HUBUNGAN KEKERABATAN FILOGENETIKA KULTIVAR PISANG
DI INDONESIA BERDASARKAN KARAKTER MORFOLOGI
Amin Retnoningsih Jurusan Biologi
Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Negeri Semarang Kampus Sekaran, Gunungpati Semarang 50229
Korespondensi: aminrn@yahoo.com
Amin Retnoningsih. 2011. Phylogenetic relationship of cultivated bananas in Indonesia based on
morpho-logical characters. Floribunda 4(2): 48–53. — Indonesia has high biodiversity of cultivated bananas. This
research objective is reveal the phylogenetic relationship of cultivated bananas in Indonesia based on mor-phological characters. This research analyzed 2 groups which are consist of ingroup namely 9 banana ac-cessions on genome groups AA, 9 AAA accession, 8 AAB accession, 8 ABB accession, and 2 BB accession,
and the other group is outgroup which representated by 2 wild bananas. The method of this research using
Cladistic program Phylogenetic Analysis Using Parsimony (PAUP) version 4.0, it used to produce
phyloge-netic trees. The study branches are tested by bootstrap 100 times. The results show that morphological cha-racters have parallel homoplasm which can be seen from the low CI score by 0.275. Taxon tested are species
and under species level (Musa spp). In general, there is no difference on changes of morphological characters
in each clade. The clade consist of Boi accession (ABB), Klutuk Wulung (BB) and Klutuk (BB) separated
from ingroup clade because they have differences of morphological characters from the others. They are
showed by high bootstrap score: 98%. The Klutuk Wulung and Klutuk are predicted to have close
phyloge-netic relationship with the wild Musa balbisiana (BB).
Keywords: Phylogenetic relationship, Indonesia, morphological characters, banana.
Amin Retnoningsih. 2011. Hubungan kekerabatan filogenetika kultivar pisang di Indonesia berdasarkan
karakter morfologi. Floribunda 4(2): 48–53. — Keanekaragaman kultivar pisang di Indonesia sangat tinggi.
Penelitian ini bertujuan mengungkap hubungan kekerabatan filogenetika kultivar pisang di Indonesia
ber-dasarkan karakter morfologi. Analisis dilakukan terhadap 9 aksesi pisang grup genom AA, 9 aksesi AAA, 8
aksesi AAB, 8 aksesi ABB, dan 2 aksesi BB sebagai ingroup, serta 2 jenis pisang liar sebagai outgroup.
Me-tode kladistika program Phylogenetic Analysis Using Parsimony (PAUP) versi 4.0 digunakan untuk
menghasilkan pohon filogenetika yang kekokohan percabangannya diuji dengan bootstrap 100 kali. Hasil penelitian menunjukkan bahwa karakter morfologi memiliki homoplasi paralel yang dapat dilihat dari nilai
CI yang rendah yaitu 0.275. Kelompok takson yang dikaji merupakan tingkat jenis dan di bawah jenis (Musa
spp) sehingga secara umum perubahan karakter morfologi di setiap klad tidak berbeda. Klad yang terdiri atas
aksesi Boi (ABB), Klutuk Wulung (BB) dan Klutuk (BB) memisah dari klade ingroup karena memiliki
karakter morfologi yang berbeda dari aksesi yang lain, ditunjukkan melalui nilai bootstrap yang tinggi yaitu 98%. Kultivar Klutuk Wulung dan Klutuk ini diduga memiliki hubungan kekerabatan yang sangat dekat
dengan jenis liar Musa balbisiana (BB).
Kata kunci: Hubungan kekerabatan filogenetika, Indonesia, karakter morfologi, pisang. Lebih dari 200 kultivar pisang dan 15 jenis
pisang liar diketahui hidup di wilayah Indonesia (Nasution & Yamada 2001). Sebagian di anta-ranya, termasuk hasil eksplorasi pisang di Irian Jaya (INIBAP 2002) merupakan kultivar yang unik. Kultivar pisang merupakan keturunan dari
perkawinan antara jenis pisang liar M acuminata
Colla (genom AA) dan M. balbisiana Colla
(genom BB) (Stover & Simmonds 1987) yang ke-mudian mengalami proses evolusi sampai menjadi pisang yang dapat dimakan. Melalui proses evolusi
ini, fertilitas yang tinggi pada jenis-jenis liar ber-angsur-angsur hilang sampai menjadi pisang tidak berbiji. Selain kehilangan bijinya, proses perubah-an ini juga menghasilkperubah-an pisperubah-ang dengperubah-an berbagai tingkat ploidi dan grup genom seperti pisang di-ploid genom AA. BB dan AB; pisang tridi-ploid ge-nom AAA, AAB, ABB, dan BBB, serta pisang tetraploid genom ABBB.
Proses seleksi dan perbanyakan vegetatif diyakini sebagai faktor utama terjadinya pisang tanpa biji (Verheij & Coronel 1992). Perubahan
pisang liar berbiji menjadi pisang tanpa biji dapat diungkap melalui analisis filogenetika. Selain itu, analisis ini juga penting karena diduga dalam waktu dekat pisang budi daya akan punah (INI-BAP 2003). Beberapa dekade terakhir ini telah terjadi penurunan produksi pisang budi daya se-cara dramatis akibat serangan hama dan penyakit. Kekhawatiran ini mendorong para peneliti me-ngumpulkan seluruh informasi kekayaan plasma nutfah yang ada untuk melacak sumber-sumber resistensi (Megia 2005). Upaya yang perlu dilaku-kan antara lain adalah mengevaluasi seluruh keka-yaan plasma nutfah. Proses evolusi genom pisang juga dipandang penting dipelajari untuk mema-hami peranan hibridisasi dan evolusi poliploidi tanaman budi daya yang lain (INIBAP 2001).
Beberapa alasan pentingnya informasi hu-bungan kekerabatan filogenetika pisang di Indone-sia adalah 1) bermacam-macam jenis pisang liar yang hidup di wilayah Indonesia seperti anak
jenis-anak jenis M. acuminata diduga sebagai tetua
pisang budi daya; 2) morfologi yang bervariasi seperti bentuk, ukuran, warna dan sifat-sifat yang lain dari batang semu, anakan, tangkai dan helai daun, braktea, bunga jantan dan buah; 3) grup ge-nom yang bervariasi dengan sterilitas yang tinggi. Kajian filogenetika diharapkan dapat memberi gambaran evolusi pisang sehingga diperoleh pema-haman bagaimana keanekaragaman pisang di Indo-nesia dapat terjadi.
Hipotesis hubungan kekerabatan filoge-netika dapat diperoleh melalui analisis kladistika (Eldenas & Linder 2000). Asumsi dasar yang digunakan adalah semua anggota kelompok yang memiliki sejarah evolusi yang sama memiliki hubungan lebih dekat satu sama lain dibandingkan dengan anggota kelompok lainnya. Hubungan antara kelompok organisme tersebut dikenali
me-lalui seperangkat karakter unik apomorf dan
sinapomorf (NCBI 2003).
Penelitian ini bertujuan mengungkap hu-bungan kekerabatan filogenetika pisang di Indone-sia berdasarkan karakter morfologi. Kekayaan plasma nutfah yang dilengkapi informasi filoge-netika akan memudahkan peneliti dalam meman-faatkan plasma nutfah tersebut khususnya dalam program pemuliaan pisang.
METODE PENELITIAN Bahan Penelitian
Tanaman yang diteliti adalah aksesi pisang koleksi Dinas Pertanian dan Kehewanan (Diperta)
Yogyakarta dan Balai Penelitian Buah (Balitbu) Tropika, Solok, Sumatera Barat. Karakterisasi morfologi dilakukan pada tanaman dewasa pada bagian vegetatif (batang semu, daun dan anakan) dan bagian generatif (bunga dan buah). Total
ak-sesi ingroup yang digunakan sebanyak 36 aksesi
yang terdiri atas 9 aksesi kultivar pisang grup ge-nom AA, 9 aksesi AAA, 8 aksesi AAB, 8 aksesi
ABB, dan 2 aksesi BB dan aksesi outgroup terdiri
atas 2 jenis pisang liar (Tabel 1). Analisis Data
Hasil skoring karakter morfologi dianalisis menggunakan PAUP versi 4.0 untuk membangun pohon filogenetika. Topologi pohon diuji statistika
menggunakan bootstrap 100 kali.
HASIL DAN PEMBAHASAN
Analisis hubungan kekerabatan filogenetika
berdasarkan karakter morfologi menghasilkan
po-hon yang parsimoni dengan tree length 378
(Gambar 1). Keseluruhan karakter yang diamati (41 karakter) merupakan karakter parsimoni
informatif. Consistensi index (CI), retention index
(RI), dan homoplasi index (HI), berturut-turut
sebesar 0.275, 0.487, dan 0.725. Hasil analisis ini
menunjukkan bahwa semua aksesi ingroup
ter-dapat dalam klad-klad yang memiliki kekokohan
rendah (bootstrap kurang dari 50%) kecuali klad
‘Klutuk Wulung’ (BB) dan ‘Klutuk’ (BB) dengan
bootsrap 98% dan klad yang terdiri atas ‘Klutuk
Wulung’ (BB), ‘Klutuk’ (BB), dan ‘Boi’ (ABB)
dengan bootstrap 67%.
Karakter morfologi yang digunakan dalam analisis filogeni ini memiliki homoplasi paralel. Hal ini terjadi antara lain karena perubahan ciri morfogi yang sama juga terjadi pada percabangan klad yang lain. Peristiwa homoplasi dapat dilihat melalui nilai HI yang tinggi yaitu 0.725 atau CI yang rendah yaitu 0.275. Kladogram dengan nilai CI sama dengan satu (CI=1) menunjukkan bahwa karakter yang digunakan dalam analisis filogeni muncul hanya satu kali (tidak terjadi homoplasi). Objek penelitian ini adalah takson yang rendah
yaitu tingkat jenis dan di bawah jenis (Musa spp.)
sehingga perubahan karakter morfologi yang terjadi pada satu aksesi dengan aksesi yang lain relatif sama. Karakter morfologi tidak cukup me-nyediakan sejumlah karakter yang memiliki sifat konvergenitas tinggi (Hillis et al. 1996).
Aksesi ‘Boi’ (ABB), ‘Klutuk Wulung’ (BB)
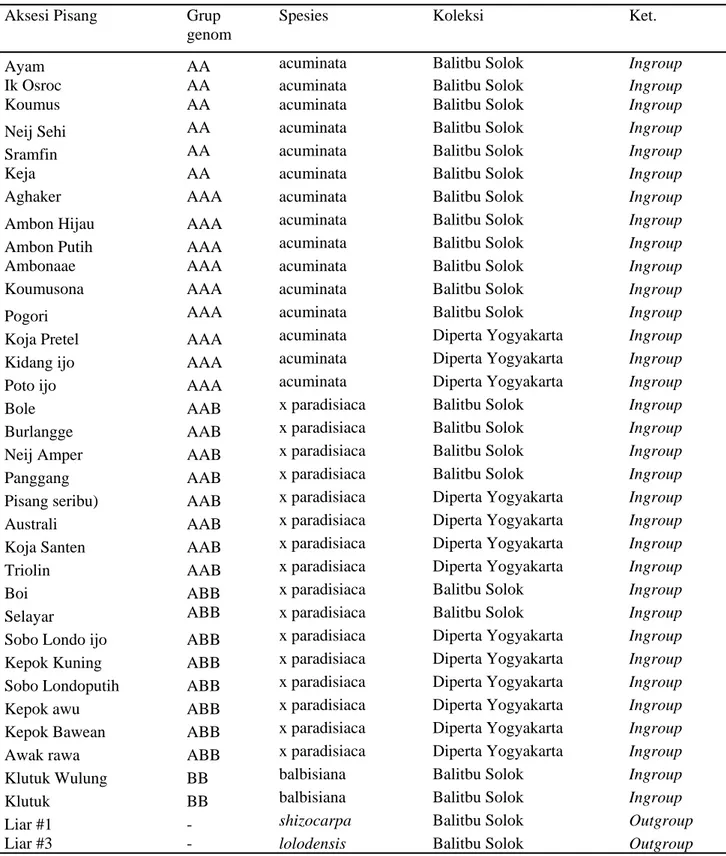
Tabel 1.Aksesi pisang yang digunakan dalam analisis filogenetika berdasarkan karakter morfologi
Aksesi Pisang Grup
genom
Spesies Koleksi Ket.
Ayam AA acuminata Balitbu Solok Ingroup
Ik Osroc AA acuminata Balitbu Solok Ingroup
Koumus AA acuminata Balitbu Solok Ingroup
Neij Sehi AA acuminata Balitbu Solok Ingroup
Sramfin AA acuminata Balitbu Solok Ingroup
Keja AA acuminata Balitbu Solok Ingroup
Aghaker AAA acuminata Balitbu Solok Ingroup
Ambon Hijau AAA acuminata Balitbu Solok Ingroup
Ambon Putih AAA acuminata Balitbu Solok Ingroup
Ambonaae AAA acuminata Balitbu Solok Ingroup
Koumusona AAA acuminata Balitbu Solok Ingroup
Pogori AAA acuminata Balitbu Solok Ingroup
Koja Pretel AAA acuminata Diperta Yogyakarta Ingroup
Kidang ijo AAA acuminata Diperta Yogyakarta Ingroup
Poto ijo AAA acuminata Diperta Yogyakarta Ingroup
Bole AAB x paradisiaca Balitbu Solok Ingroup
Burlangge AAB x paradisiaca Balitbu Solok Ingroup
Neij Amper AAB x paradisiaca Balitbu Solok Ingroup
Panggang AAB x paradisiaca Balitbu Solok Ingroup
Pisang seribu) AAB x paradisiaca Diperta Yogyakarta Ingroup
Australi AAB x paradisiaca Diperta Yogyakarta Ingroup
Koja Santen AAB x paradisiaca Diperta Yogyakarta Ingroup
Triolin AAB x paradisiaca Diperta Yogyakarta Ingroup
Boi ABB x paradisiaca Balitbu Solok Ingroup
Selayar ABB x paradisiaca Balitbu Solok Ingroup
Sobo Londo ijo ABB x paradisiaca Diperta Yogyakarta Ingroup
Kepok Kuning ABB x paradisiaca Diperta Yogyakarta Ingroup
Sobo Londoputih ABB x paradisiaca Diperta Yogyakarta Ingroup
Kepok awu ABB x paradisiaca Diperta Yogyakarta Ingroup
Kepok Bawean ABB x paradisiaca Diperta Yogyakarta Ingroup
Awak rawa ABB x paradisiaca Diperta Yogyakarta Ingroup
Klutuk Wulung BB balbisiana Balitbu Solok Ingroup
Klutuk BB balbisiana Balitbu Solok Ingroup
Liar #1 - shizocarpa Balitbu Solok Outgroup
Liar #3 - lolodensis Balitbu Solok Outgroup
terpisah dari aksesi yang lain karena memiliki beberapa karakter morfologi yang sangat berbeda.
Hal ini ditunjukkan melalui nilai bootstrap 67%
untuk klad ‘Boi’, ‘Klutuk Wulung’ dan ‘Klutuk’ dan 98% untuk klad ‘Klutuk Wulung’ dan ‘Klutuk’. Berdasarkan skoring 15 karakter
morfo-logi (Simmonds & Sheperd 1955), grup genom BB
dikelompokkan ke dalam jenis M. balbisiana. Jenis
pisang ini memiliki karakter yang berbeda secara
ekstrim dengan M. acuminata (AA). Setiap
ka-rakter M. balbisiana liar diberi nilai 5 dan M.
Gambar 1. Pohon filogeni yang diperoleh melalui analisis kladistik. Liar#1 Liar#3 KejaAA KoumusAA BoleAAA Selayar ABB KoumosonaABB BoiBBB KlutukWulungBB KlutukBB NeijAmperAAB PanggangAAB BuranggeAAB IkOsrocAAA NeijSehiAA SramfinAA SoboLondopthABB AghakerAAA AmbonPutihAAA AmbonHijauAAA AmbonaaeAAA PogoriAAA Mas40hariAA KidangijoAAA KojaPretelAAA AustraliAAB MonyetAA RejangAA PotoijoAAA KepokKuningABB SoboLondoijoABB KepokBaweanABB AwakrawaABB TriolinAAB KepokawuABB RajaSeribuAAB KojaSantenAAB AyamAA
dan ’Klutuk’ memiliki hubungan kekerabatan yang
sangat dekat dengan tipe liarnya, yaitu M.
balbisiana (BB). Karakter morfologi kultivar ’Boi’
mirip dengan kelompok Klutuk sehingga dima-sukkan grup genom BBB (INIBAP 2002). Hasil penelitian ini menunjukkan bahwa kultivar ’Boi’
tidak terdapat dalam klad dengan nilai bootstrap
yang sama seperti kelompok Klutuk. Hal ini membuktikan bahwa aksesi ’Boi’ tidak termasuk grup genom BBB. Hasil analisis filogeni ber-dasarkan karakter morfologi ini memperkuat hasil
penelitian Pillay et al. (2004) yang
memper-lihatkan bahwa BBB sangat jarang ditemukan secara alami. Kekeliruan penempatan ’Boi’ ke da-lam grup genom BBB juga dibuktikan melalui kajian analisis mikrosatelit. Berdasarkan lokus mi-krosatelit MaCIR108 aksesi tersebut dimasukkan ke dalam grup genom ABB bukan BBB karena memiliki satu alel A dan dua alel B (Retnoningsih & Karno 2007).
Meskipun hanya memiliki nilai bootstrap
yang rendah (kurang dari 50%) dapat dilihat bahwa grup genom yang sama memiliki kecen-derungan berada pada klad yang sama. Kelompok triploid AAA seperti ‘Aghaker’, ’Ambon Putih’, ’Ambon Hijau’ dan ’Ambonaee’ terdapat dalam satu klad karena secara morfologi keempat aksesi tersebut sangat mirip satu sama lain sehingga diduga memiliki sejarah evolusi yang sama (Valmayor et al. 2002). Aksesi tersebut
kemung-kinan berasal dari tetua liar subspesies Musa
acuminata yang sama. Klad yang terdiri atas
‘Kidang Ijo’ (AAA), ’Koja Pretel’ (AAA), ’Austoli’ (AAB), ’Monyet’ (AA) dan ’Rejang’ (AA) kemungkinan berasal dari tetua liar
yang memiliki kekerabatan lebih dekat dengan M.
acuminata subsp. zebrina. Menurut Nasution &
Yamada (2001), kultivar ’Monyet’ dikelompokkan
ke dalam subspesies zebrina.
Sebagian besar aksesi hibrid hasil
persi-langan M. acuminata dan M. balbisiana (M x
paradisiaca ABB) seperti ’Kepok Kuning’, ’Sobo
Londoijo’, ’Kepok Bawean’, ’Awak Rawa’, dan ’Kepok Awu’ terdapat dalam satu klad. Dalam klad ini juga ditemukan aksesi AAB yaitu ’Triolin’. Menurut Jumari & Pudjoarinto (2000), aksesi ‘Triolin’ berdasarkan karakter morfologi dikelompokkan ke dalam grup genom AB. Berda-sarkan karakter ini, ’Triolin’ memperlihatkan karakter transisi antara grup kultivar AA, AAA dan AAB dengan grup kultivar BB, ABB, dan ABBB. Grup genom AB secara alami jarang ditemukan (Valmayor et al. 2000) sehingga
eksis-tensi hibrid AB khususnya di Indonesia masih dipertanyakan. Hasil kajian mikrosatelit menun-jukkan bahwa ’Triolin’ memiliki dua alel A dan satu alel B sehingga aksesi tersebut seharusnya dikelompokkan ke dalam grup genom AAB (Retnoningsih & Karno 2007). Secara morfologi kekerabatan ‘Triolin’ lebih dekat dengan ke-lompok ABB mungkin karena kesamaan tetua liar
M. balbisiana sehingga dalam evolusinya berada
dalam klad grup genom ABB.
Jumlah dan jenis karakter morfologi yang digunakan dalam analisis filogeni ini sangat ter-batas karena banyak karakter yang tidak dapat direkam pada saat pengumpulan data. Hal ini me-rupakan salah satu kelemahan karakter morfologi yang ketersediaannya tidak maksimal. Berbeda dengan karakter morfologi yang dapat dicatat hanya pada fase pertumbuhan tertentu, karakter molekuler dapat diperoleh kapan saja dan dari bagian mana saja (Guzow-Krzeminska et al. 2001). Oleh karena itu, analisis filogenetika pisang menggunakan pendekatan molekuler akan mem-berikan dan menutupi keterbatasan karakter mor-fologi. Informasi yang disumbangkan kajian de-ngan pendekatan molekuler akan lebih berarti untuk mengungkap keanekaragaman kultivar pi-sang. Berdasarkan pendekatan molekuler, filogeni dan keanekaragaman pisang di Indonesia diha-rapkan lebih dapat dipahami karena ribuan basa nukleotida dapat diidentifikasi. Dalam hal ini jum-lah karakter yang dapat digunakan dalam análisis jauh lebih banyak dan hampir tidak terbatas (Vicente et al. 2005).
DAFTAR PUSTAKA
Eldenas PK & Linder HP. 2000. Congruence and complementarity of morphological and trnL-F sequence data and phylogeny of the Afri-can Restionaceae. Sys. Bot. 25(4): 692– 707.
Guzow-Krzeminska B, Gorniak M & Wegrzyn G. 2001. Molecular determination keys: con-struction of keys for species identification based on restriction fragment length
poly-morphism. Int. Arch. Biosci. p. 1057–1067.
Hillis MD, Moritz C & Mable BK.1996.
Molecu-lar Systematics. Second edition. Sinauer
Associates, Sunderland.
[INIBAP] International Network for the Improve-ment of Banana and Plantain. 2001. Banana diversity. http://www.inibap.org/. [12 Juli
2003]
[INIBAP] International Network for the Improve-ment of Banana and Plantain. 2002. The
exploration of Musaceae in Irian Jaya
(Papua). INIBAP.
[INIBAP] International Network for the Improve-ment of Banana and Plantain. 2003. Just how far are bananas from extinction?. http:// www.inibap.org/ [12 Juli 2003]
Jumari & Pudjoarinto A. 2000. Kekerabatan fenetik kultivar pisang di Jawa. Biologi 2(9): 531–542.
Megia R. 2005. Musa Sebagai Model Genom.
Hayati 12: 167–170.
Nasution RE & Yamada I. 2001. Pisang-pisang Liar di Indonesia. Bogor, Puslitbang Biologi-LIPI.
NCBI2003. Systematics and Molecular Phyloge-netics. http://www.ncbi.nlm. nih.gov/ Pillay M, Tenkouano A, Ude G & Ortiz R. 2004.
Molecular characterization on genomes in
Musa and its applications. In: Jain SM &
Swennen R (Eds), Banana Improvement:
Cellular, Molecular Biology, and Induced
Mutations. Science Publishers, Inc. Enfield
(NH), USA. Plymouth, UK. pp. 271–286. Retnoningsih A & Karno. 2007. Analisis variasi
mikrosatelit pisang bergenom AAB, ABB,
BB, dan BBB. Sainteknol. 4: 1–10.
Simmonds NW. 1962. The Evolution of the
Ba-nana. London: Longman Inc.
Simmonds NW & Sheperd K. 1955. The taxonomy
and origins of the cultivated bananas. J.
Linn. Soc. Lond. Bot. 55: 302–312.
Stover RH & Simmonds NW. 1987. Bananas.
New York. John Wiley and Sons.
Valmayor RV, Espino RRC & Pascua OC. 2002.
The Wild and Cultivated Bananas of the
Philippines. Los Banos. Foundation Inc.
Verheij EWM & Coronel RE. 1992. Edible Fruits
and Nuts. Prosea 2: 225–229.
Vicente MC de, Gusman FA, Engels J & Rao VR.
2005. Genetic characterization and its use
in decision making for the conservation of
crop germplasm. The role of