AKTIVITAS ANTIHIPERTENSI AKTINOMISET
ENDOFIT ASAL TANAMAN PEGAGAN DAN
BELIMBING WULUH
WAHYU EKA SARI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ABSTRAK
WAHYU EKA SARI. Aktivitas Antihipertensi Aktinomiset Endofit Asal Tanaman Pegagan dan Belimbing Wuluh. Dibimbing oleh YULIN LESTARI dan MIN RAHMINIWATI.
Aktinomiset endofit hidup di dalam jaringan tanaman tanpa menimbulkan efek negatif. Beberapa tanaman obat berasosiasi dengan mikrob endofit yang dapat menghasilkan senyawa bioaktif yang sama dengan tanaman inangnya. Inhibisi Angiotensin Converting Enzyme (ACE) merupakan salah satu mekanisme antihipertensi yang efektif. Potensi aktinomiset endofit tanaman obat sebagai penghasil inhibitor ACE perlu dikaji. Tujuan penelitian ini adalah mengisolasi aktinomiset endofit dari akar, batang, daun pegagan (Centella a sia tica ), dan buah belimbing wuluh (Averrhoa bilimbi), serta mengkaji aktivitasnya sebagai antihipertensi melalui aktivitas inhibitor ACE. Aktinomiset endofit diisolasi dari permukaan tanaman yang telah disterilisasi, serta ditumbuhkan dan disebar dalam media Humic-a cid Vita min-B a gar (HV). Purifikasi isolat ditumbuhkan pada media Interna tiona l Streptomyces P r oject No.2 aga r (ISP2), dan ekstrak kasar aktinomiset endofit digunakan untuk uji aktivitas inhibitor ACE secara in vitro. Sebanyak 12 isolat aktinomiset endofit berhasil diisolasi dari bagian daun pegagan (tiga isolat) dan buah belimbing wuluh (sembilan isolat). Data yang diperoleh menunjukkan bahwa sembilan isolat (AEP-1, AEP-2, AEP-3, AEB-1, AEB-3, AEB-5, AEB-6, AEB-7, dan AEB-8) menunjukkan keragaman aktivitas ACE inhibitor dan tergolong ke dalam Streptomyces spp.. Tiga isolat lainnya (AEB-2, AEB-4, dan AEB-9) tergolong ke dalam non-Streptomyces, dan juga menunjukkan keragaman aktivitas inhibitor ACE. Sementara itu, tanaman kultur jaringan pegagan yang bebas dari mikrob endofit tidak menunjukkan adanya aktivitas inhibisi ACE. Isolat AEP-1 memiliki persentase aktivitas inhibitor ACE tertinggi (279,2%), diikuti oleh isolat AEB-5 juga memiliki persentase aktivitas inhibitor ACE tertinggi (222,92%), melebihi aktivitas penghambatan ACE oleh captopril (0,01 mg/mL) sebesar 61,5%, dan captopril (0,02 mg/mL) sebesar 66,7%. Data tersebut mengindikasikan bahwa isolat aktinomiset endofit asal daun pegagan dan buah belimbing wuluh mampu memproduksi senyawa inhibitor ACE.
ABSTRACT
WAHYU EKA SARI. Antihypertension Activity of Endophytic Actinomycetes Isolated from Plants of Pegagan and Belimbing Wuluh. Under direction of YULIN LESTARI and MIN RAHMINIWATI.
AKTIVITAS ANTIHIPERTENSI AKTINOMISET
ENDOFIT ASAL TANAMAN PEGAGAN DAN
BELIMBING WULUH
WAHYU EKA SARI
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada
Departemen Biologi
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
Judul : Aktivitas Antihipertensi Aktinomiset Endofit Asal Tanaman Pegagan dan
Belimbing Wuluh
Nama : Wahyu Eka Sari
NRP : G34061708
Disetujui
Dr. Ir. Yulin Lestari
drh. Min Rahminiwati, Ph.D.
NIP 19620710 198803 2 002 NIP 19610528 198503 2 004
Diketahui
Ketua Departemen Biologi
Dr. Ir. Ence Darmo Jaya Supena, M.Si.
NIP 19641002 198903 1 002
Tanggal Lulus :
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala rahmat dan karunia-Nya, sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Februari 2010 sampai Oktober 2010 ini ialah aktinomiset endofit pada tanaman obat. Penelitian ini bertujuan mengisolasi dan menapis aktinomiset endofit asal tanaman pegagan dan buah belimbing wuluh sebagai penghasil senyawa antihipertensi melalui aktivitas inhibitor ACE.
Penulis mengucapkan terima kasih kepada Ibu Dr. Ir. Yulin Lestari dan Ibu drh. Min Rahminiwati, Ph.D. selaku pembimbing yang telah banyak memberikan nasehat, saran, motivasi, dan waktu konsultasi, serta solusi dari setiap permasalahan yang dihadapi penulis selama melaksanakan penelitian dan penyusunan karya ilmiah ini. Terima kasih pula penulis ucapkan kepada Ibu Ir. Agustin Wydia Gunawan, M.S. selaku dosen penguji komisi pendidikan Departemen Biologi FMIPA IPB, atas koreksi dan sumbang sarannya terhadap perbaikan karya ilmiah ini. Disamping itu, penulis juga ingin menyampaikan terima kasih kepada Ibu Heni dan Bapak Jaka selaku staf Laboratorium Mikrobiologi IPB, Mbak Wiwi dan Mas Endi selaku staf Laboratorium Uji Biofarmaka LPPM-IPB, Bapak Ir. Edi Sandra M.Si. selaku dosen kultur jaringan Departemen KSHE IPB, dan Mas Eko selaku staf Laboratorium Bersama Kimia IPB, yang telah membantu selama penelitian berlangsung. Ungkapan terima kasih juga disampaikan kepada Ayah Sumarsono, Ibu Sumiyati, dan Adik Tian, serta keluarga, atas segala doa dan kasih sayangnya. Terima kasih pula atas dukungan, motivasi, dan bantuan yang diberikan oleh sahabat-sahabat, kakak-kakak, dan adik-adik di Biologi IPB, FORCES IPB, dan wisma bintang, serta seluruh pihak yang telah memberikan doa dan dukungannya.
Semoga karya ilmiah ini dapat bermanfaat. Amin.
Bogor, Maret 2011
RIWAYAT HIDUP
Penulis dilahirkan di Cilacap pada tanggal 23 Desember 1988 dari ayah Turyadi dan ibu Sumiyati,S.Ag. Penulis merupakan putri pertama dari dua bersaudara. Tahun 2006 penulis lulus dari SMA Negeri 3 Cilacap dan pada tahun yang sama lulus seleksi masuk IPB melalui jalur Undangan Seleksi Masuk IPB (USMI). Penulis memilih mayor Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, serta minor Gizi Masyarakat, Fakultas Ekologi Manusia.
Selama mengikuti perkuliahan, penulis menjadi asisten praktikum Perkembangan Hewan dan Biologi Dasar TPB pada tahun ajaran 2008/2009, praktikum Fisiologi Tumbuhan dan Biologi Dasar TPB pada tahun ajaran 2009/2010, serta praktikum Fisiologi Prokariot dan Biologi Dasar TPB pada tahun ajaran 2010/2011. Penulis pernah menjadi delegasi IPB dalam kongres pelajar internasional Asian Science Camp di Bali pada tahun 2008, dan pada tahun 2009 penulis menerima penghargaan dari Departemen Biologi sebagai mahasiswa berprestasi bidang non-akademik 2009. Penulis juga pernah menjuarai berbagai lomba diantaranya ialah Juara I lomba essay ilmiah tingkat IPB (2007), Juara III lomba musabaqah tilawatil qur’an cabang MSQ tingkat IPB (2007), Penerima hibah DIKTI tingkat nasional untuk Program Kreativitas Mahasiswa (PKM-Artikel Ilmiah tahun 2008, Penelitian dan Pengabdian Masyarakat tahun 2009, serta PKM-Penelitian tahun 2010). Selain itu, penulis pernah menjadi finalis dari lomba karya tulis tingkat nasional bidang kesehatan di Universitas Airlangga Surabaya, dan bidang ekologi manusia di Institut Pertanian Bogor pada tahun 2009.
Penulis juga aktif dalam kegiatan organisasi unit kegiatan mahasiswa Forum for Scientific Studies (FORCES) sejak tahun ajaran 2006/2007 hingga 2009/2010, dan jabatan tertinggi sebagai wakil direktur FORCES pada tahun ajaran 2008/2009. Di samping itu, penulis juga pernah aktif dalam kegiatan organisasi LDK-DKM Al-Hurriyah IPB (tahun ajaran 2007/2008), Himpunan Mahasiswa Biologi (HIMABIO) IPB (tahun ajaran 2007/2008), dan Rohis Biologi 43 (tahun ajaran 2007/2008 hingga 2009/2010). Penulis berkesempatan menjalani Praktik Lapangan dengan judul “Biodegradasi Pestisida Carbaryl pada Kondisi Anaerobik oleh Mikroba Denitrifikasi dari Tanah Pertanian Intensif Lembang dan Dieng” di Laboratorium Mikrobiologi, Pusat Penelitian Biologi, Lembaga Ilmu Pengetahuan Indonesia (LIPI) Cibinong pada tahun 2009.
Beasiswa yang pernah penulis peroleh selama masa perkuliahan antara lain beasiswa Peningkatan Prestasi Akademik (PPA) pada tahun ajaran 2006/2007 hingga tahun 2007/2008, beasiswa Bantuan Belajar Mahasiswa (BBM) pada tahun ajaran 2008/2009, dan beasiswa Yayasan Karya Salemba Empat (KSE) pada tahun ajaran 2009/2010.
DAFTAR ISI
Halaman
DAFTAR GAMBAR ... .... vi
DAFTAR LAMPIRAN ... vi
PENDAHULUAN ... ... 1
BAHAN DAN METODE Waktu dan Tempat Penelitian ... 2
Isolasi dan Kultivasi Mikrob Endofit dari Akar, Batang, dan Daun Pegagan, serta Buah Belimbing wuluh ... 2
Purifikasi Aktinomiset Endofit dan Pengamatan Mikroskopis ... 2
Produksi Filtrat Kultur Aktinomiset Endofit Asal Daun Pegagan dan Buah Belimbing Wuluh, serta Pengukuran Biomassa... 2
Uji in vitro Inhibisi Ekstrak Aktinomiset Endofit terhadap Aktivitas ACE ... 3
HASIL Isolasi dan Kultivasi Mikrob Endofit dari Akar, Batang, dan Daun Pegagan, serta Buah Belimbing wuluh... 3
Purifikasi Aktinomiset Endofit dan Pengamatan Mikroskopis ... 4
Produksi Filtrat Kultur Aktinomiset Endofit Asal Daun Pegagan dan Buah Belimbing Wuluh, serta Pengukuran Biomassa... 5
Uji in vitro Inhibisi Ekstrak Aktinomiset Endofit terhadap Aktivitas ACE ... 5
PEMBAHASAN... ... 7
SIMPULAN DAN SARAN Simpulan... 10
Saran... 10
DAFTAR PUSTAKA ... 11
DAFTAR GAMBAR
Halaman
1 Tanaman kultur jaringan pegagan steril mikrob endofit umur 55 hari………... 3
2 Koloni aktinomiset endofit pada media HV umur 3 minggu………... 4
3 Keragaman morfologi koloni aktinomiset endofit umur 10 hari pada media ISP2. Streptomyces spp. asal daun pegagan (a) AEP-1, (b) AEP-2, (c) AEP-3, asal buah belimbing wuluh (d) AEB-1, (e) AEB-3, (f) AEB-5, (g) AEB-6, (h) AEB-7, (i) AEB-8; non-Streptomyces asal buah belimbing wuluh (j) AEB-2, (k) AEB-4, (l) AEB-9... 4
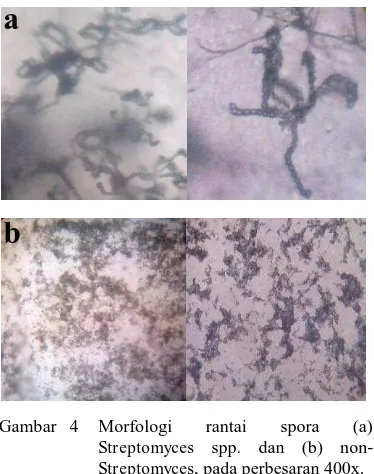
4 Morfologi rantai spora (a) Streptomyces spp. dan (b) non-Streptomyces, pada perbesaran 400x... 5
5 Pertumbuhan aktinomiset endofit pada media cair ISP2 selama 10 hari (a) isolat potensi asal daun pegagan (AEP-1), (b) isolat potensi asal buah belimbing wuluh (AEB-5)... 5
6 Persentase aktivitas penghambatan ACE ekstrak kasar isolat aktinomiset endofit asal daun pegagan (AEP-1, AEP-2, dan AEP-3), ekstrak kultur jaringan tanaman pegagan (EP1), ekstrak daun pegagan (EP2), captopril (C1 dan C2), dan media cair ISP2 (M)... 6
7 Persentase aktivitas penghambatan ACE ekstrak kasar isolat aktinomiset endofit asal buah belimbing wuluh (AEB-1, AEB-2, AEB-3, AEB-4, AEB-5, AEB-6, AEB-7, AEB-8, dan AEB-9), ekstrak buah belimbing wuluh (EBW), captopril (C1 dan C2), dan media cair ISP2 (M)... 6
DAFTAR LAMPIRAN
Halaman
1 Komposisi media agar-agar asam humat mengandung vitamin B (HV)... 142 Komposisi media agar-agar International Streptomyces Project No.2 (ISP2)... 14
3 Komposisi media Murashige & Skoog (MS)... 14
4 Nilai absorbansi hasil pengukuran spektrofotometri λ 228 nm... 15
1
PENDAHULUAN
Mikrob endofit hidup di dalam jaringan tanaman pada periode tertentu tanpa menimbulkan bahaya, serta dapat diisolasi dari jaringan tanaman yang sudah disterilisasi permukaannya atau diekstrak dari jaringan tanaman bagian dalam (Hallman et al. 1997). Mikrob ini merupakan sumber alamiah yang potensial dari dalam jaringan tanaman yang dapat dikaji manfaatnya dalam bidang obat-obatan, pertanian, dan industri (Strobel & Daisy 2003). Berdasarkan penelitian Hasegawa et al. (2006) mikrob endofit akan mengkolonisasi jaringan tanaman, serta memperoleh nutrisi dan perlindungan dari tanaman inangnya. Populasi mikrob yang melimpah di alam, baik di tanah, air, maupun yang bersifat endofit, memiliki potensi untuk dikaji kemampuannya sebagai penghasil senyawa antihipertensi.
Hipertensi merupakan suatu keadaan seseorang ketika terjadi peningkatan tekanan darah sistolik ≥ 140 mmHg atau tekanan darah diastolik ≥ 90 mmHg, penderita memiliki resiko penyakit jantung, stroke, dan gagal ginjal (Iskandar 2007; Yusuf 2008). Beberapa penyebab munculnya hipertensi antara lain penyakit gagal ginjal, kelainan endokrin, asupan garam terlalu tinggi, stres atau salah pemakaian obat (Iskandar 2007). Selain itu, tinggi rendahnya tekanan darah juga dipengaruhi oleh faktor Renin Angiotensin System (RAS), yang melibatkan pengubahan zat angiotensin I menjadi angiotensin II (Yusuf 2008). Angiotensin II berfungsi untuk sekresi aldosteron penyebab retensi sodium yang dapat meningkatkan volume cairan ekstraseluler, sehingga mengakibatkan terjadinya hipertensi. Dengan menghambat aktivitas angiotensin converting enzyme (ACE), maka angiotensin I tidak diubah menjadi angiotensin II, sehingga hipertensi dapat dicegah. Metode inhibitor ACE merupakan metode skrining antihipertensi yang efektif (Wagner et al. 1991; Hansen et al. 1995; Somanadhan et al. 1996).
Beberapa tanaman obat secara empiris digunakan sebagai obat tradisional untuk mengendalikan hipertensi. Salah satunya adalah ekstrak tanaman pegagan (Centella asiatica) dan buah belimbing wuluh (Averrhoa bilimbi) (Wijayakusuma & Dalimartha 2005). Akan tetapi, pengobatan menggunakan tanaman obat membutuhkan banyak biomassa dan waktu tumbuh yang lama, serta dapat mengganggu kelestarian
alam jika dieksploitasi secara berlebihan, sehingga diperlukan inovasi yang efektif dan efisien sebagai solusi permasalahan tersebut.
Cara inovatif untuk mengefisienkan sumber senyawa bioaktif adalah dengan memanfaatkan mikrob endofit yang berasosiasi dengan tanaman obat tersebut. Menurut Strobel & Daisy (2003) berbagai jenis senyawa bioaktif dengan beragam fungsi yang terkandung di dalam tumbuhan, diduga dapat pula dihasilkan oleh mikrob endofit pada tumbuhan tersebut. Adanya kemampuan mikrob endofit menghasilkan senyawa metabolit sekunder sesuai dengan tanaman inangnya, merupakan peluang yang dapat dioptimalkan untuk memproduksi metabolit sekunder secara efisien dan cepat.
Penelitian sebelumnya terhadap ekstrak tanaman pegagan menunjukkan bahwa tanaman tersebut mengandung senyawa bioaktif Triterpenoid (Wijayakusuma & Dalimartha 2005). Triterpenoid merupakan senyawa bioaktif yang dapat berfungsi sebagai antioksidan, sehingga dapat menangkap radikal bebas sebagai salah satu penyebab timbulnya penyakit hipertensi. Selain itu, secara tradisional, buah belimbing wuluh pada umumnya juga digunakan oleh masyarakat untuk mengobati penyakit hipertensi. Menurut Wijayakusuma & Dalimartha (2005) dan Iskandar (2007) bagian yang sering digunakan dari tanaman belimbing wuluh untuk mengobati hipertensi adalah buahnya. Buah belimbing wuluh mengandung zat kalium yang dapat melancarkan keluarnya air seni, sehingga dapat menurunkan tekanan darah tinggi (Hariana 2004).
Aktinomiset merupakan bakteri Gram positif berfilamen dan dapat berperan sebagai penghasil beragam senyawa bioaktif yang dapat berfungsi antara lain sebagai antibiotik, enzim inhibitor, dan senyawa bioaktif lainnya (Lestari 2006). Mikrob ini juga dikenal sebagai penghasil antibiotik terbesar.
2
BAHAN DAN METODE
Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan pada bulan Februari hingga Oktober 2010 di Laboratorium Mikrobiologi, Departemen Biologi, FMIPA-IPB, Laboratorium Bersama Kimia, Departemen Kimia, FMIPA-IPB, dan Laboratorium Uji Biofarmaka, Pusat Studi Biofarmaka-LPPM-IPB.
Isolasi dan Kultivasi Mikrob Endofit dari Akar, Batang, dan Daun Pegagan, serta Buah Belimbing Wuluh
Tanaman pegagan yang digunakan sebagai sumber mikrob endofit berasal dari kebun koleksi tanaman obat Biofarmaka IPB dan Desa lingkar Kampus IPB Dramaga, sedangkan buah belimbing wuluh berasal dari perumahan dosen Kampus IPB Dramaga. Bagian tanaman seperti akar, batang, dan daun pegagan umum digunakan untuk obat antihipertensi, sedangkan pada belimbing wuluh adalah buahnya. Isolasi mikrob endofit mengacu pada Coombs dan Franco (2003) yang dimodifikasi dalam hal konsentrasi NaOCl yang digunakan untuk sterilisasi permukaan. Sampel akar, batang, dan daun pegagan serta buah belimbing wuluh yang telah dipotong, disterilisasi permukaannya secara bertahap yaitu direndam dalam alkohol 70% selama 1 menit, kemudian direndam dalam natrium hipoklorit (NaOCl) 1% selama 5 menit, dan selanjutnya direndam kembali dalam alkohol 70% selama 1 menit. Langkah terakhir dibilas sebanyak 3 kali dengan akuades steril. Potongan bagian tanaman yang telah steril permukaannya digerus menggunakan mortar secara aseptik, lalu dilarutkan dalam 12.5 mM bufer fosfat (pH 7.1). Ekstrak yang diperoleh disentrifugasi pada kecepatan 3000 rpm selama 10 menit pada suhu ruang. Supernatan hasil sentrifugasi diambil sebanyak 0,1 ml, lalu disebar secara merata pada cawan Petri berisi media agar-agar asam humat yang mengandung vitamin B (media HV) (Lampiran 1). Media tersebut mengandung antibiotik sikloheksamida (50 mg/L media), dan asam nalidiksat (20 mg/L media). Selanjutnya cawan berisi media dan supernatan tersebut diinkubasi pada suhu ruang (28°C) selama 3 minggu agar pertumbuhan koloni aktinomiset endofit dapat diamati dengan baik. Isolasi aktinomiset endofit dilakukan dua kali pengulangan.
Untuk membuktikan bahwa koloni aktinomiset endofit yang tumbuh hanya berasal dari dalam jaringan kedua tanaman
tersebut, maka dilakukan uji kontrol negatif terhadap air sisa hasil rendaman terakhir ketika proses sterilisasi permukaan selesai, dengan cara menyebar 0,1 mL air sisa rendaman tersebut ke atas media uji, dan dilakukan tiga kali pengulangan.
Purifikasi Aktinomiset Endofit dan Pengamatan Mikroskopis
Mikrob endofit hasil isolasi yang telah tumbuh pada media HV, selanjutnya dimurnikan pada media agar-agar International Streptomyces Project 2 (ISP2) (Lampiran 2). Purifikasi aktinomiset endofit dilakukan dua kali pengulangan. Langkah berikutnya adalah inkubasi isolat yang dilakukan selama 7 hari pada suhu ruang. Setelah itu, morfologi keragaman rantai spora diamati secara mikroskopis. Isolat aktinomiset endofit yang telah diperoleh, diletakkan di atas kaca preparat dengan ditetesi sedikit air di atasnya, selanjutnya diamati pada perbesaran 400x menggunakan mikroskop cahaya, sampai tampak jelas morfologi rantai spora yang terbentuk pada masing-masing isolat.
Produksi Filtrat Kultur Aktinomiset Endofit Asal Daun Pegagan dan Buah Belimbing Wuluh, serta Pengukuran Biomassa
3
Aktivitas inhibitor ACE (%) = 100–[100 x (C–D)/(A–B)
adalah melakukan pengukuran biomassa menggunakan neraca analitik.
Uji in vitro Inhibisi Ekstrak A kt i no mi s e t E n do fit terhadap Aktivitas ACE
Pengukuran aktivitas inhibitor ACE pada penelitian ini mengacu pada metode Hayes et al. (2007) yang telah dimodifikasi dalam hal konsentrasi enzim, substrat, dan bahan pelarut lainnya yang digunakan untuk uji in vitro. Substrat enzim yang digunakan untuk uji in vitro penghambatan ACE adalah Hippuryl-L-Histidyl-L-Leucine (HHL) dari Sigma Co. Sebanyak 200 µl bufer HHL (2,5 mM HHL dalam 0,05 M bufer natrium borat, mengandung 0,15 M NaCl, pada pH 8.3) dicampur dengan 80 µl filtrat aktinomiset endofit (sampel), selanjutnya diinkubasi pada inkubator bergoyang selama 3 menit pada suhu 37ºC. Campuran antara bufer HHL dengan sampel akan mulai bereaksi setelah dilakukan penambahan 20 µl (ACE-A6778, Sigma Aldrich Co.) (0,03745 unit/mL). Selanjutnya campuran tersebut diinkubasi pada inkubator bergoyang selama 1 jam pada suhu 37ºC. Reaksi dihentikan dengan penambahan 250 µl 0,5 M HCl, dan selanjutnya ditambahkan 1,7 ml etil asetat sebagai pelarut untuk evaporasi. Evaporasi dilakukan menggunakan alat rotavapor R-205 BUCHI, heating bath B-490 BUCHI, untuk menghilangkan pelarut etil asetat dari fraksi yang dilarutkan serta untuk menghilangkan pengotor dari campuran reaksi. Fraksi sampel yang telah terpisah dari pelarut etil asetat setelah evaporasi, kemudian dilarutkan kembali dengan penambahan 1 ml air destilata. Langkah terakhir adalah pengukuran absorbansi aktivitas inhibisi ACE menggunakan spektrofotometer UV-1700 PharmaSpec SHIMADZU pada panjang gelombang (λ) 228 nm, dengan pengenceran sebanyak 15x. Pengukuran aktivitas inhibitor ACE ini dilakukan dengan dua kali ulangan.
Hasil absorbansi yang diperoleh selanjutnya dihitung menggunakan rumus:
Keterangan: A merupakan hasil absorbansi dengan ACE dan tanpa inhibitor ACE, B merupakan hasil absorbansi tanpa ACE dan tanpa inhibitor ACE, C merupakan hasil absorbansi dengan ACE dan inhibitor ACE, sedangkan D merupakan hasil absorbansi dengan inhibitor ACE dan tanpa ACE (Hayes et al. 2007). Sampel inhibitor ACE yang diuji aktivitas penghambatannya
terdiri atas captopril 0,01 mg/mL (C1) dan 0,02 mg/mL (C2) sebagai kontrol positif, media cair ISP2 (M) sebagai kontrol negatif, ekstrak kultur jaringan tanaman pegagan (EP1) asal Departemen Konservasi Sumber Daya Hutan dan Ekowisata, Fakultas Kehutanan IPB (Gambar 1), ekstrak daun pegagan (EP2), ekstrak buah belimbing wuluh (EBW), dan isolat aktinomiset endofit asal akar, batang, dan daun pegagan (AEP), serta aktinomiset endofit asal buah belimbing wuluh (AEB).
Semua sampel diberikan perlakuan yang sama sesuai dengan prosedur di atas. Contoh perhitungan persentase aktivitas penghambatan ACE dapat dilihat pada Lampiran 5.
Gambar 1 Tanaman kultur jaringan pegagan steril mikrob endofit umur 55 hari pada media MS.
Tanaman kultur jaringan pegagan dikatakan steril mikrob endofit karena media tanam (Murashige & Skoog (MS) (Lampiran 3) dengan penambahan auksin dan sitokinin), jaringan pegagan, serta proses pengkulturan dari awal hingga akhir, keseluruhan dilakukan secara aseptik.
HASIL
Isolasi dan Kultivasi Mikrob Endofit dari Akar, Batang, dan Daun Pegagan, serta Buah Belimbing Wuluh
Total isolat aktinomiset endofit yang berhasil diisolasi berjumlah 12 isolat. Tiga isolat berasal dari daun pegagan, sedangkan sembilan isolat lainnya dari buah belimbing wuluh. Isolat aktinomiset endofit tidak ada yang diperoleh dari akar dan batang pegagan. Semua isolat yang diperoleh dapat tumbuh dengan baik pada media HV yang merupakan media selektif bagi pertumbuhan aktinomiset, sehingga adanya koloni berwarna putih yang tumbuh pada media tersebut (Gambar 2), menandakan aktinomiset endofit.
4
a
b
c
d
e
f
g
h
i
j
k
l
hasil rendaman terakhir dari proses sterilisasi permukaan, tidak tumbuh satu koloni pun baik koloni berwarna putih yang menandakan aktinomiset endofit maupun koloni mikrob lainnya. Hal tersebut menandakan bahwa proses sterilisasi permukaan berhasil dan menguatkan dugaan bahwa aktinomiset endofit yang berhasil diisolasi pada penelitian ini berasal dari jaringan tanaman bagian dalam.
Gambar 2 Koloni aktinomiset endofit pada media HV umur 3 minggu.
Purifikasi aktinomiset endofit dan pengamatan mikroskopis
Hasil purifikasi aktinomiset endofit pada media ISP2 selama 7 hari menunjukkan keragaman karakteristik morfologi koloni dan pigmentasi yang beragam. Tiga isolat aktinomiset endofit yang diperoleh dari bagian daun pegagan ialah 1, 2, dan AEP-3 (Gambar AEP-3a-c), sedangkan sembilan isolat aktinomiset endofit lainnya hasil isolasi dari buah belimbing wuluh ialah AEB-1, AEB-2, AEB-3, AEB-4, AEB-5, AEB-6, AEB-7, AEB-8, AEB-9 (Gambar 3d-l).
Berdasarkan morfologi koloni di atas media ISP2, ke-12 isolat aktinomiset endofit tersebut dapat dibedakan antara Streptomyces spp. dan non-Streptomyces. Koloni aktinomiset sebagian besar akan tampak keras seperti tumbuh akar di dalam agar-agar, berbeda dengan koloni mikrob lainnya yang tampak lunak di atas media agar. Menurut Ghadin et al. (2008) Streptomyces spp. di atas media padat akan menunjukkan miselium dengan spora aerial berwarna putih hingga abu-abu. Genus non-Streptomyces hanya membentuk miselium vegetatif, dan akan tampak lembab di atas media agar (tidak membentuk spora aerial). Berdasarkan ciri-ciri tersebut, maka isolat AEP-1, AEP-2, AEP-3, AEB-1, AEB-3, AEB-5, AEB-6, AEB-7, dan AEB-8 termasuk ke dalam Streptomyces spp., sedangkan isolat AEB-2, AEB-4, dan AEB-9 termasuk ke dalam non-Streptomyces.
Penggolongan 12 isolat aktinomiset endofit ke dalam Streptomyces spp. dan non-Streptomyces juga didasarkan pada pengamatan rantai spora secara mikroskopis. Karakteristik morfologi rantai spora Streptomyces spp. mampu membentuk rantai spora aerial dengan morfologi yang beragam (seperti rantai, kait, hingga spiral) (Gambar 4a), sedangkan karakteristik morfologi rantai spora non-Streptomyces, tidak membentuk rantai spora aerial dan tidak berbentuk rantai, kait, atau spiral (Gambar 4b).
Gambar 3 Keragaman morfologi koloni aktinomiset endofit umur 10 hari pada media ISP2, Streptomyces spp. asal daun pegagan (a) AEP-1, (b) AEP-2, (c) AEP-3; asal buah belimbing wuluh (d) AEB-1, (e) AEB-3, (f) AEB-5, (g) AEB-6, (h) AEB-7, (i) AEB-8;
non-Streptomyces asal buah belimbing wuluh (j) AEB-2, (k) AEB-4, (l) AEB-9.
5
a
a
b
Granul-granul spora berwarna putih
Gambar 4 Morfologi rantai spora (a)
Streptomyces spp. dan (b)
non-Streptomyces, pada perbesaran 400x.
Produksi filtrat kultur aktinomiset endofit asal daun pegagan dan buah belimbing wuluh, serta pengukuran biomassa
Hasil produksi filtrat kultur aktinomiset endofit selama 10 hari menunjukkan keragaman warna filtrat yaitu dari kuning (seperti warna media cair ISP2) hingga merah bata (Gambar 5). Hal tersebut diduga karena sifat fisiologis masing-masing isolat berbeda-beda untuk memproduksi metabolit sekunder yang terkandung dalam filtrat.
Gambar 5 Pertumbuhan aktinomiset endofit pada media cair ISP2 selama 10 hari; (a) isolat potensi asal daun pegagan (AEP-1) (b) isolat potensi asal buah belimbing wuluh (AEB-5).
Produksi filtrat dilakukan menggunakan teknik aerasi dengan kecepatan 120 rpm pada suhu ruang sehingga isolat
tumbuh dengan membentuk granul-granul spora berwarna putih, tersuspensi dengan media dan mengendap jika didiamkan (Gambar 5).
Tabel 1 Nilai bobot biomassa aktinomiset endofit asal daun pegagan dan buah belimbing wuluh umur 10 hari pada media cair ISP2
Jenis isolat
Bobot biomassa total
(mg/mL) Aktinomiset endofit asal
daun pegagan
Streptomyces spp.
AEP-1 0,720 AEP-2 0,926 AEP-3 1,023 Aktinomiset endofit asal
buah belimbing wuluh
Streptomyces spp.
AEB-1 0,553 AEB-3 1,686 AEB-5 5,263 AEB-6 0,883 AEB-7 0,430 AEB-8 1,467 non-Streptomyces
AEB-2 1,320 AEB-4 2,426 AEB-9 2,413
Tabel 1 menunjukkan hasil pengukuran biomassa tiga isolat aktinomiset endofit asal daun pegagan dan sembilan isolat aktinomiset endofit asal buah belimbing wuluh. Bobot biomassa yang diperoleh ke-12 aktinomiset endofit menunjukkan keragaman antar isolat. Bobot biomassa isolat potensial aktinomiset endofit asal daun pegagan (AEP-1) dan buah belimbing wuluh (AEB-5) yang menunjukkan aktivitas penghambatan ACE tertinggi berturut-turut yaitu 0,720 mg/mL dan 5,263 mg/mL. Adanya keragaman bobot biomassa juga diduga karena kemampuan masing-masing isolat berbeda-beda dalam memproduksi metabolit sekunder yang terkandung dalam filtrat.
Uji in vitro inhibisi ekstrak ka s a r a kt i no mi s e t e n dof it terhadap aktivitas ACE
6
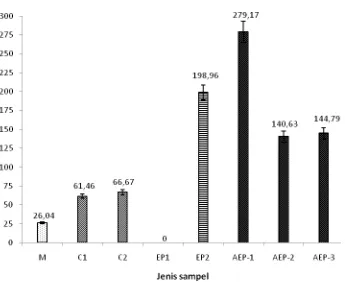
Gambar 6 Persentase aktivitas penghambatan ACE ekstrak kasar isolat aktinomiset endofit asal daun pegagan (AEP-1, AEP-2, dan AEP-3), ekstrak kultur jaringan tanaman pegagan (EP1), ekstrak daun pegagan (EP2), captopril (C1 dan C2), dan media cair ISP2 (M).
7
Gambar 6 menunjukkan bahwa persentase aktivitas penghambatan ACE yang ditunjukkan oleh ketiga isolat aktinomiset endofit asal daun pegagan (AEP-1, AEP-2, dan AEP-3) lebih tinggi dibandingkan dengan aktivitas kontrol negatif (media cair ISP2), dan kontrol positif (captopril). Persentase aktivitas penghambatan ACE oleh media cair ISP2 (M) sebesar 26,04%, lebih kecil dibandingkan dengan aktivitas tanaman inang pegagan, ketiga isolat aktinomiset endofit asal daun pegagan, dan kontrol positif captopril.
Berdasarkan data yang diperoleh, antara captopril 0,01 mg/mL (C1) dan 0,02 mg/mL (C2) menunjukkan aktivitas yang hampir sama satu sama lain. Captopril 0,01 mg/mL menunjukkan aktivitas sebesar 61,46%, sedangkan captopril 0,02 mg/mL sebesar 66,67%. Adapun persentase aktivitas penghambatan ACE oleh isolat AEP-1 sebesar 279,17%, isolat AEP-2 sebesar 140,63%, dan isolat AEP-3 sebesar 144,79%. Hal tersebut mengindikasikan bahwa nilai persentase penghambatan ACE yang dihasilkan oleh ketiga isolat aktinomiset endofit asal daun pegagan lebih tinggi dibandingkan dengan kontrol positif, sebesar dua hingga empat kali lipat. Volume semua sampel yang diujikan besarnya sama yaitu sebanyak 80 µL dari bobot biomassa yang berbeda-beda. Aktivitas penghambatan ACE yang ditunjukkan oleh ekstrak daun pegagan sebesar 198,96%. Berdasarkan perhitungan nilai absorbansi hasil pengukuran spektrofotometri pada λ 228 nm (Lampiran 4), diperoleh persentase aktivitas penghambatan ACE oleh tanaman kultur jaringan pegagan steril mikrob endofit (EP1) sebesar -14,58% (negatif), sehingga ekstrak tersebut dianggap aktivitas penghambatannya nol persen.
Gambar 7 menunjukkan bahwa persentase aktivitas penghambatan ACE oleh ke-12 isolat aktinomiset endofit asal buah belimbing wuluh hampir sama dengan aktivitas yang ditunjukkan oleh ekstrak tanaman inangnya (166,67%), bahkan ada dua isolat (AEB-4 dan AEB-5) yang aktivitasnya lebih besar hingga dua kali lipat (184,38% dan 222,92%). Isolat aktinomiset endofit asal buah belimbing wuluh yang menunjukkan aktivitas penghambatan tertinggi adalah isolat AEB-5 yaitu sebesar 222,92%, sedangkan aktivitas penghambatan ACE terendah terdapat pada isolat AEB-8 yaitu sebesar 0%. Hal tersebut menunjukkan bahwa tidak semua isolat aktinomiset endofit memiliki kemampuan untuk menghambat ACE.
Jika aktivitas penghambatan ACE yang ditunjukkan oleh Gambar 6 dan 7 dibandingkan, maka dapat dilihat adanya perbedaan aktivitas antara ekstrak daun pegagan dengan ekstrak buah belimbing wuluh. Persentase aktivitas penghambatan ACE yang ditunjukkan oleh ekstrak daun pegagan (EP2) yaitu 198,96%, sedangkan ekstrak buah belimbing wuluh (EBW) hanya sebesar 166,67%. Oleh karena itu, dapat dinyatakan bahwa aktivitas antihipertensi ekstrak daun pegagan lebih tinggi dibandingkan dengan aktivitas antihipertensi ekstrak buah belimbing wuluh.
Selain itu, aktivitas tertinggi yang ditunjukkan oleh isolat aktinomiset endofit AEP-1 asal daun pegagan sebesar 279,17% juga menunjukkan nilai yang lebih tinggi dibandingkan dengan aktivitas penghambatan ACE tertinggi oleh isolat AEB-5 asal buah belimbing wuluh (222,92%).
PEMBAHASAN
Aktinomiset endofit asal daun pegagan dan buah belimbing wuluh menunjukkan keragaman morfologi koloni pada media ISP2 (Gambar 3), bentuk rantai spora secara mikroskopis (Gambar 4), dan aktivitas penghambatan terhadap ACE (Gambar 6 & 7). Menurut Takahashi dan Omura (2003) keragaman jenis aktinomiset yang terisolasi bergantung pada asal habitatnya, jenis media yang digunakan, dan metode isolasi yang dipakai. Media yang digunakan untuk isolasi pada penelitian ini adalah media HV yang ditambahkan antibiotik sikloheksamida (50 mg/mL) untuk menekan pertumbuhan cendawan dan asam nalidiksat (20 mg/mL) untuk menekan pertumbuhan bakteri Gram negatif.
8
Koloni aktinomiset endofit pada media HV tampak dominan berwarna putih (Gambar 2). Pemurnian koloni tersebut dilakukan pada media ISP2. Aktinomiset endofit yang tumbuh pada media ISP2 tampak memiliki morfologi koloni yang beragam, demikian pula dengan bentuk rantai sporanya.
Keberadaan aktinomiset di lingkungan sangat melimpah terutama di rizosfer. Aktinomiset endofit berasosiasi dengan tanaman inang dan dapat memberikan efek yang menguntungkan, serta tidak membahayakan bagi tanaman inangnya. Sharma et al. (2005) menyatakan bahwa akar lateral merupakan bagian tumbuhan yang paling banyak dihuni oleh mikrob endofit. Hal ini dikarenakan mikrob endofit masuk ke dalam jaringan tanaman melalui akar lateral kemudian menyebar ke dalam ruang interseluler dan berkas pembuluh. Selain itu, mikrob endofit juga dapat masuk melalui bagian daun, bunga, batang, stomata, kotiledon, dan bagian tanaman yang terluka. Dalam penelitian ini aktinomiset endofit berhasil diisolasi dari daun pegagan, namun tidak diperoleh dari bagian akar dan batang pegagan, sedangkan pada belimbing wuluh hanya digunakan buahnya yang umum digunakan oleh masyarakat dan berhasil diisolasi aktinomiset endofitnya. Tidak diperolehnya aktinomiset endofit dari akar dan batang pegagan kemungkinan karena adanya faktor preferensi dalam hal mikrob endofit tersebut mengkolonisasi tanaman inangnya. Aktinomiset merupakan bakteri Gram positif berfilamen, dengan kandungan guanin dan sitosin (G+C) yang tinggi (>55%) di dalam genomnya (Miyadoh 1997). Beberapa senyawa bioaktif dihasilkan oleh kelompok aktinomiset. Menurut Raja dan Prabakarana (2011) aktinomiset dikenal sebagai penghasil antibiotik terbesar, karena dari 16.500 antibiotik yang telah ditemukan, lebih dari setengahnya dihasilkan oleh aktinomiset. Sebagian besar aktinomiset (95%) beranggotakan Streptomyces (Lachevalier et al. 1977). Koloni aktinomiset yang tergolong Streptomyces spp. membentuk miselium aerial dan secara mikroskopis memiliki morfologi rantai spora seperti kait, spiral atau heliks (Kudo 1997). Aktinomiset yang tidak membentuk miselium aerial atau hanya membentuk miselium dalam substrat merupakan kelompok non-Streptomyces. Genus yang digolongkan ke dalam non-Streptomyces antara lain Mycobacterium, Nocardia, Micromonospora, Microbispora, A ctinoplanes, dan Actinomadura (Miyadoh
1997). Menurut Miyadoh & Otoguro (2004) morfologi rantai spora, permukaan spora, warna miselium, serta pigmentasi dapat dijadikan dasar klasifikasi hingga tingkat spesies. Berdasarkan pengamatan terhadap karakteristik morfologi koloni aktinomiset pada media ISP2, tampak bahwa isolat AEP-1, AEP-2, AEP-3, AEB-AEP-1, AEB-3, AEB-5, AEB-6, AEB-7, AEB-8 termasuk Streptomyces spp. karena mampu membentuk miselium aerial dan spora berwarna putih, putih kekuningan, putih kecokelatan, hingga abu-abu (Gambar 3), serta menunjukkan rantai spora yang tersusun keriting, seperti kait atau spiral, (Gambar 4a). Isolat AEB-2, AEB-4, dan AEB-9 tergolong non-Streptomyces, karena tidak membentuk rantai spora aerial (Gambar 4b). Ghadin et al. (2008) menyatakan bahwa karakteristik Streptomyces pada media padat ditunjukkan dengan munculya substrat miselium setelah empat hari inkubasi dan formasi miselium yang lengkap setelah enam hari inkubasi, dengan warna spora aerial berwarna putih hingga abu-abu.
Interaksi antara mikrob endofit dengan tanaman inangnya merupakan hubungan simbiosis mutualisme. Koloni mikrob endofit di dalam jaringan tanaman akan memperoleh nutrisi dan perlindungan dari tanaman inangnya, sedangkan tanaman inang memperoleh senyawa bioaktif dari mikrob endofit yang dapat berfungsi sebagai antimikrob, pemacu pertumbuhan tanaman, enzim pendegradasi lignin, selulosa, hemiselulosa, kitinase, amilase, dan glukanase (Hasegawa et al. 2006). Castillo et al. (2002) berhasil mengisolasi aktinomiset endofit Streptomyces sp. strain NRRL 30562 asal tanaman Snakevine (Kennedia nigrisca) yang dipercaya suku Aborigin untuk mengobati luka dan infeksi, yang ternyata mampu menghasilkan senyawa munumbycin A-D, antibiotik peptida baru berspektrum luas.
9
stamineus) juga pada umumnya digunakan masyarakat secara tradisional untuk obat hipertensi. Tanaman tingkat tinggi dapat mengandung mikrob endofit yang mampu menghasilkan senyawa bioaktif atau metabolit sekunder yang diduga sebagai akibat koevolusi atau transfer genetik dari tanaman inangnya ke dalam mikrob endofit (Tan & Zou 2001).
Berdasarkan penelitian Tejesvi et al. (2008) pada tanaman obat Terminalia arjuna , T. chebula, Azadirachta indica , dan Holarrhena antidysenterica diperoleh isolat dominan yaitu mikrob endofit Pestalotiopsis, yang mempunyai aktivitas penghambatan ACE sebesar > 60%. Anggota aktinomiset seperti Streptomyces chromofuscus (Nakatsukasa et al. 1985) dan Micromonospora halophytica (Lima 1999) juga diketahui dapat menghasilkan inhibitor ACE.
Daya hambat aktivitas enzim ACE diukur menggunakan substrat N-Hippuril-L-histidyl-L-leucine hydrate (HHL) .yang akan terhidrolisis menjadi N-Hippuric acid dan L-histidyl-L-leucine (Kasahara & Ashihara 1981). Kondisi optimal substrat HHL untuk uji inhibisi ACE adalah pada pH 8.3 pelarut NaCl 300 mM (Cheung et a l. 1980). Dalam penelitian ini, substrat HHL dilarutkan menggunakan NaCl 150 mM, pH 8.3.
ACE merupakan enzim yang dapat mengubah angiotensin I (Asp-Arg-Val-Tyr-Ile-His-Pro-Phe-His-Leu) menjadi angiotensin II (Asp-Arg-Val-Tyr-Ile-His-Pro-Phe). Ondetti et al. (1983) melaporkan bahwa inhibitor ACE dapat berfungsi sebagai antihipertensi. Keunggulan inhibitor ACE sebagai mekanisme antihipertensi selain dapat berperan dalam proses diuretik, ACE-I juga dapat berperan melalui aktivitas saraf simpatik. Angiotensin II mempunyai efek vasokonstriksi yang kuat, meningkatkan aktivitas sistem saraf simpatik, serta dapat merangsang produksi aldosteron (Akil & Bakri 2001). Peningkatan aktivitas saraf simpatik dapat meningkatkan aktivasi β-1 adrenoreseptor jantung sehingga dapat meningkatkan cardiac output yang dapat meningkatkan tekanan darah. Mekanisme tersebut berkaitan dengan potensial aksi dalam hal pembukaan channel ion Na+ dan K+. Selain itu, peningkatan aktivitas saraf simpatik juga dapat meningkatkan aktivitas α -1 pada otot polos, yang akan meningkatkan resistensi perifer, sehingga dapat mempengaruhi elastisitas pembuluh darah yang dapat meningkatkan kontraksi otot
jantung untuk memompa darah, dan pada akhirnya akan meningkatkan tekanan darah. Penurunan tekanan darah juga akan meningkatkan aliran tekanan darah yang akan mengakibatkan pelepasan protease renin dan pemecahan dekapeptida angiotensin I menjadi angiotensin II. Peningkatan angiotensin II dapat meningkatkan produksi aldosteron yang dapat meningkatkan retensi natrium dan air. Dengan demikian volume darah akan meningkat dan pada akhirnya menyebabkan peningkatan tekanan darah (Akil & Bakri 2001; Yusuf 2008).
Captopril merupakan inhibitor spesifik yang pertama kali ditemukan pada tahun 1975 oleh Chusman dan Ondetti dan merupakan inhibitor kompetitif yang penting untuk menghambat aktivitas ACE. Captopril ((2S)-1-[(2S)-2-methyl-3-sulfanyl propanoyl] pyrrolidine-2-carboxylic acid) merupakan obat hipertensi yang berasal dari sintesis kimiawi. Mishra (2011) melaporkan bahwa sintesis captopril pertama kali diturunkan dari bisa ular. Inhibitor ini dapat menginaktivasi pengonversian dari angiotensin I menjadi angiotensin II (Lima 1999).
Penelitian ini menggunakan captopril (komersial) sebagai kontrol positif aktivitas penghambatan ACE. Persentase aktivitas penghambatan ACE oleh captopril (C1) sebesar 61,46% dan captopril (C2) sebesar 66,67% (Gambar 6 & 7) menunjukkan persentase penghambatan yang lebih rendah dibandingkan dengan aktivitas penghambatan ACE aktinomiset endofit. Persentase aktivitas penghambatan ACE oleh captopril yang ditunjukkan oleh Hayes et al. (2007) sebesar 100% dengan konsentrasi captopril yang digunakan 0,005 mg/mL. Nilai persentase tersebut lebih tinggi dibandingkan dengan hasil yang diperoleh pada penelitian ini. Perbedaan persentase aktivitas penghambatan ACE tersebut kemungkinan disebabkan oleh perbedaan kemurnian captopril. Captopril yang digunakan Hayes et al. (2007) berasal dari Sigma Co. yang memiliki kemurnian lebih tinggi dibandingkan dengan captopril komersial yang mengandung berbagai komponen bahan baku campuran obat.
Media cair ISP2 (kontrol negatif) yang digunakan sebagai media produksi filtrat menunjukkan aktivitas penghambatan yang sangat rendah (26,04%) dibandingkan kisaran aktivitas penghambatan yang ditunjukkan oleh isolat aktinomiset endofit pada penelitian ini (103,13%-279,17%).
10
menunjukkan aktivitas penghambatan ACE yang bervariasi berkisar antara 140,63%-279,17%. Isolat AEP-1 menunjukkan aktivitas penghambatan ACE yang paling tinggi (279,17%) di antara ke-3 isolat lainnya, serta melebihi aktivitas penghambatan ACE yang ditunjukkan oleh ekstrak daun pegagan (198,96%). Hal tersebut mengindikasikan bahwa ketiga isolat aktinomiset endofit asal daun pegagan mampu menghasilkan senyawa inhibitor ACE. Hasil uji in vitro terhadap tanaman kultur jaringan pegagan umur 55 hari (steril mikrob) ternyata tidak menunjukkan aktivitas penghambatan ACE. Hal ini menguatkan dugaan bahwa aktinomiset endofit pada pegagan berperan untuk menghasilkan senyawa inhibitor ACE.
Streptomyces spp. maupun non-Streptomyces asal buah belimbing wuluh menunjukkan persentase aktivitas penghambatan ACE yang bervariasi antara 0% hingga 222,92%. Isolat AEB-5 menunjukkan aktivitas penghambatan ACE tertinggi (222,92%) melebihi aktivitas penghambatan ACE yang dihasikan oleh ekstrak buah belimbing wuluh (166,67%). Hal ini mengindikasikan bahwa aktinomiset endofit asal buah belimbing wuluh juga mampu menghasilkan senyawa inhibitor ACE. Aktivitas penghambatan ACE aktinomiset endofit isolat AEP-1 asal daun pegagan menunjukkan aktivitas penghambatan yang lebih tinggi dibandingkan dengan isolat AEB-5 asal buah belimbing wuluh. Bobot biomassa kedua isolat tersebut berbeda, meskipun pada kondisi pertumbuhan yang sama. Bobot biomassa isolat AEB-5 (5,263 mg/mL) lebih besar dibandingkan dengan bobot biomassa yang dihasilkan oleh isolat AEP-1 (0,720 mg/mL). Namun demikian, aktivitas penghambatan ACE isolat AEP-1 lebih tinggi dibandingkan dengan kemampuan penghambatan ACE isolat AEB-5. Perbedaan kemampuan aktivitas penghambatan ACE dapat disebabkan oleh perbedaan kemampuan masing-masing isolat. Aktinomiset endofit yang digunakan untuk uji penghambatan ACE pada penelitian ini berupa ekstrak kasar yang masih mengandung beragam komponen media. Ekstrak kasar aktinomiset endofit tersebut kemungkinan juga mengandung beragam senyawa bioaktif lain yang dihasilkan oleh masing-masing isolat, sehingga dapat berpengaruh terhadap aktivitas penghambatan ACE.
Kemampuan mikrob endofit yang dapat menghasilkan senyawa bioaktif yang sama dengan tanaman inangnya diharapkan
dapat mengurangi kebutuhan bahan baku obat berbasis biomassa tanaman. Produksi senyawa bioaktif sebagai antihipertensi yang dihasilkan oleh aktinomiset endofit akan lebih cepat dan efisien dibandingkan dengan produksi senyawa aktif menggunakan tanaman inangnya. Penelitian ini menunjukkan bahwa pegagan dan belimbing wuluh yang secara empiris telah dikenal sebagai obat hipertensi berasosiasi dengan aktinomiset endofit yang mempunyai kemampuan menghasilkan senyawa inhibitor ACE sehingga dapat dikembangkan lebih lanjut sebagai bahan baku obat hipertensi.
SIMPULAN DAN SARAN
Simpulan
Total isolat aktinomiset endofit yang berhasil diisolasi berjumlah 12 isolat, tiga isolat berasal dari daun pegagan (Centella asiatica) yang termasuk Streptomyces spp., sedangkan sembilan isolat lainnya berasal dari buah belimbing wuluh (Averrhoa bilimbi), enam isolat termasuk Streptomyces spp. dan tiga isolat lainnya termasuk non-Streptomyces. Ekstrak kasar isolat aktinomiset endofit yang diuji menunjukkan aktivitas inhibitor ACE yang beragam berkisar antara 103,13% - 279,17%. Isolat AEP-1 asal daun pegagan menunjukkan aktivitas penghambatan ACE tertinggi (279,17%), diikuti isolat AEB-5 asal buah belimbing wuluh (222,92%). Aktivitas inhibisi kedua isolat tersebut melebihi aktivitas inhibisi kontrol positif captopril (0,01 mg/mL) 61,46% dan captopril (0,02 mg/mL) 66,67%. Sementara itu, tanaman kultur jaringan pegagan steril mikrob endofit tidak memiliki aktivitas inhibisi ACE. Dengan demikian isolat AEP-1 dan AEB-5 berpotensi sebagai penghasil inhibitor ACE dan dapat dikembangkan lebih lanjut sebagai obat hipertensi.
Saran
11
DAFTAR PUSTAKA
Akil MN, Bakri S. 2001. Angiotensin converting enzyme inhibitor (ACE-I) dan proteksi vaskular. Cermin Dunia Kedokteran 132:7-9.
Castillo et al. 2002. Munumbicins, wide spectrum antibiotics produced by Streptomyces NRRL 30562, endophytic on Kennedia nigriscans. Microbiology 148:2675-2685.
Cheung H, Wang FL, Ondetti MA, Sabo EF, Chusman DW. 1980. Binding of peptide substrates and inhibitors of angiotensin-converting enzyme. J Biol Chem 255:401-407.
Coombs JT, Franco CMM. 2003. Isolation and identification of actinobacteria from surface-sterilized wheat roots. Appl Environ Microbiol 69:5603-5608.
Crawford DL, Lynch JM, Whipps JM, Ousley MA. 1993. Isolation and characterization actinomycete antagonists of a fungal root phatogen. Appl Environ Microbiol 59:3899-3905.
Ghadin N et al. 2008. Isolation and characterization of novel endophytic Streptomyces SUK 06 with antimicrobial activity from Malaysian plant. Asian J Plant Sci 7:189-194.
Hallmann J, Quadt-Hallmann A, Mahaffee WF, Kloepper JW. 1997. Bacterial endophytes in agricultural crops. Can J Microbiol 43:895-914.
Hansen K et al. 1995. In vitro screening of traditional medicines vfor antihypertensive effect based on inhibition of the angiotensin converting enzyme (ACE). J Ethnopharmacol 48:43-51.
Hariana HA. 2004. Tumbuhan Obat dan Khasiatnya. Jakarta: Penebar Swadaya. Hasegawa S, Meguro A, Shimizu M,
Nishimura T, Kunoh H. 2006. Endophytic actinomycetes and their interaction with host plants. Actinomycetologica 20:72-81. Hayakawa MT, Nonomura H. 1987. Humic
acid-vitamin agar, a new method for the selective isolation of soil actinomycetes. J Ferment Bioeng 65:501-509.
Hayes M et al.. 2007. Fermentate of Lactobacillus animalis DPC6134 contains of range of novel propeptide angiotensin converting enzyme inhibitor. J Appl Environ Microbial 73:4658-4667.
Iskandar Y. 2007. Tanaman obat yang berkhasiat sebagai antihipertensi [karya ilmiah]. Bandung: Fakultas Farmasi Universitas Padjajaran Bandung.
Kasahara Y, Ashihara Y. 1981. Colorimetry of angiotensin-I converting enzyme activity in serum. Clin Chem 27:1922-1925.
Kudo T. 1997. Family Streptomycetaceae. In: Miyadoh S (ed). Atlas of Actinomycetes. The Society for Actinomycetes Japan. p 122-155.
Lachevalier MP, Bievre C de, Lachevalier HA. 1977. Chemotaxonomy of aerobic actinomycetes: phospholipid composition. Biochem Syst Ecol 5:249-260.
Lestari Y. 2006. Identification of indigenous Streptomyces spp. producing antibacterial compounds. J Mikrobiol Indones 11:99-101.
Lima DP de. 1999. Synthesis of angiotensin-converting enzyme (ACE) inhibitors: an important class of antihipertensive drugs. Quim Nova. 22:375-381.
Mishra BB. 2011. Natural products in drug discovery: clinical evaluations and investigations. Di dalam: Tiwari VK, editor. Opportunity, Challenge and Scope of Natural Products in Medicinal Chemistry. Ed ke-2. India: Research Signpost. hlm 1-62.
Miyadoh S. 1997. Morphology and Phylogeny of Actinomycetes. Atlas of Actinomycetes. The Society for Actinomycetes Japan. Miyadoh S, Otoguro M. 2004. Workshop on
isolation methods and classification of actinomycetes [makalah]. Biotechnology Center LIPI.
Nakatsukasa WM, Wilgus RM, Thomas DN, Mertz FP, Boeck LD. 1985. Angiotensin converting enzyme inhibitors produced by Streptomyces chromofuscus. Discovery, taxonomy and fermentation. J Antibiot 38:997-1002.
Ondetti MA, Cushman DW, Rubin B, Bindra JS, Lednicer D, editor. Ed ke-2. 1983. In Chronicles of Drugs Discovery. New York: John Wiley & Sons,Inc.
Raja A, Prabakarana P. 2011. Actinomycetes and drug-an overview. Am J Drug Discov Dev 1:75-84.
Sharma PK, Sarita S, Prell J. 2005. Isolation and characterization of an endophytic bacterium related to Rhizobium/Agrobacte rium from wheat (Triticum aestivum L.) roots. Current Sci 89:608-610.
12
Strobel G, Daisy B. 2003. Bioprospecting for microbial endophytes and their natural product. Microbiol Mol Biol Rev 67:491-502.
Takahashi Y, Omura S. 2003. Isolation of new actinomycete strains for screening of new bioactive compounds. J Gen Appl Microbiol 49:141-154.
Tan RX, Zou WX. 2001. Endophytes: a rich source of functional metabolites. Nat Prod Rep 18:448-459.
Tejesvi MV, Kini KR, Prakash HS, Subbiah V, Shetty HS. 2008. Antioxidant, antihypertensive, and antibacterial pro-
perties of endophytic Pestalotiopsis species from medicinal plants. Can J Microbiol 54:769-780.
Wagner H, Elbl G, Lotter H, Uinea M. 1991. Evaluation of natural products as inhibitors of angiotensin I-converting enzyme (ACE). Pharm Pharmacol Lett 1:15-18.
Wijayakusuma H, Dhalimartha S. 2005. Ramuan Tradisional untuk Pengobatan Darah Tinggi. Bogor Penebar Swadaya. Yusuf I. 2008. Hipertensi sekunder.
14
14
Lampiran 1 Komposisi media agar-agar asam humat mengandung vitamin B (HV) Komposisi bahan Jumlah bahan (g) per liter media
Asam humat 1,00*
CaCO3 0,02
FeSO4.7H2O 0,01
KCl 1,71
MgSO4.7H2O 0,05
Na2HPO4 0,50
B-vitamin 5,00 mL**
Sikloheksimida 0,05
Asam nalidiksat 0,02***
Agar-agar murni 18,00
Akuades 1,00 L
Ket : *) Dilarutkan dalam 40 mL 0,4% NaOH; **) Menggunakan satu butir vitamin B complex IPI, dilarutkan dalam 200 mL akuades steril (disaring menggunakan millipore 0,22 µm); ***) Dilarutkan dalam 1 mL NaOH 0,4% + 3 mL akuades steril. pH media HV adalah 7.2.
Lampiran 2 Komposisi media agar-agar International Streptomyces Project No.2 (ISP2) Komposisi bahan Jumlah bahan (g) per liter media
Yeast extract 4
Malt extract 10
Glukosa 4
Agar-agar 18
Ket : Media cair ISP2 tidak menggunakan komposisi agar-agar.
Lampiran 3 Komposisi media Murashige & Skoog (MS)
Bahan kimia Konsentrasi larutan (g/L)
Pemakaian larutan media (ml/L)
NH4NO3 82,500 20
KNO3 95,000 20
KH2PO4 34,000 5
H3BO3 1,240
KI 0,166
Na2MoO4.2H2O 0,050
CoCl2.6H2O 0,005
CaCl.2H2O 88,000 5
MgSO4.7H2O 74,000 5
MnSO4.4H2O 4,460
ZnSO4.7H2O 1,720
CuSO4.5H2O 0,005
Na2EDTA.2H2O 3,730 10
FeSO4.7H2O 2,780
Myo-Inositol 10,000 10
Thiamine 0,010 10
Niacine 0,050
Pyridoxine 0,050
Glycine 0,200
15
Lampiran 4 Nilai absorbansi hasil pengukuran spektrofotometri λ 228 nm
Jenis sampel
Absorbansi Persentase (%)
aktivitas penghambatan
ACE A1 A2
Rata-rata B1 B2
Rata-rata C1 C2
Rata-rata D1 D2
Rata-rata
16
Lampiran 5 Contoh perhitungan penentuan persentase aktivitas penghambatan ACE
Keterangan dari rumus persamaan tersebut :
A = Hasil absorbansi dengan ACE, tanpa inhibitor ACE B = Hasil absorbansi tanpa ACE, dan tanpa inhibitor ACE C = Hasil absorbansi dengan ACE dan inhibitor ACE
D = Hasil absorbansi dengan inhibitor ACE, namun tanpa ACE
Contoh perhitungan:
1). Kontrol positif captopril (0,01 mg/mL)
Aktivitas inhibitor ACE (%) = 100 – [100 x (0,3600 – 0,3415) / (0,5410 – 0,4930)] = 100 – [100 x (0,0185) / (0,048)]
= 100 – (38,54) = 61,46 %
2). Kontrol positif captopril (0,02 mg/mL)
Aktivitas inhibitor ACE (%) = 100 – [100 x (0,3500 – 0,3340) / (0,5410 – 0,4930)] = 100 – [100 x (0,016) / (0,048)]
= 100 – (33,33) = 66,67 %
3). Isolat AEP-1 (0,058 mg/mL)
Aktivitas inhibitor ACE (%) = 100 – [100 x (0,4345 – 0,5205) / (0,5410 – 0,4930)] = 100 – [100 x (- 0,086) / (0,048)]
= 100 – (- 179,17) = 279,17 %
4). Isolat AEB-5 (0,421 mg/mL)
Aktivitas inhibitor ACE (%) = 100 – [100 x (0,5135 – 0,5725) / (0,5410 – 0,4930)] = 100 – [100 x (- 0,059) / (0,048)]
= 100 – (- 122,92) = 222,92 %