TRANSFORMASI GENETIK KENTANG (Solanum tuberosum
L.) KULTIVAR BARAKA DENGAN GEN PEMBUNGAAN
Hd3a
BUSTOMI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa skripsi berjudul Transformasi Genetik Kentang (Solanum tuberosum L.) Kultivar Baraka dengan Gen Pembungaan Hd3a
adalah benar karya saya bersama komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, September 2014
Bustomi
ABSTRAK
BUSTOMI. Transformasi Genetik Kentang (Solanum tuberosum L.) Kultivar Baraka dengan Gen Pembungaan Hd3a. Dibimbing oleh SUHARSONO dan UTUT WIDYASTUTI.
Pembungaan mempunyai peranan yang sangat penting dalam pemuliaan tanaman. Pembungaan merupakan transisi dari fase vegetatif ke fase generatif. Salah satu gen yang berhubungan dengan pembungaan adalah Hd3a. Penelitian ini bertujuan untuk melakukan transformasi genetik kentang (Solanum tuberosum L.) kultivar Baraka dengan gen pembungaan Hd3a dibawah kendali promoter kuat
prolC. Tanaman kentang diperbanyak secara in vitro di media MS2. Transformasi
genetik dilakukan melalui bantuan Agrobacterium tumefaciens dengan metode ko-kultivasi menggunakan potongan daun dan buku sebagai eksplan. Seleksi tanaman transgenik dilakukan di media MS yang mengandung 10-40 mg/L higromisin. Hasil penelitian menunjukkan bahwa rata-rata efisiensi transformasi genetik relatif rendah yaitu 16.36% dengan efisiensi regenerasi yang sangat tinggi yaitu 100%. Rendahnya efisiensi transformasi kemungkinan disebabkan oleh rusaknya eksplan selama proses transformasi. Efisiensi transformasi eksplan buku lebih tinggi daripada eksplan daun. Rata-rata tunas transgenik putatif yang dihasilkan tiap kalus adalah 2.5. Di media MS0, empat dari 24 tanaman transgenik putatif
menghasilkan umbi sedangkan tunas non-transgenik tidak menghasilkan umbi. Hal ini menunjukkan bahwa Hd3a menginduksi pembentukan umbi pada tanaman kentang kultivar Baraka transgenik putatif.
Kata kunci: Solanum tuberosum L. kultivar Baraka, transformasi genetik, gen pembungaan Hd3a
ABSTRACT
BUSTOMI. Genetic Transformation of Potato (Solanum tuberosum L.) Cultivars Baraka with Hd3a Flowering Gene. Supervised by SUHARSONO and UTUT gene under control of a strong promoter prolC. Potato plants were propagated in vitro on MS2 medium. Genetic transformation was carried out by co-cultivation
method by using Agrobacterium tumefaciens LBA4404. Leaf disc and internodes were used as explants for transformation. Selection of transgenic plants was performed in MS medium containing 10-40 mg/L hygromycin. The results showed that the efficiency of transformation was relatively low. The efficiency of shoot regeneration of transgenic calli was very high. The low efficiency of transformation was caused by damage of explant during transformation process. The efficiency of transformation of internode was higher than leaf explants. Putative transgenic shoot regeneration per callus was 2.5. In MS0 media, four of
twenty-four putative transgenic plants produced tuber whereas all non-transgenic plants did not. This result showed that Hd3a induced the formation of tuber of transgenic potato cv. Baraka.
Keywords: Solanum tuberosum L. cultivar Baraka, genetic transformation, Hd3a
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains
pada
Departemen Biologi
TRANSFORMASI GENETIK KENTANG (Solanum tuberosum
L.) KULTIVAR BARAKA DENGAN GEN PEMBUNGAAN
Hd3a
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Desember 2013 hingga Mei 2014 ini ialah transformasi genetik kentang (Solanum tuberosum L.) kultivar Baraka dengan gen pembungaan Hd3a .
Terima kasih penulis ucapkan kepada Bapak Prof Dr Ir Suharsono, DEA dan Ibu Dr Ir Utut Widyastuti, MSi selaku pembimbing atas segala bimbingan, dukungan, arahan, kesabaran waktu yang disediakan serta saran yang telah diberikan selama penelitian dan penulisan karya ilmiah ini. Penulis juga mengucapkan terima kasih kepada Proyek Penelitian Desentralisasi Baru IPB
yang berjudul “Rekayasa genetika tanaman dengan gen toleransi aluminium dan pembungaan” dengan kontrak no: 48/IT3.11/LT/2014 atas nama: Prof Dr Ir Suharsono, DEA yang telah membiayai penelitian ini. Terimakasih diucapkan juga kepada Dr Berry Juliandi, MSi selaku penguji skripsi atas semua saran, masukan, dan perbaikan yang telah diberikan. Terimakasih dan penghargaan sebesar-besarnya penulis sampaikan kepada seluruh staf di laboratorium Biologi Molekular dan Selular Tanaman (BMST) dan BIORIN, khususnya pada Mbak Nia, Mbak Pepy, Mbak Ara, Pak Asep, Pak Mulya, Pak Iri, dan Pak Yanto. Terimakasih juga ditujukan kepada Bapak Diky, SP (BA Farm) yang telah membantu dalam proses aklimatisasi. Ucapan terimakasih juga penulis sampaikan kepada rekan-rekan seperjuangan S1 Aje, Carin, Dwika, Eka, Ica, Ika, Ina, dan Lisa, rekan-rekan S2 Bang Baso, Pak Ari, Cak Rudi, Mbak Wiwin, Mbak Isni, Mbak Uuf, Mbak Efah, Mbak Nadea, Mbak Tiwi, Mbak Fajri, Mbak Destik, Mas Wawan, Mbak Lia, Mbak Nurul, dan Mbak Nuril dan rekan-rekan S3 Pak Asri, Pak Ilyas, Ibu Ifah, dan Ibu Ida serta teman-teman biologi 47 dan teman-teman kost Perwira 6. Ungkapan terimakasih juga penulis sampaikan kepada seluruh keluarga atas segala doa dan dukungan yang diberikan.
Semoga karya ilmiah ini bermanfaat.
Bogor, September 2014
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vi
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
BAHAN DAN METODE 2
Waktu dan Tempat 2
Bahan 2
Metode 3
Perbanyakan planlet 3
Perbanyakan Agrobacterium tumefaciens 3
Transformasi genetik kentang kultivar Baraka 3
Aklimatisasi 3
HASIL DAN PEMBAHASAN 4
Pembentukan kalus 4
Pembentukan tunas transgenik putatif 4
SIMPULAN DAN SARAN 9
Simpulan 9
Saran 10
UCAPAN TERIMA KASIH 10
DAFTAR PUSTAKA 10
LAMPIRAN 12
DAFTAR TABEL
1 Rata-rata efisiensi transformasi kentang kultivar Baraka 7 2 Rata-rata efisiensi regenerasi kalus dan jumlah tunas setiap kalus
kentang kultivar Baraka 8
DAFTAR GAMBAR
1 Posisi Gen Hd3a pada daerah T-DNA di dalam plasmid p2K1-Hd3a 2 2 Bentuk kalus dari eksplan batang kentang kultivar Baraka hasil
transformasi dengan A. tumefaciens yang mengandung p2K1-Hd3a 4 3 Pembentukan tunas kentang kultivar Baraka di media regenerasi 5
Tunas kentang yang di tanam di media seleksi yang mengandung 40
mg/L higromisin 6
4 Uji viabilitas tunas kentang non-transgenik dan media seleksi 6 5 Tanaman kentang kultivar Baraka in vitro yang berumur 4 minggu 9 6 Tanaman kentang kultivar Baraka transgenik putatif yang ditanam di
polibag
9
DAFTAR LAMPIRAN
1 Komposisi Media MS (Murashige dan Skoog 1962) 12 2 Perhitungan persentase efisiensi transformasi dan efisiensi
PENDAHULUAN
Latar Belakang
Kentang (Solanum tuberosum L.) merupakan salah satu komoditas yang potensial untuk diversifikasi pangan. Kentang mengandung karbohidrat yang tinggi sehingga dapat digunakan sebagai alternatif makanan pokok. Selain itu kentang dapat dijadikan bahan baku industri makanan olahan.
Menurut Badan Pusat Statistik (BPS 2014), produktivitas tanaman kentang Indonesia pada tahun 2012 adalah 16.6 ton/ha dan pada tahun 2013 adalah 16.7 ton/ha. Produktivitas ini lebih rendah daripada rata-rata produktivitas dunia yang pada 2012 adalah sebesar 18.9 ton/ha dan Australia sebesar 38.3 ton/ha (FAOSTAT 2014). Rendahnya produksi kentang di Indonesia dapat disebabkan oleh beberapa faktor, diantaranya adalah rendahnya kualitas bibit, dan teknik budidaya (Wattimena 1992) serta tingginya tingkat serangan hama dan penyakit (Mahmud 1990). Penggunaan varietas yang unggul sangat penting dalam meningkatkan produksi kentang. Varietas unggul dapat diperoleh melalui persilangan konvensional, dan rekayasa genetika. Bunga sangat penting di dalam persilangan konvensional sehingga pembungaan mempunyai peranan yang sangat penting dalam pemuliaan tanaman.
Pembungaan merupakan transisi dari fase vegetatif ke fase generatif/reproduksi. Keadaan cuaca sangat berpengaruh terhadap pembungaan tanaman kentang. Kentang berbunga pada suhu 15-20°C, kelembaban udara yang tinggi dan cukup sinar matahari, serta sedikit hujan (Sahat 1991). Untuk berbunga, tanaman kentang memerlukan hari panjang yaitu sekitar 16-18 jam.
Gen Hd3a pertama kali diidentifikasi sebagai quantitative trait locus (QTL) yang proteinnya dapat menginduksi pembungaan padi pada kondisi hari pendek (Kojima et al. 2002). Gen Hd3a yang berasal dari padi telah diisolasi oleh Tamaki
et al. (2007) yang mempunyai homologi dengan gen flowering locus T (FT) dari
Arabidopsis thaliana. FT/Hd3a merupakan sinyal florigen pembungaan bertipe cepat (Tamaki et al. 2007). Protein Hd3a berinteraksi dengan faktor transkripsi bZIP untuk menginduksi transisi dari fase vegetatif ke fase reproduktif di dalam meristem apikal (Tsuji et al. 2008). Komiya et al. (2008) menunjukkan bahwa supresi Hd3a dengan RNA interference (RNAi) dapat menunda pembungaan. Introduksi gen Hd3a dapat menginduksi pembungaan pada tanaman seperti
Saussurea involucrata (Li et al. 2011), Jatropha curcas L (Sulistyaningsih 2012) dan Nicotiana benthamiana (Syafitri 2012; Senjaya 2013).
Transformasi genetik kentang dengan menggunakan Agrobacterium tumefaciens telah banyak dilakukan, diantaranya yang dilakukan oleh Nurhasanah
et al. (2003) dan Manguntungi (2014). Navarro et al. (2011) telah mengekspresikan Hd3a secara berlebihan di kentang kultivar Andigena yang merupakan kentang hari pendek. Kentang transgenik yang dihasilkan dapat berbunga dan berumbi di daerah hari panjang, sedangkan tanaman kentang non-transgenik tidak berbunga dan tidak berumbi. Hal ini menunjukkan bahwa Hd3a
2
Tujuan Penelitian
Penelitian ini bertujuan untuk melakukan transformasi genetik kentang (Solanum tuberosum L.) kultivar Baraka dengan gen pembungaan Hd3a di bawah kendali promoter kuat prolC.
BAHAN DAN METODE
Waktu dan Tempat
Penelitian ini dilaksanakan dari bulan Desember 2013 sampai dengan bulan Mei 2014 di Laboratorium Biotechnology Research Indonesia-The Netherlands
(BIORIN), dan Laboratorium Biologi Molekuler dan Seluler Tanaman (BMST), Pusat Penelitian Sumberdaya Hayati dan Bioteknologi (PPSHB), Lembaga Penelitian dan Pengabdian kepada Masyarakat (LPPM), Institut Pertanian Bogor.
Bahan
Kentang (Solanum tuberosum L.) kultivar Baraka in vitro digunakan sebagai bahan tanaman untuk transformasi genetik. Agrobacterium tumefaciens galur LB A4404 yang mengandung plasmid p2K1 yang membawa gen Hd3a di bawah kendali promoter prolC (Tamaki et al. 2007) yang dipautkan dengan gen penanda
GFP serta gen resistensi terhadap higromisin (hpt) digunakan untuk melakukan transformasi genetik. Peta fisik daerah T-DNA dari plasmid p2K1-Hd3a disajikan pada Gambar 1. Plasmid atau vektor ekspresi ini adalah pemberian Prof. Ko Shimamoto (Nara Institute of Science & Technology, Jepang).
Gambar 1 Posisi Gen Hd3a pada daerah T-DNA di dalam plasmid p2K1-Hd3a.
3 Metode
Perbanyakan tanaman in vitro
Tanaman kentang in vitro diperbanyak dengan stek ruas (node) tunggal yang mengandung satu mata tunas di media MS2 makro. Media MS2 makro adalah
media MS yang mengandung 2x konsentrasi unsur hara makro ditambah 2 ml/L
pantothenic acid ditumbuhkan selama 4 minggu di ruang kultur dengan suhu 24o -25oC dan pencahayaan 2000-3000 lux serta fotoperiode 16 jam.
Perbanyakan Agrobacterium tumefaciens
Agrobacteruim tumefaciens strain LBA4404 yang mengandung plasmid p2K1-Hd3a yang mengandung gen Hd3a di bawah kendali promoter prolC
(Tamaki et al. 2007) dibiakkan selama 36 jam pada media LB (1% tripton, 0,5%
yeast extract, 1% NaCl) cair yang mengandung 50 mg/L kanamisin dan 25 mg/L rifampisin, dengan penggoyangan pada kecepatan 150 rpm, suhu 28oC, di dalam gelap. Sebanyak 100 µL dari biakan tersebut kemudian dibiakkan kembali di dalam 10 mL media LB cair dengan kondisi yang sama dengan sebelumnya hingga mencapai nilai OD600 sebesar 0,5-0,7.
Transformasi genetik kentang kultivar Baraka
Transformasi dilakukan dengan metode ko-kultivasi yang menggunakan bantuan A. tumefaciens. Sebelum ko-kultivasi, biakan A. tumefaciens
disentrifugasi pada kecepatan 5000 rpm selama 15 menit dan endapan yang diperoleh diresuspensi dengan 10 ml media ko-kultivasi cair (media MS cair yang mengandung 0.5 mg/L BAP, 0.1 mg/L IAA, dan 20 mg/L Asetosiringon) hingga OD600 0.5-0.7. Eksplan yang ditransformasi adalah potongan daun berukuran 1
cm2 dan buku (internode) berukuran 0.5-1 cm. Ko-kultivasi dilakukan dengan merendam eksplan di dalam suspensi A. tumefaciens, selama 10 menit, digoyang dengan kecepatan 100 rpm, pada suhu ruang. Eksplan dikeringkan dengan tisu steril, kemudian ditanam di media ko-kultivasi padat selama 3 hari di ruang gelap. Setelah itu eksplan dicuci dengan akuades steril sebanyak empat kali dan pada bilasan terakhir ekplan direndam selama 10 menit dengan menambahkan 100 mg/l larutan cefotaxime. Eksplan dikeringkan dengan tisu steril dan kemudian ditanam di media induksi kalus yang juga merupakan media regenerasi tunas yaitu media MS yang mengandung 3 mg/L BAP, 2 mg/L IAA, 1 mg/L GA3, 100 mg/L
cefotaxime, dan 3 g/L agar gelrite. Subkultur dilakukan setiap 12 hari. Tunas yang terbentuk, dipindahkan ke media seleksi yaitu media MS yang mengandung 10 mg/L higromisin. Pada subkultur selanjutnya, konsentrasi higromisin ditingkatkan hingga 40 mg/L. Eksplan yang menghasilkan tunas transgenik putatif dihitung untuk mengetahui efisiensi transformasi dan efisiensi regenerasi (Lampiran 2).
Eksplan non transgenik (kontrol) yang tidak ditransformasi dengan A. tumefaciens ditanam dalam media seleksi higromisin yang sama dengan eksplan yang di transformasi dengan A. tumefaciens sebagai kontrol efisiensi media seleksi.
Aklimatisasi
4
berukuran 3 cm x 3 cm x 5 cm yang berisi coco-peat. Setelah agak besar, sekitar 3 minggu, tanaman dipindah ke dalam polibag yang mengandung tanah, pasir dan pupuk kandang dengan perbandingan 1:1:1 (volume).
HASIL DAN PEMBAHASAN
Pembentukan kalus
Eksplan yang digunakan dalam proses transformasi adalah potongan daun yang berukuran 1 cm2 dan buku batang tanpa mata tunas yang berukuran 0.5-1
cm. Penggunaan buku tanpa mata tunas bertujuan untuk menghindari tumbuhnya tunas palsu dari bagian buku eksplan ketika ditanam di media regenerasi. Induksi kalus dan regenerasi menggunakan media yang sama yaitu media MS yang mengandung 3 mg/L BAP, 2 mg/L IAA, 1 mg/L GA3, 100 mg/L cefotaxime, dan 3 g/L agar gelrite, seperti yang digunakan oleh Puspita (2002) dan Manguntungi (2014).
Hasil penelitian menunjukkan bahwa eksplan buku lebih cepat membentuk kalus daripada eksplan daun yaitu pada minggu ke-2 setelah tanam. Kalus terbentuk mulai dari kedua ujung buku batang yang dilukai kemudian merata ke seluruh bagian eksplan (Gambar 2). Pada eksplan daun, kalus mulai terbentuk pada minggu ke-3 setelah tanam, dari bagian tangkai lalu kemudian menyebar ke seluruh permukaan daun. Penambahan IAA dan BAP pada penelitian ini dapat memacu pembentukan kalus seperti yang dilakukan oleh Li et al. (2007). Menurut Hidayat (2009) jaringan meristematik pada batang lebih banyak daripada di daun sehingga proses pembelahan dan pemanjangan sel di batang lebih cepat dari pada di daun.
Gambar 2 Bentuk kalus dari eksplan batang kentang kultivar Baraka hasil transformasi dengan A. tumefaciens yang mengandung p2K1-Hd3a Pembentukan tunas transgenik putatif
Setiap 12 hari setelah penanaman di media induksi kalus, eksplan disubkultur secara berulang pada media yang sama. Subkultur yang dilakukan setiap 12 hari bertujuan untuk memacu pembentukan tunas. Pada bagian yang terluka pada saat infeksi, kalus menghasilkan tonjolan-tonjolan yang kemudian
5 peranan yang penting dalam memicu terbentuknya tunas. Buku batang kemungkinan mengandung zat pengatur tumbuh lebih banyak dari pada daun.
Gambar 3 Pembentukan tunas kentang kultivar Baraka di media regenerasi. Regenerasi tunas dari kalus yang terdapat pada: (a) ujung buku batang, (b) tengah buku batang, dan (c) tangkai daun
Ko-kultivasi eksplan kentang dengan A. tumefaciens yang mengandung gen
Hd3a yang dipautkan dengan gen hpt telah menghasilkan tanaman yang resisten terhadap higromisin. Hal ini menunjukkan bahwa proses transformasi kentang dengan gen Hd3a yang dipautkan dengan gen hpt telah berhasil dan menghasilkan tanaman kentang transgenik putatif. Transformasi genetik dengan menggunakan perantara A. tumefaciens merupakan salah satu metode yang banyak digunakan karena menghasilkan tanaman transgenik yang stabil (Li et al. 2007).
6
transgenik dilakukan pada media yang mengandung 30 mg/L higromisin ( Syafitri 2012).
(a) (b)
Gambar 4 Tunas kentang yang ditanam di media seleksi yang mengandung 40 mg/L higromisin. (a) tunas transgenik, (b) tunas non-transgenik
Untuk membuktikan bahwa tanaman kentang non-transgenik mempunyai viabilitas yang baik dan media seleksi berfungsi dengan baik, tanaman non-transgenik ditanam di dua macam media MS yang berbeda yaitu: (1) media yang tidak mengandung higromisin dan (2) media yang mengandung higromisin sebagai media seleksi. Pada media yang tidak mengandung higromisin, tunas kentang non-transgenik tumbuh dengan baik, sedangkan pada media yang mengandung higromisin, tunas kentang mengalami kematian (Gambar 5). Hal ini menunjukkan bahwa tanaman kentang non-transgenik mempunyai viabilitas yang baik, dan media seleksi berfungsi dengan baik.
(a) (b)
Gambar 5 Uji viabilitas tunas kentang non-transgenik dan media seleksi. (a) tunas non-transgenik yang berumur 5 minggu yang ditanam di media yang tidak mengandung higromisin, (b) tunas non-transgenik berusia 5 minggu yang ditanam di media seleksi yang mengandung higromisin
1 cm
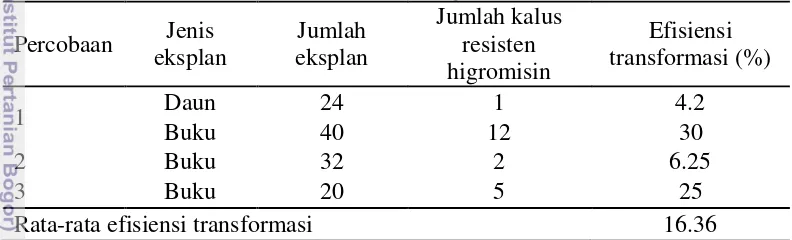
7 Pada penelitian ini, efisiensi transformasi kentang kultivar Baraka berkisar antara 4.2% dan 27% dengan rata-rata 16.36% (Tabel 1). Efisiensi transformasi ini lebih tinggi dibandingkan dengan transformasi genetik kentang kultivar Russet Burbank, Lehmi Russet, dan Atzimba dengan gen Bar (Dinarti 1999), kultivar Atlantik dengan gen hordothionin (Nurhasanah 2003), dan relatif sama dengan transformasi genetik kentang kultivar Atlantik dengan gen c-lysozyme
(Manguntungi 2014), tetapi lebih kecil dibandingkan dengan transformasi pada
Jatropha curcas yaitu 26.67% (Sulistyaningsih 2012) dan pada Nicotiana benthamiana yaitu 86% (Syafitri 2012). Efisiensi transformasi dipengaruhi oleh spesies dan kultivar yang digunakan. Rendahnya efisiensi transformasi pada penelitian ini disebabkan oleh banyaknya eksplan yang mati karena terinfeksi A. tumefaciens yang sulit dihilangkan dengan perlakuan cefotaxime. Rusaknya jaringan dari eksplan selama proses transformasi dapat menyebabkan kematian eksplan. Selain itu, perlakuan cefotaxime yang terus menerus untuk menghilangkan A. tumefaciens juga dapat menghambat pertumbuhan dan bahkan mematikan eksplan kentang.
Tabel 1 Rata-rata efisiensi transformasi kentang kultivar Baraka Percobaan Jenis
Efisiensi transformasi dari eksplan daun lebih kecil daripada eksplan buku (Tabel 1). Hal ini terjadi karena eksplan daun mempunyai jaringan yang sangat tipis sehingga mudah rusak pada saat proses transformasi. Cadangan makanan di daun lebih sedikit daripada di batang (Santoso dan Nursandi 2001) sehingga daun lebih sulit untuk beregenerasi daripada batang.
8
meningkatkan frekuensi pembentukan tunas maupun jumlah tunas yang dihasilkan (Sulistiyaningsih 2012).
Tabel 2 Rata-rata efisiensi regenerasi kalus dan jumlah tunas setiap kalus kentang kultivar Baraka
Rata-rata efisiensi regenerasi 100 48 2.5
Jumlah tunas transgenik putatif yang dihasilkan tiap kalus kentang kultivar Baraka pada penelitian ini adalah 2.5 (Tabel 2). Hasil ini relatif sama dengan yang diperoleh Listanto et al. (2009).
Di media MS0, empat dari 25 tanaman transgenik putatif menghasilkan
umbi sedangkan tunas non-transgenik di media yang sama tidak menghasilkan umbi. Umbi terbentuk pada kentang transgenik putatif yang berumur 4 minggu (Gambar 6). Hasil ini menyerupai hasil penelitian Navarro et al. (2011) yang menunjukkan bahwa Hd3a selain dapat menginduksi pembentukan bunga juga dapat menginduksi terbentuknya umbi pada kentang kultivar Andigena, yang merupakan kentang hari pendek dan berumur genjah, bila ditanam di hari panjang. Kentang Andigena non-transgenik tidak berbunga dan tidak berumbi bila ditanam di daerah yang mempunyai hari panjang. Kentang kultivar Andigena berbeda dengan Baraka yang merupakan kultivar yang berumur dalam. Walaupun kultivar Andigena dan Baraka berbeda tipe umurnya, tetapi Hd3a mempunyai pengaruh yang relatif sama yaitu menginduksi pembentukan umbi yang lebih awal dibandingkan dengan tanaman non-transgenik. Hal ini menunjukkan bahwa Hd3a dapat menginduksi pembentukan umbi pada kentang walaupun harus dibuktikan lagi pada kultivar kentang lainnya.
9
(a) (b)
Gambar 6 Tanaman kentang kultivar Baraka in vitro yang berumur 4 minggu. (a) tanaman transgenik putatif, (b) tanaman non-transgenik. = umbi
Gambar 7 Tanaman kentang kultivar Baraka transgenik putatif berumur 3 bulan yang ditanam di polibag
Menurut Navarro et al. (2011), gen Hd3a dtranskripsikan dan translasikan di dalam jaringan daun kentang, kemudian protein Hd3a tersebut disalurkan melalui floem ke daerah pucuk apikal untuk menginduksi pembungaan dan disalurkan ke daerah stolon untuk menginduksi pembentukan umbi. Protein Hd3a memiliki kemampuan untuk mempercepat pengumbian melalui mekanisme aktivasi gen-gen pengumbian yang homolog dengan gen pembungaan FT pada A. thaliana.
SIMPULAN DAN SARAN
Simpulan
Transformasi genetik kentang kultivar Baraka telah berhasil dilakukan dan menghasilkan 48 tunas transgenik putatif. Efisiensi transformasi pada penelitian ini relatif rendah yaitu 16.36%, dan efisiensi regenerasinya sangat tinggi yaitu 100%. Efisiensi transformasi pada eksplan buku lebih tinggi daripada eksplan
1 cm
10
daun. Di media MS0, empat dari 24 tanaman transgenik putatif menghasilkan
umbi, yang menunjukkan bahwa Hd3a menginduksi pembentukan umbi.
Saran
Tanaman transgenik putatif perlu dianalisis untuk mengetahui integrasi gennya.
UCAPAN TERIMA KASIH
Terima kasih disampaikan kepada Proyek Penelitian Desentralisasi Baru
IPB yang berjudul “Rekayasa genetika tanaman dengan gen toleransi aluminium
dan pembungaan” dengan kontrak no: 48/IT3.11/LT/2014 atas nama: Prof Dr Ir Suharsono, DEA yang telah membiayai penelitian ini.
DAFTAR PUSTAKA
Dinarti D. 1999. Efisiensi transfer gen penanda nptII atau bar menggunakan isolat
Agrobacterium oncogenik dan disarmed pada tiga kultivar kentang. [tesis]. Bogor (ID): Program Pascasarjana, IPB
BPS [ Badan Pusat Statistik ]. 2014. Luas Panen, Produktivitas dan Produksi Kentang. [Internet]. [diunduh 2014 Juni 4]. Tersedia pada: http:www.bps.go.id.
FAOSTAT [Food and Agriculture Organization of the United States]. 2012. The Agricultural Production of Potatoes. [Internet]. [diunduh 2014 Juli 4]. Tersedia pada: http:www.faostat3.foo.org/faostat-gateway/gp/to/download/Q/QC/E.
Gaba V, Schlarman E, Elman C, Sagee O, Watad AA, Gray DJ. (1998) In vitro studies on the anatomy and morphology of bud regeneration in melon cotyledons. In Vitro Cellular and Developmental Biology - Plant 35: 1-7 Hidayat O. 2009. Kajian penggunaan hormon IBA, BAP dan kinetin terhadap
multiplikasi Tunas Tanaman Penghasil Gaharu (Gyrinops versteegii (Gilg) Domke) secara In Vitro. [skripsi]. Bogor (ID): Fakultas Kehutanan, IPB. Kojima et al. 2002. Hd3a, a rice ortholog of the arabidopsis FT gene, promotes
transition to flowering downstream of Hd1 under short-day conditions.
Plant Cell Physiol 43:1096–1105.
Komiya R, Ikegami A, Tamaki S, Yokoi S, Shimamoto K. 2008. Hd3a and RFT1
are essential for flowering in rice. Development 135:767-774. doi:10.1242/dev.008631
Li M, Li H, Jiang H, Pan X, Wu G. 2007. Establishment of an Agrobacterium
mediated cotyledon disc transformation method for Jatropha curcas. Plant
11 Li M, Li H, Hu X. 2011. Genetic transformation and overexpression of a rice
Hd3a induces early flowering in Saussurea involucrata Kar.et Kir. ex Maxim. Plant Cell Tiss Organ Cult. 106: 363-371.
Listanto E, Wattimena GA, Armini NM, Sinaga MS, Sofiari E, Herman M. 2009. Regenerasi beberapa kultivar kentang dan transformasi kentang dengan Gen RB melalui Agrobacterium tumefaciens. J. Hort. 19(2): 137-147. Mahmud M. 1990. Penyakit Bakteri Tanaman Pangan dan Hortikultura di
Indonesia. Jakarta (ID): PT Agricon.
Manguntungi AB. 2014. Transformasi genetik kentang ‘Atlantik’ dengan gen
penyandi lisozim melalui perantara Agrobacterium tumefaciens. [makalah seminar]. Bogor: Sekolah Pascasarjana, IPB.
Murashige T, Skoog F. 1962. A revised medium for rapid growth and bioassays with tobacco culture. Physiol Plant. 15: 473-497.
Navarro C, Abelanda JA, Cruz EO, Carlos A, Tamaki S, Silva J, Shimamoto K, Prat S. 2011. Control of flowering and storage organ formation in potato by flowering locus T. Nature 478: 119-123. doi: 10.1038/nature 10431. Nurhasanah, Wattimena GA, Purwito A, Wiendi NMA, Suharsono. 2003.
Transformasi genetik tanaman kentang cv. Atlantik dengan mengintroduksikan gen hordothionin untuk mendapatkan ketahanan terhadap penyakit bakteri. Buletin Agronomi 31 (2): 63-67.
Puspita T. 2002. Studi regenerasi kentang (Solanum tuberosum) kultivar desiree secara in vitro. [skripsi]. Bogor (ID): Fakultas Pertanian, IPB.
Sahat S. 1991. Pengaruh cara stimulasi perbungaan terhadap produksi bunga, buah, dan biji beberapa kultivar kentang (Solanum tuberosum L.). Bull Penel Hort. 20:105-111.
Senjaya SK. 2013. Pewarisan genetik transgen Hd3a pada tanaman Nicotiana benthamiana transgenik. [skripsi]. Bogor (ID): Fakultas Matematika dan Ilmu Pengetahuan Alam, IPB.
Smith O. 1968. Potatoes: Production, storing, processing. London (GB): Avi Publishing Company.
Sulistyaningsih YC. 2012. Rekayasa ekspresi gen pembungaan Hd3a pada tanaman jarak pagar (Jatropha curcas L.). [disertasi]. Bogor: Sekolah Pascasarjana, IPB.
Syafitri LN. 2012. Transformasi genetik Nicotiana benthamiana dengan gen pembungaan Hd3a dari padi. [skripsi]. Bogor (ID): Fakultas Matematika dan Ilmu Pengetahuan Alam, IPB.
Tamaki S, Matsuo S, Wong H, Yokoi S, Shimamoto K. 2007. Hd3a protein is a mobile flowering signal in rice. Science 36:1033-1036.
Tsuji H, Tamaki S, Komiya R, Shimamoto K. 2008. Florigen and the photoperiodic control of flowering in rice. Rice 1:25-35.
12
Lampiran 1 Komposisi Media MS (Murashiege dan Skoog 1962)
Bahan Konsentrasi
Senyawa dalam media (mg/L)
Volume yang dipipet (ml/L)
NH4NO3 1650 20
KNO3 1900 20
KH2PO4 170 5
H3BO3 6.2
Na2MoO4.2H2O 0.25
CoCl2.6H2O 0.025
KI 0.83
CaCl2.2H2O 440 5
MgSO4.7H2O 370 5
MnSO4.4H2O 22.3
ZnSO4.7H2O 8.6
CuSO4.5H2O 0.025
Na2EDTA 37.3 5
FeSO4.7H2O 27.8
Thiamine-HCl 0.1 5
Niacin (asam nikotinat)
0.5 Pyridoxine-HCl 0.5
Glycine 2.0
Myo inositol 100 10
Gula pasir 30 g/L
13 Lampiran 2 Perhitungan persentase efisiensi transformasi dan efisiensi regenerasi
% efisiensi transformasi = ℎ ℎ
ℎ � � %
14
RIWAYAT HIDUP
Penulis dilahirkan di jakarta pada tanggal 24 Februari 1993 dari ayah Tirsat bin Langen (Alm) dan ibu Sumaliyah (Alm). Penulis merupakan anak tunggal.
Tahun 2010 penulis lulus dari SMA Negeri 52 jakarta dan pada tahun yang sama lulus seleksi masuk IPB melalui jalur Undangan Seleksi Masuk IPB (USMI). Penulis terpilih masuk Program Studi Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam dan mendapatkan beasiswa dari Dikti yaitu beasiswa bidik misi.
Selama mengikuti perkuliahan, penulis aktif di Koperasi Mahasiswa (KOPMA) sebagai staf Riset dan Pengembangan pada tahun 2011/2012 dan aktif di Himpunan Mahasiswa Biologi (HIMABIO) sebagai staf divisi Pengembangan sumber daya manusia (PSDM) pada tahun 2012/2013. Penulis pernah melaksanakan studi lapang di Kebun Raya Cibodas, Taman Nasional Gunung Gede-Pangrango pada tahun 2012, dengan judul makalah “Invasif Alien Species di
Kebun Raya Cibodas”. Penulis juga pernah melakukan magang dengan topik isolasi DNA kedelai transgenik di BB Biogen Bogor pada bulan Januari-Februari 2013. Pada bulan Juni-Juli 2013 penulis melakukan praktik lapangan di Taman