbandingannya 1 : 2. Perbandingan antara Ca dan P dalam tepung limbah udang sendiri jauh lebih besar yaitu hampir 10 : 1. Oleh sebab itu, bila digunakan dalam ransum domba, perbandingan antara Ca dan P perlu diperhatikan karena yang dapat ditolerir hanya sampai 7 : 1 (NRC 1985). Perbandingan komposisi mineral antara tepung limbah udang dengan tepung ikan dan bungkil kedelai disajikan pada Tabel 3.
Tabel 3 Kandungan mineral tepung kepala udang, tepung ikan, dan bungkil kedelai
Mineral Tepung kepala udang1) Tepung ikan2) Bungkil kedelai3)
Ca (%) 15.30 4.20 0.3 Na (%) 2.05 0.97 -K (%) 0.20 0.68 2.1 Mg (%) 0.95 0.22 -P (%) 1.66 2.80 0.71 Sr (%) 0.22 - -Mn (ppm) 29 - -Fe (ppm) 82 - -Cu (ppm) 13 10.24 -Zn (ppm) 21 14.76 45 As (ppm) 27 - -Ba (ppm) 54 -
-1)Shahidi dan Synowiecki (1992)
2)
Hartadi et al. (1997)
3)Parakkasi (1999)
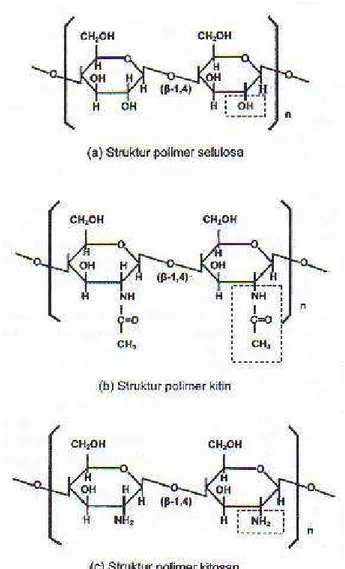
Selain potensi di atas, dalam limbah udang juga terdapat kitin antara 20 sampai dengan 30% (Wanasuria 1990) yang di dalamnya terkandung nitrogen (N) antara 6.6 sampai dengan 6.7% (Stelmoch et al. 1985). Bila digunakan dalam ransum ternak ruminansia, N tersebut berpotensi sebagai sumber N bukan protein (NPN) bagi mikroba rumen. Begitu juga kitinnya yang berupa polisakarida dengan bentuk molekulnya mirip selulosa (Gambar 1), apabila lolos dari rumen atau tidak dimanfaatkan oleh mikroba rumen, maka dalam pencernaan pascarumen akan mengikat asam empedu karena dianalogikan sebagai serat. Dengan demikian asam lemak yang diemulsi oleh asam empedu ikut terikat, karena serat (kitin) tidak dapat diabsorpsi pada usus halus (Piliang dan Djojosubagio 2006). Serat pakan selain lignin dan selulosa juga mengandung hemiselulosa, gum, dan pektin serta beberapa karbohidrat lain yang biasanya tidak dapat dicerna; maka bersama asam empedu dan asam lemak, diekskresikan bersama feses. Kondisi ini dapat menurunkan kolesterol serum darah dan daging pada ayam (Supadmo 1997).
Gambar 1 Struktur polimer selulosa (a), kitin (b), dan kitosan (c) (Hawab 2006).
Hidrolisis dan Pemanasan Protein
Hidrolisis protein diartikan sebagai pemecahan banyak ikatan menjadi satu ikatan atau putusnya ikatan peptida yang menghubungkan asam-asam amino (Girindra 1986). Pada limbah udang, hidrolisis yang baik untuk meningkatkan kecernaannya adalah dengan HCl 6% disertai pemanasan tekanan tinggi menggunakan pressure cooker selama 45 menit (Sudibya 1998).
Pemanasan mengakibatkan terjadinya perubahan pada suatu protein yang dikenal sebagai denaturasi (Lehninger 1982). Denaturasi dapat diartikan suatu perubahan atau modifikasi terhadap struktur sekunder, tersier, dan kuartener terhadap molekul protein, tanpa terjadinya pemecahan ikatan-ikatan kovalen.
Denaturasi dapat pula diartikan suatu proses terpecahnya ikatan hidrogen, interaksi hidrofobik, ikatan garam, dan terbukanya lipatan molekul (Winarno 1991).
Dua macam denaturasi, yaitu (1) pengembangan rantai peptida dan (2) pemecahan protein menjadi unit yang lebih kecil tanpa disertai pengembangan molekul. Terjadinya kedua jenis denaturasi ini tergantung pada keadaan molekul. Denaturasi yang pertama terjadi pada rantai polipeptida, sedangkan yang kedua terjadi pada bagian-bagian molekul yang tergabung dalam ikatan sekunder (Winarno 1991). Jadi proses denaturasi tidak merusak ikatan peptida yang terdapat antara asam amino dalam struktur primer (Girindra 1986).
Lehninger (1982) mengemukakan bahwa jika protein mengalami denaturasi, tidak ada ikatan kovalen pada kerangka rantai polipeptida yang rusak. Jadi deret asam amino khas protein tersebut tetap utuh setelah denaturasi. Selanjutnya dikemukakan bahwa rantai polipeptida yang berikatan kovalen pada protein asli (natif) melipat dalam tiga dimensi dengan suatu pola yang khas bagi tiap jenis protein. Jika suatu protein terdenaturasi, susunan tiga dimensi khas dari rantai polipeptida terganggu dan molekul ini terbuka menjadi struktur acak, tanpa adanya kerusakan pada struktur kerangka kovalen (Gambar 2).
Gambar 2 Sketsa proses denaturasi protein (Brandts 1967 disitir Winarno 1991).
Denaturasi dan koagulasi protein yang terjadi selama pemanasan mengakibatkan menurunnya kelarutan protein (Cheftel et al. 1985). Besarnya tingkat kelarutan protein setelah pemanasan dipengaruhi oleh suhu dan lama pemanasan yang digunakan (Hultin 1985).
Proses pemanasan protein yang tidak sampai merusak kandungan nutrisinya dilakukan dengan maksud agar kurang soluble dalam rumen. Cara ini biasa disebut heat treated protein (HTP) dan cara pemanasan yang sedang populer disebut dry extrusion process, yaitu pemanasan yang tidak memakai sumber panas dari luar. Panas terjadi akibat friksi (friction) dan diteruskan dengan expansion melalui extrusi (extrusion) dan tekanan (pressure). Konsep ini dilakukan karena protein tidak dapat dipenuhi dari mikroba rumen (terutama pada ternak yang berproduksi tinggi) maka tambahan asam-asam amino akan dapat dipenuhi dengan pemberian HTP yang langsung dapat digunakan pada pascarumen (Prawirokusumo 1994).
Proses Pencernaan pada Ruminansia
Pencernaan adalah serangkaian proses perubahan fisik dan kimia dari bahan makanan di dalam alat pencernaan sampai memungkinkan terjadinya proses penyerapan. Hewan ruminansia memiliki empat bagian perut dengan fungsi yang berbeda yaitu rumen, retikulum, omasum dan abomasum (Gambar 3). Sistem pencernaan pada ruminansia melibatkan interaksi dinamis antar pakan, populasi mikroba dan ternak itu sendiri (Mertens 1993). Prosesnya terdiri atas pencernaan mekanis di mulut, pencernaan fermentatif oleh mikroba di rumen dan pencernaan hidrolisis oleh enzim pencernaan pasca rumen (Sutardi 1977).
Gambar 3 Alat pencernaan pada ruminansia.
Pada hewan ruminansia, rumen dan retikulum tidak terpisah secara sempurna sehingga dipandang sebagai satu kesatuan (retikulorumen). Retikulorumen berfungsi sebagai tempat fermentasi makanan melalui aktivitas sejumlah mikroba dengan produk akhirnya berupa amonia (NH3), asam lemak
terbang (volatile fatty acid atau VFA), gas metan, dan air. Omasum fungsinya belum jelas, tetapi pada organ ini terjadi penyerapan air, NH3, dan VFA, diduga
juga memproduksi VFA dan NH3. Abomasum fungsinya sama dengan perut
monogastrik (Church dan Pond 1982; Forbes dan France 1993).
Makanan yang masuk ke dalam mulut akan mengalami proses pengunyahan atau pemotongan secara mekanis sehingga membentuk bolus. Pada proses ini makanan bercampur dengan saliva, kemudian masuk ke dalam rumen melalui esofagus untuk selanjutnya mengalami proses pencernaan fermentatif. Di rumen bolus dicerna oleh enzim mikroba, hasil pencernaan fermentatif berupa VFA, NH3
dan air. Selama di rumen makanan yang masih kasar dikembalikan lagi ke mulut (regurgitasi dan remastikasi). Partikel makanan yang tidak tercerna di rumen dialirkan ke abomasum dan dicerna secara hidrolitik oleh enzim pencernaan yang sama pada monogastrik. Hasil pencernaan tersebut diserap oleh usus halus dan selanjutnya masuk dalam sistem peredaran darah (Sutardi 1979).
Pencernaan dan Absorpsi Karbohidrat
Karbohidrat merupakan nutrien dominan dalam ransum ruminansia yaitu sekitar 60 sampai dengan 75% dari total nutrien ransum. Fungsi karbohidrat adalah sumber energi dan keambaannya bermanfaat untuk memelihara proses pencernaan. Karbohidrat berasal dari dua jenis yaitu karbohidrat dinding sel (selulosa dan hemiselulosa) banyak terkandung dalam hijauan pakan (roughage), dan karbohidrat isi sel (gula dan pati) banyak terkandung dalam pakan konsentrat. Selulosa merupakan zat penyusun tanaman, banyak mengandung unit glukosa, tersusun dalam rantai lurus, panjang dengan ikatan 1,4 dan biasanya dalam bentuk kristal. Hemiselulosa adalah karbohidrat rantai lurus terdiri atas polimer pentosa (silosa dan arabinosa), asam uronat dan galaktosa (Tillman et al. 1986).
Karbohidrat pakan di rumen akan mengalami tiga tahap pencernaan. Tahap pertama gula-gula kompleks seperti selulosa dan hemiselulosa dihidrolisis oleh enzim 1,4-glukosidase menjadi gula-gula sederhana (heksosa, pentosa dan selobiosa), sedangkan pati, dan sukrosa dihidrolisis menjadi maltosa, glukosa dan fruktosa. Tahap kedua, pemecahan gula-gula sederhana menjadi piruvat melalui siklus glikolisis Embden-Meyerhoff. Tahap tiga, piruvat oleh mikroba rumen
diubah menjadi asam lemak terbang (VFA) yaitu asetat, propionat dan butirat (Collier 1985).
Jenis karbohidrat pakan sangat besar pengaruhnya terhadap jumlah dan jenis VFA yang diproduksi. Konsentrasi VFA total di rumen bervariasi dan bergantung pada jenis ransum yang dikonsumsi (McDonald et al. 2002), pengolahan dan frekuensi pemberian makan (Sutardi 1977). Konsentrasi VFA yang optimum bagi kelangsungan hidup ternak berkisar 80 sampai dengan 160 mM, dengan proporsi 65% asetat, 20% propionat, 10% butirat dan 5% valerat serta asam lemak bercabang yaitu isobutirat, isovalerat, dan 2-metilbutirat (Czerkawski 1986). Asam lemak bercabang ini diduga berasal dari asam amino berantai cabang yaitu leusin, isoleusin, dan valin. VFA terutama yang berantai cabang, esensial bagi pertumbuhan mikroba rumen (Sutardi 1977). Proporsi VFA dapat berubah pada ransum tinggi serat, sehingga nisbah asetat/propionat lebih besar dari pada ransum tinggi konsentrat. Proporsi propionat meningkat pada ransum tinggi konsentrat, dan proporsi isobutirat serta isovalerat meningkat pada ransum tinggi protein (Sutardi 1977; Collier 1985).
Produk akhir berupa VFA dalam jaringan tubuh akan dimanfaatkan sebagai sumber energi dan bahan sintesis lemak. Asam propionat diabsorpsi melalui epitel rumen dan masuk ke sirkulasi darah, dibawa ke hati untuk selanjutnya diubah menjadi glukosa dan menjadi bagian dari cadangan glukosa hati. Asam butirat sebelum masuk ke sirkulasi darah dan dibawa ke hati bersama asetat dikonversi menjadi asam beta hidroksi butirat (-hydroxybutyric acid atau BHBA) di dalam epitel rumen (Banerjee 1978; Crampton et al. 1978). Asetat dan BHBA dari hati disalurkan ke sistem sirkulasi dan digunakan oleh jaringan sebagai sumber energi melalui siklus asam sitrat (Tillman et al. 1986; Forbes dan France 1993) dan sebagai substrat lipogenesis pada lemak susu, sedangkan propionat untuk glukoneogenesis dan lipogenesis lemak tubuh.
Pencernaan dan Absorpsi Protein
Protein yang dikonsumsi tidak seluruhnya didegradasi oleh mikroba rumen, sebagian ada yang lolos dan masuk ke abomasum, terus mengalir ke usus halus. Protein yang tidak tercerna akan mengalir ke sekum dan kolon dan difermentasi
oleh mikroba menjadi VFA dan NH3 selanjutnya diabsorpsi. Protein mikrobanya
tidak dapat dimanfaatkan oleh ternak dan keluar melalui feses.
Sebagian senyawa protein atau nitrogen bukan protein (NPN) asal ransum yang dihidrolisis oleh enzim proteolitik mikroba rumen dirombak menjadi oligopeptida dan asam amino yang merupakan produk intermediate. Selanjutnya oligopeptida dan asam amino mengalami deaminasi dan menghasilkan asam keto-, CO2, VFA, dan NH3(Sutardi 1977).
Kebanyakan mikroba rumen tidak dapat memanfaatkan asam amino secara langsung, karena mikroba terutama bakteri rumen tidak mempunyai sistem transpor untuk mengangkut asam amino ke dalam selnya. Lebih kurang 82% mikroba rumen membutuhkan N-NH3 untuk mensintesis protein selnya, oleh
karena itu mereka lebih suka merombak asam amino tersebut menjadi NH3
(Sutardi 1977).
Amonia merupakan sumber nitrogen utama untuk sintesis de novo asam amino mikroba rumen. Konsentrasi N-NH3 optimum untuk sintesis protein
mikroba rumen adalah 50 mg liter-1 atau 3.57 mM (Satter dan Slyter 1974), atau antara 4 sampai dengan 12 mM (Sutardi 1979). Kadar amonia yang optimum untuk mencapai efisiensi penggunaan energi dan protein pakan adalah 7 sampai dengan 8 mM (Erwanto et al. 1993). N-NH3 yang diproduksi tidak semua
digunakan untuk sintesis protein mikroba tetapi sisanya diserap oleh dinding rumen, masuk ke sirkulasi portal dan dibawa ke hati untuk selanjutnya diubah menjadi urea dan masuk ke sirkulasi darah (Tillman et al. 1986). Urea tersebut sebagian keluar melalui urin dan sebagian lagi masuk kembali ke rumen melalui saliva.
Pada sintesis protein mikroba rumen selain membutuhkan N-NH3 juga
membutuhkan asam keto- sebagai kerangka karbon dan VFA sebagai sumber energi. Setiap mol VFA yang dihasilkan akan terbentuk protein mikroba sekitar 14 sampai dengan 36 mg (Satter dan Slyter 1974). Mikroba rumen yang mati akan masuk ke usus dan berguna sebagai sumber protein bagi ternak. Protein mikroba bersama dengan protein ransum lolos degradasi di dalam usus mengalami pencernaan oleh protease usus dengan hasil akhir asam amino (Sutardi 1977; Leng et al. 1977).
Sumbangan protein asal mikroba rumen berkisar 40 sampai 80% (Sniffen dan Robinson 1987), sedangkan sumbangan energi asal VFA berkisar 60 sampai 80% (Ensminger et al. 1990). Pada ternak yang berproduksi tinggi, pasokan protein asal mikroba saja tidaklah cukup sehingga harus diberi protein yang tak terdegradasi oleh mikroba dalam rumen pada pakannya.
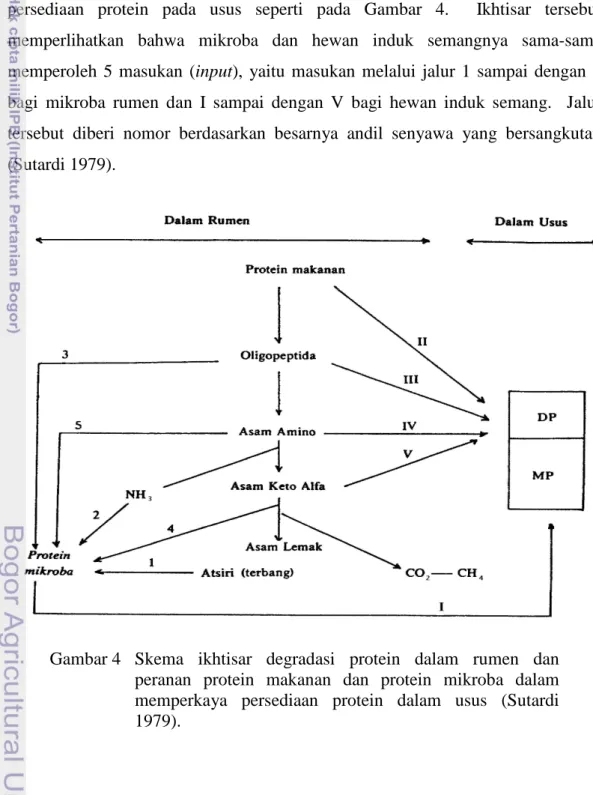
Gambaran tentang degradasi protein bahan makanan dalam rumen dan peranan bahan makanan yang tahan terhadap degradasi dalam memperkaya persediaan protein pada usus seperti pada Gambar 4. Ikhtisar tersebut memperlihatkan bahwa mikroba dan hewan induk semangnya sama-sama memperoleh 5 masukan (input), yaitu masukan melalui jalur 1 sampai dengan 5 bagi mikroba rumen dan I sampai dengan V bagi hewan induk semang. Jalur tersebut diberi nomor berdasarkan besarnya andil senyawa yang bersangkutan (Sutardi 1979).
Gambar 4 Skema ikhtisar degradasi protein dalam rumen dan peranan protein makanan dan protein mikroba dalam memperkaya persediaan protein dalam usus (Sutardi 1979).
Terdapat enam asam amino pembatas dalam pakan ruminansia yang harus diperhatikan ketersediaannya, yaitu metionin (Met), leusin (Leu), isoleusin (Ileu), valin (Val), lisin (Lys) dan treonin (Thr). Asam amino pembatas tersebut didasarkan pada adanya transfer Met dan asam amino bercabang (Leu, Ileu dan Val) ke dalam protein mikroba rumen yang cukup besar, Lys dirombak total dalam rumen, dan tidak dijumpai Thr dalam rumen (Sutardi 1997). Sebelumnya Merchen dan Titgemeyer (1992) melaporkan bahwa ternak ruminansia membutuhkan asam amino aromatik (fenilalanin dan triptofan) dan sejumlah asam amino yang bersifat semiesensial atau koesensial.
Pencernaan dan Absorpsi Lemak
Pencernaan lemak dimulai di retikulorumen. Lemak pakan mengalami dua peristiwa yaitu lipolisis dan biohidrogenasi di rumen (Scott dan Ashes 1993). Pada lipolisis oleh lipase mikroba, lemak atau minyak diurai menjadi asam lemak bebas (Free Fatty Acid atau FFA), gliserol dan galaktosa. Van Nevel dan Demeyer (1995) melaporkan bahwa semua asam lemak yang dilepaskan dari triasil gliserol terakumulasi dalam bentuk FFA, dan gliserol serta galaktosa difermentasi lebih lanjut menjadi VFA dengan proporsi terbanyak propionat (Scott dan Ashes 1993; Jenkins 1993). Asam lemak bebas tak jenuh secara cepat dihidrogenasi oleh mikroba rumen menjadi asam lemak jenuh.
Lipolisis pada beberapa lemak pakan terjadi sangat cepat di dalam rumen. Immig et al. (1993) melaporkan bahwa lipolisis minyak kedelai di rumen domba mencapai 90% selama satu jam setelah introduksi, dan di rumen sapi mencapai 85 sampai dengan 90% (Bauchart et al. 1990). Sumber lipase utama pada lipolisis berasal dari bakteri Anaerovibrio lipolytica, dan 30 persennya lipase asal protozoa (Harfoot dan Hazlewood 1988).
Liposisis menurun pada ransum berprotein rendah, atau berkadar pati atau sukrosa tinggi (Gerson et al. 1983). Menurunnya lipolisis tersebut berhubungan dengan rendahnya pH rumen. Van Nevel dan Demeyer (1995) melaporkan bahwa lipolisis minyak kedelai pada inkubasi in vitro (pH 5,25) sangat lambat akibat terhambatnya pertumbuhan mikroba lipolitik atau aktivitas lipase.
Pada proses hidrogenasi terjadi perubahan asam lemak oleat (C18:1), linoleat
(C18:2), dan linolenat (C18:3) menjadi stearat (C18:0) melalui isomerase dan
reduktase. Banyaknya linoleat yang dihidrogenasi di rumen antara 60 sampai dengan 95% dan linolenat antara 80 sampai dengan 100% (Doreau dan Ferlay 1994), proses hidrogenasi menjadi rendah pada ransum berkonsentrat lebih dari 70% (Bauchart et al. 1990). Asam lemak tak jenuh berantai lebih dari C18seperti
arakhidonat, eikosa pentanoat (EPA) dan dokosa heksaenoat (DHA) tidak mengalami hidrogenasi di rumen (Ashes et al. 1992).
Asam lemak yang telah mengalami lipolisis dan biohidrogenasi seperti C2
sampai dengan C14 dan VFA hasil pencernaan lemak langsung diserap melalui
dinding rumen, sedangkan asam lemak rantai panjang (lebih dari C14), asam lemak
jenuh hasil sintesis de novo dan lemak pakan terus mengalir ke abomasum dan diserap di usus halus.
Asam empedu mengemulsi lemak di usus halus. Partikel emulsi tersebut terutama mengandung triasil gliserol. Lipase pankreas merombak triasil gliserol menjadi mono dan diasil gliserol seperti halnya asam lemak bebas. Asam lemak bebas, mono dan diasil gliserol diserap secara difusi pasif masuk ke sel epitel brush border usus halus. Selanjutnya mono gliserol dan diasil gliserol di dalam sel mukosa usus dirakit kembali menjadi triasil gliserol dan dengan penambahan protein membentuk kilomikron (Collier 1985). Asam lemak dalam triasil gliserol tersebut semuanya mempunyai panjang rantai lebih dari 12 karbon.
Kilomikron yang terbentuk disekresikan melalui sistem limfe dan masuk ke aliran darah. Kilomikron plasma tersebut masuk ke dalam hati, jaringan adiposa atau jaringan perifer seperti ambing (Linder 1992; Collier 1985). Sel hati mengambil kilomikron langsung dari plasma, sedangkan organ lainnya harus dibantu oleh lipase lipoprotein dalam memecah kilomikron menjadi triasil gliserol. Asam lemak bebas tersebut selanjutnya masuk ke jaringan lemak dan urat daging untuk digunakan sebagai sumber energi atau disimpan kembali dalam bentuk trigliserida untuk digunakan kemudian. Kilomikron sisanya yang tidak terserap bersama-sama dengan hasil pencernaan karbohidrat dan protein dibawa ke hati melalui sistem portal untuk dimetabolisme lebih lanjut.
Domba
Domba diklasifikasikan dalam Kingdom: Animal, Phylum: Chordata (bertulang belakang), Kelas: Mamalia (menyusui), Ordo: Artiodactyla (berkuku genap), sub ordo: Ruminansia, Famili: Bovidae, Genus: Ovis, dan spesies: Ovis aries (Devendra dan Mcleroy 1992).
Domba yang dikenal di Indonesia ada tiga jenis yaitu: 1) Domba Jawa ekor kurus (JEK); 2) Domba Jawa ekor gemuk (JEG); dan 3) Domba Sumatera ekor kurus (SEK). Perbedaan masing-masing jenis domba ini dapat dilihat dari sifat-sifat luarnya, antara lain domba JEK dan SEK mempunyai ekor kurus dan panjang, warna bulu domba JEK bervariasi, pada jantan umumnya bertanduk. Domba SEK umumnya berwarna coklat muda dan pada jantan jarang yang bertanduk. Kedua jenis domba ini rata-rata bulunya (wool) kasar dan tersebar tidak teratur dibagian tubuhnya. Domba JEK mempunyai beberapa kelompok atau populasi lokal yang diberi nama sesuai dengan nama daerah atau tempat keberadaannya, seperti domba Garut dan Priangan. Domba JEG mempunyai ekor yang tebal, gemuk, dan pendek, umumnya dikenal sebagai penghasil wool, berwarna putih dan tidak bertanduk. Daerah penyebaran populasi domba ini di Jawa Timur, dipelihara terisolasi, sebagai contohnya di pulau Lombok (Tomaszewska et al. 1993).
Menurut Mason (1980), ada tiga jenis kelompok domba yang terdapat dipulau Jawa yaitu domba ekor tipis atau domba lokal, domba ekor gemuk, dan domba priangan atau dikenal sebagai domba ekor sedang. Domba ekor tipis merupakan domba asli Indonesia dikenal sebagai domba lokal, terdapat di daerah Jawa Barat dan Jawa Timur. Sumoprastowo (1993) mengemukakan bahwa domba lokal memiliki daya adaptasi yang baik pada iklim tropis. Domba jantan memiliki tanduk kecil dan melengkung kebelakang dengan bobot hidup dewasa berkisar 30 sampai dengan 40 kg. Domba betina tidak bertanduk dengan bobot hidup berkisar 15 sampai dengan 20 kg. Ukuran tubuhnya relatif kecil, warna bulunya beragam, ekor kecil dan panjangnya sedang, lambat dewasa, tidak seragam, berbulu kasar dan hasil dagingnya relatif sedikit, serta rata-rata bobot potongnya 20 kg (Edey 1983).
Pertumbuhan Ternak
Secara sederhana Butterfield (1988) mendefinisikan pertumbuhan sebagai terjadinya perubahan ukuran tubuh dalam suatu organisme sebelum mencapai dewasa, sedangkan perkembangan adalah produk hasil perbedaan pertumbuhan dari masing-masing bagian tubuh dari suatu organisme. Perubahan ukuran meliputi perubahan bobot hidup, bentuk dimensi linear dan komposisi tubuh termasuk pula perubahan pada komponen-komponen tubuh seperti otot, lemak, tulang dan organ dalam serta komponen kimia terutama air, lemak, protein dan abu (Soeparno 2005).
Pertumbuhan adalah bertambahnya bobot hingga ukuran dewasa tercapai atau lebih spesifik pertumbuhan dapat dijelaskan dengan bertambahnya produksi unit biokimia baru oleh pembagian sel, pembesaran sel atau persatuan dari bahan-bahan (material) yang berasal dari lingkungan. Perkembangan menunjukkan koordinasi berbagai proses hingga kematangan (kedewasaan) tercapai, seperti diferensiasi selular dan perubahan bentuk tubuh. Pertumbuhan pada umumnya dinyatakan dengan mengukur kenaikan bobot hidup yang mudah dilakukan dan biasanya dinyatakan sebagai pertambahan bobot hidup harian atau average daily gain (ADG). Pertumbuhan yang diperoleh dengan memplotkan bobot hidup terhadap umur akan menghasilkan kurva pertumbuhan (Tillman et al. 1986 dan Taylor 1984).
Pertumbuhan ternak terdiri atas tahap cepat yang terjadi mulai awal sampai pubertas dan tahap lambat yang terjadi pada saat kedewasaan tubuh telah tercapai (Tillman et al. 1986). Pada waktu kecepatan pertumbuhan mendekati konstan, slope kurva pertumbuhan hampir tidak berubah. Hal ini berarti pertumbuhan otot, tulang dan organ-organ penting mulai berhenti, sedangkan penggemukan (fattening) mulai dipercepat (Judge et al. 1989).
Tumbuh-kembang dipengaruhi oleh faktor genetik, pakan, jenis kelamin, hormon, lingkungan dan manajemen (Judge et al. 1989). Beberapa faktor utama yang mempengaruhi pertumbuhan domba sebelum lepas sapih adalah genotipe, bobot lahir, produksi susu induk, jumlah anak per kelahiran, umur induk, jenis kelamin anak dan umur sapih (Edey 1983). Potensi pertumbuhan dalam periode ini dipengaruhi oleh faktor bangsa, heterosis (hybrid vigour) dan jenis kelamin.
Pola pertumbuhan ternak tergantung pada sistem manajemen (pengelolaan) yang dipakai, tingkat nutrisi pakan yang tersedia, kesehatan dan iklim. Batubara et al. (1993) menyatakan bahwa pertambahan bobot hidup domba lokal Sumatera jantan muda dengan menggunakan pakan konsentrat komersial yang dicampur bungkil inti sawit (40%), molases (20%) dan urea (0.5%) adalah sebesar 106 g/ekor/hari dan konversi pakan adalah 8.2, sedangkan dengan pakan konsentrat kualitas tinggi (pakan komersial) pertambahan bobot hidup adalah 100 g/ekor/hari dan konversi pakan sebesar 9.4. Perbedaan bangsa memberikan keragaman dalam kecepatan pertumbuhan dan komposisi tubuh. Ternak dari satu bangsa tertentu cenderung tumbuh dan berkembang dalam suatu sifat yang khas dan menghasilkan karkas dengan sifat tersendiri, sehingga merupakan sifat khas bangsanya (Judge et al. 1989).
Karkas dan Komponen Karkas
Bobot karkas adalah bobot hidup setelah dikurangi bobot saluran pencernaan, darah, kepala, kulit dan keempat kaki mulai dari persendian carpus atau tarsus ke bawah. Dijumpai sedikit modifikasi, kadang-kadang dengan atau tanpa ginjal, lemak ginjal, lemak pelvis, lemak sekitar ambing, diaphragma dan ekor. Perbedaan sangat besar adalah lemak ginjal atau lemak pelvis termasuk ke dalam karkas atau tidak (Berg dan Butterfield 1976). Karkas sebagai satuan produksi dinyatakan dalam bobot karkas dan persentase karkas. Persentase karkas adalah perbandingan antara bobot karkas dengan bobot hidup saat dipotong (dikurangi isi saluran pencernaan dan urine) dikali 100% (Berg dan Butterfield 1976; Tulloh 1978; Judge et al. 1989). Persentase karkas dipengaruhi oleh bobot karkas, bobot ternak, kondisi, bangsa ternak, proporsi bagian-bagian non karkas, ransum yang diberikan dan cara pemotongan (Berg dan Butterfield 1976).
Herman (2005) menyatakan bahwa persentase karkas domba Priangan adalah sebesar 55.1 % dan domba ekor gemuk adalah sebesar 55.3% pada bobot potong 40 kg. Persentase karkas bervariasi karena umur dan perlemakan dari domba tersebut, sedangkan persentase tulang, otot dan lemak dalam karkas dipengaruhi oleh umur, bangsa dan perlemakan pada domba. Hasil penelitian Sugiyono (1997) mendapatkan bahwa bobot karkas domba lokal yang diberi
pakan konsentrat biasa adalah sebesar 7.5 kg dari bobot hidup 19.3 kg dan persentase karkasnya 39.1%. Johnston (1983) menyatakan bahwa persentase karkas pada domba yang kurus dan kondisinya buruk kurang dari 40%, sedangkan pada kondisi gemuk persentase karkas dapat melebihi 60%. Pendapat lain dikemukakan Tulloh (1978) bahwa apabila ternak tidak diberi makan atau minum untuk suatu periode tertentu (dua hari misalnya) maka persentase karkas akan meningkat karena berkurangnya jumlah urine dan feses selama periode tertentu. Komposisi pakan juga berpengaruh terhadap besarnya persentase karkas. Ternak yang mendapat pakan hijauan dengan mutu yang rendah, mengandung lebih banyak digesta didalam saluran pencernaannya dari pada ternak yang diberi pakan bermutu tinggi dengan proporsi biji-bijiannya yang tinggi. Ternak yang dipuasakan keragaman persentase karkasnya dapat mencapai 4% lebih besar (Tulloh 1978). Soeparno (2005) mengemukakan bahwa perbedaan komposisi tubuh dan karkas diantara bangsa ternak disebabkan oleh perbedaan ukuran tubuh dewasa atau perbedaan bobot pada saat dewasa.
Komponen utama karkas terdiri atas jaringan otot, tulang dan lemak. Kualitas karkas sangat ditentukan oleh ketiga komponen tersebut. Tulang sebagai kerangka tubuh, merupakan komponen karkas yang tumbuh dan berkembang paling dini, kemudian disusul oleh otot dan yang paling akhir oleh jaringan lemak (Soeparno 2005). Proporsi komponen karkas dan potongan karkas yang dikehendaki konsumen adalah karkas atau potongan karkas yang terdiri atas proporsi daging tanpa lemak (lean) yang tinggi, tulang yang rendah dan lemak optimal (Natasasmita 1978). Komponen karkas yang dapat memberikan nilai ekonomis adalah lemak, karena lemak berfungsi sebagai pembungkus daging dan memberikan keempukan pada daging (Berg dan Butterfield 1976).
Hasil penelitian Sugiyono (1997) mendapatkan bahwa domba lokal yang diberi pakan konsentrat biasa, persentase daging tanpa lemak (lean), lemak dan tulangnya berturut-turut adalah sebesar 62.63%, 5.42% dan 24% dari bobot setengah karkas. Herman (2005) dan Rachmadi (2003) menyatakan bahwa semakin tinnggi bobot potong yang diperoleh menyebabkan bobot karkas segar dan persentase karkas akan semakin tinggi. Herman (2005) menyatakan bahwa pada bobot potong 17.5 kg karkas, otot, tulang, dan lemak pada domba Priangan
berturut-turut adalah 8.290, 2.554, 720 dan 598 gram sedangkan untuk domba ekor gemuk berturut-turut 8.530, 2.521, 724 dan 794 gram. Rachmadi (2003) menyatakan bahwa domba yang diberikan pakan konsentrat yang mengadung bungkil inti sawit sebanyak 45% mempunyai bobot tubuh kosong, bobot karkas dan persentase karkasnya berturut-turut adalah sebesar 14.30 %, 6.24 % dan 43.57% dengan masa pengemukan enam bulan.
Murray dan Slezacek (1976) melaporkan bahwa tidak ada perbedaan dalam proporsi daging, tulang dan jaringan ikat maupun perlemakan pada tingkat pemberian pakan dan perbedaan pada domba, tetapi berbeda dalam depot lemak tubuhnya. Domba yang mendapatkan pakan lebih banyak mempunyai lemak subkutan lebih banyak, namun lemak intramuskuler lebih rendah. Soeparno (2005) dan Ouhayoun (1998) mengemukakan bahwa perlemakan dipengaruhi oleh bobot karkas dan konsumsi ransum, khususnya energi. Makin tinggi bobot karkas, lemak karkas makin meningkat.
Karkas domba dibagi menjadi dua bagian besar yaitu : Pertama foresaddle (bagian depan) meliputi: neck (leher), shoulder (bahu), shank (paha depan), rack (dada) dan breast (bagian bawah dada). Kedua hindsadle (bagian belakang) meliputi: leg (paha belakang), loin (pinggang) dan flank (bagian bawah perut) (Judge et al. 1989). Domba lokal jantan mempunyai komposisi potongan komersial karkas pada bobot potong 15 kg adalah sebagai berikut : leg (34.47%), loin (9.40%), rib (9.46%), shoulder (21.87%), shank (3.74%), breast (9.01%) dan neck (8.98%) (Triatmojo 1988). Judge et al. (1989) menyatakan bahwa komposisi leg (39%), loin (7%), rib (9%), shoulder (26%), shank (5%), breast (10%), flank (2%), ginjal dan lemak ginjal (2%).
Herman (2005) menyatakan bahwa pada irisan karkas utama, memperlihatkan bahwa domba Priangan mempunyai potongan shoulder yang lebih besar dengan persentase otot lebih tinggi dan lemak lebih rendah dibandingkan dengan irisan shoulder domba Ekor Gemuk. Domba Ekor Gemuk mempunyai potongan leg yang lebih besar daripada domba Priangan, tetapi persentase lemaknya lebih tinggi dan ototnya lebih rendah. Persentase potongan leg pada domba Priangan (30.8%), Ekor Gemuk (32.3%), loin pada domba Priangan (9.1%), Ekor Gemuk (10.1%), rack pada domba Priangan (9.4%), Ekor
Gemuk (8.6%) dan shoulder pada domba Priangan (28.2%), Ekor Gemuk (27.3%).
Peningkatan bobot karkas segar akan meningkatkan bobot leg, neck, loin, rack dan shoulder, dimana persentase peningkatan bobot terbaik ditunjukkan oleh potongan leg disusul oleh loin, rack dan shoulder (Beermann et al. 1986). Saparto (1981) menyatakan bahwa persentase shank meningkat dengan menurunnya bobot karkas, sebaliknya persentase loin dan rack meningkat dengan naiknya bobot karkas. Lebih lanjut dinyatakan bahwa pada domba jantan, otot pada shoulder, leg, loin, dan breast mempunyai sifat masak dini sehingga pertumbuhannya relatif lebih cepat dibandingkan dengan bagian bagian tubuh lainnya. Sugana et al. (1983) melaporkan bahwa persentase loin meningkat, persentase leg berkurang dengan meningkatnya bobot karkas.
Sifat Fisik dan Kimia Daging
Kualitas atau mutu daging ditentukan oleh keempukan (tenderness), cita rasa (flavour), tekstur, aroma, warna sari minyak atau jus daging (juiceness), lemak intramuskuler (marbling), hilangnya air selama perebusan atau susut masak (cooking loss), daya mengikat air oleh protein daging (water holding capacity, WHC), dan pH daging (Aberle et al. 2001). Gurnadi (1986) menyatakan bahwa ada 3 faktor sebagai kriteria untuk menentukan mutu daging yaitu : 1) nilai gizi (ditentukan oleh protein, lemak, karbohidrat, vitamin, mineral); 2) selera konsumen (penampilan: seperti warna, keempukan, marbling, ketegaran atau firmness, juiciness, dan tekstur); 3) teknologi penanganan atau pengolahan (daya mengikat air yang tinggi, memiliki kecenderungan lemak tertentu, kandungan jaringan ikat dan air tertentu).
Pada dasarnya ada tiga komponen yang menentukan keempukan daging, yaitu sytruktur miofibrilar dan status kontraksi otot, kandungan jaringan ikat dan tingkat ikatan silangnya, serta daya mengikat air dan jus daging (Soeparno 2005). Komponen utama yang mempengaruhi keempukan daging adalah jaringan ikat, kelompok serat otot dan lemak. Jaringan ikat terutama kolagen dan jumlah ikatan silangnya memiliki peran yang besar terhadap keempukan daging (Aberle et al. 2001).
Menurut Soeparno (2005) pH ultimat daging adalah pH yang dicapai setelah glikogen otot habis atau setelah enzim-enzim glikolitik menjadi tidak aktif pada pH rendah atau setelah glikogen tidak lagi sensitif terhadap serangan enzim-enzim glikolitik. Lawrie (2003) menyatakan bahwa penurunan pH postmortem dipengaruhi oleh faktor intrinsik dan ekstrinsik. Faktor intrinsik antara lain spesies, tipe otot, glikogen otot dan variabilitas diantara ternak, sedangkan faktor ekstrinsik antara lain adalah temperature lingkungan perlakuan sebelum pemotongan dan suhu penyimpanan. Penurunan pH otot postmortem juga bervariasi diantara ternak. Pada sejumlah ternak dapat ditemukan bahwa pH karkas atau daging hanya menurun sedikit selama beberapa jam pertama setelah pemotongan, dan pada saat tercapainya kekakuan daging, pH tetap tinggi yaitu antara 6.5 sampai dengan 6.8. Pada ternak yang lain, pH dapat menurun dengan cepat hingga mencapai 5.4 sampai dengan 5.5 selama beberapa jam pertama setelah pemotongan (Aberle et al. 2001).
Komponen air dalam jaringan daging ada 3 bentuk yaitu: 1) air bebas (free water) yang jumlahnya 4 sampai dengan 10 gram/100 gram protein; 2) air tak bergerak (immobilized water) dengan jumlah 20 sampai dengan 60 gram/100 gram protein dan 3) air yang terikat erat (tightly bound water) yang jumlahnya 300 sampai dengan 360 gram/100 gram protein. Jika dilakukan penekanan (pressur) atau sentrifugasi, maka hanya air yang dalam bentuk bebas saja yang terpisah, sementara air yang tak bergerak dan air yang terikat erat, tetap tertinggal dalam daging. Air yang tak bergerak dan air yang terikat erat, sangat menentukan water holding capacity (WHC) (Soeparno et al. 1990). Daya mengikat air (WHC) berkonstribusi terhadap tenderness (keempukan), warna, rasa dan semua penentu kualitas daging. Kehilangan cairan selama penyimpanan dan pemrosesan akan merugikan produsen daging, prosessor dan konsumen (Luts et al. 1992).
Protein daging berdasarkan jaringan asalnya dapat dibagi dalam 3 yaitu: 1) protein sarkoplasma; 2) protein dari serat-serat daging (miofibril); dan 3) protein dari jaringan pengikat. Sedangkan berdasarkan struktur molekulnya maka protein daging adalah: myosin, aktin, globulin, mioglubin,hemoglobulin, kolagen elastin dan enzim.
Sementara lemak daging sekitar 95% disusun oleh trigliserida, 5% disusun oleh kolesterol, fosfolipid dan vitamin ADEK. Trigeliserida pada lemak daging disusun oleh asam lemak jenuh dan asam lemak tidak jenuh dari molekul dengan rantai atom C5 sampai dengan C20. Asam lemak tak jenuh yang menyusun lemak
daging terdiri dari asam lemak tak jenuh tunggal (monosaturated fatty acids) dan asam lemak poli tak jenuh (polysaturated fatty acids). Makin tinggi imbangan asam lemak tak jenuh dengan asam lemak jenuh, maka lemak tersebut konsistensinya akan semakin lembek. Disamping itu, asam lemak tak jenuh cepat tengik karena ikatan rangkap pada asam lemak tak jenuh mudah dioksidasi oleh oksigen udara yang menghasilkan senyawa peroksida yang menimbulkan bau tengik (Soeparno et al. 1990).
Lemak dan Trigliserida
Lemak merupakan subtansi yang dapat ditemukan dalam jaringan tumbuhan dan hewan. Lemak tidak larut dalam air tetapi larut dalam pelarut organik seperti benzene, eter dan khloform. Lemak mengandung karbon, oksigen, dan hidrogen dengan rumus: C12H22O11 (Mc Donald et al. 2002). Lemak biasa disebut ester
lemak murni dari gliserol yaitu trigliserida. Lemak merupakan ikatan organik yang masuk ke dalam klasifikasi lipid bersama-sama dengan ikatan kimia lainnya termasuk lilin, fosfolipid dan sterol (Wahyu 1985).
Frandson (1992) menyatakan bahwa lemak digolongkan menjadi lemak sederhana, lemak gabungan dan lemak derivat. Lemak sederhana adalah ester dari asam-asam lemak dan alkohol termasuk macam-macam lemak (ester asam lemak dan gliserol) dan wax (ester asam lemak dan alkohol selain gliserol). Lemak gabungan mengandung beberapa gugus selain alkohol dan asam lemak seperti fosfor, nitrogen, dan karbohidrat. Lemak derivat merupakan senyawa yang dihasilkan oleh hidrolisa lemak sederhana ataupun lemak gabungan.
Lemak dalam daging terdapat dalam bentuk trigliserida. Trigliserida merupakan komponen utama asam lemak dalam makanan yang dibentuk dari fraksi katalisa gliserol dengan 3 molekul asam lemak. Trigliserida merupakan bentuk lemak yang paling efisien untuk menyimpan kalori (Piliang dan Djojosoebagjo 2006). Selanjutnya dikatakan bahwa kelebihan energi terjadi jika
energi melebihi kebutuhan metabolisme; kelebihan energi menyebabkan akumulasi lemak yang berlebihan sehingga disimpan dalam jaringan lemak dalam bentuk cadangan lemak. Beberapa trigliserida berbentuk butir-butir kecil pada jaringan yang digunakan untuk metabolisme energi.
Asam lemak adalah komponen terbesar dari beberapa lipida kompleks yang mengandung 12 sampai dengan 24 atom C yang umumnya sebagian besar terdapat pada jaringan hewan. Sebagai contoh adalah asam linoleat yang diketahui dapat menurunkan taraf kolesterol dalam darah juga dipertimbangkan sebagai asam lemak esensial, tetapi ternyata bahwa asam linoleat dapat merangsang pembentukan tumor dan penyebaran tumor (metastase) (Enser 1984). Konsumsi yang berlebihan dari lemak yang mengandung asam linoleat tinggi, dipercaya dapat merangsang kanker payudara, prostat dan kanker usus besar (Adnan 1994).
Trigliserida adalah komponen utama dari penyimpanan lemak atau depot lemak pada tumbuhan dan hewan; umumnya tidak dijumpai pada membran. Bagian utama trigliserida adalah asam lemak tidak jenuh dan bila disimpan dalam suhu kamar akan berbentuk cair (Lehninger 1997). Trigliserida yang ada pada tubuh hewan, 95% berasal dari makanan, dan 5 % disintesis oleh tubuh. Murray et al. (1990) menyatakan bahwa dalam saluran pencernaan, sebagian besar trigliserida dipecah menjadi monogliserida dan asam lemak, kemudian ketika melalui epitel usus, disentesis kembali menjadi molekul trigleserida yang baru dan masuk ke dalam limfe dan bentuk droplet (butiran kecil) yang tersebar (kilomikron). Selain itu, sebagian besar kolesterol dan fosfolipid diabsorbsi dari saluran pencernaan kemudian masuk ke dalam kilomikron.
Fungsi dari trigliserida adalah sebagai cadangan energi. Trigliserida merupakan lemak yang efisein untuk dipakai sebagai cadangan energi dan tidak banyak membutuhkan tempat serta dapat menghasilkan energi lebih besar dibandingkan karbohidrat dan protein dengan jumlah yang sama yaitu dengan perbandingan karbohidrat : protein : lemak sebesar 1 : 1: 2,5 (Piliang dan Djojosoebagio 2006).
Trigliserida yang disintesis dalam hati akan digunakan untuk memproduksi lipoprotein darah dimana pemenuhan kebutuhan asam lemak, berasal dari makanan, dari jaringan adiposa melalui darah atau dari biosentesis hati.
Syamsuhaidi (1997) mengemukakan bahwa imbangan energi protein ransum yang diperluas dapat meningkatkan konsentrasi trigliserida yang ada di serum darah.
Kolesterol
Kolesterol merupakan subtansi putih yang larut dalam lemak, terdapat dalam lemak hewani dan minyak, empedu, darah, jaringan urat saraf, hati, ginjal dan kelenjar adrenal serta penting dalam metabolisme. Kolesterol dalam tubuh dapat berupa kolesterol endogenus dan kolestrol eksogenus. Kolesterol endogenus merupakan kolesterol yang dibentuk sendiri oleh sel-sel tubuh terutama di dalam hati. Kolesterol eksogenus merupakan kolesterol yang berasal dari makanan. Kolesterol dibutuhkan oleh tubuh untuk tumbuh besar dan memperbaiki sel-sel yang rusak, menghasilkan asam empedu yang dibutuhkan dalam penyerapan lemak (Murray et al. 1990). Lebih lanjut dikemukakan bahwa keseimbangan kolesterol dalam jaringan dapat meningkat atau menurun. Perubahan keseimbangan ini dipengaruhi oleh beberapa faktor. Peningkatan terjadi karena (1) pengambilan lipoprotein yang mengandung kolesterol oleh reseptor, seperti LDL; (2) pengambilan lipoprotein yang mengandung kolesterol oleh lintasan yang tidak diantarai reseptor; (3) pengambilan kolesterol bebas dari lipoprotein yang kaya akan kolesterol itu oleh membran sel; (4) sintesis kolesterol; (5) hidrolisis ester kolesteril oleh enzim ester kolesteril hidrolase. Penurunan terjadi karena (1) aliran keluar kolesterol dari membran sel ke lipoprotein dengan potensial kolesterol rendah, khusus DHL3 atau HDL nasen,
yang digalakkan oleh LCAT (lesitin:kolesterol asiltransferase); (2) esterifikasi kolesterol oleh ACAR (asil-KaO: Kolesterol asiltransferase); (3) penggunaan kolesterol untuk sintesis senyawa-senyawa steroid lainnya, seperti hormon atau asam-asam empedu dalam hati. Guyton (1994) mengemukakan bahwa faktor-faktor yang mempengaruhi konsentrasi kolesterol plasma adalah kenaikan jumlah kolesterol yang dicerna tiap hari sedikit meningkatkan konsentrasi plasma. Diet lemak jenuh dapat meningkatkan konsentrasi kolesterol darah sebanyak 15-25% karena adanya penimbunan lemak dalam hati, pencernaan lemak yang mengandung asam lemak tidak jenuh yang tinggi (biasanya menekan konsentrasi kolesterol). Kolesterol darah dapat meningkat pada diabetes mellitus akibat dari
peningkatan umum mobilisasi lemak. Hormon seks wanita yaitu estrogen dapat menurunkan kolesterol darah dan hormon seks pria yaitu androgen dapat meningkatkan kolesterol dalam darah.
Jalur utama pengeluaran kolesterol tubuh adalah melalui konversi oleh hati menjadi asam empedu. Kurang lebih separuh dari kolesterol dieksresikan ke dalam feses setelah sebelumnya diubah menjadi asam-asam empedu dan sebahagian besar kolesterol yang diekskresi ke dalam empedu akan diserap kembali (Murray et al. 1990).
Arora (2007), menjelaskan bahwa kolesterol bekerja membantu mengangkut lemak yang diolah dari hati ke seluruh tubuh melalui pembuluh darah. Proses ini akan terus berjalan dan berulang-ulang. Secara garis besar, kerja kolesterol dapat dijelaskan sebagai berikut:
1. Makanan atau lemak masuk lambung untuk diemulsi oleh asam empedu kemudian diteruskan masuk ke usus halus untuk dicerna dan diserap. Sari-sari makanan yang telah diserap kemudian dikirim ke hati untuk diproses lalu dikirim ke seluruh tubuh.
2. Hati mengandung lemak (VLDL), VLDL masuk melalui pembuluh-pembuluh darah, membongkar muatannya (lemak) diseluruh tubuh. Kemudian VLDL yang kosong berubah menjadi LDL.
3. Beberapa potongan LDL dapat tersangkut di sepanjang dinding pembulun darah, sehinga mempersempit pembuluh darah.
4. HDL dalam darah atau pembuluh darah berfungsi untuk melepaskan potongan LDL yang tersangkut di dinding pembuluh-pembuluh darah dan membawanya kembali ke hati. Di hati, LDL tersebut di daur ulang menjadi VLDL atau dihancurkan dan dibuang. VLDL yang baru akan memulai kembali proses pengiriman.
Menurut Arora (2007), kolesterol dapat dikelompokan menjadi beberapa jenis, yaitu:
1. LDL (Low Density Lipoprotein)
LDL berfungsi mengangkut kolesterol dari hati ke seluruh bagian tubuh. Jika kolesterol yang tersedia lebih dari yang dibutuhkan, LDL akan beredar dalam aliran darah dan akhirnya tertimbun pada bagian dalam dinding pembuluh
darah yang kemudian dapat menyebabkan penyumbatan dan berkurangnya pasokan darah
2. HDL (High Density Lipoprotein
HDL merupakan lipoprotein yang berjalan mengikuti aliran darah dari bagian tubuh (perifer), sambil membawa kolesterol ke hati untuk dihancurkan. Selain itu, jenis kolesterol ini juga berfungsi untuk mengangkut kolesterol bebas yang terdapat dalam endotel jaringan perifer, termasuk pembuluh darah, ke reseptor HDL di dalam hati untuk keluar lewat empedu, sehingga penimbunan kolesterol di perifer berkurang. Dalimartha (2002) menyatakan bahwa HDL merupakan lipoprotein yang diharapkan tinggi dalam tubuh 3. VRDL
VRDL merupakan partikel-partikel lemak yang disebut siklomikro dan asam lemak pembentuk VLDL yang digunakan untuk energi dan pemindahan lemak.
4. Trigliserida
Trigliserida merupakan jenis lemak yang diproduksi oleh hati. Pada manusia, lebih dari 5% lemak yang dikonsumsi, dalam bentuk trigliserida.
5. Lipoprotein (a)
Lipoprotein (a) merupakan lemak yang berkaitan dengan aterosklerosis dan berbagai penyakit arteri koroner. LP (a) lebih berkaitan dengan gen.
Tingginya kolesterol dalam tubuh sering menimbulkan ganguan bagi kesehatan. Gangguan tersebut berupa ateroklerosis yang diakibatkan karena adanya penimbunan LDL dan trigliserida yang selanjutnya menjadi penyakit jantung koroner dan stroke. LDL merupakan lipoprotein yang mudah melekat pada dinding sebelah dalam pembuluh darah dan menyebabkan penumpukan lemak yang dapat menyempitkan pembuluh darah.