Regenerasi Beberapa Kultivar Kentang dan Transformasi Kentang dengan Gen RB melalui Agrobacterium tumefaciens
Teks penuh
Gambar
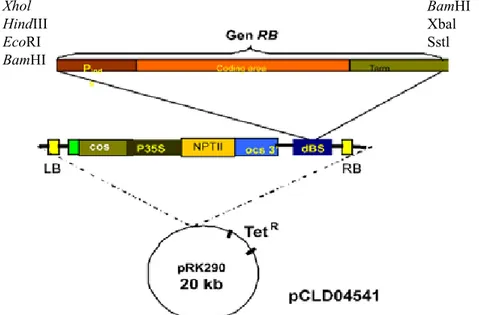

Dokumen terkait
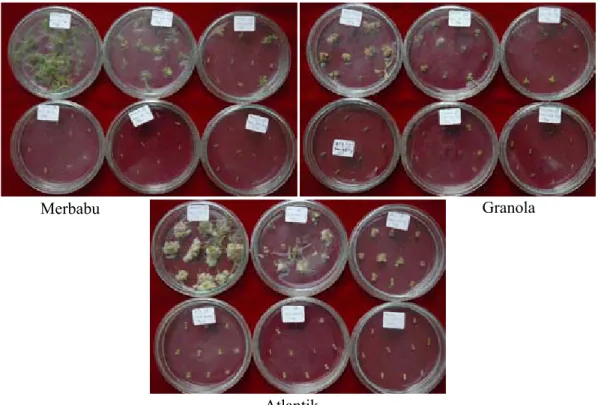
Kalus yang tumbuh dan bersih dari kontaminasi menggunakan metode ini mencapai 90%. Hasil kalus yang dapat tumbuh pada media seleksi dapat dilihat pada gambar 4. Kalus PA 183 dan
Kalus yang tumbuh dan bersih dari kontaminasi menggunakan metode ini mencapai 90%. Hasil kalus yang dapat tumbuh pada media seleksi dapat dilihat pada gambar 4. Kalus PA 183 dan
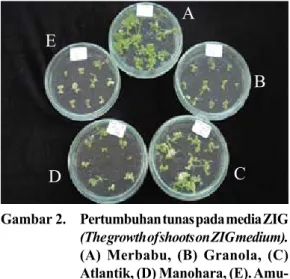
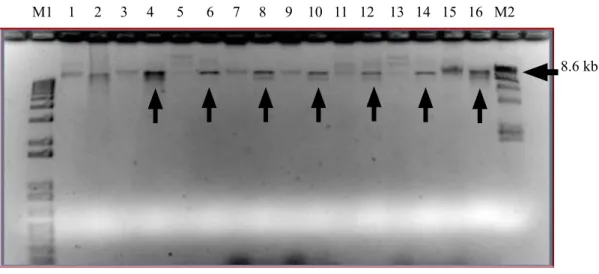
Tunas hasil regenerasi ini mampu tumbuh pada media seleksi yang mengandung 40 mg/L higromisin yang kemudian disebut dengan kentang transgenik putatif, sedangkan
Beberapa galur tanaman transgenik potensial tahan terhadap penggerek batang kuning, yaitu dengan tidak menunjukkan gejala sundep pada semua anakan yang diamati.. Pola
Berdasarkan perbandingan antara nilai AUDPC tanaman Granola yang terserang penyakit dan antara dengan galur transgenik yang menunjukkan kategori tahan dan sangat tahan
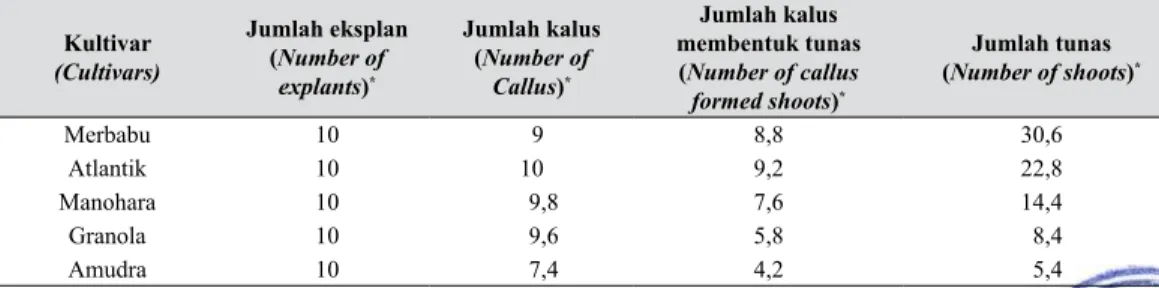
Dari sebanyak tiga kali transformasi genetik yang dilakukan pada genotipe Intan, telah diperoleh beberapa eksplan yang berhasil membentuk kalus dan tunas pada
