KINERJA PERTUMBUHAN DAN RESPONS IMUN NON
SPESIFIK UDANG VANAME Litopenaeus vannamei YANG
DIBERI PAKAN DENGAN FEED ADDITIVE β-(1,3) GLUKAN
DAN POLI-β- HIDROKSIBUTIRAT
SARMIN
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2014
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Kinerja pertumbuhan dan respons imun non spesifik udang vaname Litopenaeus vannamei yang diberi pakan dengan feed additive β-(1,3) glukan dan poli-β-hidroksibutirat adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, November 2014 Sarmin NRP C151120511
RINGKASAN
SARMIN. Kinerja pertumbuhan dan respons imun non spesifik udang vaname Litopenaeus vannamei yang diberi pakan mengandung feed additive β-(1,3) glukan dan poli-β-hidroksibutirat. Dibimbing oleh MUHAMMAD AGUS SUPRAYUDI dan DEDI JUSADI.
Penelitian dengan tiga ulangan ini dilakukan untuk menguji pakan dengan penambahan feed additive berupa β-(1,3) glukan dan poli-β-hidroksibutirat terhadap kinerja pertumbuhan dan respons imun non spesifik udang vaname Litopenaeus vannamei. Juvenil udang dengan bobot awal 2,06±0,03 g dipelihara dalam 12 unit akuarium 90x40x35 cm3 yang diisi 90 L air laut. Udang dengan kepadatan awal 20 ekor/akuarium dipelihara selama 42 hari menggunakan sistem resirkulasi dengan debit air 2,5 L/menit. Selama masa budidaya udang diberi empat jenis pakan yang berbeda, yaitu yang ditambah 0,15% β-(1,3) glukan, 1% poli-β-hidroksibutirat, kombinasi 0,15% dan 1% poli-β-hidroksibutirat, atau tanpa β-(1,3) glukan dan poli-β-hidroksibutirat (kontrol). Jumlah pakan yang diberikan didasarkan pada persentase biomassa, dengan frekwensi pemberian pakan 4 kali sehari.
Hasil penelitian menunjukkan bahwa udang yang diberi pakan mengandung 0,15% β-(1,3) glukan memiliki kinerja pertumbuhan, meliputi laju pertumbuhan spesifik, konversi pakan, retensi protein dan sintasan, yang terbaik. Di sisi lain pemberian 1% poli-β-hidroksibutirat dalam pakan memiliki dampak yang kurang baik terhadap kinerja pertumbuhan udang; hal ini terlihat dari lebih rendahnya laju pertumbuhan dan sintasan udang di perlakuan tersebut dibandingkan dengan kontrol. Kinerja pertumbuhan udang di setiap perlakuan berkorelasi dengan respons imun non spesifik udang. Udang yang diberi 0,15% β-(1,3) glukan secara signifikan memiliki nilai total haemocyte count (THC) dan nilai respiratory burst (RB) yang paling tinggi. Sedangkan udang yang diberi 1% poli-β-hidroksibutirat memiliki kadar THCyang paling rendah; bahkan lebih rendah dari udang kontrol. Pemberian β-(1,3) glukan yang dikombinasikan dengan poli-β-hidroksibutirat pada udang tidak mampu meningkatkan kadar THC melebihi perlakuan kontrol. Dengan demikian, penambahan 0,15% β-(1,3) glukan secara tunggal dalam pakan dapat meningkatkan respons imun non spesifik dan kinerja pertumbuhan udang vaname.
Kata kunci: Pertumbuhan, udang, respons imun non spesifik, Litopenaeus vannamei.
SUMMARY
SARMIN. Growth performance and non-specific imune response of Litopenaeus vannamei fed on the diet supplemented with β-(1,3) glucan dan poly-β-hydroybutyrate. Supervised by MUHAMMAD AGUS SUPRAYUDI dan DEDI JUSADI.
A triplicate experiment was conducted to determine the effect of the diet supplemented with β-(1,3) glucan dan poly-β-hydroybutyrate on growth performance and non-specific imune response of Litopenaeus vannamei. Twenty shrimps with an initial body weight of 2.06 ± 0.03 g were stocked in 90x40x35 cm3 of recirculating aquarium with water flow at 2.5 l/min. During rearing period, shrimp were fed on the diet supplemented with either 0.15 % β-(1,3) glucan, 1 % poly-β-hydroybutyrate, 0.15 % β-(1,3) glucan and 1 % poly-β-hydroybutyrate, or without β-(1,3) glucan nor 1 % poly-β-hydroybutyrate (control). The shrimp were fed on the diet at 4-8 % biomass, four times a day.
Results showed that shrimp fed on the diet supplemented with 0.15 % β-(1,3) glucan had the best growth performance, including specific growth rate, feed conversion ratio, protein retention dan survival rate. On the other hand, shrimp fed on the diet supplemented with 1 % poly-β-hydroybutyrate had poor specific growth and survival rate, worse than control. The growth performance of shrimp in all treatments related with the status of non-specific response imune. Shrimp fed on the diet supplemented with 0.15 % β-(1,3) glucan had significantly highest values of total haemocyte count (THC) and respiratory burst (RB). While those shrimp fed on the diet supplemented with 1 % poly-β-hydroybutyrate had the lowest values of THC and RB. Feeding on the diet supplemented with 0.15 % β-(1,3) glucan and 1 % poly-β-hydroybutyrate resulted into the similar THC and RB values of shrimp with those in control. Thus, supplementation of 0.15% β-(1,3) glucan into the diet, not poly-β-hydroybutyrate, increase non-specific response imune and growth performance of srimp.
Keywords: Growth, shrimp, non-specific immune response, Litopenaeus vannamei.
© Hak Cipta Milik IPB, Tahun 2014
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis ini dalam bentuk apa pun tanpa izin IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Ilmu Akuakultur
KINERJA PERTUMBUHAN DAN RESPONS IMUN NON
SPESIFIK UDANG VANAME Litopenaeus vannamei YANG
DIBERI PAKAN DENGAN FEED ADDITIVE β-(1,3) GLUKAN
DAN POLI-β- HIDROKSIBUTIRAT
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2014
SARMIN
Judul Tesis : Kinerja pertumbuhan dan respons imun non spesifik udang vaname Litopenaeus vannamei yang diberi pakan dengan feed additive β-(1,3) glukan dan poli-β-hidroksibutirat.
Nama : Sarmin NRP : C151120511
Disetujui oleh Komisi Pembimbing
Dr Ir Muhammad Agus Suprayudi, MSi Ketua
Dr Ir Dedi Jusadi, MSc Anggota
Diketahui oleh
Ketua Program Studi IlmuAkuakultur
Dr Ir Widanarni, MSi
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MSc Agr
PRAKATA
Puji syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya, sehingga penelitian dan penulisan tesis ini berhasil diselesaikan. Penelitian ini berjudul Kinerja pertumbuhan dan respons imun non spesifik udang vaname Litopenaeus vannamei yang diberi pakan dengan feed additive β-(1,3) glukan dan poli-β-hidroksibutirat. Pada kesempatan ini penulis mengucapkan terimakasih kepada Dr Ir Muhammad Agus Suprayudi, MSi dan Dr Ir Dedi Jusadi, MSc. selaku tim komisi pembimbing atas arahan, bimbingan dan masukan-masukannya sejak penyusunan rencana penelitian sampai penyusunan tesis ini. Julie Ekasari, SPi, MSi, Ph.D selaku ketua proyek yang telah membimbing dan memberikan arahan secara langsung dilapangan. Dr Ir Widanarni, MSi selaku ketua Program Studi Ilmu Akuakultur atas arahan, masukan dan perbaikan tesis ini. Dr Ir Mia Setiawati, MSi selaku penguji luar komisi, atas arahan dan masukan untuk perbaikan dalam penyusunan tesis ini.
Penulis juga mengucapkan terimakasih kepada bapak Mardi selaku teknisi di Lab terpadu Ilmu Kelautan IPB Ancol serta Alm. Ibu Ince, Erfan dan Erfin atas bantuan serta masukannya selama penelitian. Teknisi Laboratorium BDP IPB; Bapak Ranta (Lab Kesehatan Ikan FPIK IPB), Bapak Jajang (Lab Lingkungan FPIK IPB), Bapak Wasjan dan mbak Retno (Lab Nutrisi FPIK IPB) yang telah membantu penulis selama melakukan analisa laboratorium. Rekan-rekan yang telah membantu selama penelitian berlangsung: Titi, Nurlita, Dani dan asep Semua Rekan-rekan mahasiswa Program Mayor Ilmu Akuakultur angkatan 2012 atas kebersamaan dan kerjasama yang baik serta bantuannya dalam perkuliahan, penelitian dan penyelesaian karya ilmiah ini. Tidak lupa kepada kedua orang tua ku atas doa dan ridhonya, sehingga semua jenjang pendidikan yang kutempuh berjalan dengan lancar sesuai yang diharapkan, serta kaka-kakaku yang tercinta. Keluarga besar “Aljabar Al Islamiyyah” staf manajemen, dewan guru dan siswa-siswi bimbingan Al jabar.
Penelitian ini merupakan bagian dari penelitian yang berjudul “Enhancement of nutrient efficiency in integrated multi-trophic aquaculture with bioflocs as nutrient converting agent” diketuai oleh Julie Ekasari, SPi, MSi, Ph.D yang dibiayai oleh program ICP Ph.D. Scholarship Vlaamse Interuniversitaire Raad, Belgia.
Bogor, 24 November 2014 Sarmin
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vii
1 PENDAHULUAN 1 Latar Belakang 1 Perumusan Masalah 1 Tujuan Penelitian 2 Manfaat Penelitian 2 2 TINJAUAN PUSTAKA 2 Sistem Imun 2 β-(1,3) glukan 3 Poli-β-hidroksibutirat 3 3 METODE 4 Pakan Uji 4 Pemeliharaan 4 Parameter 5
4 HASIL DAN PEMBAHASAN 8
Hasil 8
Pembahasan 9
5 KESIMPULAN DAN SARAN 11
Kesimpulan 11
Saran 11
DAFTAR PUSTAKA 12
LAMPIRAN 15
DAFTAR TABEL
1 Formulasi bahan pakan (%) yang digunakan dalam pembuatan pakan uji dan
komposisi proksimatnya (%)...4
2 Kondisi air pemeliharaan udang vaname...5
3 Kinerja pertumbuhan udang vaname...8
4 Respons imun non spesifik udang vaname...8
DAFTAR LAMPIRAN
1 ANOVA dan Uji-Duncan pertumbuhan udang vaname yang diberi feed additive (1,3) glukan 0,15%; Poli-βhidroksibutirat (PHB) 1%; kombinasi β-(1,3) glukan 0,15% dan PHB 1% serta kontrol...152 ANOVA dan Uji-Duncan konversi pakan udang vanname yang diberi feed additive (1,3) glukan 0,15%; Poli-βhidroksibutirat (PHB) 1%; kombinasi β-(1,3) glukan 0,15% dan PHB 1% serta kontrol...16
3 ANOVA dan Uji-Duncan sintasan udang vaname yang diberi feed additive β-(1,3) glukan 0,15%; Poli-βhidroksibutirat (PHB) 1%; kombinasi β-β-(1,3) glukan 0,15% dan PHB 1% serta kontrol...17
4 ANOVA dan Uji-Duncan retensi protein udang vaname yang diberi feed additive (1,3) glukan 0,15%; Poli-βhidroksibutirat (PHB) 1%; kombinasi β-(1,3) glukan 0,15% dan PHB 1% serta kontrol...18
5 ANOVA dan Uji-Duncan total haemocyte count (THC) udang vaname yang diberi feed additive β-(1,3) glukan 0,15%; Poli-βhidroksibutirat (PHB) 1%; kombinasi β-(1,3) glukan 0,15% dan PHB 1% serta kontrol...19
6 ANOVA dan Uji-Duncan respiratory burst (RB) udang vaname yang diberi feed additive β-(1,3) glukan 0,15%; Poli-βhidroksibutirat (PHB) 1%; kombinasi β-(1,3) glukan 0,15% dan PHB 1% serta kontrol...20
1 PENDAHULUAN
Latar Belakang
Budidaya udang vaname secara intensif memerlukan pergantian air yang banyak dan tidak mudah dilakukan karena memerlukan biaya yang besar. Pergantian air ini juga dapat menyebabkan udang vaname stress dan beresiko terhadap masuknya penyakit baik bakteri maupun virus yang dapat menyebabkan infeksi. Teknologi bioflok (BFT) dengan pergantian air yang terbatas ternyata dapat mengurangi, bahkan menghilangkan ancaman infeksi mikroba air tersebut masuk ke dalam media budidaya dan tetap menjaga kualitas air ( Horowitz 2002). Teknologi bioflok dapat memanfaatkan limbah organik baik yang berasal dari limbah metabolisme maupun sisa-sisa pakan untuk pertumbuhan bakteri heterotrof dengan C/N ratio 10-20. Bakteri tersebut menjadi pakan tambahan yang dapat menghasilkan energi untuk meredam stress, sehingga udang vaname menjadi sehat dan dapat tumbuh dengan baik. Bioflok juga dapat meningkatkan sintasan udang vaname karena dibioflok terdapat poli-β hidroksibutirat yang berfungsi menyediakan energi (De Schryver dan Verstraete 2009) dan β-(1,3) glukan yang berfungsi mengaktifkan sel fagosit untuk melakukan fagositosis (Hunter et al. 2002).
Pada penelitian ini produk-produk penyusun bioflok yang berasal dari dinding sel bakteri seperti β-(1,3) glukan (BG) dan poli-β hidroksibutirat (PHB)
dicobakan secara langsung sebagai feed additive. Feed additive merupakan bahan pakan tambahan yang diberikan melalui pencampuran dengan bahan-bahan pakan yang lain. β-(1,3) glukan adalah feed additive yang diikat melalui ikatan β-(1,3) glukosida (Ha et al. 2002) dan banyak ditemukan pada dinding sel bakteri, dan khamir (Hunter et al. 2002). β-(1,3) glukan (BG) dapat meningkatkan kemampuan sel darah putih untuk menghancurkan benda asing yang ada pada tubuh dan melindungi mamalia dari infeksi penyakit (Vetvicka et al. 2002), β-(1,3) glukan (BG) juga dapat mengaktifkan mekanisme pertahanan non spesifik dan meningkatkan daya tahan terhadap penyakit infeksi dengan meningkatkan jumlah sel darah (Sakai 1999). Feed additive lain yang digunakan dalam penelitian ini adalah poli-β-hidroksibutirat (PHB). Poli-β-hidroksibutirat (PHB) adalah feed additive yang dapat diproduksi oleh Bacillus dan Lactobacillus (Anderson dan Dawes 1990). Poli-β-hidroksibutirat (PHB) dapat mengaktifkan mekanisme pertahanan non spesifik dengan cara melepaskan asam lemak yang dapat menyediakan energi bagi udang (Defoirdt et al. 2007).
Perumusan Masalah
Sistem pemeliharaan udang secara intensif memerlukan pergantian air yang banyak, sehingga dapat menyebabkan udang vaname menjadi stress dan beresiko terhadap masuknya penyakit baik bakteri maupun virus yang dapat menyebabkan infeksi. Teknologi bioflok (BFT) dengan pergantian air yang terbatas ternyata dapat menghasilkan energi untuk meredam stress dan mengurangi resiko masuknya penyakit yang dapat menyebabkan infeksi pada udang vaname. Pada bioflok juga terdapat poli-β-hidroksibutirat (PHB) dan β-(1,3) glukan yang dapat meningkatkan pertumbuhan dan sintasan udang vaname. Penyusun bioflok yang
2
berupa β-(1,3) glukan (BG) dan poli-β-hidroksibutirat (PHB) dalam penelitian ini digunakan sebagai feed additive untuk meningkatkan pertumbuhan dan respons imun non spesifik udang vaname.
Tujuan Penelitian
Penelitian ini bertujuan mengkaji efektifitas β-(1,3) glukan, poli-β-hidroksibutirat serta kombinasinya untuk meningkatkan kinerja pertumbuhan dan respons imun non spesifik udang vaname.
Manfaat Penelitian
Manfaat penelitian ini diharapkan dapat memberikan rekemondasi pemberian feed additive seperti β-(1,3) glukan, poli-β-hidroksibutirat serta kombinasinya yang efektif dalam pakan untuk meningkatkan kinerja pertumbuhan dan respons imun non spesifik udang vaname.
2 TINJAUAN PUSTAKA
Sistem Imun UdangMenurut Ramu and Zacharia (2000), mekanisme imun krustasea bersifat non spesifik atau kurang bisa mengembangkan sistem imun spesifik dimana memorinya sangat lemah dibandingkan vertebrata tingkat tinggi. Siwicki et al. (1998) menyatakan bahwa imunostimulan dapat mengaktifkan mekanisme pertahanan non spesifik dan respons imun spesifik. Imunostimulan meningkatkan daya tahan terhadap penyakit infeksi, dengan meningkatkan mekanisme pertahanan non spesifik (Sakai 1999).
Sistem imun non spesifik ini dapat dirangsang oleh glukan (Jorgensen and Robertsen 1995). Parameter imun diantaranya meliputi perhitungan total hemosit count (THC) dan respiratory burst (RB). Nilai THC dan RB berkaitan dengan pembentukan sel fagosit pada hemosit udang (Rodriguez dan Le Moullac 2000). Hemosit berperan penting pada pertahanan tubuh krustasea yaitu dapat menghancurkan partikel asing yang masuk ke tubuh udang yang meliputi tahap pengenalan dan fagositosis (Johansson et al. 2000).
Respiratory burst (RB) merupakan salah satu parameter imun udang berkaitan dengan reaksi fagositosis yang merupakan reaksi yang paling umum dalam pertahanan selular udang. Proses fagositosis dimulai dengan perlekatan (attachment) dan penelanan (ingestion) partikel mikroba ke dalam sel fagosit. Sel fagosit kemudian membentuk vakuola pencernaan (digestive vacuola) yang disebut fagosom (Rodriquez dan Le Moullac 2000). Lisosom (granula dalam sitoplasma fagosit) kemudian bersatu dengan fagosom membentuk fagolisosom. Mikroorganisme selanjutnya dihancurkan oleh enzim lisozim dan sisa-sisa mikroba yang telah dihancurkan dikeluarkan dari dalam sel melalui proses egestion. Pemusnahan partikel mikroba yang difagosit melibatkan pelepasan enzim lisozim ke dalam fagosom dan produksi respiratory burst (percepatan respirasi)(Rodriquez dan Le Moullac 2000).
3 β-(1,3) glukan
β-(1,3) glukan merupakan polisakarida yang diikat melalui ikatan β-(1,3) glukosida (Ha et al. 2002) dan banyak ditemukan pada dinding sel bakteri dan khamir (Hunter et al. 2002). Saccharomyces cerevisiae termasuk khamir uniseluler yang tersebar luas di alam dan merupakan galur potensial penghasil β-(1,3) glukan, karena sebagian besar dinding selnya tersusun atas β-glukan (Lee et al. 2001). β-(1,3) glukan memiliki aktivitas biologis sebagai peningkat sistem imun (Kulickle et al. 1996). β-(1,3) glukan dapat meningkatkan kemampuan sel darah putih dalam menghancurkan benda asing yang ada pada tubuh dan melindungi mamalia dari infeksi penyakit (Vetvicka et al. 2002).
β-(1,3) glukan dapat meningkatkan respons imun udang dengan cara meningkatkan aktivitas sel-sel fagosit untuk menjalankan proses fagositosis (Yin et al. 2006). Meningkatnya respons imun udang tersebut dapat mempengaruhi nilai sintasan menjadi lebih tinggi, karena resistensi udang terhadap patogen juga meningkat (Cook et al. 2003). β-(1,3) glukan terbukti secara ilmiah sebagai bahan yang tidak memiliki toksisitas atau efek samping (Ber 1997). β-(1,3) glukan dalam prosesnya bekerja dengan cara mengikat molekul reseptor yang terdapat pada permukaan sel fagosit. Pada saat reseptor diikat oleh β-(1,3) glukan, sel fagosit menjadi lebih aktif dalam melakukan fagositosis terhadap partikel asing. Pada saat bersamaan mereka mengeluarkan molekul-molekul sinyal yang merangsang pembentukan sel hemosit yang baru (Rodriguez dan Le Moullac 2000).
Poli-β-hidroksibutirat
Poli-β-hidroksibutirat (PHB) merupakan polimer intraseluler yang dihasilkan oleh mikroorganisme sebagai sumber karbon dan penyimpanan energi yang dapat meningkatkan sistem imun (Defoirdt et al. 2007). Poli-β-hidroksibutirat (PHB) dapat diproduksi oleh Bacillus dan Lactobacillus (Anderson dan Dawes 1990). Poli-β-hidroksibutirat (PHB) merupakan polimer yang melepaskan asam lemak rantai pendek, yang dapat menyediakan sumber energi ekstra, namun tidak menyediakan asam lemak rantai panjang yang diperlukan untuk metabolisme. Degradasi poli-β-hidroksibutirat (PHB) dapat terjadi melalui mekanisme enzimatik (Defoirdt et al. 2009). Poli-β-hidroksibutirat (PHB) dapat melindungi organisme akuatik dari serangan bakteri patogen (De Schryver et al. 2010). Poli-β-hidroksibutirat (PHB) akan terdegradasi menjadi asam lemak butirat yang merupakan asam lemak rantai pendek oleh enzim depolimerase. Asam lemak rantai pendek tersebut akan diubah menjadi Asil-CoA kemudian akan masuk ke dalam mitokondria.
Reaksi ß-oksidasi berlangsung dalam 4 tahap yaitu dehidrogenasi I, hidratasi, dehidrogenasi II, dan tiolasi. Dehidrogenasi I yaitu dehidrogenasi asam lemak-CoA yang sudah berada di dalam mitokondrion oleh enzim acyl-CoA dehidrogenase, menghasilkan senyawa enoyl-CoA. Reaksi ini, FAD sebagai koenzim direduksi menjadi FADH2. Satu molekul FADH2 menghasilkan dua
molekul ATP. Hidratasi, yaitu ikatan rangkap pada enoyl-CoA dihidratasi menjadi 3-hidroxyacyl-CoA oleh enzim enoyl-CoA hidratase. Dehidrogenase II, yaitu dehidrogenasi 3- hidroxyacyl-CoA oleh enzim ß-hidroxyacyl-CoA dehidrogenase dengan NAD sebagai koenzimnya menjadi ß-ketoacyl-CoA. NADH yang terbentuk dari NAD+ menghasilkan tiga molekul ATP.
4
3 METODE
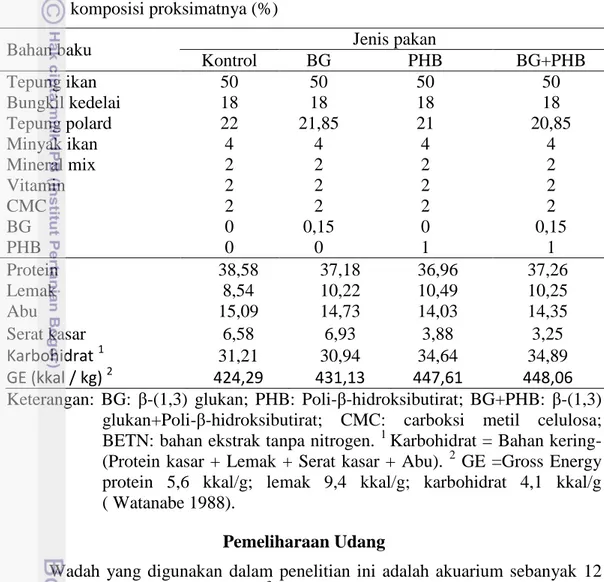
Pakan UjiPoli-β-hidroksibutirat (PHB) dan β-(1,3) glukan yang digunakan dalam penelitian ini adalah produk komersil yang dicampurkan bersama bahan pakan lain dalam pembuatan pakan. Formulasi pakan dan komposisi proksimat yang digunakan dalam penelitian ini disajikan pada Tabel 1.
Tabel 1 Formulasi bahan pakan yang digunakan dalam pembuatan pakan uji dan komposisi proksimatnya (%)
Bahan baku Jenis pakan
Kontrol BG PHB BG+PHB Tepung ikan 50 50 50 50 Bungkil kedelai 18 18 18 18 Tepung polard 22 21,85 21 20,85 Minyak ikan 4 4 4 4 Mineral mix 2 2 2 2 Vitamin 2 2 2 2 CMC 2 2 2 2 BG 0 0,15 0 0,15 PHB 0 0 1 1 Protein 38,58 37,18 36,96 37,26 Lemak 8,54 10,22 10,49 10,25 Abu 15,09 14,73 14,03 14,35 Serat kasar 6,58 6,93 3,88 3,25 Karbohidrat 1 31,21 30,94 34,64 34,89 GE (kkal / kg) 2 424,29 431,13 447,61 448,06
Keterangan: BG: β-(1,3) glukan; PHB: Poli-β-hidroksibutirat; BG+PHB: β-(1,3) glukan+Poli-β-hidroksibutirat; CMC: carboksi metil celulosa; BETN: bahan ekstrak tanpa nitrogen. 1 Karbohidrat = Bahan kering-(Protein kasar + Lemak + Serat kasar + Abu). 2 GE =Gross Energy protein 5,6 kkal/g; lemak 9,4 kkal/g; karbohidrat 4,1 kkal/g ( Watanabe 1988).
Pemeliharaan Udang
Wadah yang digunakan dalam penelitian ini adalah akuarium sebanyak 12 buah dengan ukuran 90×40×35 cm3. Akuarium diisi air laut sebanyak 90 L dan didesinfeksi dengan larutan klorin 30 mg/L serta diaerasi kuat selama 24 jam. Kemudian ditambahkan sodium tiosulfat 15 mg/L untuk menetralkan kandungan klorin dan diaerasi kuat minimal empat jam.
Udang yang digunakan adalah udang vaname bobot rata-rata 2,06 ± 0,03 g. Udang vaname diperoleh dari Balai Pengembangan Budidaya Air Payau Situbondo, Jawa Timur. Udang dengan kepadatan awal 222 ekor/m2 dipelihara selama 42 hari. Pemeliharaan menggunakan sistem resirkulasi dengan debit air 2,5 L/menit dan dilakukan pergantian air setiap hari sebanyak 20-30%. Kualitas air yang diukur selama penelitian ini seperti temperatur, pH, oksigen terlarut dan salinitas yang disajikan pada Tabel 2. Pakan yang diberikan berdasarkan feed
5 rate yang dikalikan biomassa (VanWyk 1999) dengan frekuensi pemberian pakan 4 kali sehari yaitu pukul 06.00, 10.00, 14.00 dan 20.00.
Tabel 2 Kondisi air pemeliharaan udang vaname Litopenaeus vannamei selama 42 hari
Perlakuan Kualitas air
Temperatur (°C) Oksigen terlarut (ppm) pH Salinitas (ppt)
Kontrol 28,4–28,6 6,2–6,7 8 33
BG 28,3–28,5 6,2–6,7 8 33
PHB 28,5–28,6 6,2–6,7 8 32
BG+PHB 28,4–28,5 6,4–6,6 8 33
Keterangan: BG: β-(1,3) glukan; PHB: Poli-β-hidroksibutirat; BG+PHB: β-(1,3) glukan+Poli-β-hidroksibutirat.
Parameter uji
Setelah masa pemeliharaan selama 42 hari, udang dipanen untuk dilakukan pengukuran kinerja pertumbuhan dan respons imun non spesifik. Kinerja pertumbuhan yang diukur meliputi specific growth rate (SGR), feed convertion ratio (FCR), retensi protein dan sintasan. Pengukuran respons imun non spesifik meliputi respiratory burst (RB) dan total haemocyte count (THC).
Specific Growth Rate (SGR)
Specific growth rate (SGR) dihitung menggunakan rumus (Huisman 1987).
(√ ) % Keterangan :
SGR : Specific growth rate (%)
Wo : Bobot tubuh rata-rata pada awal pertumbuhan (g) Wt : Bobot tubuh rata-rata pada akhir pertumbuhan (g) t : Waktu pemeliharaan (Hari)
Feed Convertion Ratio (FCR)
Feed Convertion Ratio (FCR) selama pemeliharaan dihitung menggunakan rumus (Zonneveld et al. 1991).
FCR = F
Bt + Bm - Bo Keterangan :
FCR : Feed convertion ratio F : Jumlah pakan (g)
Bt : Biomassa udang pada saat akhir perlakuan (g) Bm : Biomassa udang yang mati saat perlakuan (g) Bo : Biomassa udang pada saat awal perlakuan (g)
6
Retensi Protein (RP)
Retensi protein (RP) dihitung berdasarkan rumus (Takeuchi 1988) RP = F - I x 100%
P Keterangan:
RP : Retensi protein (%)
F : Jumlah protein tubuh pada akhir penelitian (g) I : Jumlah protein tubuh pada awal penelitian (g) P : Jumlah protein yang dikonsumsi (g)
Sintasan
Sintasan adalah persentase antara udang vaname yang hidup pada akhir pemeliharaan dengan jumlah yang ditebar pada awal penelitian dihitung dengan rumus (Efendi 1997). % 100 x No Nt SR Keterangan: SR : Persentase sintasan (%)
Nt : Jumlah udang vaname pada akhir penelitian No : Jumlah udang vaname pada awal penelitian Total Haemocyte Count (THC)
Tiga ekor udang diambil secara acak dari masing-masing ulangan untuk pengambilan sampel hemolim. Pengambilan hemolim dilakukan dengan menggunakan syringe berukuran 1 mL yang telah diisi dengan sepertiga antikoagulan. Sebanyak 0,6 mL hemolim diambil menggunakan jarum suntik 1 mL yang telah berisi 0,2 mL antikoagulan. Campuran hemolim-antikoagulan tersebut kemudian diteteskan pada haemocytometer. Selanjutnya THC dihitung di bawah mikroskop dengan perbesaran 40x (Blaxhall dan Daishley 1973).
Respiratory Burst (RB)
Respiratory burst dari hemosit diukur berdasarkan reduksi NBT (nitroblue tetrazolium) sebagai ukuran superoxide anion (O2-). Sebanyak 50 μL campuran hemolim-antikoagulan diinkubasi selama 30 menit pada suhu ruang. Kemudian disentrifuse 3.000 rpm selama 20 menit dan supernatan dibuang. Ditambahkan 100 μL NBT dalam larutan HBSS (hank's buffered salt solution konsentrasi 0,3 % didiamkan 2 jam pada suhu ruang. Kemudian disentrifuse 3.000 rpm 10 menit, supernatan dibuang dan ditambahkan 100 μL metanol absolut disentrifuse 3.000 rpm selama 10 menit. Endapan yang terbentuk kemudian dibilas sebanyak 2 kali dengan metanol 70 %. Selanjutnya 120 μL KOH (2M) dan 140 μL DMSO (dimethylsulfoxide) ditambahkan untuk melarutkan endapan. Endapan yang telah larut dimasukkan ke microplate diukur densitas optikal (OD) menggunakan microplate reader panjang gelombang 630 nm. Respiratory burst dinyatakan sebagai reduksi NBT per 10 μL hemolim (Cheng et al. 2004)
7 Analisa kimia
Analisis proksimat dilakukan pada pakan, udang vaname awal pemeliharaan dan udang vaname diakhir pemeliharaan. Analisis proksimat pakan perlakuan meliputi kadar protein kasar dilakukan dengan metode Kjeldahl, kadar lemak kering dengan metode Soxhlet, kadar lemak basah dengan metode Folch, kadar abu dengan pemanasan sampel pada suhu 600 °C, serat kasar menggunakan metode pelarutan sampel dengan asam, basa kuat dan pemanasan serta kadar air dengan metode pemanasan dalam oven bersuhu 105-110 °C (Takeuchi 1988).
Analisis Data
Data kinerja pertumbuhan dan respons imun non spesifik udang diolah menggunakan SPSS versi 17 dengan selang kepercayaan 95% dan untuk melihat perbedaan antar perlakuan digunakan uji lanjut Duncan.
8
4
HASIL DAN PEMBAHASAN
Hasil
Setelah dipelihara selama 42 hari, udang mengalami pertumbuhan yang signifikan. Hal ini bisa dilihat dari terjadinya peningkatan bobot tubuh udang yang meningkat 3,03-4,78 kali (Tabel 3). Udang yang diberi pakan BG 0,15% memiliki laju pertumbuhan yang paling tinggi dibandingkan dengan perlakuan lain. Namun, udang yang diberi PHB 1%, secara signifikan memiliki laju pertumbuhan yang lebih rendah. Fenomena laju pertumbuhan antar perlakuan ini sejalan dengan nilai peningkatan bobot udang, konversi pakan, dan sintasan. Di sisi lain, retensi protein udang yang diberi penambahan BG, secara signifikan berbeda nyata dengan perlakuan lain tetapi tidak berbeda nyata dengan kontrol.
Tabel 3 Kinerja pertumbuhan udang vaname Litopenaeus vannamei setelah pemeliharaan selama 42 hari
Parameter Perlakuan Kontrol BG PHB BG+PHB Bobot awal (g) 2,07±0,03 2,09±0,03 2,03±0,13 2,04±0,12 Bobot akhir (g) 8,05±0,20 9,97±0,42 7,47±0,30 8,49±0,79 SGR (%) 1,88±0,03ab 2,14±0,05c 1,80±0,06a 1,95±0,09b FCR 2,41±0,09b 2,16±0,04a 2,59±0,18b 2,53±0,05b Retensi protein (%) 14,51±0,94ab 16,08±3,09b 11,78±2,09a 12,06±0,21a Sintasan (%) 85,00±0,00b 96,67±5,77c 73,33±2,89a 75,00±5,00a Keterangan: SGR: specific growth rate; FCR: feed conversion ratio; BG: β-(1,3)
glukan; PHB: poli-β-hidroksibutirat; BG+PHB: β-(1,3) glukan+poli-β-hidroksibutirat. Angka yang diikuti huruf superskrip yang sama dalam satu baris menunjukkan tidak berbeda nyata (p >0,05).
Respons imun non spesifik yang terdiri dari THC dan RB disajikan pada Tabel 4. Pemberian BG, maupun kombinasi BG dan PHB pada udang melalui pakan mampu meningkatkan respons imun non spesifik udang tersebut. Respons imun non spesifik udang yang diberi pakan dengan penambahan BG secara signifikan berbeda nyata dengan perlakuan yang lain.
Tabel 4 Parameter imun non spesifik udang vaname Litopenaeus vannamei setelah masa pemeliharaan selama 42 hari
Parameter Perlakuan
Kontrol BG PHB BG+PHB
THC (x 107 sel/ml) 6,35±2,75b 10,87±2,24c 1,70±0,14a 7,16±2,78b RB (Abs/10μl) 0,04±0,00a 0,12±0,02b 0,05±0,00a 0,07±0,02a Keterangan: THC: total haemocyte count; RB: respiratory burst; BG: β-(1,3)
glukan; PHB: Poli-β-hidroksibutirat; BG+PHB: β-(1,3) glukan+Poli-β-hidroksibutirat. Angka yang diikuti huruf superskrip yang sama dalam satu baris menunjukkan tidak berbeda nyata (p>0,05).
9
Pembahasan
Hasil penelitian menunjukkan bahwa penambahan β-(1,3) glukan (BG) di dalam pakan menghasilkan pertumbuhan udang yang paling tinggi. β-(1,3) glukan yang diberikan kepada udang dapat diurai dalam proses pencernaan secara enzimatis untuk menghasilkan energi (Manoppo 2011). Energi tersebut digunakan untuk meredam stress pada pemeliharaan udang secara intensif, sehingga udang tidak mudah terserang penyakit dan dapat tumbuh dengan optimal. Hal ini sesuai dengan pernyataan (Lopez et al. 2003) yang mengatakan bahwa penambahan BG 2 g/kg dalam pakan yang diberikan pada udang vaname selama 48 hari dapat meningkatkan laju pertumbuhan spesifik sampai 14% lebih tinggi dibandingkan kontrol. Penelitian (Sang dan Fotedar 2010) juga menunjukan laju pertumbuhan Cherax tenuimanus yang diberi BG 0,1-0,2 mg/kg pakan selama 84 hari lebih tinggi dibandingkan dengan yang tidak diberikan BG. Namun demikian, udang yang diberi pakan PHB memiliki pertumbuhan yang rendah dibandingkan perlakuan yang lain. Pertumbuhan yang rendah ini disebabkan oleh meningkatnya produksi asam lemak dalam usus, sehingga terjadi peneurunan nilai pH dalam usus udang vaname. Hal ini bertolak belakang dengan hasil penelitian (De Schryver et al. 2010) pada ikan European sea bass Dicentrarchus labrax PHB yang terdegradasi oleh enzim depolimerase menjadi asam lemak butirat menyebabkan meningkatnya produksi asam lemak rantai pendek dalam usus, yang dicirikan dengan terjadinya penurunan pH dari 7,7 menjadi 7,2. Potensial hidrogen (pH) usus yang turun menyebabkan udang kehilangan banyak energi, sehingga pertumbuhanya rendah hal ini juga berakibat terhadap rendahnya sintasan udang vaname. Udang vaname juga diduga tidak memeiliki enzim carnitine palmitoyltransferase, sehingga asam lemak tersebut tidak bisa masuk ke mitokondria untuk menghasilkan energi. Energi yang seharusnya untuk meredam stress tidak mencukupi, sehingga udang mudah terserang penyakit, pertumbuhan rendah, retensi protein rendah, fagositosis tidak berjalan sempurna ditandai dengan nilai THC dan RB yang rendah hal ini juga menyebabkan sintasan rendah.
Udang vaname yang diberi pakan dengan penambahan BG memiliki nilai FCR yang berbeda nyata dengan perlakuan lain maupun kontrol. Hal ini diduga karena adanya proses penguraian BG secara enzimatis yang menghasilkan energi menyebabkan kebutuhan energi udang lebih cepat terpenuhi. Faktor ini yang menyebabkan udang vaname yang diberi pakan dengan BG lebih cepat terpenuhi kebutuhan energinya, sehingga lebih cepat berhenti makan yang menyebabkan jumlah pakan yang dibutuhkan lebih sedikit. Semakin rendah FCR maka semakin baik dan efektif karena semakin sedikit pakan yang diperlukan untuk menghasilkan 1 kg daging (Zonneveld et al. 1991). Namun demikian, udang yang diberi PHB memiliki nilai FCR yang tinggi karena udang memerlukan banyak energi untuk mempertahankan pH usus, sehingga udang banyak memerlukan pakan untuk memenuhi energi tersebut.
Nilai retensi protein udang yang diberi BG pada pakan memiliki nilai yang tertinggi. Tingginya retensi protein disebabkan oleh adanya tambahan energi dari penguraian BG, sehingga diduga tidak ada protein yang dipecah menjadi energi, hal ini yang mengakibatkan retensi proteinya tinggi. Udang vaname yang diberi
10
PHB memiliki nilai retensi protein yang terendah. Rendahnya nilai retensi protein disebabkan oleh banyaknya protein yang dipecah menjadi energi karena energi dari lemak tidak bisa digunakan untuk mempertahankan pH usus, sehingga protein yang diretensi menjadi sedikit. Namun demikian, udang yang diberi BG juga tidak berbeda nyata dengan kontrol. Hal ini diduga karena ada protein yang dipecah menjadi energi untuk meredam sterss.
Nilai total haemocyte count (THC) udang vaname yang diberi BG memiliki nilai yang tertinggi dan berbeda nyata dengan perlakuan lain maupun kontrol. Tingginya nilai THC disebabkan oleh terurainya β-(1,3) glukan yang menghasilkan energi untuk mendukung berjalanya proses fagositosis berjalan sempurna, sehingga produksi THC udang vaname menjadi tinggi. Selain itu, β-(1,3) glukan juga bekerja dengan cara mengikat molekul reseptor yang terdapat pada permukaan sel fagosit, sehingga sel fagosit menjadi lebih aktif dalam melakukan fagositosis terhadap partikel asing dan merangsang pembentukan sel-sel hemosit yang baru (Rodriguez dan Le Moullac 2000). Penelitian sebelumnya yang dilaporkan oleh Sahoo et al. (2008) udang galah yang diberi pakan dengan tambahan BG dengan kadar 1,5 g/kg memiliki nilai THC yang lebih tinggi dibandingkan dengan perlakuan lain. Sahoo et al. (2008) menjelaskan tingginya nilai THC pada pada udang galah yang diberi BG diakibatkan oleh tingginya mobilisasi sel hemosit dalam tubuh udang, sehingga dapat meningkatkan imunitas dan pengenalan mengenai benda asing yang masuk dalam tubuh udang.
Udang vaname yang diberi PHB memiliki nilai THC yang rendah, hal ini disebabkan oleh banyak energi yang hilang untuk mempertahankan pH usus, sehingga proses fagositosis tidak berjalan dengan baik yang berakibat pada rendahnya nilai THC. Nilai THC yang rendah menyebabkan udang vaname menjadi rentan terhadap patogen (Le Moullac et al. 1998). Nilai THC yang rendah juga dapat menyebabkan terjadi infeksi akut yang dapat berakibat pada kematian (Rodriguez dan Le Moullac 2000). Nilai THC sejalan dengan nilai respiratory burst (RB), udang vaname yang diberi BG memiliki nilai tertinggi dan berbeda nyata antar perlakuan maupun kontrol. Udang vaname yang diberi BG memiliki nilai RB tertinggi karena BG dapat mengaktifkan sel fagosit dan BG juga dapat diurai menjadi energi untuk mendukung proses fagositosis, hal ini yang menyebabkan nilai RB menjadi lebih tinggi. Respiratory burst berkaitan dengan mekanisme fagositosis, sehingga semakin tinggi nilai RB maka sistem imun udang menjadi semakin baik (Rodriguez dan Le Moullac 2000). Namun demikian, udang yang diberi PHB memiliki nilai RB yang rendah karena udang banyak kehilangan energi untuk mempertahan pH usus dan tidak memiliki enzim carnitine palmitoyltransferase yang dapat merubah asam lemak butirat menjadi energi. Hal ini yang menyebabkan udang vaname tidak memiliki energi yang cukup untuk fagositosis, sehingga fagositosis tidak berjalan sempurna yang berakibat pada rendahnya nilai RB.
Sintasan udang vaname yang tertinggi terjadi pada perlakuan dengan penambahan β-(1,3) glukan dalam pakan. β-(1,3) glukan yang diurai secara enzimatis menghasilkan tambahan energi untuk meredam stress dan meningkatkan proses fagositosis, sehingga udang vaname tidak mudah terserang penyakit yang berakibat pada sintasan tinggi. Hal ini sesuai dengan pernyataan (Yin et al. 2006) yang menyatakan bahwa β-(1,3) glukan dapat meningkatkan
11 respons imun udang dengan cara meningkatkan aktivitas sel-sel fagosit untuk menjalankan proses fagositosis. Meningkatnya respons imun udang tersebut dapat mempengaruhi nilai sintasan menjadi lebih tinggi, karena resistensi udang terhadap patogen juga meningkat (Cook et al. 2003). Hasil penelitian (Chang et al. 2003b) memperlihatkan bahwa pemberian oral BG selama 20 hari secara efektif meningkatkan sistem imun udang windu, sehingga sintasanya meningkat. Chang et al. (2003a) juga melaporkan bahwa suplementasi BG 2 g/kg pakan selama 40 hari meningkatkan sintasan udang windu.
Namun demikian, udang vaname yang diberi PHB dan kombinasi BG dengan PHB memiliki sintasan yang rendah. Hal ini diduga karena PHB yang terdegredasi menjadi asam lemak butirat dapat menurunkan pH usus, sehingga banyak energi yang digunakan untuk mempertahan pH tersebut. Energi untuk proses fagositosis tidak mencukupi, sehingga udang mudah terserang penyakit yang berakibat pada rendah sintasan. Sistem pemeliharaan udang vaname dengan kepadatan tinggi atau intensif juga dapat menyebabkan stress sedangkan energi untuk meredam stress tersebut tidak cukup, sehingga mudah terserang penyakit dan dapat menyebabkan kematian. Poli-β-hidroksibutirat dan kombinasi BG dengan PHB rendah juga diduga karena udang vaname tidak memiliki enzim carnitine palmitoyltransferase yang dapat mengubah asam lemak butirat menjadi energi, sehingga energi untuk meredam stress dan fagositosis tidak mencukupi yang berakibat pada sintasan yang rendah. Selain itu, kombinasi BG dengan PHB sintasanya rendah diduga karena tidak hanya BG dan PHB yang berperan penting dalam biofok tetapi ada nutrien lain yang berperan dalam bioflok tersebut.
5
KESIMPULAN DAN SARAN
Kesimpulan
Pemberian β-(1,3) glukan sebagai feed additive melalui pakan memiliki kemampuan yang terbaik didalam meningkatkan kinerja pertumbuhan dan respons imun non spesifik. Namun demikian, feed additive yang berupa poli-β-hidroksibutirat memiliki kinerja pertumbuhan dan respons imun non spesifik yang rendah. Udang vaname yang diberi pakan kombinasi antara β-(1,3) glukan dan poli-β-hidroksibutirat juga belum bisa meningkatkan kinerja pertumbuhan dan respons imun non spesifik.
Saran
Perlu dilakukan penelitian lebih lanjut mengenai dosis poli-β-hidroksibutirat yang tepat dan faktor lain yang menyebabkan teknologi bioflok dapat meningkatkan pertumbuhan dan sintasan udang vaname.
12
DAFTAR PUSTAKA
Anderson, AJ, Dawes, EA. 1990. Occurence, Metabolism, Metabolic Role and Industrial Uses of Bacterial Polyhydroxyalkanoates. Microbiological Reviews. 54(4): 450-472.
Avnimelech Y. 1999. Carbon/nitrogen ratio as a controlelement in aquaculture systems. Aquaculture 176: 227-235.
Ber L. 1997. Yeast derived beta-1,3-D-Glucan: an adjuvant concept. American Journal of Natural Medicine 4 (3). www.anma.com/mon43.html.
Blaxhall PC, Daysley KW. 1973. Routine haemotological methods for use with fish blood. Journal Fish Biology 5:577-581.
Chang CF, Chen HY, Su MS, Liao IC. 2003a. Immunomodulation by dietary β-1,3-glucan in the brooders of the black tiger shrimp Penaeus monodon. Fish Shellfish Immunology 10: 505-514.
Cheng W, Liu CH, Yeh ST, JC. Chen. 2004. The immune stimulatory effect of sodium alginate on the white shrimp Litopenaeus vannamei and its resistance against Vibrio alginolyticus. Fish and Shellfish Immunology 17: 41-51.
Cook MT, Hayball PJ, Hutchinson W, Nowak BF, Hayball JD. 2003. Administration of a commercial immune-stimulan preparation, EcoActiva as a feed supplement enhances macrophage respira-tory burst and the growth rate of snaper Pagurus auratus Sparidae (Bloch and Schneider) in winter. Fish and Shellfish Immunology 14: 333-345.
Defoirdt T, Halet D, Vervaeren H, Boon N, Van de WT, Sorgeloos P, Bossier P, Verstraete W. 2007. The bacterial storage compound of poly- β-hydrobutyrate protects Artemia fransiseana from pathogenic Vibrio campbellii. Environmental Microbiology 9: 445-452.
Defoirdt T, Boon N, Sorgeloos P, Verstraete W, Bossier P. 2009. Short-chain fatty acids and poly-β-hydroxyalkanoates: (new) biocontrol agents for a sustainable animal production (review). Biotechnology Advances 27: 680-685.
De Schryver and Verstraete, Willy. 2009. Nitrogen removal from aquaculture pond water by heterotrophic nitrogen assimilation in lab-scale sequenching batch reactor. Bioresource Technology 100 (2009) 1162-1167.
De Schryver P, Sinha AK, Kunwarr PS, Baruah K, Verstraete W, Boon N, De Boeck G, Bossier P. 2010. Poly-beta-hydroxybutyrate (PHB) increases growth performance and intestinal bacterial range-wighted richness in juvenile European sea bass, Dicentrarchus labrax. Appl. Microbio. Biotecnol 86: 1535-1541.
Ha C, K. Lim, Y. Kim, S. Lim, C. Kim, and H. Chang. 2002. Analysis of alkali-soluble glucan produced by Saccharomyces cerevisiae wild-type and mutants. Applied Microbiology and Biotechnology 58 (3): 370-377.
Horowitz S, Horowitz A. 2002. Microbial intervention in aquaculture. In: Lee, C.-S., O’Bryen, P. (Eds.), Proceedings of Microbial Approaches to Aquatic Nutrition within Environmentally Sound Aquaculture Production Systems. The World AquacultureSociety, Baton Rouge, Louisiana, USA pp. 119– 131. Huisman EA.1987. Principles of fish production. Netherland:Department of Fish
Culture and Fisheries, Wageningen Agriculture University.Waganingen.Netherland.170p.
13 Hunter KW, Gault RA, Berner MD. 2002. Preparation of microparticulate
β-glucan from Saccharomyces cerevisiae for use in immune potentiation. Lett Appl Microbiol 35: 267-271.
Johansson M, Keyser P, Sritunyalucksana K, Soderhall K. 2000. Crustacean haemocytes and haemotopoiesis. Aquaculture 191: 45-52.
Jorgensen J , Robertsen B. 1995. Yeast β-glucan stimulates respiratory burst activity of Atlantic salmon (Salmo salar L.) macrophages. Developmental and Comparative Immunology 19: 43–57.
Kulickle WM, AL. Lettau, H. Thielking. 1996. Correlation between immunological activity, molar mass, and molar structure of different (1,3)-β-D-glucans. Carbohydrate Research 297: 135-143.
Lee JN, Lee DY, In-Hye J, Gi-Eun K, Kim HN. 2001. Purification of soluble β Glucan with immuno-enhancing activity from the cell wall of yeast. Bioscience Biotechnology and Biochemistry. 65 (4): 837-841.
Le Moullac G, Soyez C, Saulnier D, Ansquer D, Avarre JC, Levy P. 1998. Effect of hypoxic stress on the immune response and the resistance to vibriosis of the shrimp Penaeus stylirostris. Fish Shellfish and Immunology 8: 621-629.
Lopez N, Cuzon G, Gaxiola G, Taboada G, Valenzuela M, Pascual C, Sanches A, Rosas C. 2003. Physiological, nutritional, and immunological role of dietary β-glukan and ascorbic acid 2-monophospate in Litopenaeus vannamei juveniles. Aquaculture 224: 223-243.
Manoppo H. 2011. Peran nukleotida sebagai imunostimulan terhadap respons imun nonspesifik dan resistensi udang vaname (Litopenaues vannamei) [Disertasi]. Bogor(ID): Institut Pertanian Bogor.
Ramu K, Zakaria. 2000. Defence mechanism in crustacean. Infofish International 5 : 30 – 32.
Rodriguez L, Le Moullac G. 2000. State of the art of immunological tools and health control of penaeid shrimp. Aquaculture 191: 109-119.
Sahoo PK, Das A, Mohanty S, Mohanty BK, Pilai BR, Mohanty J. 2008. Dietary β-1,3 glucan improve the immunity and disease resistance of freshwater prawn Macrobrachium rosenbergii challenged with Aeromonas hydrophyla. Aquaculture Research 39: 1574-1578.
Sakai M. 1999. Current research status of fish immunostimulants. Aquaculture 172: 63-92.
Sang H, Fotedar R. 2010. Effects of dietary β-1,3-glukan on the growth, survival, physiological, and immune response of marron, Cherax tenuimanus (Smith 1912). Fish and Shellfish Immunology 28: 957-960.
Siwicki AK, Morand M, Terech-Majewska E, Niemczuk W, Kazun K, Glombski E.1998. Influence of immunostimulants on the effectiveness of vaccine in fish: in vitro and in vivo study. J. Appl. Ichthyology 14: 225-227.
Takeuchi T. 1988. Laboratory work-chemical evaluation of dietary nutrients, In: Watanabe T (ed). Fish Nutrition and Mariculture. Japan: Kanagawa international fisheries training centre. Japan International Cooperation Agency (JICA). pp 179-233.
VanWyk P, Scarpa J. 1999.Water Quality and Management. In: Van Wyk P. et al. (Eds.), Farming Marine Shrimp in Recirculating Freshwater Systems. Florida Department of Agriculture and Consumer Services, Tallahassee pp. 128–138. Vetvicka V, Terayama K, Mandeville R, Brousseau P, Kournikakis B, Ostroff G.
14
2002. Orally-administered yeast beta-1,3-glukan prophylactically protects against anthrax infection and cancer in mice. The Journal the American Nutraceutical Association 5: 1-5.
Watanabe T. 1988. Fish nutrition and mariculture. Department of aquatic Bioscience. Tokyo University of Fisheries. JICA. 233 p.
Yin G, Jeney G, Racs T, Xu P, Jun X, Jeney Z. 2006. Effect of two Chinese herbs (Astragalus radix and Scutellaria radix) on nonspecific immune system of tilapia, Oreochromis niloticus. Aquaculture 253: 39-47.
Zonneveld N, Huisman EA, Boon JH. 1991. Prinsip-Prinsip Budidaya Ikan. Jakarta(ID): PT Gramedia Pustaka Utama.
15 Lampiran 1 Hasil analisis statistik laju pertumbuhan spesifik (SGR) udang vaname Litopenaeus vannamei yang diberi feed additive berbeda (β-(1,3) glukan 0% dan PHB 0%; β-(β-(1,3) glukan 0,15%; PHB 1% dan kombinasi β-(1,3) glukan 0,15% dan PHB 1%) selama 42 hari. Huruf yang sama menunjukkan tidak ada perbedaan nyata (P>0,05).
Descriptives SGR N Mean Std. Deviation Std. Error 95% Confidence Interval for Mean Minimum Maximum Lower Bound Upper Bound 1.00 3 1.8833 .02517 .01453 1.8208 1.9458 1.86 1.91 2.00 3 2.1367 .04726 .02728 2.0193 2.2541 2.10 2.19 3.00 3 1.7967 .05508 .03180 1.6599 1.9335 1.74 1.85 4.00 3 1.9500 .09539 .05508 1.7130 2.1870 1.89 2.06 Total 12 1.9417 .14064 .04060 1.8523 2.0310 1.74 2.19 ANOVA SGR Sum of
Squares df Mean Square F Sig.
Between Groups .188 3 .063 16.673 .001 Within Groups .030 8 .004 Total .218 11 SGR Duncana Perlakuan N
Subset for alpha = 0.05
1 2 3 3.00 3 1.7967 1.00 3 1.8833 1.8833 4.00 3 1.9500 2.00 3 2.1367 Sig. .121 .219 1.000
Means for groups in homogeneous subsets are displayed. a. Uses Harmonic Mean Sample Size = 3.000.
16
Lampiran 2 Hasil analisis statistik rasio konversi pakan (FCR) udang vaname Litopenaeus vannamei yang diberi feed additive berbeda (β-(1,3) glukan 0% dan PHB 0%; β-(1,3) glukan 0,15%; PHB 1% dan kombinasi β-(1,3) glukan 0,15% dan PHB 1%) selama 42 hari. Huruf yang sama menunjukkan tidak ada perbedaan nyata (P>0,05).
Descriptives FCR N Mean Std. Deviation Std. Error 95% Confidence Interval for Mean
Minimum Maximum Lower Bound Upper Bound 1.00 3 2.4100 .09165 .05292 2.1823 2.6377 2.31 2.49 2.00 3 2.1667 .03512 .02028 2.0794 2.2539 2.13 2.20 3.00 3 2.5900 .18083 .10440 2.1408 3.0392 2.40 2.76 4.00 3 2.5333 .05132 .02963 2.4059 2.6608 2.49 2.59 Total 12 2.4250 .19252 .05558 2.3027 2.5473 2.13 2.76 ANOVA FCR Sum of
Squares df Mean Square F Sig.
Between Groups .318 3 .106 9.422 .005 Within Groups .090 8 .011 Total .408 11 FCR Duncana Perlakuan N
Subset for alpha = 0.05
1 2 2.00 3 2.1667 1.00 3 2.4100 4.00 3 2.5333 3.00 3 2.5900 Sig. 1.000 .081
Means for groups in homogeneous subsets are displayed. a. Uses Harmonic Mean Sample Size = 3.000.
17 Lampiran 3 Hasil analisis statistik sintasan udang vaname Litopenaeus vannamei yang diberi feed additive berbeda (β-(1,3) glukan 0% dan PHB 0%; β-(1,3) glukan 0,15%; PHB 1% dan kombinasi β-(1,3) glukan 0,15% dan PHB 1%) selama 42 hari. Huruf yang sama menunjukkan tidak ada perbedaan nyata (P>0,05).
Descriptives Sintasan N Mean Std. Deviation Std. Error 95% Confidence Interval for Mean
Minimum Maximum Lower Bound Upper Bound 1.00 3 85.0000 .00000 .00000 85.0000 85.0000 85.00 85.00 2.00 3 96.6667 2.88675 1.66667 89.4956 103.8378 95.00 100.00 3.00 3 73.3333 2.88675 1.66667 66.1622 80.5044 70.00 75.00 4.00 3 75.0000 5.00000 2.88675 62.5793 87.4207 70.00 80.00 Total 12 82.5000 10.11300 2.91937 76.0745 88.9255 70.00 100.00 ANOVA Sintasan Sum of
Squares df Mean Square F Sig.
Between Groups 1041.667 3 347.222 33.333 .000 Within Groups 83.333 8 10.417 Total 1125.000 11 Sintasan Duncana Perlakuan N
Subset for alpha = 0.05
1 2 3 3.00 3 73.3333 4.00 3 75.0000 1.00 3 85.0000 2.00 3 96.6667 Sig. .545 1.000 1.000
Means for groups in homogeneous subsets are displayed. a. Uses Harmonic Mean Sample Size = 3.000.
18
Lampiran 4 Hasil analisis retensi protein udang vaname Litopenaeus vannamei yang diberi feed additive berbeda (β-(1,3) glukan 0% dan PHB 0%; β-(1,3) glukan 0,15%; PHB 1% dan kombinasi β-(1,3) glukan 0,15% dan PHB 1%) selama 42 hari. Huruf yang sama menunjukkan tidak ada perbedaan nyata (P>0,05).
Descriptives R.Protein N Mean Std. Deviation Std. Error 95% Confidence Interval for Mean
Minimum Maximum Lower Bound Upper Bound 1.00 3 14.5100 .94016 .54280 12.1745 16.8455 13.56 15.44 2.00 3 16.0833 3.08923 1.78357 8.4093 23.7574 14.25 19.65 3.00 3 11.7833 2.08433 1.20339 6.6056 16.9611 10.56 14.19 4.00 3 12.0600 .21000 .12124 11.5383 12.5817 11.85 12.27 Total 12 13.6092 2.47958 .71579 12.0337 15.1846 10.56 19.65 ANOVA R.Protein Sum of
Squares df Mean Square F Sig.
Between Groups 38.000 3 12.667 3.420 .073 Within Groups 29.632 8 3.704 Total 67.631 11 R.Protein Duncana Perlakuan N
Subset for alpha = 0.05
1 2 3.00 3 11.7833 4.00 3 12.0600 1.00 3 14.5100 14.5100 2.00 3 16.0833 Sig. .135 .346
Means for groups in homogeneous subsets are displayed. a. Uses Harmonic Mean Sample Size = 3.000.
19 Lampiran 5 Hasil analisis statistik total haemocyte count (THC) udang vaname
Litopenaeus vannamei yang diberi feed additive berbeda (β-(1,3) glukan 0% dan PHB 0%; β-(1,3) glukan 0,15%; PHB 1% dan kombinasi β-(1,3) glukan 0,15% dan PHB 1%) selama 42 hari. Huruf yang sama menunjukkan tidak ada perbedaan nyata (P>0,05).
Descriptives THC N Mean Std. Deviation Std. Error 95% Confidence Interval for Mean Minimum Maximum Lower Bound Upper Bound 1.00 3 6.4133 2.86151 1.65209 -.6950 13.5217 4.35 9.68 2.00 3 10.8700 2.23658 1.29129 5.3140 16.4260 9.48 13.45 3.00 3 1.6467 .17786 .10269 1.2048 2.0885 1.52 1.85 4.00 3 5.1200 1.54211 .89034 1.2892 8.9508 3.96 6.87 Total 12 6.0125 3.83808 1.10796 3.5739 8.4511 1.52 13.45 ANOVA THC Sum of
Squares df Mean Square F Sig.
Between Groups 130.839 3 43.613 11.183 .003 Within Groups 31.201 8 3.900 Total 162.040 11 THC Duncana Perlakuan N
Subset for alpha = 0.05
1 2 3 3.00 3 1.6467 4.00 3 5.1200 5.1200 1.00 3 6.4133 2.00 3 10.8700 Sig. .063 .446 1.000
Means for groups in homogeneous subsets are displayed. a. Uses Harmonic Mean Sample Size = 3.000.
20
Lampiran 6 Hasil analisis statistik respiratory burst (RB) udang vaname Litopenaeus vannamei yang diberi feed additive berbeda (β-(1,3) glukan 0% dan PHB 0%; β-(1,3) glukan 0,15%; PHB 1% dan kombinasi β-(1,3) glukan 0,15% dan PHB 1%) selama 42 hari. Huruf yang sama menunjukkan tidak ada perbedaan nyata (P>0,05).
Descriptives RB N Mean Std. Deviation Std. Error 95% Confidence Interval for Mean Minimum Maximum Lower Bound Upper Bound 1.00 3 .0400 .00000 .00000 .0400 .0400 .04 .04 2.00 3 .1200 .02000 .01155 .0703 .1697 .10 .14 3.00 3 .0600 .01000 .00577 .0352 .0848 .05 .07 4.00 3 .0667 .02082 .01202 .0150 .1184 .05 .09 Total 12 .0717 .03353 .00968 .0504 .0930 .04 .14 ANOVA RB Sum of
Squares df Mean Square F Sig.
Between Groups .010 3 .003 15.000 .001 Within Groups .002 8 .000 Total .012 11 RB Duncana Perlakuan N
Subset for alpha = 0.05
1 2 1.00 3 .0400 3.00 3 .0600 4.00 3 .0667 2.00 3 .1200 Sig. .074 1.000
Means for groups in homogeneous subsets are displayed. a. Uses Harmonic Mean Sample Size = 3.000.
21
RIWAYAT HIDUP
Penulis bernama Sarmin, dilahirkan di Cilacap, tanggal 07 Juni 1988 sebagai anak Keenam dari pasangan Bapak Rasmuki dan Ibu Dasem. Saat ini penulis bertempat tinggal Desa Sadabumi Kecamatan Majenang Kabupaten Cilacap. Penulis menyelesaikan pendidikan dasar di SD 03 Sadabumi (1995 - 2001), jenjang tingkat sekolah menengah pertama di SMP N 1 Majenang (2001 - 2004), jenjang pendidikan menengah atas di SMA N 1 Majenang (2004 - 2007), dan melanjutkan pendidikan pada perguruan tinggi Universitas Jenderal Soedirman Fakultas Sains dan Teknik, Jurusan Perikanan dan Kelautan, Program Studi Budidaya
Perairan tahun 2007 dan lulus pada tahun 2012. Pada tahun 2012, penulis
melanjutkan studinya dengan menempuh Program Magister pada program studi Ilmu Akuakultur, Sekolah Pascasarjana, IPB.