Soil Biology & Biochemistry 38 (2006) 1734–1744
Effects of tree harvesting, forest floor removal, and compaction on
soil microbial biomass, microbial respiration, and N availability
in a boreal aspen forest in British Columbia
Lucero Mariani
a,, Scott X. Chang
a, Richard Kabzems
b aDepartment of Renewable Resources, 442 Earth Sciences Building, University of Alberta, Edmonton, Alta., Canada T6G 2E3
b
Ministry of Forests, 9000–17th St., Dawson Creek, BC, Canada V1G 4A4
Received 30 May 2005; received in revised form 27 November 2005; accepted 29 November 2005 Available online 17 February 2006
Abstract
The effects of timber harvesting and the resultant soil disturbances (compaction and forest floor removal) on relative soil water content, microbial biomass C and N contents (Cmic and Nmic), microbial biomass C:N ratio (Cmic-to-Nmic), microbial respiration,
metabolic quotient (qCO2), and available N content in the forest floor and the uppermost mineral soil (0–3 cm) were assessed in a
long-term soil productivity (LTSP) site and adjacent mature forest stands in northeastern British Columbia (Canada). A combination of principal component analysis and redundancy analysis was used to test the effects of stem-only harvest, whole tree harvest plus forest floor removal, and soil compaction on the studied variables. Those properties in the forest floor were not affected by timber harvesting or soil compaction. In the mineral soil, compaction increased soil total C and N contents, relative water content, and Nmicby 45%, 40%,
34% and 72%, respectively, and decreased Cmic-to-Nmic ratio by 29%. However, these parameters were not affected by stem only
harvesting or whole tree harvesting plus forest floor removal, contrasting the reduction of white spruce and aspen growth following forest floor removal and soil compaction reported in an earlier study. Those results suggest that at the study site the short-term effects of timber harvesting, forest floor removal, and soil compaction are rather complex and that microbial populations might not be affected by the perturbations in the same way as trees, at least not in the short term.
r2006 Elsevier Ltd. All rights reserved.
Keywords:Available N; Boreal forest; Long-term soil productivity (LTSP); Microbial respiration; Microbial biomass;qCO2; Redundancy analysis
1. Introduction
Forest harvesting has been reported to decrease soil evapotranspiration, increase soil temperature and its diurnal fluctuations, and create a large amount of debris and dead roots that are easily decomposed by soil biota (Greacen and Sands, 1980;Lenhard, 1986;Williamson and Neilsen, 2000). In addition, forest-harvesting machinery may cause soil compaction and uneven forest floor displacement or disturbance, and common site preparation practices such as harvest residue removal and forest floor
scalping can affect site organic matter (OM) content and soil porosity, the two ecosystem properties most likely to impact long-term soil productivity (Powers et al., 2005).
Assessing the effects of forest harvesting and site preparation practices on soil biological properties and processes is a crucial step towards the conservation of forest ecosystem functions and productivity (Li et al., 2004). Soil organic matter (SOM) content and quality, soil microbial biomass, microbial respiration, and N avail-ability have been used as indices to assess soil biological activity and health (Nambiar, 1996; Burger and Kelting,
1999;Schoenholtz et al., 2000). SOM content and quality
are generally considered as key indicators of forest soil quality and sustainability because SOM is linked to the fundamental attributes of the soil: water-holding capacity, nutrient availability, soil buffering capacity, rates of gas
www.elsevier.com/locate/soilbio
0038-0717/$ - see front matterr2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.soilbio.2005.11.029
Corresponding author. Present address: Muse´um National d’Histoire
Naturelle, 4 avenue du Petit Chaˆteau, 91 800 Brunoy, France. Tel.: +33 1 60 47 92 14; fax: +33 1 60 46 57 19.
exchange, and soil biological activities (Powers et al., 2005; Nambiar, 1996). Soil microbial biomass is a small but very active part of the SOM and has frequently been suggested as a sensitive indicator of change, whereas total SOM content appears to be quite resilient to soil disturbance (Johnson et al., 1991;Bauhus et al., 1998;Stone and Elioff,
1998; Piatek and Allen, 1999; Li et al., 2004). Microbial
respiration is a measure of soil biological activity that can be combined with microbial biomass to derive specific microbial respiration (qCO2) that can be used to assess the
efficiency of soil micro-organisms in utilizing C substrates (Insam, 1990;Pietikainen and Fritze, 1995;Bauhus et al., 1998). Being at the core of plant nutrition, soil N availability is also a critical index for evaluating the sustainability of soil productivity (Marshall, 2000;Li et al., 2003).
Reports on the impact of forest harvesting, soil compaction, and forest floor removal on biological proper-ties of forest soils are still scarce and some experiments were designed in such a way that compaction and forest floor removal treatments are confounded (i.e.,Dick et al.,
1988; Pietikainen and Fritze, 1995; Corns and Maynard,
1998). In addition, some published studies report contra-dictory results, such as those summarized inTable 1.Table 1indicates that stem only harvest and forest floor removal either cause no effect or decrease the parameters listed; while whole tree harvest and soil compaction can cause no effect, increase, or decrease the measured properties. A most recent publication reports that compaction alters soil
physical properties and tree growth but not the biological indices of soil health, indicating a strong resilience of the microbial communities (Shestak and Busse, 2005). While tree growth can be impaired by the decrease of large pores due to compaction, microbes were found to take advantage of the increased volume of small pores (Shestak and Busse, 2005). Forest floor removal could also alter mineral soil properties in a contrasting way for plants and microbes. Therefore, no definitive conclusions can be made regarding the impact of forest harvesting and soil disturbances on soil biological properties, particularly in the boreal region in North America.
The objective of this study was to investigate the effects of soil compaction and forest floor removal on microbial biomass C and N contents (Cmic and Nmic), microbial
respiration, and soil N availability in a long-term soil productivity (LTSP) experiment established in a represen-tative mesic aspen ecosystem in northeastern British Columbia, Canada. Under boreal aspen forests, SOM contents, total soil N and S contents, available N and P, and other nutrients are significantly higher in the forest floor and the first few centimetres of the mineral soil than in deeper horizons (Neville et al., 2002) and the highest concentration of roots are at the organic/mineral–soil interface (Strong and La Roi, 1985). We therefore sampled the forest floor and the uppermost mineral soil (0–3 cm) in order to study the most biologically active soil horizons. Forest floor removal and soil compaction on the studied site have been shown to reduce white spruce and aspen
Table 1
A summary of published studies on the effects of timber harvest, soil compaction, and forest floor removal on soil biological properties
Stem only harvest Whole tree harvest Soil compactiona Forest floor removala No effect Decrease No effect Increase Decrease No effect Increase Decrease No effect Decrease
Forest floor
Cmicb 9c
Resp. 9
qCO2 9
Corg 5 5 5 5 5; 12d
Navail 1; 9 1
Ntot 9 1 1 12
Surface mineral soil(0–15 cm)
Cmic 2; 4; 7; 10 6 4
Nmic 4 4; 7
Cmic:Nmic 7 7
Resp. 11 11; 10
Corg 5 5 5 2; 6 5; 12 8; 6
Navail 8 8 4; 6 6 8; 6 4
Ntot 8 8 6 2 8; 12 6
WC 8 8 4; 12; 3 6; 3 8 4
aAdditional effects over timber harvest.
bCmic, microbial biomass C; Nmic, microbial biomass N; Resp., microbial respiration;qCO2, specific microbial respiration; Corg, soil organic C; Navail, available N; Ntot, total soil N; WC, water content.
cReferences: (1)Corns and Maynard (1998), (2)Dick et al. (1988), (3)Gomez et al. (2002), (4)Jordan et al. (2000), (5)Laiho et al. (2003), (6)Li et al.
(2003), (7)Li et al. (2004), (8)Piatek and Allen (1999), (9)Pietikainen and Fritze (1995), (10)Shestak and Busse (2005), (11)Startsev et al. (1998), (12)
Stone and Elioff (1998). d
growth which will likely delay future stand development and reduce site productivity (Stone and Kabzems, 2002). However we expected contrasting results between tree growth and soil processes (Shestak and Busse, 2005). Consequently, we hypothesized that tree harvesting, soil compaction, and forest floor removal do not change mineral soil microbial biomass, microbial respiration and N availability, and that tree harvesting and soil compaction do not affect these properties in the forest floor.
2. Materials and methods
2.1. Study site
The research site is located about 40 km northeast of Dawson Creek (551580N, 1201280W; 720 masl) in the Peace
Forest District, in northeastern British Columbia. The site is representative of mesic aspen (Populus tremuloides
Michx.) ecosystems in the moist, warm subzone of the Boreal White and Black Spruce (BWBS) biogeoclimatic zone (DeLong et al., 1990, 1991). The site has a south aspect and a 4% slope gradient. The overstory tree species were dominated by aspen, with small amounts of white spruce (Picea glauca(Moench) Voss), cottonwood (Populus balsamifera L.) and lodgepole pine (Pinus contorta ex Loud. var. latifoliaEngelm.). Soils on the study site have 20–30 cm of silt loam veneer over clay loam and are dominated by Luvic Gleysols (Soil Classification Working Group, 1998). The pre-treatment forest floor thickness averaged 7.5 cm (Kabzems, unpublished data) and the humus form was classified as a Lamimoder (Fons et al.,
1998). The pre-treatment forest floor contained
1350 kg ha1
of total N (SE of the mean: 71 kg ha1
; Kabzems, unpublished data). Annual precipitation ranges between 395 and 660 mm, with 33–63% of this falls as snow, mean annual temperature is 1.11C, and mean frost free period is 150 days (DeLong et al., 1990).
2.2. Experimental design and sample collection
Forest floor and 0–3 cm mineral soil samples were collected with a Dutch auger in September 2002 from all three replications (replicated at the plot level) of four treatments of the LTSP experiment and adjacent mature forests. The sampled four treatments in the LTSP experiment were: (1) stem-only harvest (forest floor intact), no compaction (treatment code: NcFi), with two of the replications installed in 1995 and the other in 1999; (2) stem-only harvest (forest floor intact), heavy compaction (HcFi), with two of the replications installed in 1995 and the other in 1999; (3) whole-tree harvest plus forest floor removal, no compaction (NcFr), with one of the replica-tions installed in 1995 and the other two in 1998; and (4) whole-tree harvest plus forest floor removal, heavy compaction (HcFr). One of the HcFr replications was installed in 1995 and the other two in 1998. The treatment plots are randomly arranged. Due to the installation of the
experiment over a 4 year period (1995–1999), as it was impossible to have all the plots installed in one single year for an experiment of this size, there is no blocking in the design. The time since installation was used as a covariate in the analysis to remove it as a confounding factor as in
Kabzems and Haeussler (2005)andTan et al. (2005). It was
found that plots installed in 1995 did not differ from those installed in 1998 or 1999 for a wide range of soil properties measured (Kabzems and Haeussler, 2005;Tan et al., 2005). In the mature forest stands adjacent to the LTSP plots, three transects were randomly located and they represented the 3 replications of the mature forest treatment. In each plot or transect, we collected one composite sample that consisted of 18 sub-samples collected around 3 randomly selected trees (6 sub-samples around each tree). The samples were placed on ice in a cooler and transported back to the laboratory.
2.3. Laboratory analyses
Fresh soil samples were passed through a 4 mm sieve. Cmic and Nmic were measured on fresh samples by the
fumigation–extraction method (Brookes et al., 1985;Vance et al., 1987). Each sample (20 g fresh mineral soil and 5 g fresh forest floor sample) was fumigated in duplicates for 24 h with alcohol-free chloroform and then extracted in 60 ml 0.5 m K2SO4. Extracts of the duplicates were
combined prior to the measurement of their C and N concentrations. Total N concentrations in K2SO4extracts
was determined by Kjeldahl digestion (Voroney et al., 1993) followed by colorimetric determination of the NH4+–N (industrial method 98–70 W) concentration on a
Technicon Autoanalyzer II (Tarrytown, New York). Total soluble organic carbon concentration in extracts was determined with an ASTRO 2001 system 2 (Leagne City, Texas). Extractability factors of 0.38 for Cmic(Vance et al.,
1987) and 0.54 for Nmic(Brookes et al., 1985;Bauhus et al.,
1998) were used to calculate Cmic and Nmic, respectively.
Available N (Navail) was measured by determining the
NH4 +
and NO3 (industrial method 487–77 A)
concentra-tions in K2SO4extracts of the non-fumigated soil samples
colorimetrically on a Technicon Autoanalyzer II following the method inMaynard and Kalra (1993).
To measure microbial respiration, fresh soil samples were pre-incubated for 7 days in the dark at 251C in 1 L mason jars fitted with butyl rubber septa on the lids. For the mineral soil, 6–8.5 g dry soil equivalent (in triplicates) was used and for the forest floor samples 0.8–1.4 g dry mass equivalent (in triplicates) was used. After the pre-incuba-tion period, we ventilated the jars, closed the lids tightly and immediately measured the initial CO2concentration in
the jars. Concentrations of CO2in the headspace of each
concentration in the headspace again. The difference in CO2 concentrations between the initial and the
post-incubation measurements was used to calculate microbial respiration.
Fresh samples of mineral soil (20 g) and forest floor (5 g) were dried for 24 h at 1051C to determine soil moisture content gravimetrically. The relative soil water content was expressed as water content as a percent of water holding capacity. The water-holding capacity of the soils was determined by the wet funnel method (Jenkinson and Powlson, 1976). A sub-set of soil samples was air-dried and ground to pass a 40-mesh sieve. Total C and N contents (Ctot and Ntot) were determined by dry combustion
(Tabatabai and Bremner, 1991) with a Costech ECS 4010 elemental combustion analyzer (Milan, Italy). Carbonates are absent from the soil. The results for all the measure-ments were expressed on an oven-dry soil weight basis and converted to kg ha1
for each cm of depth of the material using soil bulk densities of forest floor and mineral soil. Data analyses were performed on the values expressed in kg ha1
cm1
.
2.4. Data analyses
2.4.1. Redundancy analysis (RDA)and permutation tests
The aim of the data analysis was to test the effects of merchantable bole harvest, forest floor removal, and soil compaction on the quantity and quality of SOM, including Ctotand Ntot, soil C:N ratio (Ctot-to-Ntot), Cmicand Nmic,
Cmic-to-Nmic ratio, Navail, rates of microbial respiration, qCO2, and relative water content.
We chose to run multivariate analyses because of the strong correlations between the variables. Analysis of the data by parametric MANOVA was not possible because of heterogeneity of within-group variance (tested by Bar-tlett’s, Log-ANOVA, Cochran’sC, and Box’sMunivariate tests) and a high ratio of the number of variables to the number of objects. In this paper, if the lowest variance is greater than 33% of the highest variance, within-group variance was considered to be homogeneous. Data transformations were not able to correct the heterogeneity of variance (heteroscedasticity).
RDA associated with permutation testing is an appro-priate non-parametrical alternative to MANOVA for ecological multivariate data sets (Verdonschot and ter
Braak, 1994; Legendre and Anderson, 1999; Anderson,
2001). We ran RDA on the correlation matrix (obtained from data centred and standardized) and calculated the multivariate within-group variances from the coordinate of the soil samples. Permutation testing is impaired by heteroscedasticity (Hayes, 1996; Legendre and Legendre, 1998). In case of heteroscedasticity, we have a two-part null hypothesis: the multivariate locations (means) do not differ and the within group variances do not differ. Conse-quently, a significant test might mean that the within group variances are different but that the means do not differ. In presence of heteroscedasticity, we used the ordination
biplots to check the cause of a significant test (M.J. Anderson pers. comm.).
We performed 3 sets of analyses: (1) for the 9 plots (including the three mature forest transects and six forest floor intact harvested plots) where both the forest floor and mineral soil layers were present, we compared the forest floor and the mineral soil by RDA with a dummy variable indicating the layer of the sample (LAYER) as an independent variable and dummy variables indicating if the sample has been harvested (HARVEST) or compacted (COMPACTION) as covariates. The 2 layers from the same plot, autocorrelated, were treated as repeated measures in the analysis. As we found a significant difference between the 2 layers, we performed the subsequent analyses for each layer separately; (2) to analyse the timber harvesting effect, we first reduced the number of biological variables by principal component analysis (PCA). Then we used RDA as a form of inverse analysis related to discriminant analysis (Legendre and
Legendre, 1998; Lepsˇ and Sˇmilauer, 2003) with the
resulting principal components (biological-PCs) as expla-natory variables and HARVEST as dependent variable; and (3) within the LTSP experiment, we tested the effect of COMPACTION and organic matter removal by RDA. The effect of organic matter removal was tested with OMR, a dummy variable indicating if the organic matter removal is low or severe. The time since installation was used as a covariate. These 3 analyses were performed separately for bulk density, because it is a parameter of a different kind. All these RDA were tested by 999 permutations under the reduced model.
As RDA is a constraint PCA, we then used PCA to evaluate the percentage of variation extracted by the RDA by comparing it to the variation extracted by the first and second principal components of the related PCA (Lepsˇ and Sˇmilauer, 2003). We regard these variables that have a correlation coefficient with the canonical axes greater than the mean of all the correlation coefficients as making meaningful contributions to the canonical axes (Heuer and Smalla, 1997).
2.4.2. Spearmann R correlation coefficients
Spearmann’s R correlation coefficients represent the proportion of common variation between the ranks of the 2 variables being compared. For the 10 variables under study, 45 simultaneous correlation tests were performed. We applied Hochberg’s method to correct for multiple testing (Hochberg, 1988;Legendre and Legendre, 1998).
2.4.3. Statistical software
The Bartlett’s, Log-ANOVA, Cochran’sC, and Box’sM
correlation was analysed with the STATISTICA package developed by StatSoft Inc. (Tulsa, OK, USA).
3. Results
3.1. Correlations between soil C and N contents and microbial properties
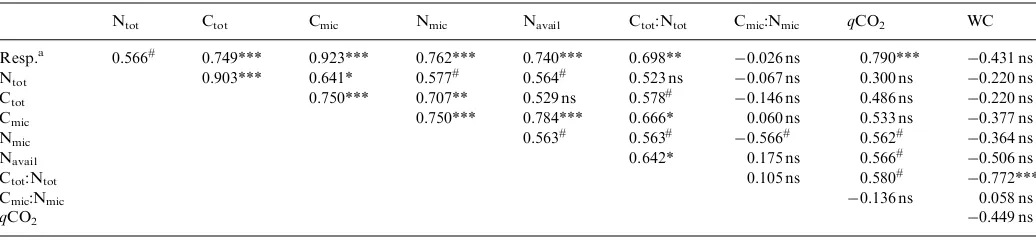
When all 24 data points (five treatments with three replications and one or two depths) were considered, soil microbial respiration, Ctot, Ntot, Ctot-to-Ntot ratio, Cmic,
Nmic, and Navailwere significantly and positively correlated
with each other, with 2 exceptions: between Navailand Ctot
and between Ctot-to-Ntotratio and Ntot(Table 2). Cmic
-to-Nmic ratio was not significantly correlated with any of the
other studied variables except for Nmic. Metabolic quotient
(qCO2) was significantly and positively correlated with
microbial respiration, Nmic and Navail, and Ctot-to-Ntot
ratio. Relative water content was significantly and nega-tively correlated only with Ctot-to-Ntotratio (Table 2).
3.2. Soil biological activities in forest floor vs. mineral soil and effects of timber harvesting
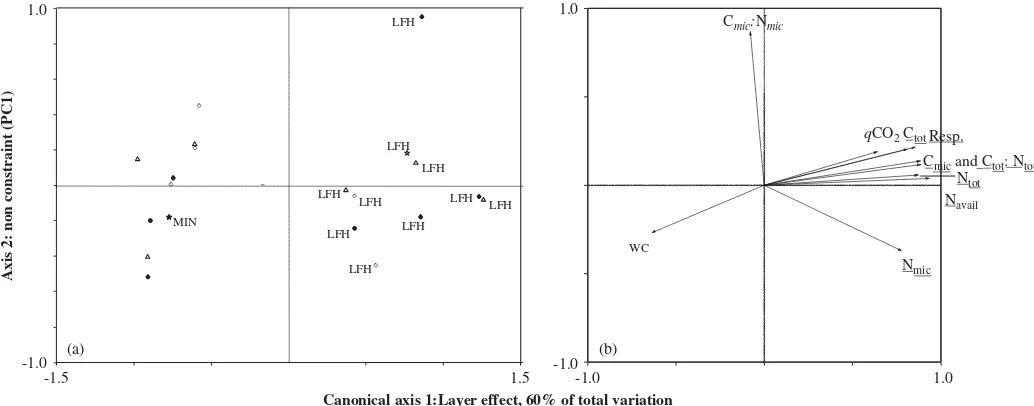
For the layer effect, the multivariate within-group variances were heterogeneous with a variance for the mineral samples reaching only 22% of the variance of the forest floor samples. However, the plot of the samples in the space of the dependant variables shows clearly that the 2 groups have different means (Fig. 1).
Reflecting the large differences between the forest floor and mineral soil in the 10 studied variables (Tables 3 and 4), the difference between forest floor and mineral soil was highly significant and explained 60% of the total variance of the 10 variables under study (F ¼26:8,P¼0:005; Fig. 1). This corresponds to 78% of the variance extracted by the first 2 PCs of the related PCA. All the variables contributed to the layer effect exceptqCO2, relative water
content, and Cmic-to-Nmicratio (Fig. 1). Ctot, Ntot, Navail,
microbial respiration and Cmic-to-Nmic and Ctot-to-Ntot
ratios were 2, 2, 4, 4, 3, 3 times and 30% greater, respectively, in the forest floor than in the mineral soil (Tables 3 and 4).
For the forest floor, no effect of harvesting was found for any of the studied variables (data not shown). The PCA of the 10 variables under study yielded biological-PCs that explained 31%, 20% and 19.5% of the total variation, for the first, second and third PC, respectively (data not shown). The discriminant analysis with these 3 biological-PCs as independent variables and HARVEST as depen-dant variable explained 41% of the variability of two grouping variables and was not significant (F ¼3:5,
P¼0:502; data not shown). In the ordination plot, the samples from the mature forest were mixed with the heavy compaction-forest floor intact plots (graph not shown).
For the mineral soil, no effect of harvesting was found either (Table 4). The PCA of the 10 variables under study yielded PCs that explained 37%, 24%, 14% and 12% of the total variation, for the first, second, third and fourth biological-PC, respectively (data not shown). The discri-minant analysis with these 4 biological-PCs as the independent variable and HARVEST, as dependant variable explained 43% of the variability of the two grouping variables and was not significant (F ¼1:9,
P¼0:175; data not shown). In the ordination plot, the samples from the mature forest were mixed with all the other groups of samples (data not shown).
3.3. Effects of compaction and forest floor removal in the LTSP experiment
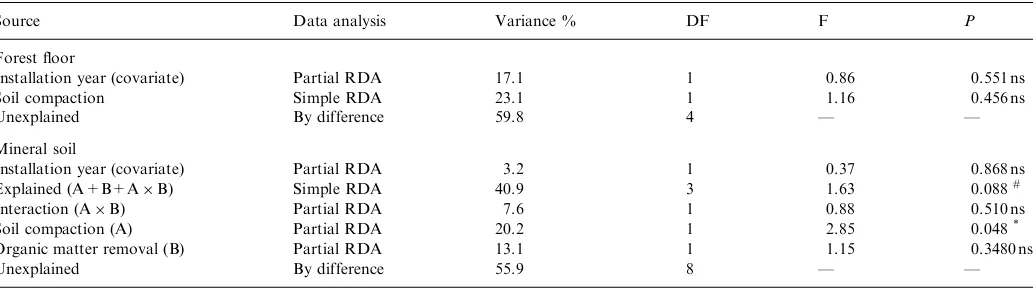
The time since installation had no effect on the 10 variables under study in the forest floor and the mineral soil: it explained 17% and 3% of the total variations for the forest floor and the mineral soil, respectively (Table 5).
For the forest floor, the multivariate analysis did not detect a significant effect of the compaction treatment or year since installation on the 10 variables under study (Table 5). The proportion of the variance explained by soil compaction (23%) was less than half of the fraction
Table 2
SpearmannRrank correlation coefficients among the 10 studied variables for two depths, including both the mature forest and LTSP plots (n¼24)
Ntot Ctot Cmic Nmic Navail Ctot:Ntot Cmic:Nmic qCO2 WC
Resp.a 0.566# 0.749*** 0.923*** 0.762*** 0.740*** 0.698** 0.026 ns 0.790*** 0.431 ns
Ntot 0.903*** 0.641* 0.577# 0.564# 0.523 ns 0.067 ns 0.300 ns 0.220 ns
Ctot 0.750*** 0.707** 0.529 ns 0.578# 0.146 ns 0.486 ns 0.220 ns
Cmic 0.750*** 0.784*** 0.666* 0.060 ns 0.533 ns 0.377 ns
Nmic 0.563# 0.563# 0.566# 0.562# 0.364 ns
Navail 0.642* 0.175 ns 0.566#
0.506 ns
Ctot:Ntot 0.105 ns 0.580#
0.772***
Cmic:Nmic 0.136 ns 0.058 ns
qCO2 0.449 ns
#
Test significant at 10%; *test significant at 5%; **test significant at 1%; ***test significant atp0.5%; ns, test non-significant at 10%. a
extracted by the first 2 PCs of the related PCA (64%, data not shown). Therefore, we did not look further at the forest floor results.
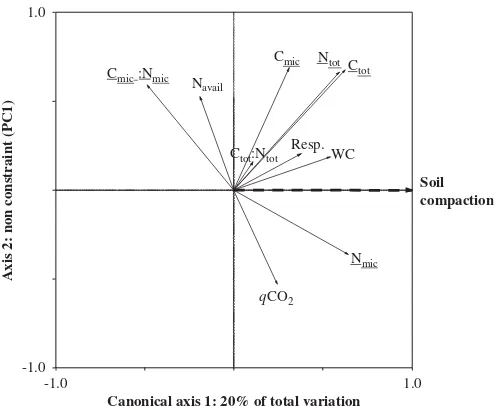
In contrast, for the mineral soil, the full model (Soil compaction+OM removal+Interaction) significantly
ex-plained 41% of the variability of the 10 variables under study in the experimental plots, after removing the effect of the covariate (Table 5). This is a substantial fraction (63%) of the total variability (66%) extracted by the first 2 PCs of the related PCA (data not shown). Soil compaction was the only factor significant in the RDA, explaining 20% of the total variability of the data (Table 5).
The mineral soil had higher Ctot, Ntot, Nmic, and water
content, but a lower Cmic-to-Nmic ratio in the compacted
than in the non-compacted plots; CmicandqCO2were only
slightly higher in the compacted plots and did not contribute meaningfully to the treatment effect (only 10% of the total variability for Cmic and 6% for qCO2
were explained by the compaction factor; data not shown); Navailand Ctot-to-Ntotratio were not affected by
compac-tion (Fig. 2,Table 4).
3.4. Bulk density
For bulk density, the only significant difference was between the forest floor and the uppermost mineral soil (91% of the total variability explained by the layer effect,
F ¼145, P¼0:005). For the uppermost mineral soil, the tests were non-significant, with 3% (F ¼0:4, P¼0:533) and 24% (F ¼2:2,P¼0:578), respectively, of variability explained by harvesting, and the treatments in the LTSP experiment, i.e., compaction+forest floor removal+inter-action term). For the forest floor, similarly, the effects of harvesting and compaction were non-significant, with 39% (F ¼11:2, P¼0:107) and 21% (F¼1:8, P¼0:236) of variability explained, respectively.
Canonical axis 1:Layer effect, 60% of total variation
Axis 2: non constraint (PC1)
-1.0
-1.0 -1.0
1.0 1.0
1.0
-1.5 1.5
MIN
LFH LFH
LFH LFH
LFH
LFH
LFH LFH LFH
LFH
Ntot
Nmic Navail
WC
Cmic:Nmic
qCO2 Ctot
Cmic and Ctot: Ntot
(a) (b)
Resp.
Fig. 1. Differences between forest floor and mineral soil layers for the 9 plots where the 2 layers were present (RDA on raw data centred and standardized). (a) RDA ordination plot representing the soil samples (coordinates are linear combination of the dependent variables). (b) RDA ordination plot. Variables underlined contributed more than the mean to canonical axis 1. The canonical analysis yielded only one canonical axis because there is only one independent variable.
Table 3
Means and coefficients of variation in percent (in parentheses) of the measured parameters in the forest floor (n¼3)
Measurement MFa NcFi HcFi
Bulk density (g cm3
) 0.24 (17) 0.16 (15) 0.22 (21) Ctotb(g kg1
) 366 (8) 403 (2) 367 (6)
(kg ha1
) 4815 (21) 3424 (12) 4278 (21) Ntot(g kg1
) 17.0 (7) 19.3 (4) 17.6 (6) (kg ha1
) 220 (17) 165 (15) 205 (20) Ctot:Ntot 21.6 (4) 20.9 (5) 20.8 (2) Navail(mg kg1
) 61 (9) 70 (8) 50 (7)
(kg ha1
) 8 (6) 6 (10) 6 (21)
WC (as a % of WHC) 25 (2) 26 (2) 26 (5) Resp. (mg CO2–C kg1d1) 451 (7) 374 (3) 505 (8)
(kg CO2ha1d1) 58 (15) 32 (16) 59 (23)
qCO2(g CO2–C kg1Cmicd1) 40 (4) 30 (5) 48 (4) Cmic(mg kg1) 9639 (13) 12074 (5) 13057 (10)
(kg ha1) 1460 (14) 1085 (12) 1243 (24) Nmic(mg kg1) 896 (10) 1300 (15) 1128 (21) (kg ha1) 118 (23) 109 (13) 128 (28) Cmic:Nmic 12.8 (8) 10.1 (10) 11.1 (38)
a
MF, mature forest; NcFi, no compaction with forest floor intact; HcFi, heavy compaction with forest floor intact.
b
4. Discussion
4.1. Forest floor and mineral soil properties
Cmicand Nmiccontents, Cmic-to-Nmic, Cmic-to-Corg(total
soil organic C content) and Nmic-to-Ntot ratios, and
microbial specific respiration rates (qCO2) were all within
the range reported for boreal forest soils (Smolander et al.,
1994; Pietikainen and Fritze, 1995; Scheu and Parkinson,
1995;Bauhus et al., 1998;Startsev et al., 1998). Navailwas
also within the range of reported values for forest soils
(Corns and Maynard, 1998;Maynard and MacIsaac, 1998; Li et al., 2003), with extractable N values averaging around 0.3% and 0.2% of Ntot in the forest floor and the
uppermost mineral soil, respectively. The Cmic-to-Corg
ratios averaged 3.%1 and 3.2% and the Nmic-to-Ntotratios
averaged 6.2% and 4.7% in the forest floor and the mineral soil, respectively.
The fact that Cmic-to-Nmicratio was not correlated with
any of the other variables measured except Nmic implies
that there was no causal link between bacteria:fungi ratio and indices of microbial activity, OM characteristics and N
Table 4
Means and coefficients of variation in percent (in parentheses) of the measured parameters in the 0–3 cm deep mineral soil (n¼3)
Measurement MFa NcFr NcFi HcFr HcFi
Bulk density (g cm3
) 0.83 (11) 0.98 (3) 0.82 (5) 0.85 (11) 0.86 (15)
Ctotb(g kg1
) 43.8 (21) 26.4 (21) 35.7 (5) 49.6 (26) 41.1 (8)
(kg ha1) 1044 (10) 770 (20) 874 (0) 1289 (21) 1091 (5)
Ntot(g kg1) 2.8 (25) 1.8 (19) 2.2 (12) 3.1 (22) 2.6 (10)
(kg ha1) 66 (15) 54 (19) 53 (8) 81 (17) 70 (6)
Ctot:Ntot 16.0 (7) 14.3 (8) 16.6 (8) 15.9 (5) 15.7 (1)
Navail(mg kg1) 5 (19) 3 (4) 4 (16) 4 (11) 4 (13)
(kg ha1) 1 (7) 1 (4) 1 (12) 1 (6) 1 (7)
WC (as a % of WHC) 37 (12) 55 (6) 41 (2) 36 (8) 28 (12)
Resp. (mg CO2–C kg1d1) 33 (1) 29 (7) 23 (21) 33 (10) 31 (2)
(kg ha1d1) 8 (6) 8 (6) 6 (17) 9 (7) 8 (14)
qCO2(g CO2–C kg1 Cmicd1
) 33 (20) 25 (14) 21 (3) 24 (4) 26 (6)
Cmic(mg kg1
) 1207 (21) 896 (13) 784 (23) 2025 (15) 1364 (5)
(kg ha1
) 262 (14) 327 (10) 268 (12) 372 (15) 305 (10)
Nmic(mg kg1
) 84 (27) 94 (16) 80 (20) 147 (27) 176 (26)
(kg ha1
) 20 (26) 27 (17) 19 (16) 40 (29) 41 (11)
Cmic:Nmic 13.9 (16) 12.3 (14) 14.3 (8) 11.2 (33) 7.8 (18)
aNcFi, no compaction with forest floor intact; HcFi, heavy compaction with forest floor intact; NcFr, no compaction with forest floor removed; HcFr, heavy compaction with forest floor removed.
bCmic, microbial biomass C; Nmic, microbial biomass N; Cmic:Nmic, microbial biomass C:N ratio; Ctot, total soil C; Ntot, total soil N; Ctot:Ntot, soil C:N ratio; Navail, available N; Resp., microbial respiration;qCO2, specific microbial respiration; WC, water content; WHC, water-holding capacity.
Table 5
Variance decomposition of the effect of the treatments and the covariate on the measured variables in the forest floor and mineral soil layers. RDA on raw data centred and standardized (multivariate analyses)
Source Data analysis Variance % DF F P
Forest floor
Installation year (covariate) Partial RDA 17.1 1 0.86 0.551 ns
Soil compaction Simple RDA 23.1 1 1.16 0.456 ns
Unexplained By difference 59.8 4 — —
Mineral soil
Installation year (covariate) Partial RDA 3.2 1 0.37 0.868 ns
Explained (A+B+AB) Simple RDA 40.9 3 1.63 0.088#
Interaction (AB) Partial RDA 7.6 1 0.88 0.510 ns
Soil compaction (A) Partial RDA 20.2 1 2.85 0.048*
Organic matter removal (B) Partial RDA 13.1 1 1.15 0.3480 ns
Unexplained By difference 55.9 8 — —
availability. Li et al. (2004) also found that Cmic-to-Nmic
ratio was not correlated with Ctot, Ntot, or Ctot-to-Ntot
ratio, whileSrivastava and Singh (1991)reported that Cmic
-to-Nmicratio was only negatively correlated with mineral N
content. On the contrary, the significant correlations between microbial respiration, Ctot, Ntot, Ctot-to-Ntotratio,
Cmic, Nmic, and Navail reflect that microbial activity, OM
quantity and quality, and available N are linked in the soil nutrient cycle.
The forest floor has a microbial population that was 50% less efficient than the mineral soil (as theqCO2data
shows), perhaps as a consequence of the abundance of food that decreased competition among microbial populations in the forest floor, allowing microbes to be less efficient. It is to be noted however that the differences in microbial efficiency did not contribute meaningfully to the difference between the forest floor and mineral soil layers in the multivariate analysis. In addition, the similarity in Cmic
-to-Nmic ratio between the forest floor and the mineral soil
suggests that the fungi:bacteria ratio (Marumoto et al., 1982) were very similar in those two soil material types.
Even though the plots were installed over a 4-year period, the plots installed in 1995 did not differ from these installed in 1998 or 1999 for the 10 chemical variables measured in this study. At the study site, others reported as well no effect of the age of the plots, on numerous parameters, i.e., bulk density, nitrogen mineralization and nitrification of 0–10 cm mineral soil and forest floor, air porosity of the 0–2 cm mineral soil, tree growth and tree regeneration (Kabzems and Haeussler, 2005; Tan et al., 2005).
4.2. Effects of forest harvesting and soil compaction
We found no significant effects of forest harvesting on the studied parameters in the forest floor and the surface mineral soil 3–7 years after the treatments were applied. Harvesting was conducted in winter on frozen ground, which is reported to have little effect on soil physical and biological properties within one (Bock and Van Rees, 2002) or 3–5 years after harvesting (Stone and Elioff, 1998;Block et al., 2002). Harvesting clearly has been shown to have immediate impacts on water evaporation at the soil surface, soil temperature and its diurnal fluctuations, and the size of easily decomposable OM pool (Greacen and
Sands, 1980; Lenhard, 1986; Williamson and Neilsen,
2000). However, our results suggest that 3–7 years after harvesting various impacts on the soil ecosystem either compensated each other, subsided, or were not yet notice-able on the measured parameters.
Severe soil compaction had no effect on any of the 10 measured chemical properties in the forest floor. In addition, the bulk density of the forest floor was the same in the LTSP experiment and the mature forest. These results probably indicate that physical and biological properties of the forest floor recovered from effects of compaction 3–7 years after the treatment.
In the surface mineral soil, heavy compaction increased Ctotand Ntotby 45% and 40%, Nmicby 72% and relative
water content by 34% and decreased Cmic-to-Nmicratio by
29%. Changes in both the plant community composition and in soil physical properties may be contributing to these responses. The rhizomatous grassCalamagrostis canadensis
expanded to 4–6 times its original cover on compacted plots where the forest floor was retained (Haeussler and Kabzems, 2005). Severe compaction decreased significantly the aeration porosity in the 0–2 cm deep mineral soil at the study site (Kabzems and Haeussler, 2005) which reflect the increase of volume of small pores at the expense of large pores (Greacen and Sands, 1980;Shestak and Busse, 2005). Thus, increases in soil C and N contents may be due to the increase of small pores that protect the OM and bacteria from protozoa grazing (Hassink et al., 1993; Shestak and Busse, 2005). This should have reduced microbial activities and hence the rates of mineralization and microbial respiration (Elliott et al., 1980). The increased microbial respiration rates were probably an artefact due to sieving that disturbed the soil and caused more soil C to become accessible to micro-organisms.
4.3. Effects of forest floor removal on mineral soil properties
Forest floor removal caused no significant effects on any of the parameters measured in this study. A detailed survey of the literature shows that forest floor removal has been reported to cause no effect as frequently as negative effects on these parameters (Table 1). In contrast, SOM quantity and soil microbial activity of the surface mineral soil were increased by compaction but were not affected by scalping. Ctot
Ntot Cmic
Nmic Navail
Ctot:Ntot Cmic :Nmic
qCO2
Soil compaction
Canonical axis 1: 20% of total variation
Axis 2: non constraint (PC1)
-1.0 1.0
-1.0 1.0
Resp. WC
Moreover, stem only harvesting did not affect SOM quantity and soil microbial activity. In contrast with these findings, forest floor removal and soil compaction on the studied site have been shown to reduce white spruce and aspen growth which will likely delay future stand develop-ment and reduce site productivity (Stone and Kabzems,
2002;Kabzems and Haeussler, 2005).
Differences between plant communities in the different treatment plots were clear (Haeussler and Kabzems, 2005). The forest floor removal plots had the greatest change in vegetation composition and structure, featuring a promi-nent moss layer and reduced abundance of herbs and shrubs. In contrast, Calamagrostis canadensis dominated heavily compacted plots where the forest floor had been retained (Haeussler and Kabzems, 2005). Competition between tree seedlings and early successional understory species is of main concern for forest regeneration (Steijlen
et al., 1995;Robinson et al., 2002;Hangs et al., 2003). Due
to the seasonal dynamic of N cycling (Li et al., 2003), a lack of synchronicity between soil N availability and the N needs of regenerating trees could also impact the relation-ships between the studied ecosystem properties.
Most importantly, large quantities of biomass and nutrients were exported in the forest floor removal plots, with the forest floor containing 1350 kg ha1
of N (Kabzems, pers. comm.). In the stem only harvested treatment 9% less wood biomass were exported as wood residues (Kabzems, pers. comm.). The forest floor in boreal forests is generally a thick layer of partially decomposed recalcitrant OM which immobilizes nutrients and its removal is expected to promote short-term site productiv-ity by changing the site’s biophysical conditions and encouraging decomposition but has the potential to compromise long-term site productivity (Prescott et al., 2000). At the study site, we observed little mixing of the forest floor with the surface mineral soil in the forest floor removal plots. We suggest that the effects of forest floor removal on the uppermost layer of mineral soil were partially compensated for by the changes in the composi-tion of the plant community which regenerated in the short period (3–7 years) after disturbance.
5. Conclusions
At 3–7 years after treatment application, biological activities, Ctotand Ntotin the uppermost mineral soil were
significantly increased by compaction but not affected by harvesting or forest floor removal; biological activities, Ctot
and Ntot in the forest floor were neither affected by
compaction nor by harvesting. Our results contrast with other studies that reported negative impacts of forest harvesting, soil compaction and forest floor removal on tree regeneration at the study site. Further studies are needed to determine if these contradictions resulted from an incomplete picture of the interactions between the soil environment, plant communities and tree species or if they describe the reality of soil microbial communities being
more resistant than trees to changes imposed by the treatments.
Acknowledgements
We thank Tony Hunt, Brian Farwell and Todd Thomson, foresters and coop student, respectively, British Columbia Ministry of Forests, for their help with field sampling, and Monica Molina and Miwa Matsushima for valuable technical help in the laboratory. We are also grateful to Drs. Pierre Legendre (Universite´ de Montre´al, Canada), Marti J. Anderson (University of Auckland, New Zealand), and Y. Feng (University of Alberta), for help with statistical analysis and to Dr. Robert F. Powers, PSW Research Station, USDA Forest Service, for providing information on LTSP experiments. Lucero Mariani was funded by a research grant and a postdoctoral fellowship from the Alberta Ingenuity Fund.
References
Anderson, M.J., 2001. A new method for non-parametric multivariate analysis of variance. Austral Ecology 26, 32–46.
Bauhus, J., Pare, D., Cote, L., 1998. Effects of tree species, stand age and soil type on soil microbial biomass and its activity in a southern boreal forest. Soil Biology and Biochemistry 30, 1077–1089.
Block, R., Van Rees, K.C.J., Pennock, D.J., 2002. Quantifying harvesting impacts using soil compaction and disturbance regimes at a landscape scale. Soil Science Society of America Journal 66, 1669–1676. Bock, M.D., Van Rees, K.C.J., 2002. Forest harvesting impacts on soil
properties and vegetation communities in the Northwest Territories. Canadian Journal of Forest Research 32, 713–724.
Brookes, P.C., Landman, A., Pruden, G., Jenkinson, D.S., 1985. Chloroform fumigation and the release of soil-nitrogen—a rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biology and Biochemistry 17, 837–842.
Burger, J.A., Kelting, D.L., 1999. Using soil quality indicators to assess forest stand management. Forest Ecology and Management 122, 155–166.
Corns, I.G.W., Maynard, D.G., 1998. Effects of soil compaction and chipped aspen residue on aspen regeneration and soil nutrients. Canadian Journal of Soil Science 78, 85–92.
DeLong, C., MacKinnon, A., Jang, L., 1990. A field guide for identification and interpretation of ecosystems of the northeast portion of the Prince George forest region. Land Management Handbook, vol. 22. B.C. Ministry of Forests, Victoria, BC.
DeLong, C., Annas, R.M., Stewart, A.C., 1991. Boreal white and black spruce zone. In: Meidinger, D., Pojar, J. (Eds.), Ecosystems of British Columbia. B.C. Ministry of Forests, Victoria, BC, pp. 237–250. Dick, R.P., Myrold, D.D., Kerle, E.A., 1988. Microbial biomass and soil
enzyme activities in compacted and rehabilitated skid trail soils. Soil Science Society of America Journal 52, 512–516.
Elliott, E.T., Anderson, R.V., Coleman, D.C., Cole, C.V., 1980. Habitable pore space and microbial trophic interactions. Oikos 35, 327–335. Fons, J., Kabzems, R.D., Klinka, K., 1998. Humus forms of trembling
aspen (Populus tremuloidesMichx) ecosystem in north-eastern British Columbia. Forest Ecology and Management 105, 241–250.
Gomez, A., Powers, R.F., Singer, M.J., Horwath, W.R., 2002. Soil compaction effects on growth of young Ponderosa pine following litter removal in California’s Sierra Nevada. Soil Science Society America Journal 66, 1334–1343.
Haeussler, S., Kabzems, R., 2005. Aspen plant community response to organic matter removal and soil compaction. Canadian Journal of Forest Research 35, 2030–2044.
Hangs, R.D., Knight, J.D., Rees, C.J.V., 2003. Nitrogen uptake characteristics for roots of conifer seedlings and common boreal forest competitor species. Canadian Journal of Forest Research 33, 156–163.
Hassink, J., Bouwan, L.A., Zwart, K.B., Bloem, J., Brussaard, L., 1993. Relationships between habitable pore space, soil biota and miner-alization in grassland soils. Soil Biology and Biochemistry 25, 47–55. Hayes, A.F., 1996. The permutation test is not distribution-free: testing
H0:r¼0. Psychological Methods 1, 184–198.
Heuer, H., Smalla, K., 1997. Evaluation of community-level catabolic profiles using BIOLOG GN microplates to study microbial community changes in potato phyllosphere. Journal of Microbial Methods 30, 49–61.
Hochberg, Y., 1988. A sharper Bonferroni procedure for multiple tests of significance. Biometrika 75, 800–802.
Insam, H., 1990. Are the soil microbial biomass and basal respiration governed by the climatic regime? Soil Biology and Biochemistry 22, 525–532.
Jenkinson, D.S., Powlson, D.S., 1976. The effects of biocidal treatments on metabolism in soil. V. A method for measuring soil biomass. Soil Biology and Biochemistry 8, 209–213.
Johnson, C.E., Johnson, A.H., Huntington, T.G., Siccama, T.G., 1991. Whole-tree clear-cutting effects on soil horizons and organic-matter pools. Soil Science Society of America Journal 55, 497–502. Jordan, D., Hubbard, V.C., Ponder Jr., F., Berry, E.C., 2000. The
influence of soil compaction and the removal of organic matter on two native earthworms and soil properties in an oak-hickory forest. Biology and Fertility of soils 31, 323–328.
Kabzems, R., Haeussler, S., 2005. Soil properties, aspen, and white spruce responses 5 years after organic matter removal and compaction treatments. Canadian Journal of Forest Research 35, 2045–2055. Laiho, R., Sanchez, F., Tiarks, A., Dougherty, P.M., Trettin, C.C., 2003.
Impacts of intensive forestry on early rotation trends in site carbon pools in the southeastern US. Forest Ecology and Management 174, 177–189.
Legendre, P., 2000. Homoge´ne´ite´ des variances—Guide. De´partement de sciences biologiques, Universite´ de Montre´al, 10 pp., available from
http://www.fas.umontreal.ca/BIOL/Casgrain/fr/labo/hv.html. Legendre, P., Anderson, M.J., 1999. Distance-based redundancy analysis:
testing multispecies responses in multifactorial ecological experiments. Ecological Monographs 69, 1–24.
Legendre, P., Legendre, L., 1998. Numerical ecology. Developments in Environmental Modeling, vol. 20, second English ed. Elsevier, Amsterdam, 853 pp.
Lenhard, R.J., 1986. Changes in void distribution and volume during compaction of a forest soil. Soil Science Society of America Journal 50, 462–464.
Lepsˇ, J., Sˇmilauer, P., 2003. Multivariate Analysis of Ecological Data using CANOCO. Cambridge University Press, Cambridge 269pp. Li, Q., Allen, H.L., Wilson, C.A., 2003. Nitrogen mineralization dynamics
following the establishment of a loblolly pine plantation. Canadian Journal of Forest Research 33, 364–374.
Li, Q., Allen, H.L., Wollum II, A.G., 2004. Microbial biomass and bacterial functional diversity in forest soils: effects of organic matter removal, compaction, and vegetation control. Soil Biology and Biochemistry 36, 571–579.
Marshall, V.G., 2000. Impacts of forest harvesting on biological processes in northern forest soils. Forest Ecology and Management 133, 43–60. Marumoto, T., Anderson, J.P.E., Domsch, K.H., 1982. Ecoposition of 14C- and 15N-labelled microbial cells in soil. Soil Biology and Biochemistry 14, 469–475.
Maynard, D.G., Kalra, Y.P., 1993. Nitrate and exchangeable ammonium nitrogen. In: Carter, M.R. (Ed.), Soil Sampling and Methods of Analysis. Canadian Society of Soil Science, Lewis Publishers, pp. 25–38.
Maynard, D.G., MacIsaac, D.A., 1998. Soil nutrient and vegetation response to patch clear-cutting of an aspen forest near Meadow Lake, Saskatchewan. Canadian Journal of Soil Science 78, 59–68.
Nambiar, E.K.S., 1996. Sustained productivity of forests is a continuing challenge to soil science. Soil Science Society of America Journal 60, 1629–1642.
Neville, J., Tessier, J.L., Morrison, I., Scarratt, J., Canning, B., Klironomos, J.N., 2002. Soil depth distribution of ecto- and arbuscular mycorrhizal fungi associated withPopulus tremuloideswithin a 3-year-old boreal forest clear-cut. Applied Soil Ecology 19, 209–216. Piatek, K.B., Allen, H.L., 1999. Nitrogen mineralization in a pine
plantation fifteen years after harvesting and site preparation. Soil Science Society of America Journal 63, 990–998.
Pietikainen, J., Fritze, H., 1995. Clear-cutting and prescribed burning in coniferous forest: comparison of effects on soil fungal and total microbial biomass, respiration activity and nitrification. Soil Biology and Biochemistry 27, 101–109.
Powers, R.F., Scott, D.A., Sanchez, F.G., Voldseth, R.A., Page-Dumroese, D., Elioff, J.D., Stone, D.M., 2005. The North American long-term soil productivity experiment: findings from the first decade of research. Forest Ecology and Management 220, 31–50.
Prescott, C.E., Maynard, D.G., Laiho, R., 2000. Humus in northern forests: friend or foe? Forest Ecology and Management 133, 23–26. Robinson, D.E., Wagner, R.G., Swanton, C.J., 2002. Effects of nitrogen
on the growth of jack pine competing with Canada blue-joint grass and large-leaved aster. Forest Ecology and Management 160, 233–242.
Scheu, S., Parkinson, D., 1995. Successional changes in microbial biomass, respiration and nutrient status during litter decomposition in an aspen and pine forest. Biology and Fertility of Soils 19, 327–332.
Schoenholtz, S.H., Van Miegroet, H., Burger, J.A., 2000. A review of chemical and physical properties as indicators of forest soil quality: challenges and opportunities. Forest Ecology and Management 138, 335–356.
Shestak, C.J., Busse, M.D., 2005. Compaction alters physical but not biological indices of soil health. Soil Science Society of America Journal 69, 236–246.
Smolander, A., Kurka, A., Kitunen, V., Malkonen, E., 1994. Microbial biomass C and N, and respiratory activity in soil of repeatedly limed and N-and P-fertilized Norway spruce stands. Soil Biology and Biochemistry 26, 957–962.
Soil Classification Working Group, 1998. The Canadian System of Soil Classification. Agriculture and Agri-Food Canada Publication 1646 (Revised), 187pp.
Srivastava, S.C., Singh, J.S., 1991. Microbial C, N and P in dry tropical forest soils: effects of alternate land-uses and nutrient flux. Soil Biology and Biochemistry 23, 117–124.
Startsev, N.A., McNabb, D.H., Startsev, A.D., 1998. Soil biological activity in recent clearcuts in west-central Alberta. Canadian Journal of Soil Science 78, 69–76.
Steijlen, I., Nilsson, M.C., Zackrisson, O., 1995. Seed regeneration of Scots pine in boreal forest stands dominated by lichen and feather moss. Canadian Journal of Forest Research 25, 713–723.
Stone, D.M., Elioff, J.D., 1998. Soil properties and aspen development five years after compaction and forest floor removal. Canadian Journal of Soil Science 78, 51–58.
Stone, D.M., Kabzems, R., 2002. Aspen development on similar soils in Minnesota and British Columbia after compaction and forest floor removal. The Forestry Chronicle 6, 886–891.
Strong, W.L., La Roi, G.H., 1985. Root density–soil relationships in selected boreal forests of central Alberta, Canada. Forest Ecology and Management 12, 233–251.
Tan, X., Chang, S.X., Kabzems, R., 2005. Effects of soil compaction and forest floor removal on soil microbial properties and N transformations in a boreal forest long-term soil productivity study. Forest Ecology and Management 217, 158–170.
ter Braak, C.J.F., 1988. CANOCO: an extension of DECORAMA to analyze species-environment relationships. Vegetation 75, 159–160. Vance, E.D., Brookes, P.C., Jenkinson, D.S., 1987. An extraction method
for measuring soil microbial biomass-C. Soil Biology and Biochemistry 19, 703–707.
Verdonschot, P.F.M., ter Braak, C.J.F., 1994. An experimental manip-ulation of oligochaete communities in mesocosms treated with chlorpyrifos or nutrient additions—multivariate analyses with Monte-Carlo permutation tests. Hydrobiologia 278, 251–266.
Voroney, R.P., Winter, J.P., Beyaert, R.P., 1993. Soil Microbial Biomass C and N. In: Carter, M.R. (Ed.), Soil Sampling and Methods of Analysis. Canadian Society of Soil Science, Lewis Publisher, Boca Raton, pp. 277–286.
Williamson, J.R., Neilsen, W.A., 2000. The influence of forest site on rate and extent of soil compaction and profile disturbance of skid trails during ground-based harvesting. Canadian Journal of Forest Research 30, 1196–1205.