L
Journal of Experimental Marine Biology and Ecology, 241 (1999) 67–80
Ontogeny of shell-related behaviors and transition to land in
the terrestrial hermit crab Coenobita compressus H. Milne
Edwards
* Renae J. Brodie
Department of Zoology, University of Washington, Seattle, WA 98195-1800, USA Received 14 September 1998; received in revised form 8 April 1999; accepted 5 May 1999
Abstract
The marine megalopa of the terrestrial hermit crab Coenobita compressus wears a small gastropod shell that it carries onto land before metamorphosing into a juvenile crab. This paper gives the first detailed account of temporal shifts in shell-related behaviors that occur as the megalopa approaches metamorphosis. Megalopae placed in individual containers shortly after metamorphosing from the last zoeal stage exhibited three behaviors: shell inspection, followed by shell wearing in water, and finally by shell wearing on land. Megalopae experimentally denied access to shells exited the water at about the same time as megalopae with shells, but they wandered about on land for shorter periods of time before burying themselves in sand and attempting to metamorphose. Once buried, megalopae without shells often emerged and re-buried themselves at other locations in their containers; almost all shell-less megalopae died without metamorphosing to the first juvenile crab stage. Once buried, megalopae with shells rarely emerged without first metamorphosing to the juvenile crab stage, and they metamorphosed significantly more frequently than those denied shells. 1999 Elsevier Science B.V. All rights reserved.
Keywords: Behavior; Land hermit crab; Megalopae; Metamorphosis; Shell; Coenobita
1. Introduction
The larvae of many benthic invertebrates swim and feed in the plankton before they settle to the sea floor and metamorphose into juveniles (Thorson, 1950). In the
*Corresponding author. Tel.: 11-206-543-1629; fax: 11-206-543-3041.
E-mail address: [email protected] (R.J. Brodie)
laboratory, these larvae often change their behaviors shortly before metamorphosis when, under more natural conditions, they would be moving from plankton to benthos. For example, many invertebrates begin early larval life positively phototactic, but later show diminished responses to light or become photonegative (Thorson, 1964; Lang et al., 1979; Botero and Atema, 1982; Marsden, 1984; Miller and Hadfield, 1986). They may also respond differently to pressure changes over time (Cragg, 1980; Sulkin, 1984). The investigators who have characterized and measured these behavioral changes have considered them to be the means by which larvae leave the plankton and encounter a suitable benthic habitat.
Like their marine counterparts, land hermit crabs pass through several zoeal stages in the plankton after hatching (Hartnoll, 1988). Then, once they metamorphose into megalopae (hereafter also referred to as postlarvae; Harvey, 1992a), they begin their journey towards land where they will bury themselves and later emerge as terrestrial juveniles (Reese and Kinzie, 1968; Harvey, 1992b). To succeed in the plankton-to-land transition, megalopae must perform several tasks: 1) shift from the plankton to the benthos and move towards an appropriate terrestrial habitat; 2) find a shell; and finally, 3) crawl onto land with it and metamorphose. Thus, by changing their behaviors over time, megalopae destined to become terrestrial adults maximize their chances of arriving on a suitable beach.
Although the literature is sparse, shifts in behavior by megalopae approaching metamorphosis have been anecdotally noted for both marine and terrestrial hermit crab postlarvae. For example, laboratory-reared megalopae of the land crab Coenobita purpureus entered shells after ten days as postlarvae (Nakasone, 1988). Thompson (1903) observed that megalopae of Eupagurus sp. generally had a short free swimming period and then became increasingly interested in shells as they approached metamor-phosis. In keeping with Thompson’s observation, Reese (1968) noticed that older megalopae of Birgus latro swam less as they aged.
These shifts in behavior are accompanied by changes in internal and external morphology. The megalopal stage is a period of profound internal rearrangement: the positions of the digestive glands, gonads, and green glands shift, and the circulatory system and musculature are modified (for Eupagurus in Thompson, 1903). Color patterns resembling those of the adults begin to appear in later megalopae and they become less translucent (pers. obs.). Thus, while the outer form remains static between the metamorphic molts, a cascade of changes during the megalopal stage presages the adult.
2. Methods and materials
2.1. Larval rearing
I collected 324 newly hatched zoeal stage larvae from nine different female terrestrial hermit crabs for this study. To accomplish this, gravid Coenobita compressus were intercepted just above the surf zone on Culebra beach, Naos Island, Panama, shortly before high tide on the evenings of April 1–3, 1997. These crabs were placed inside plastic washtubs partly filled with damp sand where each was provided a Plexi-glass dish filled with filtered ocean water. On each of the three nights, three adult female crabs shed thousands of zoeae larvae into their water dishes; these zoeae were combined in a larger Plexi-glass bowl and 108 individuals out of the thousands of larvae in the bowl were randomly removed and placed in plastic rearing containers. All subsequent rearing and experimentation took place at the Smithsonian Tropical Research Institute’s marine facilities on Naos Island, Panama.
The zoeae were housed individually in compartmentalized plastic boxes holding about 20 ml of filtered ocean water per compartment and incubated at 278C on a 12:12 L:D light cycle, until they reached the megalopal stage after 3 or 4 molts and the experiment commenced. Zoeae were transferred to clean compartments and fed Artemia nauplii daily; the larvae consumed newly hatched Artemia for the first two weeks, after which they fed upon larger 1–2 day old Artemia.
2.2. Experimental procedure
When individual larvae molted to the megalopal stage they were transferred the next day to experimental containers and relocated outside to an open but covered arena, where they experienced natural daily fluctuations in temperature and filtered sunlight. Each experimental container had both an ocean and land environment: a plastic cup holding 10 ml of sea water was sunk into the damp sand filling a larger clear plastic cup (diameter59 cm; Fig. 1). A clear plastic top covered the container. The sand used in the plastic cups was collected from a beach inhabited by hundreds of adult C. compressus. A small amount of this sand (less than 1 g) was also added to the cups’ ‘‘oceans’’ to ensure that the megalopae were exposed to adult odors which might be necessary for metamorphosis (Crisp, 1974). Megalopae were transferred to the water-filled cups from which they could emerge onto land via any of four symmetrically placed lines of hot melt glue extending from the bottom of the cup to the edge of the terrestrial environment. Megalopae were fed fish flakes in the water and on land.
Fig. 1. Megalopal habitat. Megalopae were introduced into the water dishes of the individual containers one day after molting; they could leave the water at any time by climbing up one of the four lines of hot melt glue.
To ascertain whether megalopae without shells would delay land transition and metamorphosis, I provided one half of the megalopae access to shells (hereafter referred to as the shell-treatment; total of 108 replicates), while the other half were denied access (the shell-less treatment; total of 107 replicates). To ensure that megalopae in the shell-treatment would have adequate access to shells in both environments, I placed one cleaned Nerita modesta shell on the damp sand in each container and two N. modesta shells in the water. The N. modesta shells were on average 4.160.2 mm (mean6SD; N515) in height with average aperture lengths of 2.460.1 mm (N515; for measuring shell dimensions see Reid, 1996).
beneath their bodies. In contrast, early megalopae extend their abdomens which bear swimming appendages (the pleopods) and often swim or rest on the substratum with abdomens straight and lifted slightly above the rest of the body. Although this is not a shell-related behavior, I measured it to determine if megalopae show other, more general, adult-like behaviors as they approach metamorphosis. This behavior was only scored if the megalopa was in the water and not wearing a shell. Megalopae in the shell-less treatment were also scored for number 4.
To determine if megalopae deprived of shells would delay or forgo land transition, individuals in both treatments were additionally scored for either occupying their water dishes or the damp sand outside of the dishes. I controlled for possible endogenous behavioral responses to changes in tidal height (Tankersley and Forward, 1994; Tankersley et al., 1995; Zeng and Naylor, 1996), by observing the postlarvae no sooner than one hour before nor later than one hour after the daylight high tide. I terminated the observations for each megalopa when it either buried itself on land in preparation for the metamorphic molt to the first young crab stage or died.
I observed megalopae in their containers through a Field Model Questar telescope (adjusted to 803) from a distance of about 2 m, while standing about the same distance above the containers on a table. To examine individual animals, I scanned each habitat until I located the megalopa, then observed it for 2–3 s and recorded its behaviors. Megalopae positioned near the proximal edge of either the water dish or the container could not be seen from the table; I approached these individuals very cautiously at the end of the observation period and examined them (13% of the data were collected this way). Container tops were removed 10–15 min before the observations began and replaced after all of the megalopae had been scrutinized. The containers were cleaned and megalopae fed shortly after the daily data collection. Individual containers were assigned positions haphazardly regardless of the treatment and these positions were shuffled during the daily cleaning and feeding sessions.
2.3. Statistical analysis
3. Results
3.1. Larval rearing and introduction into the experiment
The zoea larvae molted to megalopae 22–35 days after the first ovigerous adult females shed them. Of the total, fifty-two passed through four zoeal stages lasting 21–28 days (mean6SD524.1761.35) while 163 passed through five zoeal stages lasting 26–32 days (28.2761.07). Sixty-nine percent of the zoeae reached the megalopal stage. The first 215 megalopae to appear were placed in individual experimental containers where 108 were given shells and 107 were not.
3.2. Emigration from ‘‘ocean’’ to ‘‘land’’
Megalopae were first observed to leave their water dishes after two days, whether or not they had access to shells. By day twelve all of the surviving megalopae had come onto land at least once (Fig. 2). Megalopae in both treatments left their water dishes after an average of 6 days (shell treatment56.362.4, N589; shell-less treatment5
6.562.5, N589; U-test: p50.68).
Once on land, megalopae often returned to the water; of the 56 that eventually attempted to metamorphose, only 15 buried themselves on the day they emigrated and 9 remained continuously on land for more than one day before burying themselves. The remainder were observed to shift from water to land 2–6 times before burrowing. However, because I checked the megalopae only once a day, I undoubtedly under-estimated the number of returns to water. Older megalopae were especially mobile and often scrambled back into the water when disturbed during the daily cleanings.
3.3. Postlarval shell-related behaviors
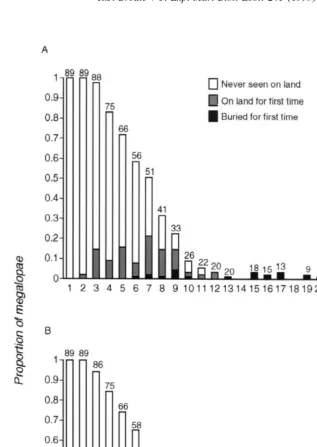
A number of megalopae with access to shells displayed interest in the shells during their first day of postlarval life. They then progressed from probing and manipulating the shells (scored as shell inspection) and donning them in water to emigrating with them onto land (Fig. 3). Postlarvae inspected their shells most often during their first week. After day eight no megalopae were observed inspecting shells during the daily data collections; however, shell inspection probably occurred albeit at a much lower frequency because megalopae occasionally changed the shells that they wore. Only one water dish-dwelling megalopa wore a shell on the first day and three megalopae wore shells on days three and four. The incidence of shell wearing in water greatly increased on day five, when more than 20% of the postlarvae wore the shells in their water dishes, and remained high through day twelve. One megalopa left its water dish wearing a shell on day three but the larger exodus did not begin until day seven. Thereafter, more megalopae wore shells on land than in water, except on day twelve when the numbers were equal (Fig. 3).
Fig. 3. Proportion of megalopae engaging in three shell-related behaviors. Proportions were calculated by dividing the number of megalopae engaged in specific behaviors on a specific day by the total number of megalopae alive on that day. Numbers above columns are the number of megalopae remaining at that age. Megalopae were removed from the study as they died or metamorphosed; as in Fig. 1, buried megalopae were included in the tabulation on the day that they buried themselves but were not included in the subsequent tallies.
suggesting that older postlarvae are more likely to achieve a successful transition from ocean to land. Further evidence of this is discussed below.
3.4. Postlarval expression of crab stance
Megalopae expressed the crab-like stance early in both the shell (on day 3) and shell-less (on day 2) treatments. Thereafter, megalopae assumed this posture sporadically and did not tend to decrease or increase expression of this behavior with age. Thus, adult-like postures are not expressed more frequently as megalopae approach metamor-phosis.
3.5. The final transition from ocean to land: death and metamorphosis
Most postlarval deaths occurred on land; of the 215 megalopae in the experiment, 102 2
died on land (before burying themselves) vs. 53 in the water (X 515.5, df51, 84
]
p,0.001). The death rates before attempted metamorphosis- (108) shell treatment vs. 71
]
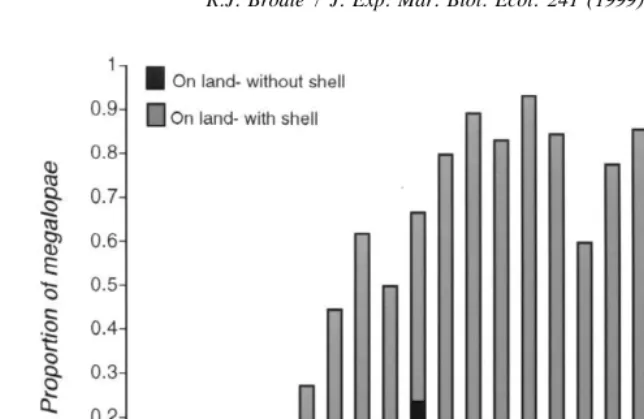
Fig. 4. Proportion of megalopae in shell treatment on land with or without their shells. Proportions were calculated by dividing the number of shelled and unshelled megalopae out of water on a specific day by the total number of megalopae in that treatment alive on that day (total numbers are as in Fig. 3).
left their water dishes during the latter part of the general exodus. The megalopae from the shell treatment that eventually metamorphosed left the water for the first time later (8.761.8 days; N515) than those that died before attempting to metamorphose (5.762.4 days; N565; U-test: p,0.001). Casual observations of older megalopae not used in this study supported this conclusion; younger megalopae appeared sluggish on land and tended to collapse; in contrast, older ones easily climbed small pieces of wood placed on the sand in their containers. Unfavorable conditions within the experimental containers may also have contributed to mortality.
The survival advantage of older postlarvae was also apparent following metamor-phosis. Four eight-day-old megalopae in the shell treatment buried themselves without their shells; older postlarvae that attempted metamorphosis did not make this mistake (Table 1). The fifteen postlarvae with shells that successfully metamorphosed to juvenile crabs burrowed at an average age of 18 days. Nine significantly younger megalopae attempted to metamorphose after about nine days but were unsuccessful (Table 1; U-test: 0.02.p.0.01).
Survival through metamorphosis of megalopae denied access to shells was poor. Only three of the thirty-two megalopae re-emerged as juvenile crabs, although four megalopae that had buried themselves were still alive at the end of the experiment. Furthermore,
17 ]
about half (32) of the naked megalopae emerged and re-buried without metamorphosing 3
]
versus about 15% (24) of those with access to shells.
Table 1
Relative success of metamorphosis for Coenobita compressus megalopae with and without shells
Treatment Metamorphosed Not metamorphosed Still megalopa
With access Buried Buried
with shells before burying themselves once they had exited the water: postlarvae without access to shells spent less time wandering after first exiting the water (3.263.9 days; N532) than megalopae with access to shells (7.065.6 days; N524).
Megalopae with shells remained underground for one to six days before re-emerging as juvenile crabs (N515). Of the shell-less megalopae, one took six days and the other two were under for an undetermined amount of time.
4. Discussion
The terrestrial hermit crab Coenobita compressus has a complex life history spanning both ocean and land environments. The oceanic phase begins when terrestrial adult females bearing embryos approach the sea at night shortly before high tide and release the hatching larvae into the swash zone. These zoeae larvae are washed by the receding tide from the swash zone into the plankton where they develop and molt 3–4 times, then metamorphose into postlarvae (also called megalopae). Like many marine invertebrates, C. compressus postlarvae move to the benthos after developing in the plankton; soon after this descent, however, they acquire a shell and continue their journey into the terrestrial environment.
In my experiments, C. compressus megalopae were able to wear shells from the first day of this stage. This is consistent with other studies of shell-entering behaviors of marine and terrestrial hermit crab megalopae (Reese 1962, 1968; Hazlett and Proven-zano, 1965), but the present study is the first to characterize the temporal sequence of shell-related behaviors.
While this experiment clearly demonstrated the sequence of megalopal shell-related behaviors, the physico-chemical environment may have influenced the absolute timing of their expression. Other studies of anomuran development have noted that the external environment can alter behaviors similar to those examined in this study. For example, Reese (1968) found that Birgus latro postlarvae (also family Coenobitidae) exited their water containers more readily and with fewer mistakes (leaving the shell behind) when surrounded by more natural beaches, complete with sand, coconut husk, soil, plants, meat, and coral pebbles, as opposed to only sand. Jensen (1991) described behavioral changes in the comparable megalopal stage of the porcellanid anomuran crabs, Petrolisthes cinctipes and P. eriomerus. He noted that megalopae exposed to adults very quickly lost the ability to swim. Following their assumption of a more benthic existence, their abdominal swimming appendages, the pleopods, degenerated irreversibly. Megalopae not exposed to adults followed the same process but delayed the physical and behavioral changes for several weeks.
The megalopae in my study were exposed to sand from an adult habitat, but the absolute timing of the behavioral events that I recorded may have been shorter had there been a more elaborate set of cues in the containers. It is also possible that the onset of the behaviors that I described were triggered by the laboratory environment (Wolcott and De Vries, 1994). Under natural conditions it would be highly maladaptive for a land hermit crab megalopa to enter a shell and crawl towards shore if it were still far out at sea; it is possible that all of the behaviors that I described begin after a megalopa senses that it is close to shore and appropriate adult habitat.
Most marine invertebrates will delay metamorphosis if they do not encounter the appropriate environmental cues (Crisp, 1974; Pechenik, 1990). These cues can be chemicals released by conspecific adults (e.g. Larman et al., 1982; Pechenik and Eyster, 1989; Gebauer et al., 1998), or some other physical or chemical indicator of a good habitat (e.g. Chia and Koss, 1978; Fitt et al., 1990; Pechenik and Cerulli, 1991; Davis, 1994). The ability to delay metamorphosis when the proper cues are absent increases a larva’s or postlarva’s chances of settling into a suitable environment for growth and reproduction (Pechenik, 1990).
shell-less C. compressus megalopae attempting metamorphosis sooner than those with shells was unexpected. Possibly it was due to the fact that the C. compressus in this experiment were housed outside and the fluctuating environmental conditions made it more difficult to survive without a shell. A quick attempt to metamorphose may have been a temporary, but usually unsuccessful, escape from truly dangerous conditions until the postlarva could achieve a more durable form. Survival without a shell in the real world where predators abound is probably even more perilous. Postlarvae with shells that metamorphosed remained subterranean for 1–6 days, similar to the 1–2 days taken by C. variabilis (Harvey, 1992b), but considerably shorter than the 2–4 weeks needed by Birgus latro (Reese and Kinzie, 1968).
Megalopae that successfully metamorphosed generally emigrated to land later and spent more time wandering before burying than those that were unsuccessful. Older megalopae were also less likely to leave their water dishes without their shells and were generally more mobile; after a couple of weeks they could walk or run over sand of all consistencies. Furthermore, I observed a number of laboratory-reared megalopae not used in this experiment and found that older individuals could easily climb pieces of wood placed in their containers. These results suggest that older megalopae do better at each transition point whether they are moving from water to land or from land to the metamorphic burrow.
Many field studies have examined how the postlarval behaviors of marine crustaceans lead to their transport into suitable habitats for settlement (e.g. Tankersley and Forward, 1994; Tankersley et al., 1995). Some of these studies have either noted changes in crustacean megalopal behaviors or considered the possibility that megalopae may change their behaviors as they approach settlement and metamorphosis (e.g. Sulkin and Van Heukelem, 1986; Lipcius et al., 1990; Olmi, 1994; Welch et al., 1997). The results of this study emphasize that the crustacean megalopa does not behave in a fixed manner throughout this single stage, but rather changes its behavior as it maneuvers out of the plankton where it passed through its larval stages and makes the transition to one of the many disparate habitats populated by adult crustaceans.
Acknowledgements
I am deeply appreciative for the intellectual and logistical support provided by John Christy and his lab during my stay in Panama. I also thank Alan Kohn, Richard Strathmann, and an anonymous reviewer for critically reading this manuscript and providing many helpful comments. This work was generously supported by a Predoctor-al Fellowship granted by the Smithsonian TropicPredoctor-al Research Institute.
References
Botero, L., Atema, J., 1982. Behavior and substrate selection during larval settling in the lobster Homarus
Chia, F.S., Koss, R., 1978. Development and metamorphosis of the planktotrophic larvae of Rostanga pulchra (Mollusca: Nudibranchia). Mar. Biol. 46, 109–119.
Cragg, S.M., 1980. Swimming behavior of the larvae of Pecten maximus (L.) (Bivalvia). J. Mar. Biol. Ass. UK 60, 551–564.
Crisp, D.J., 1974. Factors influencing the settlement of marine invertebrate larvae. In: Grant, P.T., Mackie, A.M. (Eds.), Chemoreception in Marine Organisms, Academic Press, London, pp. 177–277.
Davis, M., 1994. Short-term competence in larvae of queen conch Strombus gigas: shifts in behavior, morphology and metamorphic response. Mar. Ecol. Prog. Ser. 104, 101–108.
Fitt, W.K., Coon, S.L., Walch, M., Weiner, R.M., Colwell, R.R., Bonar, D.B., 1990. Settlement behavior and metamorphosis of oyster larvae (Crassostrea gigas) in response to bacterial supernatants. Mar. Biol. 106, 389–394.
Gebauer, P., Walter, I., Anger, K., 1998. Effects of substratum and conspecific adults on the metamorphosis of
Chasmagnathus granulata (Dana) (Decapoda: Grapsidae) megalopae. J. Exp. Mar. Biol. Ecol. 223,
185–198.
Harms, J., 1992. Larval development and delayed metamorphosis in the hermit crab, Clibanarious erythropus (Latreille) (Crustacea, Diogenidae). J. Exp. Mar. Biol. Ecol. 156, 151–160.
Hartnoll, R., 1988. Evolution, systematics, and geographical distribution. In: Burggren, W.W., McMahon, B.R. (Eds.), Biology of the Land Crabs, Cambridge University Press, New York, pp. 6–54.
Harvey, A.W., 1992a. Costs and benefits of delayed metamorphosis in the hermit crab, Clibanarius longitarsus. Am. Zool. 32, 114A.
Harvey, A.W., 1992b. Abbreviated larval development in the Australian terrestrial hermit crab Coenobita
variabilis McCulloch (Anomura: Coenobitidae). J. Crustac. Biol. 12, 196–209.
Harvey, A.W., Colasurdo, E.A., 1993. Effects of shell and food availability on metamorphosis in the hermit crabs, Pagurus hirsutiusculus (Dana) and Pagurus granosimanus (Stimpson). Exp. Mar. Biol. Ecol. 165, 237–249.
Hazlett, B.A., Provenzano, A.J., 1965. Development of behavior in laboratory reared hermit crabs. Bull. Mar. Sci. 15, 616–633.
Jensen, G.C., 1991. Competency, settling behavior, and postsettlement aggregation by porcelain crab megalopae (Anomura: Porcellanidae). J. Exp. Mar. Biol. Ecol. 153, 49–61.
Lang, W.H., Forward, R.B., Miller, D.C., 1979. Behavioral responses of Balanus improvisus nauplii to light intensity and spectrum. Biol. Bull. 157, 166–181.
Larman, V.A., Gabbott, P.A., East, J., 1982. Physico-chemical properties of settlement factor proteins from the barnacle Balanus balanoides. Comp. Biochem. Physiol. 72, 329–338.
Lipcius, R.N., Olmi, E.J., vanMontfrans, J., 1990. Planktonic availability, molt stage and settlement of blue crab postlarvae. Mar. Ecol. Prog. Ser. 58, 235–242.
Marsden, J.R., 1984. Swimming in response to light by larvae of the tropical serpulid Spirobranchus
giganteus. Mar. Biol. 83, 13–16.
Miller, S.E., Hadfield, M.G., 1986. Ontogeny of phototaxis and metamorphic competence in larvae of the nudibranch Phestilla sibogae Bergh (Gastropoda: Opisthobranchia). J. Exp. Mar. Biol. Ecol. 97, 95–112. Nakasone, Y., 1988. Larval stages of Coenobita purpureus Stimpson and C. cavipes Stimpson reared in the laboratory and survival rates and growth factors of three land hermit crab larvae (Crustacea: Anomura). Zool. Sci. 5, 1105–1120.
Olmi, E., 1994. Vertical migration of blue crab Callinectes sapidus megalopae: implications for transport in estuaries. Mar. Ecol. Prog. Ser. 113, 39–54.
Pechenik, J.A., 1990. Delayed metamorphosis by larvae of benthic marine invertebrates: does it occur? Is there a price to pay? Ophelia 32, 63–94.
Pechenik, J.A., Cerulli, T.R., 1991. Influence of delayed metamorphosis on survival, growth, and reproduction of the marine polychaete Capitella sp. I. J. Exp. Mar. Biol. Ecol. 151, 17–27.
Pechenik, J.A., Eyster, L.S., 1989. Influence of delayed metamorphosis on the growth and metabolism of young Crepidula fornicata (Gastropoda) juveniles. Biol. Bull. 176, 14–24.
Reid, D.G., 1996. In: Systematics and Evolution of Littorina, The Ray Society, The Dorset Press, Dorchester, p. 463.
Reese, E.S., 1963. The behavioral mechanism underlying shell selection by hermit crabs. Behavior 21, 78–126.
Reese, E.S., 1968. Shell use: an adaptation for emigration from the ocean by the coconut crab. Science 161, 385–386.
Reese, E.S., Kinzie, R.A., 1968. The larval development of the coconut or robber crab, Birgus latro (L.), in the laboratory (Anomura, Paguroidea). Crustaceana 15, 117–144.
Sulkin, S.D., 1984. Behavioral basis of depth regulation in the larvae of brachyuran crabs. Mar. Ecol. Prog. Ser. 15, 181–205.
Sulkin, S.D., VanHeukelem, W.F., 1986. Variability in the length of the megalopal stage and its consequence to dispersal and recruitment in the portunid crab Callinectes sapidus Rathbun. Bull. Mar. Sci. 39, 269–278. Tankersley, R.A., Forward, R.B., 1994. Endogenous swimming rhythms in estuarine crab megalopae:
implications for flood-tide transport. Mar. Biol. 118, 415–424.
Tankersley, R.A., McKelvey, L.M., Forward, R.B., 1995. Responses of estuarine crab megalopae to pressure, salinity, and light: implications for flood-tide transport. Mar. Biol. 122, 391–400.
Thompson, M.T., 1903. The metamorphosis of the hermit crab. Proc. Boston Soc. Nat. Hist 31, 147–209, pls. 1–10.
Thorson, G., 1950. Reproductive ecology of marine bottom invertbrates. Biol. Rev. 25, 1–45.
Thorson, G., 1964. Light as an ecological factor in the dispersal and settlement of larvae of marine bottom invertebrates. Ophelia 1, 167–208.
Welch, J.M., Rittschof, D., Bullock, T.M., Forward, R.B., 1997. Effects of chemical cues on settlement behavior of blue crab Callinectes sapidus postlarvae. Mar. Ecol. Prog. Ser 154, 143–153.
Wolcott, D.L., DeVries, M.C., 1994. Offshore megalopae of Callinectes sapidus: depth of collection, molt stage and response to estuarine cues. Mar. Ecol. Prog. Ser. 109, 157–163.
Zar, J.H., 1996. In: Biostatistical Analysis, Prentice-Hall, Englewood Cliffs, New Jersey, p. 662, third ed. Zeng, C., Naylor, E., 1996. Occurrence in coastal waters and endogenous tidal swimming rhythms of late