SIFAT FISIOLOGI DAN AGRONOMI PADI RATUN
DENGAN SISTEM SALIBU PADA BUDIDAYA
SYSTEM OF RICE INTENSIFICATION
(SRI)
PINTA OMAS PASARIBU
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Sifat Fisiologi dan Agronomi Padi Ratun dengan Sistem Salibu pada Budidaya System of Rice Intensification (SRI) adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Agustus 2016
RINGKASAN
PINTA OMAS PASARIBU. Sifat Fisiologi dan Agronomi Padi Ratun dengan Sistem Salibu pada Budidaya System of Rice Intensification (SRI). Dibimbing oleh TRIADIATI dan ISWANDI ANAS.
Upaya peningkatan produktivitas padi dapat dilakukan dengan meningkatkan produktivitas lahan melalui pemanfaatan tanaman ratun dengan sistem salibu yang dibudidayakan dengan metode System of Rice Intensification (SRI). Budidaya padi System of Rice Intensification (SRI) merupakan suatu metode dalam pengelolaan tanaman, tanah, air, dan unsur hara untuk meningkatkan pertumbuhan dan perkembangan tanaman padi. Ratun adalah kemampuan tanaman padi dalam menghasilkan anakan baru setelah tanaman pertama dipanen. Keunggulan ratun dapat memberikan tambahan produksi padi per musim tanam, hemat input produksi, tenaga, dan waktu. Sistem salibu merupakan modifikasi pada tanaman ratun yang berkembang di daerah Sumatera Barat yang mampu menghasilkan produksi lebih baik dibandingkan dengan sistem yang umum dilakukan oleh petani (non-salibu).
Penelitian ini bertujuan untuk menganalisis faktor internal yang mendukung fisiologi dan agronomi padi ratun pada sistem salibu yang dibudidayakan dengan metode SRI, sehingga diperoleh informasi dan data mengapa sistem salibu dapat menghasilkan produksi lebih baik dibanding sistem non-salibu. Penelitian ini menggunakan rancangan acak kelompok (RAK) untuk mengkaji metode SRI dan konvensional pada tanaman pertama, sedangkan pada tanaman ratun menggunakan rancangan acak kelompok (RAK) dua faktor yaitu teknik pemotongan dan metode budidaya dengan lima ulangan. Teknik pemotongan terdiri dari sistem salibu dan sistem non-salibu, sedangkan metode budidaya terdiri dari metode SRI dan konvensional. Data pada tanaman pertama dianalisis secara statistik menggunakan Independent t-test dan pada tanaman ratun dianalisis dengan ANOVA pada tingkat kepercayaan α = 5%.Pengamatan pada penelitian ini meliputi pengamatan peubah vegetatif, generatif, dan fisiologi pada padi tanaman pertama dan tanaman ratun.
Peubah agronomi meliputi pertumbuhan vegetatif dan pertumbuhan generatif pada tanaman pertama dan tanaman ratun. Peubah pertumbuhan vegetatif yang diamati yaitu tinggi tanaman, jumlah anakan, jumlah daun, bobot kering tajuk, dan akar umur 105 HSS dan 75 HSP, jumlah anakan produktif per rumpun, serta jumlah anakan produktif per m2. Peubah
pertumbuhan generatif yang diamati yaitu bobot 1000 gabah, bobot gabah kering per rumpun, bobot gabah kering panen per m2, dan bobot gabah kering giling per m2.
kinetin, dan auksin) pada jaringan meristem interkalar pada saat panen tanaman pertama dan tujuh hari setelah panen tanaman pertama.
Hasil penelitian menunjukkan bahwa pertumbuhan vegetatif dan pertumbuhan generatif, laju fotosintesis, serapan hara (N dan P) pada tanaman pertama pada metode SRI lebih tinggi dibandingkan dengan metode konvensional. Hasil gabah pada metode SRI lebih tinggi (sekitar 24.2%) dibandingkan dengan metode konvensional. Pertumbuhan vegetatif dan pertumbuhan generatif, laju fotosintesis, serapan hara (N dan P) pada tanaman ratun lebih tinggi pada sistem salibu dan metode SRI dibandingkan dengan perlakuan lainnya. Hasil gabah pada sistem salibu dan metode SRI lebih tinggi (sekitar 50.3% dari tanaman pertama) dibandingkan dengan perlakuan lainnya. Kemampuan padi dalam menghasilkan ratun dipengaruhi oleh kandungan karbohidrat total dan fitohormon seperti giberelin, kinetin, dan auksin yang terdapat pada meristem interkalar. Kandungan fitohormon yang ditemukan pada sistem salibu dan metode SRI lebih tinggi sehingga produktivitas ratun meningkat dibandingkan dengan perlakuan lainnya, namun kandungan karbohidrat total yang ditemukan lebih sedikit dibandingkan dengan non-salibu.
SUMMARY
PINTA OMAS PASARIBU. Physiological Characteristics and Agronomy of Ratoon Rice Under Salibu System in System of Rice Intensification (SRI). Supervised by TRIADIATI dan ISWANDI ANAS.
Rice productivity can be increased by improving land productivity with a ratoon crop of salibu system cultivated with System of Rice Intensification (SRI) method. Rice cultivation of SRI is a method to increase the rice growth and development by managing the plants, soil, water, and nutrients. Ratooning, the ability of the rice plant to regenerate new tillers after harvested.The beneficial aspects of ratoon provide the increase of rice productivity at once in harvesting and efficient in time, labor, and cost. Local people in West Sumatra commonly re-cut the rice stalk at seven days after first harvesting. This method is called as salibu system which is a modification of ratoon crop and produce a higher yield than non-salibu system (no cutting after first harvesting).
The aim of this study was to analyze internal factors affecting the physiological traits of ratoon rice under the salibu system with the SRI method, in order to obtain information and data why the salibu system can generate the higher production than the non-salibu system. Randomized Block Design (RBD) was used for first crop to compared SRI and conventional methods, while RBD with factorial cutting techniques (salibu and non-salibu system) and cultivation methods was used for ratoon crop. There were five replications for each treatment. All first crops’data were statistically analyzed using Independent t-test and ratoon crop was analyzed by ANOVA with α = 5% level of probability. This research observed the parameters of vegetative, generative, and physiology of first and ratoon crop.
Agronomical parameters observed were the vegetative and generative growth parameters of first and ratoon crop. The vegetative growth parameters measured were plant height, tiller number, leaf number, shoot dry weight at 105 days after sowing (DAS), and at 75 days after harvest (DAH), number of productive tillers per hill, and number of productive tillers per m2. The generative growth parameters observed were
weight of 1000 grains, grain dry weight per hill, grain dry weight at harvest per m2, and grain dry weight (yield) per m2.
Physiological parameters observed were the photosynthetic rate (A) at two phases of growth (the peak stage of vegetative and generative of first and ratoon crop), N, P, and K uptake in leaves at first and ratoon crop, carbohydrate contents in the stubbles’ intercalary meristem tissues of first crop and seven days after frist harvesting, and the phytohormones content (gibberellins, cytokinins, and auxins) in the stubbles’ intercalary meristem tissues of first crop and seven days after first harvesting.
using SRI method (approximately 24.2%) was also higher than that of conventional method. The vegetative and generative parameters, photosynthetic rate, and nutrient (N and P) uptake of ratoon crop using salibu system and SRI method were higher than those of other treatments. The grain yield of ratoon crop using salibu system with SRI method was higher (approximately 50.3 % of first crop production) than that of other treatments. The ability of rice plants to produce a ratoon crop was influence by their carbohydrate content and the phythormones such as gibberellins, cytokinins, and auxins that remain in the intercalary meristem tissues of stubbles after being harvested. The higher rice productivity of salibu system and SRI method was determined by higher phytohormones instead of lower carbohydrate content.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Biologi Tumbuhan
SIFAT FISIOLOGI DAN AGRONOMI PADI RATUN
DENGAN SISTEM SALIBU PADA BUDIDAYA
SYSTEM OF RICE INTENSIFICATION
(SRI)
PINTA OMAS PASARIBU
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Judul Tesis : Sifat Fisiologi dan Agronomi Padi Ratun dengan Sistem Salibu pada Budidaya System of Rice Intensification (SRI) Nama : Pinta Omas Pasaribu
NIM : G353130151
Disetujui oleh Komisi Pembimbing
Dr Dra Triadiati, MSi
Ketua Prof Dr Ir Iswandi Anas, MSc Anggota
Diketahui oleh
Ketua Program Studi Biologi Tumbuhan
Dr Ir Miftahudin, MSi
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
PRAKATA
Puji syukur penulis ucapkan ke hadirat Tuhan Yang Maha Esa yang selalu melimpahkan rahmat dan karuniaNya, sehingga penulis dapat menyelesaikan karya ilmiah ini dengan judul Sifat Fisiologi dan Agronomi Padi Ratun dengan Sistem Salibu Pada Budidaya System of Rice Intensification (SRI). Penelitian ini dilaksanakan pada bulan Juni 2014 sampai Februari 2015, di Sindang Barang Jero dan Laboratorium Fisiologi, Departemen Biologi Fakultas MIPA, Institut Pertanian Bogor.
Terimakasih penulis ucapkan kepada Dr Dra Triadiati, MSi dan Prof Dr Ir Iswandi Anas, MSc selaku pembimbing atas bimbingan, masukan dan arahan yang diberikan. Demikian pula, penulis ucapkan terima kasih kepada penguji luar komisi Dr Ir Sugiyanta, MSi dan Dr Ir Miftahudin, MSi selaku Ketua Program Studi Biologi Tumbuhan IPB, yang telah memberikan masukan pada saat ujian sidang tesis untuk membuat karya ilmiah ini menjadi lebih baik. Kepada DIKTI melalui Beasiswa Unggulan 2013/2014 terima kasih atas kepercayaannya untuk memberikan beasiswa kuliah selama menempuh pendidikan pascasarjana di IPB. Kepada teman-teman Pascasarjana Biologi Tumbuhan IPB 2013 terima kasih atas kebersamaan yang singkat dan sangat indah. Ungkapan terimakasih yang tak terhingga juga penulis ucapkan kepada orang tua tercinta Bapak Sumihar Pasaribu (Alm), Ibu Lamsetiur Sihite, abang dan kakak, serta seluruh keluarga besar, atas segala doa, dukungan, motivasi dan kasih sayangnya selama ini.
Semoga karya ilmiah ini bermanfaat.
Bogor, Agustus 2016
DAFTAR ISI
DAFTAR TABEL xi
DAFTAR GAMBAR xi
DAFTAR LAMPIRAN xi
1 PENDAHULUAN
Latar Belakang 1
Tujuan Penelitian 2
Tahapan Penelitian 2
2 TINJAUAN PUSTAKA
System of Rice Intensification (SRI) 3
Sistem Pertanian Konvensional 4
Ratun 4
Sistem Salibu 5
Fisiologi Padi Ratun 5
3 METODE
Waktu dan Tempat Penelitian 7
Bahan dan Alat Penelitian 7
Rancangan Penelitian 7
Analisis Tanah 7
Pelaksanaan Penelitian di Lapang 7
Pengamatan 9
Analisis Data 10
4 HASIL DAN PEMBAHASAN
Hasil 11
Pembahasan 16
5 SIMPULAN DAN SARAN
Simpulan 21
Saran 21
DAFTAR PUSTAKA 22
LAMPIRAN 27
DAFTAR TABEL
1 Jumlah anakan produktif/rumpun, jumlah anakan produktif/m2,
bobot kering tajuk/rumpun, dan bobot kering akar/rumpun (g) pada
tanaman pertama dan ratun 12
2 Bobot 1000 gabah (g), bobot gabah kering/rumpun (g), bobot gabah kering panen (g m-2), dan bobot gabah kering giling (g m-2) pada
tanaman pertama dan tanaman ratun 13
3 Perbandingan produksi tanaman ratun terhadap tanaman pertama 14 4 Laju fotosintesis pada fase puncak vegetatif dan generatif pada
PAR 2000 µmol CO2 m-2 s-1 pada tanaman pertama dan tanaman
ratun 14
5 Serapan hara N, P, dan K pada daun tanaman pertama dan tanaman
ratun 15
6 Pengaruh teknik pemotongan dan metode budidaya terhadap kandungan karbohidrat total dan fitohormon pada meristem
interkalar padi 16
DAFTAR GAMBAR
1 Tahapan penelitian pada tanaman pertama dan tanaman ratun 2 2 Tinggi tanaman, jumlah daun total/rumpun, dan jumlah
anakan/rumpun pada tanaman pertama dan tanaman ratun 11
LAMPIRAN
1 Tata letak satuan percobaan di lapang 29
2 Hasil analisis kimia dan fisik tanah percobaan di persawahan
1
PENDAHULUAN
Latar Belakang
Salah satu teknologi budidaya dengan inovasi baru pada tanaman padi, yang dikenal dengan System of Rice Intensification (SRI) telah dilaporkan mampu meningkatkan produksi padi (Kabir & Uphoff 2007; Sato & Uphoff 2007; Thakur et al. 2010). SRI merupakan sebuah inovasi yang masih berkembang, namun konsep dan praktiknya telah terbukti mampu meningkatkan produksi padi dan pendapatan petani, sekaligus mengurangi input seperti benih, pupuk sintetik, pestisida, dan air. Metode SRI dapat meningkatkan produksi padi dengan cara mengubah pengelolaan tanaman, tanah, air, dan unsur hara (Ahmed et al. 2015). Metode SRI terdiri dari empat komponen penting, yaitu penanaman bibit muda (8-12 hari), penanaman bibit tunggal (satu lubang tanam untuk satu benih), jarak tanam lebar (25 cm x 25 cm atau lebih), dan kondisi tanah lembap (irigasi berselang). Selain empat komponen tersebut biasanya metode SRI dianjurkan menggunakan pupuk organik untuk memperbaiki sifat tanah agar tanaman padi dapat tumbuh dengan baik. Pengembangan metode SRI ini ada yang diarahkan pada pertanian organik (Barison & Uphoff 2011). Walaupun demikian, metode SRI dengan menggunakan pupuk sintetik atau campuran antara pupuk sintetik dan pupuk organik juga berkembang di tempat lain yang bertujuan untuk mengoptimalkan pertumbuhan tanaman dan produksi.
Metode budidaya padi SRI dikembangkan pertama sekali di Madagaskar pada tahun 1980an dan menghasilkan produksi padi yang lebih tinggi dibandingkan dengan metode konvesional (Laulanie 1993). Praktik penerapan metode SRI ini telah diterapkan oleh lebih dari 54 negara termasuk oleh produsen beras besar di dunia seperti India, Cina, Vietnam, dan Filipina (Katambara et al. 2013). Metode SRI yang diterapkan di Afganistan dapat meningkatkan produksi padi sebesar 66% dibandingkan metode konvensional (Thomas & Ramzi 2010). Demikian juga di Irak, produksi padi juga meningkat sebesar 42% dengan metode SRI (Hameed et al. 2013). Praktik metode SRI yang diterapkan di Indonesia bagian timur (Nusa Tenggara) mampu meningkatkan produksi padi sebesar 78% (Sato et al. 2011). Praktik metode SRI yang telah diterapkan di kota Bogor juga mampu meningkatkan produksi padi berkisar antara 24% sampai 32.6% (Bakrie et al. 2010; Hutabarat 2011; Hidayati 2015).
2
et al. 1985; Oad et al. 2002a,b), sehingga hasil yang diperoleh lebih rendah jika dibandingkan dengan tanaman pertamanya. Akan tetapi, produksi ratun dapat ditingkatkan melalui teknologi yang dikenal dengan sistem salibu. Sistem salibu merupakan teknik pemotongan kembali sisa batang panen padi pada tanaman pertama yang dapat menghasilkan tunas baru. Erdiman (2013) melaporkan bahwa ratun dengan sistem salibu telah dikembangkan di daerah Sumatera Barat dan menghasilkan produksi yang lebih tinggi dibandingkan dengan sistem yang umum digunakan oleh petani (non-salibu). Dengan demikian, pemanfaatan ratun dapat memberikan keuntungan bagi petani dari segi waktu untuk mencapai panen lebih singkat, biaya produksi menjadi lebih murah, tanpa pengolahan tanah, dan tanpa penyemaian (Nair & Rosamma 2002; Sanni et al. 2009; Tari 2011; Faruq et al. 2014). Sistem salibu pada ratun dengan metode budidaya SRI belum banyak dilakukan oleh petani di Indonesia dan karakter fisiologi dan agronomi tanaman ratun dengan sistem salibu pada metode SRI juga masih belum banyak diketahui.
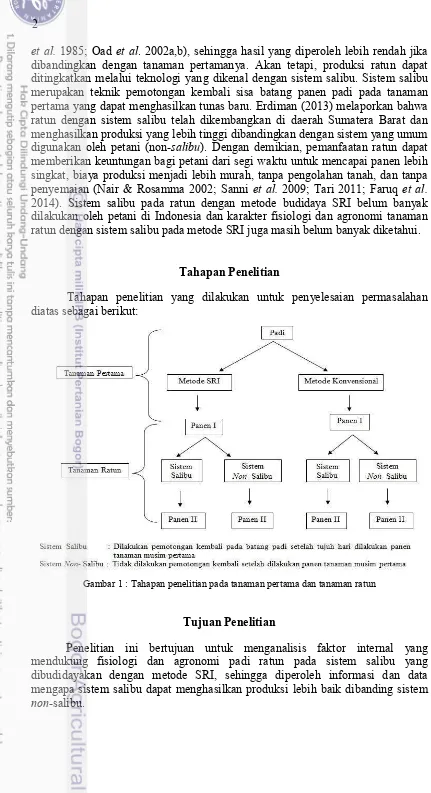
Tahapan Penelitian
Tahapan penelitian yang dilakukan untuk penyelesaian permasalahan diatas sebagai berikut:
Gambar 1 : Tahapan penelitian pada tanaman pertama dan tanaman ratun
Tujuan Penelitian
3
2 TINJAUAN PUSTAKA
System of Rice Intensification (SRI)
Salah satu cara yang dapat digunakan untuk meningkatkan produksi padi adalah dengan menggunakan metode System of Rice Intensification (SRI). Metode SRI merupakan sistem pertanian budidaya padi yang pertama kali dibudidayakan di Madagaskar pada tahun 1980 oleh Fr. Henry de Laulanie. Metode SRI mampu memberikan hasil berlipat dibanding dengan metode konvensional (Laulanie 1993). Penerapan metode SRI telah terbukti berhasil meningkatkan produksi padi sebesar 50% bahkan di beberapa tempat mencapai lebih dari 100% (Setiajie et al. 2008). Metode SRI pertama sekali diperkenalkan di Indonesia oleh Prof Dr N Uphoff dari Cornell University, USA pada tahun 1997 dengan memberikan seminar di Lembaga Penelitian dan Pengembangan Padi di Sukamandi Jawa Barat. Metode SRI ini diterapkan dengan menghasilkan rata-rata hasil panen sebesar 7.61 ton/ha, sedangkan dengan metode konvensional rata-rata hanya 4.27 ton/ha (Uphoff 2011).
Pengaturan air sangat diperhatikan pada metode SRI yaitu kondisi tanah lembap (tidak tergenang) untuk memperbaiki kondisi perakaran tanaman padi. Pada dasarnya tanaman padi tidak membutuhkan keadaan tergenang selama proses pertumbuhannya, karena tanaman padi bukan tanaman air. Penggenangan secara terus menerus dianggap suatu pemborosan pemakaian sumberdaya air. Penggenangan secara terus menerus selain menyebabkan terjadinya peningkatan gas metan juga menyebabkan meningkatnya jaringan aerenkim yang menghambat proses penyerapan air dan unsur hara oleh akar tanaman padi, yang konsekuensinya menghambat pertumbuhan dan perkembangan tanaman (Sumardi 2007).
Prinsip dasar dari budidaya SRI menurut Barison dan Uphoff (2011) sebagai berikut:
1. Umur bibit dipindahkan (transplantasi) ke lahan lebih awal. Pada metode SRI bibit yang digunakan pada saat berumur 8-12 hari setelah semai.
2. Penanaman bibit tunggal yaitu satu lubang untuk satu bibit. Hal ini bertujuan agar tanaman memiliki cukup ruang untuk menyebar dan memperdalam perakarannya.
3. Jarak tanam lebar. Pada metode SRI dianjurkan jarak tanam lebar dengan jarak minimal 25 cm x 25 cm agar akar tanaman mempunyai cukup ruang untuk berkembang sehingga anakan maksimum dapat tercapai.
4. Kondisi tanah tetap lembap, tidak tergenang air (irigasi berselang). Pada metode SRI dianjurkan teknik irigasi berselang agar tercipta kondisi perakaran yang teroksidasi, untuk meningkatkan kesuburan tanah dan mendapatkan akar tanaman yang panjang dan lebat. Pada metode SRI ini kondisi tidak tergenangi dipertahankan selama pertumbuhan vegetatif dan reproduktif.
4
Sistem Pertanian Konvensional
Pertanian secara konvensional menggunakan tanah sawah untuk pertumbuhan padi. Dalam aplikasinya, pertanian konvensional menggunakan prinsip sebagai berikut:
1. Umur bibit dipindahkan (transplantasi) ke lahan sawah saat berumur 25-30 hari.
2. Penanaman bibit sebanyak 5-10 bibit untuk satu lubang tanam, sehingga terjadi persaingan dalam memperoleh cahaya, unsur hara dan ruang tumbuh. 3. Penggunaan jarak tanam yang lebih sempit daripada metode SRI yaitu 20 cm
x 20 cm.
4. Kondisi lahan yang selalu tergenang air sehingga terjadi pembororsan air dan perakaran padi tidak teroksidasi dengan baik.
5. Pemupukan pada metode konvensional sebagian besar menggunakan pupuk sintetik.
Ratun
Peningkatan produksi padi dapat dicapai melalui peningkatan indeks panen dan peningkatan produksi tanaman setiap musim melalui teknologi budidaya atau dengan menanam varietas padi baru yang memiliki sifat unggul. Padi ratun merupakan salah satu alternatif yang dapat dikembangkan oleh petani sebagai tanaman setelah padi pertama dipanen, karena padi ratun lebih hemat sumberdaya dan lebih singkat. Ratun atau singgang (Jawa) atau turiang (Sunda) merupakan tunas yang tumbuh dari tunggul batang yang telah dipanen dan menghasilkan anakan baru hingga dapat dipanen kembali (Krishnamurthy 1988). Budidaya ratun dapat dijadikan sebagai alternatif untuk meningkatkan indeks tanam per tahun, misalnya dari 1 kali menjadi 2 kali atau dari 2 kali menjadi 3 kali tanam dalam satu tahun.
Ratun bukanlah hal yang baru bagi petani, praktek budidaya tanaman padi ratun telah lama dilakukan petani di daerah tropis dan di daerah beriklim sedang. Manfaat ratun telah dipelajari dalam banyak negara seperti India, Jepang, Amerika Serikat, Filipina, Brasil, Kolombia, Swaziland, Thailand, dan Taiwan. Secara genetik, setiap jenis padi memiliki kemampuan menghasilkan ratun yang berbeda-beda. Beberapa varietas padi dilaporkan menghasilkan ratun antara 1.4-3.8 t/ha. Varietas-varietas tersebut adalah IR8 menghasilkan ratun 1.4 t/h di India (Mahadevappa 1988), IR 29 menghasilkan ratun 3.8 t/ha di China, IR28 dan IR 42 masing-masing menghasilkan ratun 2.1 t/ha dan 2.9 t/ha di Filipina (Krishnamurthy 1988).
Keuntungan penerapan ratun adalah cepat, mudah dan murah serta dapat meningkatkan produktivitas padi per unit area dan per unit waktu (Nair & Rosamma 2002). Beberapa keuntungan ratun lainnya antara lain : (a) tidak melakukan pengolahan tanah, penyemaian, dan penanaman kembali, (b) tenaga kerja yang dibutuhkan lebih sedikit, (c) waktu untuk mencapai panen singkat, (d) kebutuhan air irigasi lebih sedikit (Krishnamuthy 1988; Santoso 2014).
5 pertumbuhan ratun tidak baik dan hasilnya sangat rendah. Oleh karena itu, untuk mendapatkan hasil yang baik, perlu diterapkan teknik budidaya yang baik. Baik atau tidaknya padi ratun sangat bergantung kepada pengelolaan tanaman padi pertamanya, misalnya pengolahan tanah, perataan, sistem tanam, dan pengelolaan gulma. Perataan tanah yang baik akan memudahkan pengelolaan air, sehingga tanaman tumbuh seragam (Santoso 2014).
Sistem Salibu
Salah satu sistem budidaya ratun yang sedang dikembangkan saat ini adalah sistem salibu. Sistem salibu pada padi sudah berhasil dikembangkan di daerah Sumatera Barat. Padi salibu merupakan tanaman padi yang tumbuh kembali setelah batang sisa panen dipotong, tunas akan muncul dari ruas yang ada di dalam tanah. Tunas ini akan mengeluarkan akar baru, sehingga suplai hara tidak lagi tergantung pada batang lama, tunas ini bisa membelah atau bertunas lagi seperti pada padi tanaman pertama. Padi salibu berbeda dengan padi ratun. Ratun adalah padi yang tumbuh dari batang sisa panen tanpa dilakukan pemotongan batang kembali, tunas akan muncul pada ruas paling atas, suplai hara tetap dari batang lama. Pertumbuhan tunas setelah dipotong sangat dipengaruhi oleh ketersedian air tanah dan pada saat panen sebaiknya kondisi air tanah dalam keadaan kapasitas lapang.
Sistem salibu akan meningkatkan indeks panen, karena tidak lagi melakukan pengolahan tanah, persemaian, dan penanaman, sehingga rentang waktu produksi lebih pendek. Budidaya ini secara tidak lansung juga dapat menanggulangi keterbatasan varietas unggul, karena pertumbuhan tanaman selanjutnya terjadi secara vegetatif, maka mutu varietas tetap sama dengan tanaman pertama. Hasil uji coba padi salibu pada beberapa daerah di Sumatera Barat cukup bagus antara lain; di Nagari Pauh, Kecamatan Matur, Kabupaten Agam hasil (7,2 t/ha) meningkat 20 % dibanding tanaman pertamanya, di Lima Kaum, Kabupaten Tanah Datar hasil (6,4 t/ha) meningkat (10 -15 %) dibanding tanaman pertama dan didaerah ini sudah ada petani yang mensalibukan padinya lebih 2 kali, berarti 1 kali tanam telah 3 kali panen dan hasilnya tetap stabil. Di Kota Nan Ampek Payakumbuh hasil padi salibu juga sama dengan tanam pertamanya (Erdiman 2013). Budidaya padi salibu meningkatkan indeks panen (IP), karena waktu produksi menjadi lebih pendek, hanya membutuhkan 80-90 % waktu dibandingkan tanaman pertamanya.
Fisiologi Padi Ratun
6
Beberapa karakter agronomi lainnya yang merupakan prasyarat tanaman ratun adalah vigoritas sistem perakaran dan konsentrasi karbohidrat yang tinggi pada batang saat panen tanaman pertama. Kondisi tanaman setelah panen tanaman pertama menunjukkan bahwa kelebihan asimilat yang tersusun dalam bentuk karbohidrat, lipid, dan protein, akan dimanfaatkan tanaman sebagai cadangan makanan dan sebagian akan ditranslokasikan ke daerah pemanfaatan vegetatif. Akar dan batang adalah bagian pemanfaatan hasil fotosintesis selama pertumbuhan ratun. Penyimpanan hasil fotosintesis ke bagian akar dan batang sangat diperlukan, agar batang tanaman padi yang telah dipanen tetap berwarna hijau. Dengan demikan, asimilat yang dihasilkan dapat dimanfaatkan untuk pertumbuhan tunas ratun (Chauhan et al. 1985; Vergara et al. 1988).
Secara morfologi, tanaman ratun pada beberapa varietas padi tumbuh dari semua ruas yang ada pada rumpun, tetapi ditemukan juga tunas yang keluar dari ruas yang tinggi atau yang rendah. Jumlah tunas yang tumbuh ditentukan oleh tinggi pada pemotongan batang tanaman pertama, tetapi kodisinya sangat dipengaruhi oleh sisa asimilat yang terdapat pada sisa batang padi saat panen tanaman pertama yang berfungsi sebagai cadangan pada batang, sehingga dimanfaatkan untuk pertumbuhan tanaman ratun dan tingkat vigorratun (Vergara et al. 1988). Tinggi pemotongan dapat mempengaruhi jumlah anakan, periode pertumbuhan, vigor ratun dan hasil biji (De Datta dan Bernasor 1988).
Umur tanaman ratun umumnya lebih pendek dibandingkan tanaman pertama, hal ini disebabkan ratun memiliki fase pertumbuhan yang berbeda dengan tanaman pertama. Pada tanaman pertama terdapat tiga fase pertumbuhan, yaitu fase vegetatif, reproduktif, dan pemasakan, sedangkan ratun memiliki dua fase, yaitu fase reproduktif dan pemasakan. Kedua fase ini umumnya berlangsung sama pada semua genotipe padi. Fase yang lebih pendek disebabkan munculnya anakan ratun sering diikuti atau bersamaan dengan keluarnya malai atau bunga (Susilawati et al. 2012).
7
3
METODE
Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan dari bulan Juni 2014 sampai dengan Februari 2015 di Desa Sindang Barang Jero, Kecamatan Bogor Barat, Kota Bogor dan Laboratorium Fisiologi, Departemen Biologi Fakultas MIPA, Institut Pertanian Bogor.
Bahan dan Alat Penelitian
Bahan yang digunakan adalah benih padi varietas Ciherang, pupuk Urea (45.7% N), pupuk SP-36 (36.3% P2O5), pupuk KCl (61.1% K2O) dan pupuk
kompos merk CV. Laksmi (0.69% N; 0.34% P2O5; 0.20% K2O). Pengukuran laju
fotosintesis dilakukan dengan menggunakan LICOR 6400XT (Nebraska, USA).
Rancangan Penelitian
Penelitian dirancang menggunakan Rancangan Acak Kelompok (RAK) faktorial dengan perlakuan yang terdiri dari 2 faktor. Faktor pertama adalah teknik pemotongan pada batang padi yang meliputi dua taraf perlakuan yaitu sistem salibu dan sistem non-salibu. Faktor kedua adalah metode budidaya yang meliputi dua taraf perlakuan yaitu metode SRI dan konvensional. Masing-masing perlakuan terdiri atas lima ulangan sehingga terdapat 20 unit percobaan. Petak percobaan berukuran 2 m x 2.5 m (Lampiran 1).
Analisis Tanah
Kualitas tanah yang digunakan dalam penelitian ini disajikan pada Lampiran 2. Analisis tanah dilakukan sebelum penanaman padi dilaksanakan. Sampel tanah diambil secara komposit pada empat titik yang berbeda dari seluruh petakan pada kedalaman 0-20 cm dengan menggunakan bor tanah. Analisis tanah meliputi sifat kimia dan fisik tanah. Sifat kimia tanah meliputi pH-tanah, C-organik, N-total, P, Ca, Mg, K, Na, KTK, Al, H, dan Fe, sedangkan sifat fisik tanah meliputi tekstur tanah. Analisis tanah dilakukan di Balai Besar Sumberdaya Lahan Pertanian Balai Penelitian Tanah, Cimanggu, Bogor.
Pelaksanaan Penelitian di Lapang Tanaman Pertama
8
diratakan untuk mempermudah mengontrol dan mengendalikan air. Benih varietas Ciherang direndam pada air biasa selama 24 jam, ditiriskan, dan diperam dalam keadaan gelap selama 48 jam sampai benih berkecambah. Pada metode SRI, benih disemai pada media tanah dan pupuk organik dalam baki selama 10 hari. Pindah tanam bibit dilakukan pada saat bibit berumur 10 hari setelah semai dengan jarak tanam 25 cm x 25 cm sebanyak 1 bibit per lubang tanam. Pengaturan air diberikan dengan kondisi yang basah dan air diberikan hanya di parit sekeliling petakan. Penggenangan dilakukan hanya pada saat penyiangan dengan ketinggian 1-2 cm. Penyiangan dilakukan pada saat 10, 20, dan 30 hari setelah tanam dengan menggunakan alat penyiang padi (landak) yang berfungsi juga untuk memberikan aerasi di bagian topsoil.
Pada metode konvensional, benih disemai dengan menebarkan benih yang telah berkecambah langsung di lahan sawah sampai berumur 25 hari. Pindah tanam bibit dilakukan pada saat bibit berumur 25 hari setelah semai dengan jarak tanam 20 cm x 20 cm sebanyak 5 bibit per lubang tanam. Pengaturan air diberikan secara tergenang terus-menerus dengan ketinggian 5 cm sampai pemasakan bulir. Penyiangan dilakukan pada saat 10 dan 20 hari setelah tanam dengan cara mencabut gulma menggunakan tangan. Pemupukan budidaya metode SRI dan konvensional persis sama yaitu dengan pemberian 50% dosis pupuk sintetik yang umum digunakan di daerah tersebut (125 kg urea/ha, 100 kg SP 36/ha, dan 50 kg KCl/ha atau setara dengan 250 g urea/petak, 200 g SP 36/petak, dan 100 g KCl/petak) dan 50% pupuk organik (2.5 ton/ha setara dengan 5 kg/petak). Pupuk SP 36, KCl diberikan pada saat penanaman, sedangkan pupuk urea diberikan pada saat tanam dan pada umur 42 hari setelah semai. Pemberian pupuk organik diberikan pada saat tanam di sekitar lubang tanam. Pemberian insektisida hanya dilakukan apabila terdapat gejala serangan hama penyakit. Pada metode SRI dan konvensional air dikeringkan saat pemberian pupuk dan 5 hari sebelum panen. Pemanenan pada tanaman pertama dilakukan dengan memotong batang padi menggunakan pisau arit saat bulir padi telah menguning sekitar 80%.
Tanaman Ratun
Sistem Salibu. Setelah panen lahan segera digenangi air setinggi 5 cm selama 3 hari dan kemudian lahan dikeringkan kembali. Empat hari kemudian, sisa batang panen padi dipotong kembali dengan ketinggian 5 cm di atas permukaan tanah. Pengaturan air pada lahan diberikan setelah tujuh hari kemudian sama seperti halnya dengan tanaman pertama. Penyiangan dan penyulaman dilakukan sepuluh hari setelah pemotongan. Pemberian pupuk diberikan dengan dosis 50% pupuk sintetik (125 kg Urea/ha, 100 kg SP 36/ha, dan 50 kg KCl/ha). Pemberian Pupuk SP 36 dan KCl diberikan pada saat umur 10 hari setelah pemotongan, sedangkan pupuk Urea diberikan pada saat umur 10 dan 30 hari setelah pemotongan. Pemanenan dilakukan dengan memotong batang padi menggunakan pisau arit saat bulir padi telah menguning sekitar 80%.
9 sudah tumbuh merata sama seperti halnya dengan tanaman pertama. Pemberian pupuk Urea, SP 36 dan KCl diberikan secara bersamaan pada hari ke-5 setelah panen tanaman pertama dengan dosis 50% pupuk sintetik (125 kg Urea/ha, 100 kg SP 36/ha, dan 50 kg KCl/ha).
Pengamatan Pengamatan Agronomi Padi
Pertumbuhan Vegetatif. Peubah pertumbuhan vegetatif yang diamati
pada tanaman pertama dan tanaman ratun adalah meliputi tinggi tanaman, jumlah daun, dan jumlah anakan pada umur 38, 53, 68 hari setelah semai dan umur 15, 30, 45 hari setelah panen.
Pertumbuhan Generatif. Peubah pertumbuhan generatif yang diamati adalah jumlah anakan produktif per rumpun, jumlah anakan produktif per m2,
bobot 1000 gabah, bobot gabah kering per rumpun, bobot gabah kering panen per m2, bobot gabah kering giling per m2 dilakukan pada saat panen.
Pengamatan Fisiologi Padi
Laju Fotosintesis. Laju fotosintesis pada tanaman pertama dan tanaman ratun diamati dengan LI-COR Biosciences (Nebraska, USA) yaitu laju asimilasi bersih/tingkat fotosintesis (A). Laju fotosintesis diukur pada PAR 2000 µmol CO2
m-2 s-1. Pengukuran fotosintesis dilakukan sebanyak 2 kali pada saat fase puncak vegetatif dan generatif. Pengukuran fotosintesis menggunakan daun lebar penuh (daun bendera).
Serapan Hara N, P, dan K pada Daun. Serapan hara N, P, dan K pada
daun dianalisis pada panen tanaman pertama dan panen tanaman ratun. Contoh daun padi diambil dari seluruh bagian atas tanaman dan dikeringkan pada oven suhu 80oC sampai bobotnya konstan. Kemudian daun kering dihaluskan menjadi bubuk. Untuk analisis kandungan N pada daun ditentukan dengan metode Kjeldhal, kandungan P dan K dianalisis dengan metode pengabuan basah menggunakan HNO3 dan HClO4, dan diukur menggunakan spektrofotometer UV VIS di Balai Besar Sumberdaya Lahan Pertanian Balai Penelitian Tanah, Cimanggu, Bogor.
Karbohidrat Total pada Meristem Interkalar Padi. Pengukuran
karbohidrat total pada meristem interkalar padi dilakukan pada saat panen tanaman pertama dan tujuh hari setelah panen tanaman pertama. Perhitungan kadar karbohidrat total pada meristem interkalar menggunakan metode Asam fenol suflat/Spektrofotometri (Dubois et al. 1956). 0.1 g batang padi ditimbang kemudian dimasukkan ke dalam erlenmeyer, ditambahkan 200 ml larutan HCl 3% dan didihkan selama 3 jam, setelah dingin dinetralisir dengan NaOH 30% dan ditambahkan sedikit CH3COOH 3% agar suasana larutan sedikit asam. Larutan
10
Analisis Fitohormon (Giberelin, Kinetin, dan Auksin) pada Meristem Interkalar Padi. Analisis giberelin, kinetin, dan auksin pada meristem interkalar dilakukan pada saat panen tanaman pertama dan tujuh hari setelah panen tanaman pertama dengan mengikuti metode yang digunakan Unyayar et al. (1996). Sampel batang yang diambil secara komposit dari setiap ulangan seberat ± 5 gr diekstrak dengan cara digerus, kemudian dilarutkan dengan metanol, kloroform, dan 2N ammonium hidroksida (12:5:3 v/v/v) sebanyak 100 ml. Ekstrak yang diperoleh kemudian disaring. Air destilat sebanyak 22.4 ml ditambahkan ke dalam ekstrak yang diperoleh, selanjutnya ekstrak dimasukkan ke dalam corong pemisah dan didiamkan selama 24 jam. Fase cair diambil sedangkan fase kloroform dibuang. Selanjutnya, pH pada fase air diatur menjadi 2.5 dengan menambahkan HCL 5N, kemudian diekstraksi sebanyak 3 kali dengan menggunakan 15 ml etilasetat. Ekstrak didiamkan beberapa menit agar fase air terpisah dari fase etilasetat. Fase etilasetat diambil dan fase air dilanjutkan untuk perlakuan berikutnya. Fase etilasetat yang diperoleh diharapkan mengandung IAA dan GA3. Fase air berikutnya diatur pH nya menjadi 7 dengan menambahkan 4N NaOH kemudian diekstraksi kembali sebanyak 3 kali menggunakan 15 ml etilasetat. Perlakuan ini akan menghasilkan kinetin bebas. Ekstrak IAA, GA3, dan kinetin yang diperoleh selanjutnya dievaporasi. Ekstrak kering yang diperoleh kemudian ditimbang ± 0.1-0.5 mg dan dilarutkan menggunakan 10 ml metanol kemudian diukur menggunakan spektrofotometri UV-VIS pada panjang gelombang 253 untuk GA3, 280 untuk IAA, dan 269 untuk kinetin.
Analisis Data
11
4
HASIL DAN PEMBAHASAN
Hasil
Pengamatan Agronomi PadiPertumbuhan Vegetatif Tanaman Pertama. Hasil penelitian
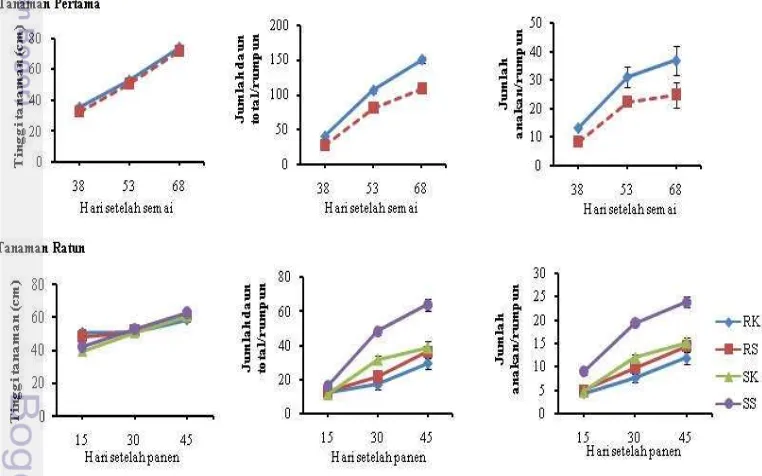
menunjukkan bahwa perlakuan metode budidaya SRI pada tanaman pertama menghasilkan tinggi tanaman, jumlah daun total per rumpun, dan jumlah anakan per rumpun berbeda nyata (P<0.05) dibandingkan metode konvensional pada umur 38 sampai 68 hari setelah semai (HSS). Jumlah anakan pada metode SRI pada umur 38, 53, dan 68 HSS berturut-turut sebesar 13.3, 31.1, dan 37, sedangkan jumlah anakan pada metode konvensional berturut-turut sebesar 8.4, 22.5, dan 24.9. Jumlah daun total per rumpun pada metode SRI pada umur 38, 53, dan 68 HSS berturut-turut sebesar 40.1, 105.8, dan 149.8, sedangkan jumlah daun total per rumpun pada metode konvensional berturut-turut sebesar 26.9, 80.7, dan 108.8 (Gambar 1).
Budidaya padi dengan metode SRI mampu meningkatkan secara nyata bobot kering tajuk dan akar tanaman padi pada umur 105 HSS berturut-turut sebesar 34.3% dan 82.5%. Jumlah anakan produktif per rumpun pada metode SRI juga nyata lebih banyak (P<0.05) dibanding dengan metode konvensional. Jumlah anakan produktif per rumpun pada metode SRI menghasilkan 24.9 anakan, sedangkan pada metode konvensional hanya 14.6 anakan, sehingga penggunaan metode SRI mampu meningkatkan jumlah anakan produktif per rumpun sebesar 71.1%, tetapi jumlah anakan produktif per m2 tidak berbeda nyata (Tabel 1).
12
Pertumbuhan Vegetatif Tanaman Ratun. Perlakuan teknik pemotongan
dan metode budidaya serta interaksinya tidak mempengaruhi tinggi tanaman ratun pada umur 15, 30, dan 45 hari setelah panen (HSP), namun memberikan pengaruh yang nyata terhadap jumlah anakan dan jumlah daun total per rumpun. Jumlah anakan dan jumlah daun total per rumpun nyata lebih banyak (P<0.05) pada perlakuan sistem salibu dengan metode budidaya SRI, semakin banyak jumlah anakan maka semakin banyak pula jumlah daun. Jumlah anakan pada perlakuan sistem salibu dengan metode budidaya SRI pada umur 15, 30, dan 45 hari setelah panen berturut-turut sebesar 9, 19.3, dan 23.8. Jumlah daun total per rumpun pada umur 15, 30, dan 45 hari setelah panen pada perlakuan sistem salibu dengan metode budidaya SRI berturut-turut sebesar 16.6, 48.7, dan 64.1 (Gambar 1).
Perlakuan teknik pemotongan dan metode budidaya masing-masing mempengaruhi jumlah anakan produktif per rumpun dan bobot kering tajuk per rumpun. Jumlah anakan produktif per rumpun dan bobot kering tajuk per rumpun nyata lebih banyak (P<0.05) pada teknik pemotongan sistem salibu dengan metode budidaya SRI. Perlakuan metode budidaya mempengaruhi bobot kering akar per rumpun. Bobot kering akar per rumpun nyata lebih banyak (P<0.05) pada metode budidaya SRI yaitu sebesar 8.3 g dibanding dengan metode konvensional yaitu sebesar 5.3 g. Jumlah anakan produktif per m2 dipengaruhi oleh perlakuan
teknik pemotongan. Jumlah anakan produktif per m2 nyata lebih banyak (P<0.05) pada teknik pemotongan sistem salibu sebesar 216.9 g dari pada sistem non -salibu yaitu sebesar 165.1 g (Tabel 1).
Tabel 1 Jumlah anakan produktif/rumpun, jumlah anakan produktif/m2, bobot
kering tajuk/rumpun (g), dan bobot kering akar/rumpun (g) pada tanaman pertama dan tanaman ratun
Perlakuan produktif/rumpun Jumlah anakan Jumlah anakan produktif/m2 tajuk/rumpun Bobot kering (g)
13
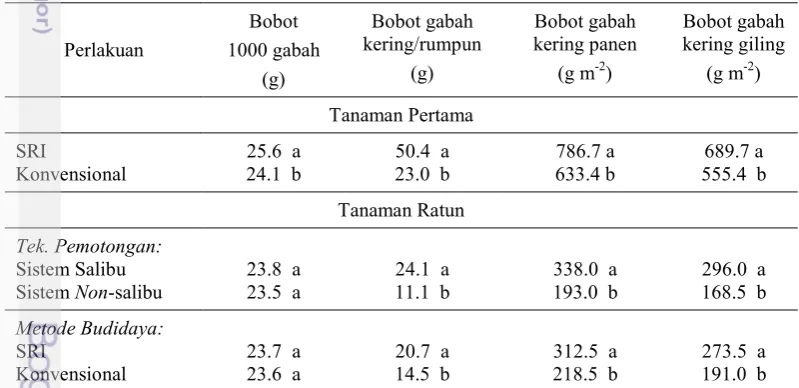
Pertumbuhan Generatif Tanaman Pertama. Budidaya padi dengan
metode SRI secara signifikan menghasilkan bobot gabah kering per rumpun yang lebih tinggi (P<0.05) dibanding dengan metode konvensional. Bobot gabah kering per rumpun pada metode SRI sebesar 50.4 g, sedangkan bobot gabah kering per rumpun pada metode konvensional sebesar 23 g. Oleh karena itu, metode SRI dapat meningkatkan bobot gabah kering per rumpun sebesar 119.3%. Bobot 1000 gabah pada metode SRI juga nyata lebih tinggi (P<0.05) dibanding metode konvensional. Bobot 1000 gabah pada metode SRI sebesar 25.6 g, sedangkan bobot 1000 gabah pada metode konvensional sebesar 24.1 g. Bobot gabah kering panen per m2 pada metode SRI nyata lebih tinggi (P<0.05) yaitu 786.7 g m-2
dibanding metode konvensional yaitu 633.4 g m-2. Begitu pula dengan bobot gabah kering giling per m2 pada metode SRI nyata lebih tinggi (P<0.05) yaitu
689.7 g m-2 dibanding metode konvensional yaitu 555.4 g m-2. Perlakuan metode
SRI dapat meningkatkan bobot gabah kering panen per m2 dan bobot gabah kering
giling per m2 sebesar 24.2% (Tabel 2).
Pertumbuhan Generatif Tanaman Ratun. Perlakuan teknik pemotongan
dan metode budidaya masing-masing serta interaksinya tidak mempengaruhi (P>0.05) bobot 1000 gabah. Perlakuan teknik pemotongan dan metode budidaya masing-masing mempengaruhi bobot gabah kering per rumpun, bobot gabah kering panen per m2, dan bobot gabah kering giling per m2. Bobot gabah kering
per rumpun, bobot gabah kering panen per m2, dan bobot gabah kering giling per m2 nyata lebih banyak (P<0.05) pada teknik pemotongan sistem salibu dengan
14
Perbandingan Produksi Tanaman Ratun terhadap Tanaman
Pertama. Hasil penelitian ini menunjukkan bahwa tanaman ratun yang
diperlakukan dengan teknik pemotongan sistem salibu dan metode budidaya SRI mampu menghasilkan persentase produksi sebesar 50.3% dari tanaman pertama (Tabel 3). Adanya serangan dari burung mempengaruhi nilai persentase produktivitas pada ratun.
Tabel 3 Perbandingan produksi tanaman ratun terhadap tanaman pertama
Produksi Tanaman Pertama Produksi Tanaman Ratun % Peningkatan produksi
Metode Konvensional
Laju Fotosintesis. Laju fotosintesis pada tanaman pertama pada fase puncak vegetatif dan generatif pada metode SRI menunjukkan hasil yang nyata (P<0.05) lebih tinggi dibandingkan dengan metode konvensional. Laju fotosintesis tanaman padi pada metode SRI saat fase puncak vegetatif dan generatif berturut-turut 40.1 dan 29.1 µmol CO2 m-2 s-1, sedangkan laju
fotosintesis tanaman padi pada metode konvensional berturut-turut 36.4 dan 17.6 µmol CO2 m-2 s-1 (Tabel 4 ).
Laju fotosintesis pada tanaman ratun dipengaruhi oleh perlakuan metode budidaya. Laju fotosintesis ratun pada saat fase puncak vegetatif dan generatif pada metode budidaya SRI nyata lebih tinggi (P<0.05) dibanding dengan metode konvensional. Laju fotosintesis ratun pada metode SRI saat fase puncak vegetatif dan generatif berturut-turut 13.7 dan 10.5 µmol CO2 m-2 s-1, sedangkan laju
fotosintesis ratun pada metode konvensional berturut-turut 12.2 dan 9.4 µmol CO2
m-2 s-1 (Tabel 4).
Tabel 4 Laju fotosintesis pada fase puncak vegetatif dan generatif pada PAR 2000 µmol CO2 m-2 s-1 pada tanaman pertama dan tanaman ratun
Tanaman Pertama Laju fotosintesis (µmol CO2 m
-2 s-1)
Fase puncak vegetatif
(70 hari setelah semai) (90 hari setelah semai) Fase generatif
SRI 40.1 a 29.1 a
Konvensional 36.4 b 17.6 b
Tanaman Ratun Laju fotosintesis (µmol CO2 m
-2 s-1)
Fase puncak vegetatif
(35 hari setelah panen) (60 hari setelah panen) Fase generatif
SRI 13.7 a 10.5 a
Konvensional 12.2 b 9.4 b
15
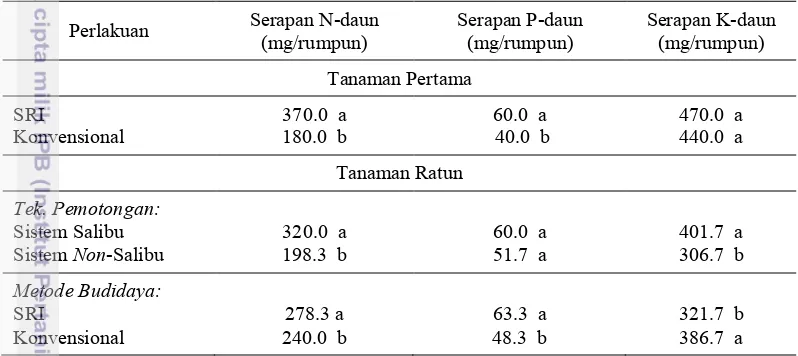
Serapan Hara Tanaman Pertama. Serapan hara N dan P pada daun padi
umur 105 HSS pada metode SRI nyata lebih tinggi (P<0.05) dibanding dengan metode konvensional. Namun, serapan hara K pada metode SRI dan metode konvensional tidak berbeda nyata (P>0.05) (Tabel 5).
Serapan Hara Tanaman Ratun. Perlakuan metode budidaya
mempengaruhi serapan hara P pada daun padi. Serapan hara P pada metode budidaya SRI berbeda nyata dibandingkan dengan konvensional (P<0.05). Perlakuan teknik pemotongan dan metode budidaya masing-masing mempengaruhi serapan hara N dan K pada daun padi ratun (Tabel 5).
Tabel 5 Serapan hara N, P, dan K pada daun tanaman pertama dan tanaman ratun Perlakuan Serapan N-daun (mg/rumpun) Serapan P-daun (mg/rumpun) Serapan K-daun (mg/rumpun)
Tanaman Pertama
masing faktor tidak berbeda nyata menurut uji Duncan (α = 0.05).
16
Tabel 6 Pengaruh teknik pemotongan dan metode budidaya terhadap kandungan
karbohidrat total dan fitohormon pada meristem interkalar padi Perlakuan Karbohidrat total
masing faktor tidak berbeda nyata menurut uji Duncan (α = 0.05).
Pembahasan
Perubahan Fisiologi Padi Tanaman Pertama dengan Metode Budidaya
System of Rice Intensification (SRI)
Metode SRI telah mampu meningkatkan produksi padi dengan cara memperbaiki pertumbuhan tanaman dari fase vegetatif sampai fase generatif. Tinggi tanaman, jumlah daun total per rumpun, dan jumlah anakan per rumpun padi lebih tinggi pada metode SRI dibandingkan dengan metode konvensional (Gambar 2). Hal ini disebabkan umur bibit yang digunakan lebih muda yaitu umur 10 HSS dibandingkan dengan metode konvensional menggunakan bibit umur 25 HSS. Penanaman bibit umur 10 HSS dapat meminimalisasi shock pada tahap pertumbuhan awal padi ketika dipindah tanamkan (Stoop et al. 2002). Penanaman bibit yang lebih muda memungkinkan tanaman lebih mudah beradaptasi dengan lingkungan dan tidak mudah stress. Bibit padi umur 10 HSS juga memiliki perakaran yang belum panjang, sehingga dapat mengurangi guncangan saat pindah tanam dan meningkatkan kemampuan tanaman padi dalam memproduksi batang dan akar selama pertumbuhan vegetatif (Ramli et al. 2012). Selain itu, penanaman dengan satu bibit padi per lubang tanam dan jarak tanam yang lebih lebar pada metode SRI juga dapat mengurangi persaingan dalam menyerap unsur hara, air, dan udara sehingga dapat meningkatkan pertumbuhan individu tanaman padi dibandingkan dengan metode konvensional (Thakur et al. 2010). Selain itu, jarak tanam bibit padi pada metode SRI yang lebih lebar dibanding metode konvensional memberi ruang yang lebih leluasa untuk pertumbuhan perakaran padi. Penggunaan jarak tanam yang lebih lebar memungkinkan cahaya yang ditangkap oleh daun padi akan lebih banyak. Dengan demikian proses fotosintesis dapat berjalan lebih optimum, sehingga menghasilkan pertumbuhan dan produksi tanaman padi yang tinggi (Nurlaili 2011).
17 didukung dengan seiring meningkatnya serapan hara N dan P dalam daun. Serapan hara N dan P dalam daun pada metode SRI lebih tinggi dibandingkan metode konvensional (Tabel 5).
Peningkatan laju fotosintesis dan peningkatan serapan hara pada metode SRI mungkin berperan dalam mengkonversi sebagian besar anakan menjadi anakan produktif (Tabel 1). Peningkatan jumlah anakan produktif dapat meningkatkan bobot 1000 gabah, bobot gabah kering per rumpun, bobot gabah kering panen per m2, dan bobot gabah kering giling per m2 (Tabel 2). Morfologi dan fisiologi tanaman padi pada metode SRI lebih baik dibandingkan dengan metode konvensional sehingga menyebabkan peningkatan hasil gabah (Thakur et al. 2011). Peningkatan pertumbuhan dan perkembangan padi tanaman pertama pada penelitian ini menunjukkan hasil yang sama dengan penelitian yang dilakukan oleh Hidayati (2015) bahwa tanaman padi yang dibudidayakan dengan metode SRI menghasilkan pertumbuhan vegetatif dan generatif lebih tinggi dibandingkan dengan metode konvensional. Pertumbuhan dan perkembangan yang baik pada tanaman pertama akan mempengaruhi keberhasilan pertumbuhan pada ratun (ratun) (Oliver et al. 2014).
Perubahan Fisiologi Tanaman Ratun dengan Sistem Salibu dengan Metode Budidaya System of Rice Intensification (SRI)
Hasil penelitian menunjukkan bahwa jumlah daun, jumlah anakan (Gambar 2), dan jumlah anakan produktif pada ratun lebih sedikit dibandingkan pada tanaman pertama (Tabel 1). Hal ini dikarenakan jumlah karbohidrat tersedia di batang padi yang diperlukan untuk pengisian biji pada ratun lebih sedikit, sehingga hasil panen ratun lebih rendah dari hasil tanaman pertama. Oad et al. (2002a) melaporkan bahwa pertumbuhan dan produktivitas padi pada ratun berbeda dengan tanaman pertama. Secara morfologi, jumlah anakan dan jumlah anakan produktif padi ratun umumnya lebih sedikit dibandingkan dengan tanaman pertama (Vergara et al. 1988; Oad et al. 2002a; Akhgari & Niyaki 2014; Faruq et al. 2014). Penelitian sebelumnya (Chauhan et al. 1985; Sanni et al. 2009; Tari 2011; Liu et al. 2012) juga telah melaporkan bahwa hasil panen ratun lebih sedikit dibandingkan dengan tanaman pertama. Namun, dengan menggunakan sistem salibu dan metode SRI mampu memperbaiki pertumbuhan padi ratun dari fase vegetatif sampai generatif dari pada sistem non-salibu yang umum dilakukan oleh petani.
18
Perlakuan teknik pemotongan dan metode budidaya masing-masing mempengaruhi jumlah anakan produktif per rumpun, bobot kering tajuk per rumpun, sedangkan bobot kering akar per rumpun dipengaruhi oleh metode budidaya. Sistem salibu dan metode budidaya SRI mampu meningkatkan jumlah anakan produktif per rumpun, bobot kering tajuk per rumpun dan bobot kering akar per rumpun dibandingkan dengan perlakuan lainnya (Tabel 1). Hal ini disebabkan banyak tunas yang membentuk anakan baru, sehingga meningkatkan bobot kering tajuk dan bobot kering akar.
Laju fotosintesis pada ratun juga dipengaruhi oleh perlakuan metode budidaya. Laju fotosintesis pada metode SRI lebih tinggi daripada metode konvensional pada fase puncak vegetatif dan generatif (Tabel 4). Daun yang tetap hijau selama pengisian biji dapat menyebabkan fotosintesis tetap tinggi, sehingga translokasi fotosintat ke biji meningkat dan menyebabkan produksi padi juga meningkat (Fu & Lee 2009). Laju fotosintesis meningkat seiring dengan meningkatnya kandungan N di daun. N berperan dalam efisiensi fotosintesis, transportasi asimilasi dan distribusi pada tanaman (Olszewski et al. 2014). Kandungan N yang tinggi dalam jaringan daun memungkinkan klorofil berada dalam jumlah yang banyak dan Rubisco sehingga dapat memicu laju fotosintesis lebih tinggi (Osaki et al. 1995). Perlakuan teknik pemotongan dan metode budidaya masing-masing mempengaruhi serapan hara N dan K pada daun, sedangkan serapan hara P dipengaruhi oleh perlakuan metode budidaya. Serapan hara N oleh daun pada sistem salibu dengan metode SRI lebih banyak dibanding sistem non-salibu dengan metode konvensional, sedangkan serapan hara K oleh daun pada sistem salibu dengan metode konvensional lebih tinggi dibandingkan sistem non-salibu dengan metode SRI. Serapan hara P pada metode SRI lebih tinggi dibandingkan metode konvensional (Tabel 5). P merupakan unsur penting dalam tumbuhan yang berperan dalam proses fotosintesis (Reich et al. 2009), begitu juga dengan K ikut berperan dalam proses fotosintesis (Motaghi & Nejad 2014).
Bobot gabah kering per rumpun, bobot gabah kering panen per m2, dan
bobot gabah kering giling per m2 dipengaruhi oleh perlakuan teknik pemotongan dan metode budidaya. Sistem salibu dan metode SRI mampu meningkatkan bobot gabah kering per rumpun, bobot gabah kering panen per m2, dan bobot gabah
19
Peran Fisiologi Padi dalam Menghasilkan Ratun
Padi merupakan salah satu jenis tanaman dari famili Poaceae yang memiliki keistimewaan mampu untuk tumbuh kembali setelah di panen. Hal yang sering dilakukan oleh petani setelah panen biasanya membiarkan lahannya begitu saja, sehingga nilai produktivitas lahan menurun, sementara mereka dapat memanfaatkan tanaman ratun. Batang padi sisa panen mampu menghasilkan rumpun baru, sehingga dapat melanjutkan pertumbuhan kembali lebih dari satu siklus panen. Kemampuan padi dalam menghasilkan ratun dapat ditentukan oleh sifat genetik dan lingkungan, seperti ketersediaan air, tingkat kesuburan tanah, sinar matahari, suhu, keadaan hama, penyakit tanaman, dan tinggi pemotongan (Mahadevappa 1988). Lebih lanjut Vergara et al. (1988) melaporkan bahwa vigoritas sistem perakaran dan konsentrasi residu karbohidrat yang tinggi pada batang saat panen tanaman pertama merupakan prasyarat pembentukan tanaman ratun.
Hasil penelitian ini menunjukkan bahwa perlakuan teknik pemotongan yang dilakukan pada batang padi tanaman pertama mempengaruhi kandungan karbohidrat total yang terdapat pada jaringan meristem interkalar sisa panen tanaman pertama. Persentase karbohidrat total yang terdapat pada meristem interkalar sisa panen tanaman pertama pada sistem salibu ditemukan lebih sedikit dibandingkan dengan sistem non-salibu (Tabel 6). Karbohidrat merupakan salah satu hasil fotosintesis yang memiliki peranan penting dalam metabolisme. Selain sebagai hasil utama fotosintesis, karbohidrat berperan sebagai substrat dalam proses respirasi yang digunakan untuk membentuk bahan baru tanaman. Karbohidrat yang tersedia cukup pada bagian akar dan batang ditranslokasikan untuk inisiasi tunas sehingga menjadi tanaman ratun. Pemotongan kembali dilakukan untuk merangsang tumbuhnya tunas sehingga dapat meningkatkan jumlah anakan dan jumlah daun.
Pemotongan kembali pada batang padi yang dilakukan pada sistem salibu juga mempengaruhi kandungan fitohormon yang terdapat pada tumbuhan. Fitohormon tumbuhan diduga berperan dalam pembentukan tunas baru. Kurepin et al. (2007) menyatakan bahwa pertumbuhan tunas pada tumbuhan dipengaruhi oleh interaksi lingkungan dengan beberapa fitohormon tumbuhan. Fitohormon dapat memberikan pengaruh yang besar pada berbagai proses fisiologis apabila fitohormon tersebut terdapat pada konsentrasi yang tepat dan pada sel yang tepat (sel target) (Srivastava 2002). Pertumbuhan tunas ratun terbentuk karena adanya kerjasama dari ketiga fitohormon tumbuhan, yaitu peran dari giberelin, kinetin, dan auksin. Hal ini dikarenakan beberapa jenis fitohormon dapat bekerja pada satu respon yang sama (apparent redundancy) (Srivastava 2002).
20
ke fase generatif (Thomas et al. 2005). Begitu juga dengan sitokinin berperan dalam merangsang pembentukan tunas baru. Sitokinin memacu pembelahan sel dan mengkontrol pertumbuhan tunas dan meristem akar (Kyozuka 2007).
Tunas-tunas baru yang membentuk ratun berasal dari jaringan meristem interkalar yang terdapat pada ruas padi. Jaringan meristem interkalar tersusun dari sel-sel yang aktif membelah dan tumbuh (Taiz & Zeiger 2010) dan pembelahan serta pemanjangan sel yang terjadi di jaringan meristem interkalar batang dipengaruhi oleh kandungan giberelin pada tumbuhan (Srivastava 2002). Meristem interkalar merupakan target dari kerja giberelin dalam merangsang pemanjangan internodus batang padi (Taiz & Zeiger 2010).
Selain giberelin dan kinetin, ditemukan juga kandungan auksin pada batang padi sisa panen. Auksin yang ditemukan pada batang padi berperan sebagai pemicu pertumbuhan tunas (Srivastava 2002) dan berperan dalam proses pembelahan dan pemanjangan sel (Campanoni & Nick 2005). Selain itu, auksin juga berperan dalam mengatur dominansi apikal. Transportasi auksin berlangsung secara basipetal, yaitu dari ujung ke basal, sehingga konsentrasi auksin banyak ditemukan didaerah ujung batang (apikal). Dominansi apikal adalah pertumbuhan pada ujung batang (apikal) sehingga menghambat pertumbuhan tunas lateral. Efek dominansi apikal dapat dihambat dengan cara memotong ujung batang sehingga tanaman akan membentuk cabang lateral (Srivastava 2002). Pemotongan yang dilakukan pada sisa batang panen tanaman pertama menyebabkan terjadinya proses dominansi apikal pada batang padi, sehingga memacu munculnya pertumbuhan tunas-tunas baru yang selanjutnya membentuk ratun.
21
5
SIMPULAN DAN SARAN
Simpulan
Pertumbuhan vegetatif dan generatif pada tanaman pertama yang dibudidayakan dengan metode SRI lebih tinggi dibandingkan dengan metode konvensional. Budidaya padi dengan metode SRI menghasilkan berat kering gabah sebesar 24.2% lebih tinggi daripada metode konvensional. Sistem salibu dan metode budidaya SRI pada ratun mampu menghasilkan pertumbuhan vegetatif dan generatif lebih tinggi dibanding dengan perlakuan lainnya. Pertumbuhan yang lebih baik pada ratun dengan sistem salibu yang dibudidayakan dengan metode SRI menghasilkan berat kering gabah sebesar 50.3% dari tanaman pertama. Kemampuan padi dalam menghasilkan ratun dipengaruhi oleh kandungan karbohidrat total dan fitohormon seperti giberelin, kinetin, dan auksin yang terdapat pada meristem interkalar padi. Kandungan fitohormon (giberelin, kinetin, dan auksin) ditemukan lebih banyak pada perlakuan teknik pemotongan sistem salibu dan perlakuan metode budidaya SRI sehingga produktivitas ratun meningkat dibandingkan dengan perlakuan lainnya.
Saran
22
DAFTAR PUSTAKA
Ahmed AR, Dutta BK, Ray DC. 2015. Response of some rice varieties to different crop management practices towards morphological and yield parameters. Int J Sci Res Pub. 5(2):1–6.
Akhgari H, Niyaki SAN. 2014. Effects of first harvest time on total yield and yield components in twice harvesting of rice (Oryza sativa L.) in Rasht, Iran. Int J Biosci. 4(5):210–215.
Bakrie MM, Anas I, Sugiyanta, Indris K. 2010. Aplikasi pupuk anorganik dan organik hayati pada budidaya padi SRI (System of Rice Intensification). J Tanah Lingk. 12:25-32.
Barison J, Uphoff N. 2011. Rice yield and its relation to root growth and nutrient- use efficiency under SRI and conventional cultivation: an evaluation in Madagascar. Paddy Water Environ. 9:65–78.
Campanoni P, Nick P. 2005. Auxin-dependent cell division and cell elongation. Plant Physiol. 137:939-948.
Chauhan JS, Vergara BS, Lopez FSS. 1985. Rice Ratooning. IRRI Res Paper Ser. 102.
Dai X, Mashiguchi K, Chen Q, Kasaharas H, Kamiya Y, Ojha S, Dubois J, Ballou D, Zhao Y. 2013. The biochemical mechanism of auxin biosynthesis by an Arabidopsis YUCCA flavin-containing monooxygenase. J Biol Chem. 288(3):1448-1457
De Datta SK, Bernasor PC. 1988. Agronomic principles and practice of rice ratooning. Di dalam: Smith WH, Kumble V, Cervantes EP, editor. Rice Ratooning, IRRI, Los Banos. Philippines (PH). hlm 163-176.
Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith F. 1956. Colorimetric method for determination of sugars and related substances. An Chem. 28:350-356.
Erdiman. 2013. Teknologi salibu meningkatkan produktivitas lahan dan pendapatan petani [Internet]. [diunduh 2014 23 Juni]. Tersedia pada: http: // sumbar. litbang. deptan. go. id/ ind/ images/ pdf/ padisalibu. pdf.
Faruq G, Taha RM, Prodhan ZH. 2014. Rice ratoon crop: A sustainable rice production system for tropical hill agriculture. Sustainability. 6(9):5785– 5800.
Fu JD, Lee BW. 2009. Changes in photosynthetic characteristics during grain filling of a stay-green rice SNU-SG1 and its F1 hybrids. J Crop Sci Biotech. 11:75-82.
Hameed KA, Mosa AKJ, Jaber FA. 2013. Irrigation water reduction using system of rice intensification compared with conventional cultivation methods in Iraq. Paddy Water Environ. 9:121-127.
Hardjowigeno S. 1995. Ilmu Tanah. Edisi Revisi. Jakarta: Akademika Pressindo Pr.
Harrell DL, Bond JA, Blanche S. 2009. Evaluation of first-crop stubble height on ratoon rice growth and development. Field Crops Res. 114:396-403. Hidayati N. 2015. Fisiologi, anatomi dan sistem perakaran pada budidaya padi
23 Hutabarat TR. 2011. Populasi mikrob tanah emisi metan dan produksi padi
dengan kombinasi pemupukan pada budidaya padi SRI (System of Rice Intensification) [tesis]. Bogor (ID) : Institut Pertanian Bogor.
Ichii M. 1983. The effect of water management on ratoon ability of rice plants. Tech Bull Fac Agri Kagawa Univ. 34(2):123–128.
Kabir H, Uphoff N. 2007. Results of disseminating the system of rice intensification with farmer field school methods in northern Myanmar. Expl Agric. 43:463-476.
Katambara Z, Kahimba FC, Mahoo HF, Mbungu WB, Mhenga F, Reuben P, Maugo M, Nyarubamba A. 2013. Adopting the system of rice intensification (SRI) in Tanzania: A review. Agric Sci. 4:369-375.
Krishnamurthy K. 1988. Rice ratooning as an alternative to double cropping in Asia. Di dalam: Smith WH, Kumble V, Cervantes EP, editor. Rice Ratooning, IRRI, Los Banos. Philippines (PH). hlm 3-15.
Kurepin LV, Emery RJN, Pharis RP, Reid DM. 2007. The interaction of light quality and irradiance with gibberellins, cytokinins and auxin in regulating growth of Helianthus annus hypocotyls. Plant Cell Environ. 30:147-155. Kyozuka J. 2007. Control of shoot and root meristem function by cytokinin.
Curr Opin Plant Biol. 10:442-446.
Laulanie HD. 1993. System of rice intensification. Tropicultura. 11:1-19.
Liu K, Qin J, Zhang B, Zhao Y. 2012. Physiological traits, yields and N translocation of ratoon rice in response to different cultivations and planting periods. Afr J Agric Res. 7(16):2539-2545.
Mahadevappa M. 1988. Rice ratooning practices in India. Di dalam: Smith WH, Kumble V, Cervantes EP, editor. Rice Ratooning, IRRI, Los Banos. Philippines (PH). hlm 69-78.
Mahmoody M, Noori M. 2014. Effect of gibberellic acid on growth and development plants and its relationship with abiotic stress. Int J Farm Alli Sci. 3(6):717-721.
Mobasser HR, Rostaei Kh, Qasempour AM. 2012. The effect of first crop harvesting time on rice-ratoon and berseem clover (Trifolium alexanderinum L .) yield as intercropping system. Int J Agron Plant Prod. 3(10):414–418.
Motaghi S, Nejad TS. 2014. The effect of diffrent levels of humic acid and potassium fertilizer on physiological indices of growth. Int J Biosci. 5(2):99-105.
Nair AS, Rosamma CA. 2002. Character association in ratoon crop of rice (Oryza sativa L.). J Tropic Agric. 40:1-3.
Nurlaili. 2011. Optimalisasi cahaya matahari pada pertanaman padi (Oryza sativa L) System of Rice Intensification (SRI) melalui pendekatan pengaturan jarak tanam. Agronobis. 3(5):22-27.
Oad F, Cruz PS, Memon N, Oad NL, Hassan ZU. 2002a. Rice ratooning management. Pak J Applied Sci. 2(1):29-35.
24
Olivier K, Cherif M, Kone B, Emmanuel D, Firmin KK. 2014. Growth, yields, and ratooning performance of lowland rice nerica L14 as affected by different fertilizers. Ind J Sci Res Tech. 2(2):18-24.
Olszewski J, Makowska M, Pszczolkowska A, Okorski A, Bieniaszewzki T. 2014. The effect of nitrogen fertilization on flag leaf and ear photosynthesis and grain yield of spring wheat. Plant Soil Environ. 60(12):531-536.
Osaki M, Iyoda M, Tadano T. 1995. Effect of N application and sink manipulation on the contents of ribulose - 1,5 bisphosphate carboxylase / oxygenase, phosphoenolpyruvate carboxylase, and chlorophyll in leaves of maize during the maturation phase. Soil Sci Plant Nutr. 41:295-303.
Ramli, Kaharuddin, Samaria. 2012. Pengaruh umur transplanting terhadap pertumbuhan vegetatif berbagai varietas padi. J Agrisistem. 8(1):1-10. Reich PB, Oleksyn J, Wright IJ. 2009. Leaf phosphorus influences the
photosynthesis-nitrogen relation: a cross-biome analysis of 314 species. Oecologia. 160:207-212.
Sanni KA, Ojo DK, Adebisi MA, Somado EA, Ariyo OJ, Sie M, Akintayo I, Tia DD. 2009. Ratooning potential of interspecific NERICA rice varietas. Int J Bot. 5(1):112-115.
Santoso MB. 2014. Budidaya padi ratun [Internet]. Balai besar pelatihan pertanian binuang [diunduh 2016 17 Maret]. Tersedia pada: http://bbppbinuang.info/get.php?file=52212budidayapadiratun.
Sato S, Uphoff N. 2007. A review of on-farm evaluations of system of rice intensification methods in Eastern Indonesia. CAB Reviews: Perspectives Agric Veterinary Sci Nutr Natur Res. 2:54-65.
Sato S, Yamaji E, Kuroda T. 2011. Strategies and engineering adaptions to disseminate SRI methods in large-scale irrigation systems in eastern Indonesia. Paddy Water Environ. 9:79-88.
Setiajie, Wardana IP, Sumedi I. 2008. Gagasan dan Implementasi System of Rice Intensification (SRI) dalam Kegiatan Budidaya Padi Ekologis (BPE). J Analisis Kebijakan Pertanian. 6:75-99.
Srivastava, LM. 2002. Plant Growth and Development: Hormones and Environment. USA: Elsevier Science.
Stoop WA, Uphoff N, Kassam A. 2002. A review of agricultural research issues raised by the system of rice intensification (SRI) from Madagascar: opportunities for improving farming systems for resource-poor farmers. Agric Syst. 71:249-274.
Sumardi. 2007. Respon padi sawah pada teknik budidaya secara aerobik dan pemberian bahan organik. J Akta Agro. 7:65-70.
Susilawati, Purwoko BS, Aswidinnoor H, Santosa E. 2012. Tingkat produksi ratun berdasarkan tinggi pemotongan batang padi sawah pada saat panen. J Agron Indonesia.40 (1):1-7.
Suwandi, Ammar M, Irsan C. 2012. Aplikasi ekstrak kompos meningkatkan hasil dan menekan penyakit padi sistem ratun di sawah pasang surut Kabupaten Banyuasin. Lahan Suboptimal. 1(2):116–122.
Taiz L, Zeiger E. 2010. Plant Physiology. California : The Benjamin/Cumming. Takahashi H. 2013. Auxin biology in roots. Plant Root (JSRR). 7:49-64.
25 Thakur AK, Rath S, Roychowdhury S, Uphoff N. 2010. Comparative
performance of rice with system of rice intensification (SRI) and conventional management using different plant spacings. Expl Agric. 196:146–159.
Thakur AK, Rath S, Patil DU, Kumar A. 2011. Effects on rice plant morphology and physiology of water and associated management practices of the system of rice intensification and their implications for crop performance. Paddy Water Environ. 9:13–24.
Thomas SG, Rieu I, Steber CM. 2005. Gibbrellin metabolism and signaling. Vitam Horm. 72:289-338.
Thomas V, Ramzi AM. 2010. SRI contributions of rice production dealing with water management constraints in northeastern Afganistan. Paddy Water Environ. 9:101-109.
Unyayar S, Topcuoglu SF, Unyayar A. 1996. A modified method for extraction and identification of indole-3- acetic acid (IAA), gibberellic acid (GA3), abcisic acid (ABA) and zeatin produced by Phanerochaetechrysosporium ME446. Bulg J PlantPysiol.22 (3-4):105-110.
Uphoff N. 2011. A review of spread in Asia [Internet]. [diunduh 2015 Feb 10]. Tersedia pada: http://www.springerlink.com/.
27
29 Lampiran 1 Tata letak satuan percobaan di lapang
Petak percobaan 2 m x 2.5 m
Saluran air masuk
Saluran air keluar
U
Keterangan : M0 : Konvensional M1 : SRI
T0 : Sistem Non-Salibu T1 : Sistem Salibu
M0T0
(1) M0T1 (1)
M1T0
(1) M1T1 (1)
M0T0
(2) M0T1 (2)
M1T0
(2) M1T1 (2)
M0T0
(3) M0T1 (3)
M1T0
(3) M1T1 (3)
M0T0
(4) M0T1 (4)
M1T0
(4) M1T1 (4)
M0T0
(5) M0T1 (5)
M1T0
(5) M1T1 (5)
Saluran air masuk
30
Lampiran 2 Hasil analisis sifat kimia dan fisik tanah percobaan di persawahan Sindang Barang Jero Bogor sebelum tanam
Sifat tanah Satuan Kriteria
pH 1:5 H20 5.0 (rendah)
Walkley & Black C-org (%) 1.88 (rendah)
Kjeldhal N-Total (%) 0.19 (rendah)
Bray I P (ppm) 7.4 (sangat rendah)
Ca 8.98 (sedang)
NH4-Asetat Mg (me/100g) 1.29 (sedang)
K 0.07 (sangat rendah)
Na 0.64 (sedang)
KTK 21.42 (sedang)
NKCl Al (me/100g) 0.05
H 0.07
Fe Tidak terukur
Tekstur Pasir (%) 23
Debu 36
Liat 41