EKOLOGI, MORFOLOGI DAN VARIABILITAS GENETIK KODOK (GENUS RANA) DI WILAYAH TIMOR BARAT
NUSA TENGGARA TIMUR
O l e h
ALFRED 0. M.
DIMA
PROGRAM PASCASARJANA INSTITUT PERTANIAN BOGOR
Motto :
Orang-orang yang m w h r d q a n mencucu~an air mata,
akan menuai d q a n 6ersora&sorai Orang yang 6q'al;ln ma& d q a n menuqis
sam Mmenuhr Geniti,
parti pulhng d q a n sora&sorai
Alfred O.M. Dima. Ekologi, Morfblogi dan Variabilitas Genetik Kodok (Genus
Rana) Di Wilayah Timor Barat Nusa Tenggara Timur (Dibawah bimbingan Prof.
Dr. Nawangsari Sugiri, sebagai keiua dan Dr. Ir. M. F. Rahrrdjo, DEA sebagai anggota).
Fragmentasi habitat dan pencemaran lingkungan perairan merupakan
faktor lingkungan yang diduga dapat mempengaruhi kelangsungan hidup genus
R a m di wilayah Timor-Barat Nusa Tenggara Timur. Mengingat betapa
pentingnya keberadaannya terhadap ekosistem terutama sebagai predator yang
mengontrol secara biologis serta potensial untuk dijadikan sebagai salah satu
komoditas, maka informasi yang lengkap secara simultan melalui penelitian yang
meliputi aspek ekologi, morfologi dan keragaman genetik pada berbagai daerah
menj adi sangat bermanfaat.
Tujuan penelitian ini antara lain untuk mengetahui keadaan habitat
alami, keragaman morfologi, dimorfisme seksual dan variabilitas genetik genus
Rana di wilayah Timor Barat Nusa Tenggara Timur.
Dari tiga wilayah yang dijadikan lokasi pengambilan sampel kodok
yaitu Kupang, Timor Tengah Selatan (TTS) dan Belu berhasil diidentifikasi s pesies Rana cancrivora, Rana erythraea dan Rana limnocharis. Kajian aspek
ekologi yang meliputi analisis kualitas air, tumbuhan dan iklim habitat kodok
genus Rana pada areal persawahan dan sungai dengan vegetasi yang didominasi
oleh padi dan rumput liar menunjukkan bahwa derajat keasaman (pH) berkisar
antara 7-8; oksigen terlarut berkisar antara 3,12-5,40 ppm; kandungan
karbondioksida bebas antara 4,29-15,02 ppm; suhu air antara 1 8 - 2 0 ' ~ dan jumlah
Total sampel 251 individu yang berasal dari tiga lokasi di wilayah
Timor Barat yaitu Kupang, TTS dan Belu digunakan untuk analisis morfometri,
sedangkan 20 individu digunakan untuk analisis kariotip dan pola pita protein.
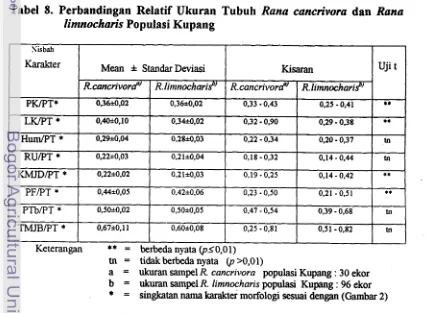
Hasil analisis perbandingan relatif delapan karakter ukuran tubuh
(morfometri) antara R cancrivora dengan R. limnocharis pada populasi dari Kupang menunjukkan bahwa nilai rataan empat karakter morfometri Rana
cancrivora lebih besar dibandingkan dengan Rana limnocharis. Analisis uji t
antara kedua spesies tersebut menunjukkan perbedaan sangat nyata (p<0,01).
Dengan demikian keempat ciri yaitu pajang kepala (PK), lebar kepala (LK),
panjang karpal-metakarpal jari depan (KMJD) dan panjang femur (PF) secara relatif terhadap panjang tubuh (PT) dapat dijadikan sebagai tambahan ciri
pembeda untuk identifikasi kedua spesies tersebut.
Analisis perbandingan morfometri berdasarkan letak geografis juga
dilakukan terhadap Rana cancrivora yang berasal dari tiga daerah (Kupang, TTS
dan Belu) menunjukkan bahwa ketiga karakter morfologi yaitu panjang tubuh,
panjang humerus dan panjang karpal metakarpus jari depan mempunyai
kecenderungan semakin panjang secara berturut-turut dari Belu, TTS dan Kupang.
Pada sisi lain dimorfisme seksual Rana cancrivora di wilayah Timor Barat
menunjukkan adanya variasi ukuran linier tubuh kodok sawah betina dewasa lebih
besar dari pada jantan dewasa. Analisis statistik terhadap 15 karakter
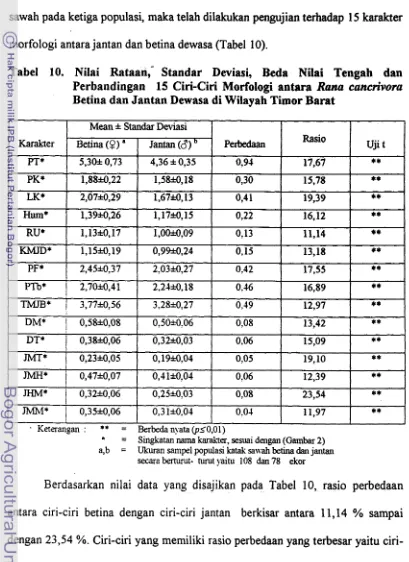
menunjukkan perbedaan sangat nyata (p< 0.01). Rasio perbedaan nilai rataan 15
karakter morfologi menujukkan bahwa nilai rataan karakter jarak hidung mulut
Rana limnocharis dan Rana cancrivora memiliki 13 pasang kromosom
(2n = 26), 9 pasang kromosom memiliki susunan yang sama dengan tipe kromosom submetasentrik, sedangkan 4 pasang kromosom lainnya memiliki
susunan yang berbeda antara kedua spesies tersebut.
Hasil analisis protein serum darah menunjukkan bahwa kedua spesies
mempunyai jumlah pita protein yang sama yaitu 1 pita protein, dengan jarak
EKOLOGI, MORFOLOGI DAN VARIABILITAS GENETlK KODOK (GENUS RANA) DI WILAYAH TIMOR BARAT
NUSA
TENGGARA TIMURO l e h
ALFRED 0 . M. D I M . 99422lBIOLOGI
Tesis sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada Program Pascasarjana, Institut Pertanian Bogor bidang keahlian Biologi/ Zoologi
PROGRAM PASCASARJANA INSTITUT PERTANIAN BOGOR
Judul Tesis : EKOLOGI, MORFOLOGI DAN VARIABILITAS
GENETIK KODOK (GENUS RANA) DI WILAYAH
TIMOR BARAT NUSA TENGGARA TIMUR
Nama Mahasiswa : Alfred 0. M. Dima
Nomor Pokok : 99422
Menyetujui;
Komisi Pembimbing,
/
Dr. Ir. M. F. Rahard-io. DEAAnggota
Mengetahui,
Ketua Program Studi Biologi,
n
1
-
Dr. Ir. Dedy Duryadi Solihin. DEA
Tanggal lulus : 12 April 2002
Penulis lahir pada tahun 1970 di Batuplat, Nusa Tenggara Timur
(NTT), dari Mama Rahel. A. Dima-Here dan Bapa J. Dima. Penulis
menyelesaikan pendidikan tingkat dasar pada SD Inpres Bakunase pada tahun
1982, Sekolah Menengah Tingkat Pertama (SMP) Negeri 3, Kupang tahun 1985
dan Sekolah Menengah Tingkat Atas (SMA) Negeri 2, Kupang tahun 1988..
Penulis memperoleh gelar sarjana peternakan pada Fakultas Peternakan
Universitas Nusa Cendana pada tahun 1993. Sejak bulan Desember 2000 menjadi
staf pengajar pada jurusan Produksi Ternak, Fakultas Peternakan Universitas Nusa
Cendana Kupang NTT. Pada tahun1999 penulis mendapat Beasiswa Magister
Dalam Negeri bagi dosen baru melalui proyek DUE DIKTI Jakarta dengan
sumber dana dari Bank Dunia dan mengikuti pendidikan Magister (S2) pada
Program Studi Biologi Program Pascasarjana Institut Pertanian Bogor (IPB).
Akhirnya pada tanggal 12 April 2002 penulis dapat menyelesaikan jenjang
KATA PENGANTAR
Salah satu syarat yang diperlukan untuk menyelesaikan jenjang pendidikan
strata 2 (Magister) yaitu melakukan penelitian ilmiah dan dipertanggung-
jawabkan secara ilmiah dalam bentuk tulisan ilmiah secara teratur dan sistimatis.
Sehubungan dengan itu, maka telah dilakukan sebuah penelitian yang berjudul
"Ekologi, Morfologi dan Variabilitas Genetik Kodok (Genus Rana) Di Wilayah Timor Barat Nusa Tenggara Timur".
Pemilihan topik ini didasarkan pada pertimbangan bahwa informasi
tentang keberadaan genus Rana di wilayah Timor Barat belum banyak terungkap
(hanya terbatas pada informasi penyebaran genus Rana di seluruh wilayah Propinsi Nusa Tenggara Timur). Semoga hasil penelitian ini dapat dijadikan
sebagai sumbangan pemikiran ilmiah bagi semua pihak yang terkait dengan upaya
pemanfaatan maupun pelestarian kodok genus Rana di wilayah Timor Barat.
Penulis menyadari bahwa dalam menyampaikan tulisan ilmiah ini masih
terdapat banyak kekurangan dan keterbatasan, baik secara teknis maupun kajian
ilmiah. Sehubungan dengan ha1 tersebut, maka dengan kerendahan hati penulis
sangat mengharapkan saran dan kritikan membangun dari semua pihak.
Bogor, 12 April 2002
Penulis,
UCAPAN TERIMA KASIH
Sebagai hambaNya, pertama-tama penulis menyampaikan syukur kepada
Tuhan Yesus Kristus yang menjadi sumber kehidupan dan keselamatanku serta
telah mengerjakan segala perkara yang baik dalam hidupku.
Melalui kesempatan ini ucapan terima kasih dan penghargaan yang
sebesarnya kepada :
1. Ibu Prof. Dr. Nawangsari Sugiri, selaku Ketua Komisi Pembimbing dan
Bapak Dr. Ir. M. F. Rahardjo, DEA selaku Anggota Pembimbing yang
telah membimbing dan mengarahkan hingga selesainya tulisan ini
2. Pimpinan Proyek DUE Magister Dalam Negeri, Dirjen Pendidikan Tinggi-
Departemen Pendidikan Nasional Jakarta beserta staf yang telah
memberikan bantuan moril maupun materil selama penulis menjalani
pendidikan di IPB
3. Kepala Laboratorium Biologi Hewan P S M IPB beserta seluruh staf yang telah membantu dan menyediakan berbagai fasilitas dalam penelitian
4. Ibu Tini Wahyuni yang telah banyak membantu dan menyumbangkan
pikiran, gagasan dan tenaga selama bekerja di laboratorium
5. Karyawan perpustakaan di lingkungan IPB dan Museum Zoologi LIP1
Cibinong yang telah memberikan pelayanan yang memuaskan kepada
penulis dalam menggunakan fasilitas perpustakaan dan laboratorium
6. Bung Rodialek Polo dan istri, Bung Vino, Bung Roy, Bung Tian, Efrit,
Om Jhon Malelak dan istri, BP dan anggota Gainanusratim, rekan-rekan
pemuda Jemaat Rehoboth Kupang dan Zebaoth Bogor untuk kasih, doa
7. Om Daddy dan Kak Yumi dengan pikiran, waktu dan tenaga yang
senantiasa bersama dalam susah dan senang untuk menolong penulis
8. Rekan-rekan seangkatan Jurusan Zoologi (Dian, Jusmaldi, Emi, Ibu Wida)
untuk kebersamaan selama kuliah hingga penelitian ini rampung
9. Ade Sandra Salamoni tersayang, yang walaupun dalam masa pendidikan
di Jakarta sempat untuk memberikan perhatian, dorongan dan kasihnya.
Semoga jalinan kasih ini tetap terbina dan senantiasa diberkati Tuhan
Yesus.
10. Bapa dan Mama, Kak Djemmie, Kak Linda, Brenda dan Ari, Kak Inna,
Kak Ongqy, Theos, Yati, Arna, Ikke, Marthen, Diana, Kak Djibrael, dan
Keluarga Besar Dima, Keluarga Salamoni, Kak Min, Kak Loudwiq dan
Om Edu Mauboi atas kasih, dukungan dan doanya kepada penulis selama
pendidikan hingga penyelesaian tugas akhir (You are best families in my
life)
11. Kepada semua pihak yang tidak dapat penulis sebutkan satu per satu atas
DAFTAR IS1
Halaman
...
...RINGKASAN 111
...
RIWAYAT HIDUP ... v ~ i i KATA PENGANTAR ... ix
UC APAN TERIMA KASM ... x
.
.
DAFTAR IS1 ... xi1
...
DAFTAR TABEL xiv
DAFTAR GAMBAR ... xv DAFTAR LAMPIRAN ... xvi I . PENDAHULUAN ... 1
A . Latar Belakang ... 1 B . Perumusan Masalah
...
- 3 C . Tujuan dan Manfaat Penelitian...
511 . TINJAUAN PUSTAKA
...
6 A . Klasifikasi dan Ciri-Ciri Morfologi...
6 B . Habitat dan Penyebaran ... 10 C . Keragaman Kariotip Genus Rana ... 14 D . Polimorfisme Protein ... 16 1II.METODE PENELITIAN ... 21 A . Tempat dan Waktu Penelitian ... 21 B . Bahan dan Alat Penelitian ... 21 C . Metode Kerja ... -221 . Pengamatan Pendahuluan ... 22 2 . Pengambilan Contoh ... 22 3 . Kualitas Air ... 23 4 . Morfologi ... -24 5 . Kariotip ... -26 6 . Pemeriksaan Pola Protein ... -28 D . Analisis Data ... 30
1 . Kualitas Air ... 30 ..
2 . Morfometri ... 31 3 . Variabilitas Genetik ... i. ... 31
IV . HASIL DAN PEMBAHASAN ... 33 A . Habitat ... 33 1 . Habitat Kodok Genus Rana di Kupang ... 33
...
1 . 1 Kualitas Air di Kupang 35
2 . Habitat Kodok Genus Rana di Timor Tengah Selatan
...
36 2.1 Kualitas Air diTTS
... 38...
3
.
Habitat Kodok Genus Rana di Belu 393.1 Kualitas Air di Belu
...
40 B . Iklim Habitat Kodok Genus Rana di Kupang,TTS
dan Belu...
42...
C . Morfologi Genus Ram -44
...
.
1 Ram cancrivora -44
...
2 . Rana limnocharis 46
...
3 Morfometri Genus Rana 48
3.1 Morfometri antara R limnocharis dengan R cancrivora
...
48 3.2 Morfometri Rana cancrivora secara Geografis...
50 3.3 Dimorfisme Seksual Rana cancrivora di Timor Barat...
51... .
D Kromosom Genus Rana 54
...
.
1 Rana cancrivora 54
...
2 . Ram limnocharis 56
... .
E Analisis Pola Protein 59
...
V. KE S IMPULAN DAN SARAN 63
A . Kesimpulan ... 63 ...
B . Saran -64
...
DAFTAR PUSTAKA -65
...
DAFTAR
TABEL
Nomor Teks Halaman
1 . Komposisi Genotip dari Sembilan Lokus Polimorf Kodok Sawah
di Jawa Tengah ... 19
2 . Komposisi Genotip dari Lima Lokus Polimorf Kodok Sawah
...
di Jawa Barat 19
3 . Tingkat Variabilitas Tiga Spesies Genus Bipes
...
20 4 . Sifat Fisik Kimiawi Air yang Diukur, Metode dan Alat Ukur...
30 5 . Beberapa Sifat Fisik Kimiawi Perairan di Wilayah Timor Barat...
33 6 . Jumlah Curah Hujan Habitat Genus Rana di Wilayah Timor Barat...
42...
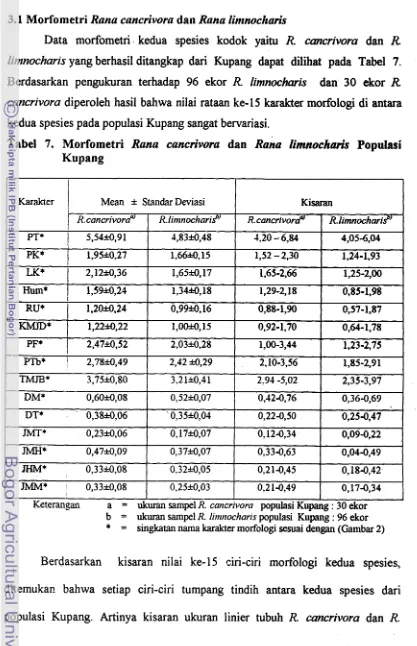
7
.
Morfometri Rana cancrivora dan Rana lintnocharis 48 8 . Perbandingan Relatif Ukuran Tubuh Rana cancriwra danRam linznocharis Populasi Kupang
...
49 9 . Morfometri Rana cancrivora pada Populasi Kupang. TTS dan Belu...
5110 . Nilai Rataan, Standar Deviasi. Beda Nilai Tengah dan Perbandingan 15 Karakter Morfologi antara Rana cancrivora Betina dan Jantan Dewasa Di Wilayah Timor Barat ... -52 1 1 . Panjang Relatif, Nilai Numerik Letak Sentromer dan Tipe Kromosom
Rana cancrivora dalam Metafase. n = 7 ... 55
12 . Panjang Relatif, Nilai Numerik Letak Sentromer dan Tipe Kromosom
Raua lintnocharis dalam Metafase. n = 7 ... 57
.
...
13 Jarak Migrasi (em) Pita Protein Standar dan Protein Sampel Genus Raw 60 ... 14 . Nilai Rf Protein Standar dan Protein Sampel Genus Rana 61
... .
15 Nilai Berat Molekul Protein Standar dan Protein Sampel Genus Rana 62
DAFTAR GAMBAR
Nomor Teks Halaman
1 . Diagram Alir Rumusan Masalah
...
- 4 2.
Karakter-Karakter Morfologi Genus Rana yang Diukur...
24...
3 . Peta Lokasi Penelitian 25
4
.
Salah Satu Sudut Pemandangan Habitat GenusRana
di Kupang...
345
.
Salah Satu Sudut Pemandangan Habitat Genus Rana diTTS...
37 6 . Salah Satu Sudut Pemandangan Habitat Genus Rana di Bdu...
40...
7 . Morfologi Rana Zimnocharis -45
...
8 . Morfologi Rana cancrivora 47
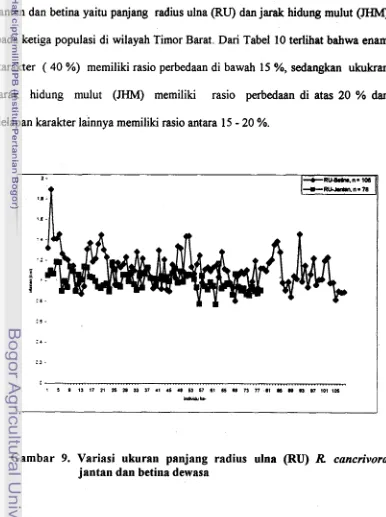
9 . Variasi Ukuran Panjang Radius Ulna (RU) Rana cancrivora Jantan dan ...
Betina di Wilayah Timor Barat 53
10
.
Variasi Ukuran Jarak Hidung Mulut (JHM) Rana cancrivora Jantan dan Betina di Wilayah Timor Barat...
54...
...
1 1 . Kromosom Rana cancrivora >. 56
...
12 . Kromosom Rana limnocharis 58
I. PENDAHULUAN A. Latar Belakang
Katak genus R a m sebagai anggota kelas Arnphibia dikenal sangat dekat
dengan kehidupan manusia. Keberadaan genus Rana ini memberikan kontribusi yang sangat besar dengan peranannya, antara lain (1) dalam aspek ekologi
sangat membantu dalam menjaga terciptanya keseimbangan ekosistem secara alami pada ekosistem daratan karena mengkonsumsi serangga seperti nyamuk,
ketam, wereng dan hama pengganggu lainnya (Inger, 1966; Sugiri et al,, 1998);
( 2 ) sebagai hewan laboratorium yang sering digunakan untuk menunjang perkembangan ilmu pengetahuan; (3) sebagai salah satu sumber pendapatan bagi masyarakat karena dapat dijadikan sebagai bahan makanan sumber protein.
Pada beberapa tahun terakhir, penelitian tentang ekologi, morfologi dan
variasi genetik genus Rana pada beberapa tempat di Indonesia telah memberikan
gambaran tentang keberadaan beberapa spesies seperti Ram macrodon, Rana
chalco~lota, Rana erythraea, Rana hosey, Rana blythi, R a m limnocharis dan Raila cnllcri~)ora.
Dilihat dari penyebarannya pada beberapa daerah di Indonesia, beberapa jenis katak genus Rana seperti Rana cancrivora, 'Rana lintnocharis dan Rana
mac~.oci'on memiliki potensi yang tinggi untuk dikembangkan atau dibudidayakan
dibandingkan dengan spesies lainnya. Berdasarkan kelimpahannya, Pratomo
(1997) dan Nasaruddin (1998) melaporkan bahwa jumlah Rana cancrivora
Demikian juga yang dilaporkan oleh Sugiri et al.(1998) bahwa dari beberapa
spesies yang diidentifikasi di daerah Jawa dan Sumatera, kodok sawah (Rana
cancrivora) tergolong spesies yang paling banyak dikumpulkan.
Bagi masyarakat Indonesia sementara ini, kodok selain dapat dijadikan
sebagai hewan percobaan laboratorium, beberapa spesies seperti Rana
cancrivora, Rana linznocharis dan Rai~a nzacrodon merupakan spesies yang
banyak dikonsumsi (edible), baik untuk kebutuhan dalam negeri maupun
diekspor. Keadaan ini akan mendorong masyarakat untuk melakukan penangkapan secara tidak terkendali. Daerah penangkapan selama ini lebih banyak dilakukan di daerah Jawa dan Sumatera. Konsekuensinya, jika
kegiatan penangkapan tersebut tidak dibatasi ataupun tidak diimbangi dengan usaha budidaya, maka akan mengakibatkan jumlah kodok genus Runa pada
kedua daerah tersebut semakin berkurang. Pada sisi lain usaha budidaya kodok genus Rmla untuk tujuan komersial selalu dihadapkan dengan tingginya resiko kegagalan, baik karena sulitnya memperoleh bibit unggul secara kontinu, sifat
kanibalisme maupun adanya serangan dari predator yang dapat meningkatkan mortalitas. Kecenderungan yang sama terlihat pada kegiatan penelitian tentang
keragaman genetik kodok genus Rana di Indonesia yang selama ini lebih banyak dilakukan di Pulau Jawa dan Sumatera, walaupun sebarannya sangat luas dan relatif merata di seluruh wilayah Indonesia. Dengan demikian perlu dilakukan
penelitian pada daerah lain sehingga dapat diperoleh gambaran secara
menyeluruh tentang aspek bioiogi dari kodok genus Rana yang tersebar di berbagai wilayah Indonesia.
belum banyak diungkapkan. Sehubungan dengan ha1 di atas, maka dilakukan
penelitian dasar yang mengkaji secara simultan mengenai ekologi, morfologi dan variabilitas genetik genus Rana di wilayah Timor Barat -Nusa Tenggara Timur.
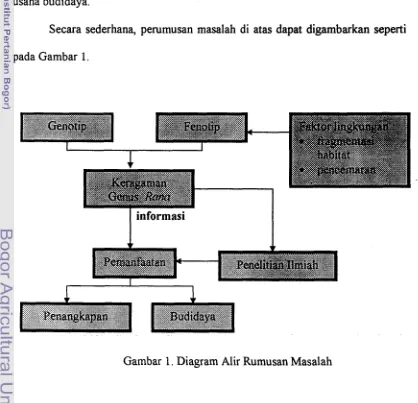
B. Perurnusan Masalah
Keberadaan kodok genus Rana selalu mengalami perubahan, baik karena
peristiwa evolusi alami maupun karena adanya aktivitas manusia yang tidak terkendali. Dengan demikian tingkat keragaman kodok genus Rana mempunyai
hubungan dengan kemampuannya dalam menyesuaikan diri dengan lingkungan sekitarnya.
Fragmentasi habitat akibat pertambahan penduduk maupun perubahan tata
guna lahan dan pencemaran perairan diduga merupakan tekanan lingkungan paling besar yang pada akhirnya dapat mempengaruhi kelangsungan hidup kodok genus Rana di wilayah Timor Barat. Apabila kondisi ini berlangsung terus dalam
waktu yang panjang akan berimplikasi pada penampilan ukuran dan bentuk serta
tingkat keragaman genetik.
Untuk mendapatkan Qambaran keanekaragaman genus Rnna dapat dilakukan dengan mengacu pada barakter fenotip dan genotip. Karakter fenotip biasanya menggunakan beberapa penanda seperti morfometri dan identifikasi warna dan bentuk tubuh. Walaupun praktis, pendekatan morfologi
memiliki kelemahan, antara lain ekspresi fenotip tidak selamanya mencerminkan
Dalam penelitian ini, karakter fenotip menggunakan morfologi dan morfometri yang dilakukan terhadap seluru h spesies dari kodok genus Rana,
sedangkan karakter genotip menggunakan penanda kromosom dan pola pita protein.
Penggunaan kedua jenis pendekatan serta dilengkapi dengan kajian
ekologi dapat memberikan gambaran yang lebih menyeluruh mengenai keberadaan kodok genus Rana di wilayah Timor Barat sehingga akan sangat berguna bagi pihak yang berkepentingan dalam memanfaatkannya secara lestari
melalui penangkapan yang terkendali maupun sebagai pengetahuan dasar bagi usaha budidaya.
Secara sederhana, perumusan masalah di atas dapat digambarkan seperti pada Gambar 1.
[image:116.530.74.492.313.716.2]informasi
C. Tujuan dan Manfaat ~enelitian
Penelitian ini bertujuan untuk mengetahui : 1) kondisi lingkungan atau
habitat alami kodok genus Ratla di wilayah Timor Barat; 2) keragaman morfologi spesies genus Rana secara geografis; 3) derajat dimorftsme seksual kodok genus Rana dan 4) keragaman genetik kodok genus Rana di wilayah Timor Barat.
Identifikasi habitat atau lingkungan yang dikaitkan dengan morfologi dan
keragaman genetik merupakan komponen yang saliag terkait. Oleh karena itu hasil penelitian ini diharapkan dapat bermanfaat sebagai bahan informasi bagi
pengembangan ilmu pengetahuan maupun dalam rangka mendukung upaya mempertahankan keberadaan serta mengembangkan potensi kodok genus Rana di wilayah Timor Barat khususnya dan Kawasan Indonesia Timur pada
11. TINJAUAN PUSTAKA
A. Klasifikasi dan Ciri-Ciri Morfologi
Katak tennasuk kelas Amphibia, yaitu vertebrata pertama yang beralih
dari kehidupan air ke kehidupan di darat. Dalam daur hidupnya, hewan ini
mernbutuhkan dua ternpat yaitu lingkungan perairan dan daratan (Dillon, 1965;
Hickman, C. P. Hickman Jr, and F. M. Hickman, 1974). Lebih lanjut dijelaskan
bahwa kehidupan kodok di kedua lingkungan berrnula atau diawali di air tawar
pada stadium larva dan dilanjutkan ke kehidupan di daratan pada stadium anakan
(juvenil) hingga dewasa untuk mencari kehidupan yang lebih menguntungkan.
Kampen (1923) menyatakan bahwa kelas Amphibia yang terdapat di
kepulauan Indo-Australia terdiri atas tujuh farnili antara lain Caecilidae,
Pelobatidae, Brecipitidae, Hylidae, Bufonidae dan Ranidae. Selanjutnya Iskandar
(1996) menyebutkan bahwa sebagai anggota dari famili Ranidae, genus Rnna di
Indonesia menyebar secara kosmopolitan meliputi Pulau Sumatera (30 jenis),
Jawa (1 5 jenis), Kalimantan (42 jenis), Nusa Tenggara (1 1 jenis), Sulawesi (16
jenis) dan Maluku (9 jenis). Sugiri et a1.(1998) dan Tritawani (1998) berhasil
rnengidentifikasi sebanyak tujuh spesies yang terdapat di daerah Jawa dan
Sumatera Barat seperti Rana hosei, Rana blythi, Rana limnocharis, Rana
chalconota, Rana ntacrodon, Rana erythraea dm1 Rana cancrivora.
Genus Rana menyebar secara kosmopolit. Biasanya telur diletakkan di air
dan mempunyai larva yang bebas berenang. Pada rahang atas terdapat gigi,
sedangkan pada rahang bawah spesies tertentu terdapat sepasang tonjolan tulang
seperti gigi, tidak mempunyai tulang rusuk, langit-langit rnulut terdapat vomer
yang terletak di antara koarla. Timpanum terletak berpadanan dengan kulit; pupil
mata berbentuk seperti ketupat dan terletak horizontal; jari-jari tungkai depan
biasanya tidak berselaput; jari tungkai belakang antara satu dengan lainnya
terdapat selaput renang (Kampen, 1923 ; Inger, 1966). Raw tidak mempunyai
kelenjar bisa pada kulit; pada perrnukaan bawah jari tangan ke empat antara
tuberkel dan cakram pada ujung jari terdapat paling banyak dua tuberkel. Ujung
jari tangan dan kaki membesar membentuk cakram, dan pinggiran cakram tidak
terdapat celah yang melingkar secara horizontal. Metatarsus luar 213 bagian
terpisah oleh selaput renang (Kampen, 1923).
Rnna cnncrivortl. Ratla cancrivora memiliki ciri-ciri antara lain moncong tumpul bulat atau agak runcing; ukuran panjang kepala sama dengan
lebar atau lebih panjang dari lebar kepala; tonjolan tulang di ujung rahang bawah
tidak berkembang (Inger,1966); timpanum jelas dengan diameter 112
-
2/3diameter mata, pada punggung terdapat banyak lipatan kulit (plika longitudinal),
terdapat lipatan supratimpani yang jelas dari mata ke aksila (Kampen,1923;
Susiri er n1.,1997); warna pada permukaan atas tubuh abu-abu, coklat dengan
bercak benvarna hitam atau kelabu tua yang susunannya tidak teratur, kulit
bagian belakang punggung berbintii; tungkai depan banyak bercak berwarna
hitam atau kelabu, bagian bawah tubuh berwarna putih; jari-jari tangan dan kaki
runcing dengan ujung jari tidak membesar, selaput renang hampir mencapai
ujung jari di sisi sebelah luar pada jari 1, 2, 3 dan sisi sebelah dalam pada jari kelima. Pada jari keempat selaput renang mencapai tuberkel subartikular distal
(Inger, 1966; Kampen, 1923; Sugiri et al., 1998). Ciri-ciri sekunder : katak betina
terdapat sepasang kmtung suara dan bantalan kopulasi di bawah jari tangan
pertama.
Rann nzncrodon Dumeril dan Bilbron. Tubuh gemuk, kaki kokoh
dengan ukuran agak panjang (tibia 0,45
-
0,56 panjang tubuh); panjang tubuh dapat lebih dari 125 mm; ukuran panjang kepala sedikit lebih besar dari padaukuran lebar kepala; moncong tumpul; kantus rostra1 turnpul; daerah pipi sangat
miring dan sedikit cekung; gendang telinga (timpanum) selalu kelihatan; gigi
vomer terletak dalam dua kelompok miring dan sudut anteriomedian koana;
mandibula bagian depan dengan sepasang penonjolan tulang; dewasa kulit
pung,oung halus; katak dengan ukuran 75 mm dengan bercak berbentuk huruf V
di antara kedua kaki depan dan tubuhnya berbintil; terdapat plika temporal; tanpa
plika dorsolateral. Ujung jari kaki depan bulat; jari pertarna lebih panjang dari jari
kedua; rigi kulit terdapat di sepanjang tepi dalam jari kedua dan ketiga. Bentuk
ujung jari kaki belakang bulat dan tanpa lekukan; selaput renang mencapai ujung
kaki, akan tetapi melekuk sampai ke pangkal tuberkulum subartikular tengah
antara jari keempat dan kelima. Gelambir terdapat di sepanjang sisi luar jari
pertama: tuberkulum subartikular berbentuk oval; tuberkulum metatarsal dalam
berbentuk oval, tanpa tuberkulum metatarsal luar. Warna pada waktu hidup
coklat sampai merah, biasanya dengan tot01 atau lorek hitam pada tungkai
belakang: Aurang lebih 1/6 jumlah spesimen mempunyai garis punggung
(Inger, 1966).
besar dari pada lebar kepala; moncong runcing; kantus rostra1 jelas; daerah pipi
hampir vertikal dan cekung; gigi vomer seperti pada Rana macrodon; pada
mandibula bagian depan terdapat sepasang penonjolan tulang. Terdapat plika
temporal; keadaan kaki sama dengan Rana macrodon; selaput renang mencapai
ujung cakram dan lebih penuh dari Rana ntacrodon. Warna h l i t coklat sampai
merah. Katak muda kulitnya dengan tot01 hitam di punggung; terdapat garis
interorbital; bibir dengan lorek hitam; terdapat garis loreal antara mata dan
hidung; terdapat garis hitam antara mata dan timpanum. Warna kaki sama dengan
Ranu n~cicrodon (Inger, 1966).
Rntw erythmea. Tubuh kecil; katak dewasa berukuran panjang 30
-
75 mm; panjang kepala lebih panjang dari lebar kepala; timpanum terlihat jelas ( kuranglebih 314 dari diameter mata). Ujung-ujung jari melebar bempa cakram; jari pertama sama dengan atau lebih panjang dari pada jari kedua; sebelah luar jari
bergelambir; tibia 0,50
-
0,57 dari panjang tubuh. Kulit lebih halus; pada sebuah sisi terdapat lipatan dorsolateral yang panjang; dengan sebuah lipatansuprarimpanurn yang lemah; permukaan ventral halus. Hewan hidup biasanya
benvarna hijau terang; lipatan dorsolateral berwarna kuning; bibir atas dan
permukaan ventral berwarna putih (Inger, 1966; Matsui, 1987).
Rnnn chnlconotn. Tubuh dan kaki ramping (kecil); katak dewasa berukcran panjang 33
-
60 mm; kepala segitiga; lebar kepala 0,27 - 0,30 dari panjang tubuh; kepala 0,37-
0,42 dari panjang tubuh. Ujung-ujung jari melebar, mempunyai tuberkel metatarsal luar. Kulit lebih kasar atau bergranula; hewanhidup benvarna kuning pucat atau hijau kekuningan; bagian bawah berwarna
merahan. Dalam alkohoi biasanya berwarna coklat kemerah-merahan (Inger,
1966; Matsui, 1979).
Rann Izosei. Tubuh kuat; kaki panjang dan kecil; katak dewasa berukuran 50- 100 mm; kepala berbentuk segitiga dan bagian ujung tumpul; lebar kepala
0,30
-
0,40 dari panjang tubuh; panjang kepala 0,36-
0,40 dari panjang tubuh;mata besar; timpanum jelas 215
-
314 dari diameter mata; panjang tibia 0,57-
0,66dari panjang tubuh; terdapat sepasang lipatan dorsolateral, tanpa tuberkel
metatarsal luar. Hewan hidup berwarna hijau gelap, kecoklatan pada sisi dan pada
permukaan dorsal anggota badan; bibir atas dan bawah berwarna putih keperak-
perakan. Dalam alkohol berwarna abu-abu gelap (Inger, 1966).
B. Habitat dan Penyebaran
Katak genus Rana terdapat di kolam-kolam berumput atau tumbuhan lain
di tepian sungai berbatu, serta pinggiran air yang ditumbuhi alang-alang. Selain
itu cgenus ini juga ditemukan di daerah pegunungan pada ketinggian 1400 m dari
permukaan laut dan di tepian sungai kecil dan sungai-sungai lebar (Menzies,
1975). Genus ini juga ditemukan di tempat-tempat basah pada daerah dataran
rendah dan selalu di dekat air (Villee, 1968).
Inger (1966) menyatakan bahwa, beberapa spesies dari genus Rana ditemukan di daerah persawahan, rawa, kolam, selokan, tempat-tempat yang
berair di hutan yang telah diubah menjadi daerah pertanian dan dataran rendah di
secara tetap di air payau, dimana Rana cancrivora dapat mempertahankan
keseimbangan osmotik dengan lingkungan sekitarnya (air garam) dengan cara
menekan urea yang terdapat ddam cairan tubuh maupun pada jaringan.
Yanaihara et a1.(1995) melaporkan bahwa R a m cancrivora yang hidup di Ang
Sila dekat Bangkok dapat hidup normal di rawa-rawa mangrove dengan salinitas
33 %.
Di Mexico, ditemukan dua spesies dari genus Rana (R. magnoacularis dan
R. be]-landieri forreri) yang menyebar secara luas mulai dari pesisir gersang yang
sempit di bagian barat laut "Mexico" hingga lereng bagian barat dari Sierra
Mudre, memperlihatkan kondisi yang sama baik pada habitat yang berbeda.
Misalnya, spesies yang telah diidentifikasi pada dua daerah yang berbeda yaitu
pada kolam air hujan dan sungai yang terletak dekat pesisir memperlihatkan
kondisi yang sama baik dengan spesies yang hidup di sungai di daerah
pegunungan dengan ketinggian 1500 m. Selanjutnya di daerah dengan ketinggian
yang lebih rendah dengan air yang tersedia secara tetap, R ntagnoacularis rnampu melampaui habitat R. berlandieri forreri yang dibatasi oleh sungai utama
dan area rawa-rawa dekat pesisir dan hidup secara bersama atau simpatrik (Frost
and Basnara, 1976).
Hal yang sama juga terjadi pada Ratla yipen dan Rana blairi, meskipun
R a m pipell hidup dengan normal di daerah dengan ketinggian 1800
-
2750 mdari permukaan laut, sedangkan Ratla blniri hidup di ketinggian 1200
-
1800 m.Kedua populasi spesies tersebut dapat ditemui hidup bersama atau simpatrik di
Di Indonesia, penyebaran spesies dari genus Rana cukup bervariasi antara
satu daerah dengan daerah lain. Rana cancrivora di Indonesia hidup di sekitar
perairan air tawar, di sawah atau rawa, koIam, selokan (Sugiri et al., 1998).
Berdasarkan ketinggian habitatnya, Pratomo (1997) menyatakan bahwa Rana
car~crivora yang terdapat di daerah Jawa Barat dapat hidup dan menempati lokasi
dengan ketinggian antara 120 m sampai dengan 1000 m di atas permukaan laut.
Habitat alami katak dewasa genus Raila pada umumnya adalah daratan
perairan tawar antara lain persawahan dan rawa. Lingkungan perairan utamanya
digunakan untuk kegiatan reproduksi, yaitu kawin, bertelur dan berkembangnya
berudu sampai menjadi anak katak setelah proses metamorfosis.
Setiap hewan ektotermik mempunyai kisaran toleransi suhu y ang memungkinkan metabolisme dan proses fisiologi lainnya dapat berjalan secara
normal. Apabila hewan berada pada suhu tubuh di atas titik tertentu
(CTM
max = suhu kritis maksimum) atau suhu tubuh di bawah titik tertentu(CTM
min = suhu kritis minimum) maka hewan tidak mampu untuk menampilkan pergerakan yangterkoordinasi. Pada beberapa amfibia, CTM max yang tertinggi hampir mencapai
4 2 ' ~ : termasuk B~rfo marir~ns, katak tropika daerah savana terbuka. Arnfibia
cenderung untuk melakukan fungsinya pada suhu lingkungan yang lebih rendah
dibandingkan dengan reptilia. Batas toleransi kritis dari salamander antara - 2 ' ~
hingga mencapai 2 7 ' ~ ~ sedangkan Anura antara 30- 4 1 ' ~ (Goin et al.. 1978).
Hasil penelitian Putnam dan Bennett (1981) tentang kinerja Bzrfo boreas
dan Rciim yipen, dengan menempatkan kedua spesies tersebut pada suhu
lingkungan yang konstan ( 3°,90,150,200,300~) untuk B. boreas dan untuk R.
0
meningkat seiring dengan peningkatan suhu tubuh dan mencapai maksimum pada
suhu 2 8 ' ~ untuk B. boreas dan 20-29'~ untuk R. pipen.
Susanto (1994) menyebutkan bahwa secara umum katak hidup di habitat
perairan dengan lingkungan yang bersuhu 20
-
3 5 ' ~ , sedangkan kisaran yangideal untuk pembiakan dan penetasan telur Rana catesbiana adalah 24
-
27%. Kisaran derajat keasaman (pH) yang ideal untuk hewan akuatik (termasukbemdu) adalah 6,5
-
8,5 (Pescod,l973), sedangkan pH optimal untuk Ranatenrporia di daerah Nottingham Inggris adalah 6. Berudunya hidup pada perairan
dengan kisaran pH 4,5
-
6. Pada pH h a n g dari 4,5 terjadi banyak kegagalantelur yang menetas dan letal ataupun dapat menyebabkan pertumbuhan yang
kerdil anak-anak Rana tenporaria (Tyler-Jones and Beattie, 1992). Rana
catesbiana di Ontario Canada mempunyai pH letal untuk embrio dan berudu pada
kisaran 3,5
-
3,8, sedangkan penurunan daya tahan hidup terjadi pada pH 4-
4,s(Licht and Grant, 1993).
Pengujian toleransi asiditas dari embrio dan larva kodok Boreae (Bufo
boreas) yang dipelihara dalam dua buah medium yaitu medium perairan pertambangan Argo dengan perlakuan pH 3,l
-
7 dan medium perairan danau"Black" dengan pH 7,l sebagai kontrol dengan jumlah masing-masing 100 zigot.
Hasil penelitian tersebut menunjukkan bahwa larva yang dipelihara di medium
perairan pertambangan pada pH 3 , l seluruhnya mati dalam waktu 24 jam. Sementara pada cairan yang mempunyai pH di atas 3,l memperlihatkan bahwa
semua larva dapat bertahan hidup dan mengalami metamorfosis secara lengkap
setelah mencapai hari ke 59 (Porter and Hakanson, 1976).
penting sekali untuk metabolisme organisme akuatik aerob dan organisme yang
hidup di sekitarnya seperti macam-macam hewan air dan amfibia (Wetzel, 1983).
Oleh karena itu di perairan yang tercemar hngisida bermerkuri di Dharward
India te jadi banyak kematian Rarla cyarrophylities dan bemdunya yang diakibatkan oleh ke j a racun saraf dari merkuri klorida dan diduga juga karena
minimnya kandungan oksigen pada perairan tersebut (Saidapur and Kanamadi,
1991).
Karbondioksida bebas dalam perairan sangat penting untuk proses
fotosintesis oleh tumbuhan air yang pada akhirnya akan menghasilkan oksigen
untuk keperluan pernapasan dan proses metabolisme.
C. Keragaman Kariotip Genus Rana
Kariotip merupakan gambaran lengkap kromosom pada tingkat metafase
suatu sel yang tersusun secara teratur dan merupakan pasangan-pasangan pada sel
diploid yang normal (Elridge, 1985). Pembuatan preparat kromosom diambil dari
sel dalam tahapan metafase dengan pemberian kolkisin. Studi pada berbagai
spesies dari kelas Arnphibia dan Reptilia telah banyak dilakukan guna melihat
keragaman antar spesies maupun untuk mengetahui hubungan kekerabatannya.
Berdasarkan hasil penelitian yang dilakukan oleh Menzies dan Tippett
(1976) terhadap kromosom dari 22 spesies Litoria di Papua dan 6 Nyctinzystes ditemukan bahwa dengan satu pengecualian (L. ir?fraJi.enata yang rnempunyai
jumlah kromosom haploid sebanyak 12), seluruh spesies lainnya ditemukan
Berdasarkan jumlah kromosom, Sugiri (1979) dan Tritawani (1996)
mengelompokkan kodok genus Rana ke dalam 3 (tiga) kelompok yaitu : 1)
+ spesies yang mepunyai jumlah kromosom diploid (2n) sebanyak 26 buah seperti
Rana cailcrivora, Rana erythraea, Rana limnocharis; 2) spesies yang mempunyai jumlah kromosom diploid (2n) sebanyak 24 buah seperti Rana hosei, Rana
n~acrodon dan Rana biythi; 3) spesies yang mempunyai jumlah kromosom
diploid (2n) sebanyak 22 buah yaitu Rana chalconota. Meskipun terdapat
kesamaan jumlah kromosom dari dua atau lebih spesies katak di atas, tetapi
secara empiris memperlihatkan adanya perbedaan dalam ukuran, susunan
maupun bentuk kromosom. Sebagai contoh, pada kelompok satu walaupun Rana
cnllcri~ora, Rana erythraea, Rana linlnocharis mempunyai ukuran kromosom
yang berbeda. Rana erythraea mempunyai empat pasang kromosom berukuran
besar dan sembilan pasang kromosom berukuran kecil. Tipe kromosom yang
ditentukan berdasarkan letak sentromer terdiri atas tipe metasentrik sebanyak
lima pasang kromosom dan delapan pasang kromosom yang bersifat
submetasentrik. Selanjutnya Rana lintwocharis memiliki dua pasang kromosom
berukuran besar yang masing-masing terdiri atas satu pasang kromosom
metasentrik dan submetasentrik. Dari sebelas pasang kromosom yang berukuran
kecil dapat dikelompokkan menjadi dua bagian yaitu enam pasang kromosom
metasentrik dan lima pasang kromosom submetasentrik.
Berdasarkan uraian di atas, maka dalam kaitannya dengan penentuan
keeratan hubungan filogeni semakin Sanyak terdapatnya kesamaan bentuk,
ukuran maupun susunan dari kromosom yang dimiliki oleh masing-masing
D. Polimorfisme Protein
Sebagaimana telah disebutkan sebelumnya bahwa selain menggunakan
penanda morfologi, keragaman genus Rana dapat juga diidentifikasi dengan
menggunakan penanda molekuler seperti kariotip, DNA mitokondrion dan
polimorfisme protein.
Protein dan enzim merupakan senyawa organik yang tersusun atas satu
atau lebih untai polipeptida melalui ikatan kovalen peptida Urutan asam amino
di dalam suatu polipeptida ditentukan oleh triplet nukleotida (kodon) di dalam
gen. Perbedaan susunan asam amino mencerminkan perbedaan kodon di dalam
gen. Dengan demikian perbedaan protein dipengaruhi oleh bentuk, ukuran, besar
muatan dan sifat kimia. Berbagai komponen protein serum pada pH di atas dan di
bawah tit ik isoelektriknya akan bergerak dengan kecepatan yang berbeda karena
adanya perbedaan muatan permukaannya (Harper, A. W. Rodwel dan P. A. Moyes, 1980).
Bila arus listrik dialirkan pada suatu medium penyangga yang berisi
protein plasma, maka migrasi komponen-komponen protein tersebut dimulai.
Sebagai contoh, molekul albumin yang mempunyai berat molekul yang lebih
kecil, memiliki laju migrasi yang paling cepat. Jadi dalam suatu medan listrik
dengan konsentrasi gel dan garam-garam tertentu protein akan bergerak ke arah
kutub muatan yang berlawanan dengan laju'yang propsional terhadap muatan
dan konformasinya (Guttman, 1985; Murphy et al., 1990).
Dalam dekade terakhir, metode elektroforesis telah banyak digunakan
untuk mempelajari populasi hewan (Nei, 1997; Murphy et a/., 1990). Pemisahan
teknik pewarnaan histokimia telah membawa revolusi besar dalam studi genetik
populasi. Metode elektroforesis protein terutama analisis isozim, selain mampu
mengungkapkan proses-proses evolusi, masalah-masalah yang berkenaan dengan
efisiensi enzim, keragaman genetik populasi alami, aliran gen, hibridisasi, jarak
genetik dan diferensiasi populasi bisa ditelaah (Murphy et al., 1990). Di pihak
lain, penggunaan polimorfisme protein sebagai penanda untuk mempelajari
genetik populasi mempunyai banyak kelemahan yang harus dipahami dengan
sungguh-sungguh, antara lain 1) adanya perbedaan pada sekuen DNA yang
mengkode asam amino yang bermuatan netral tidak mempengaruhi mobilitas
protein; 2) subtitusi asam amino yang bermuatan sama tidak mempengaruhi
mobilitas protein; 3) subtitusi nukleotida pada basa ketiga pada suatu kodon tidak
selalu membawa perubahan pada asam amino yang dihasilkan dan 4) sekuen-
sekuen DNA pada daerah intron tidak mengkode protein (Solihin, 1994).
Dari hasil penelitian Sugiri (1979) pada kodok batu (Rana blythi) dan kodok raksasa diperoleh masing-masing lima buah pita protein yaitu albumin,
globulin alfa 1, globulin alfa 2, globulin beta dan globulin gama dengan
konsentrasi yang berbeda. Pengukuran pada beberapa spesies dari genus Rana juga dilakukan oleh Tritawani (1997) dengan menganalisis pola pita protein dari
enam spesies katak memperlihatkan adanya perbedaan jarak migrasi pita protein
walaupun keenam spesies tersebut memiliki enam pita protein.
Berdasarkan hasil analisis tersebut dapat ditarik kesimpulan bahwa masing-
masing spesies yang diamati tersebut berbeda.
Nei (1987) menyatakan bahwa pendugaan heterogenitas genetik melalui
pendugaan yang minimum. Pendugaan akan mendekati keadaan populasi alaminya bila menggunakan jumlah sampel yang besar atau menganalisis banyak lokus. Beberapa ukuran keragaman genetik yang biasa digunakan, antara lain
nilai proporsi lokus polimorf (Ppoly). Nilai Ppoly yang dihasilkan sangat dipengaruhi oleh jumlah lokus dan jumlah sampel. Pada analisis keragaman genetik yang menggunakan jumlah lokus yang kecil serta ukuran sampel yang kurang dari 50 &an menyebabkan hasil perhitungannya tidak terlalu akurat.
Studi tentang tingkat keragaman genetik dari spesies
Rana
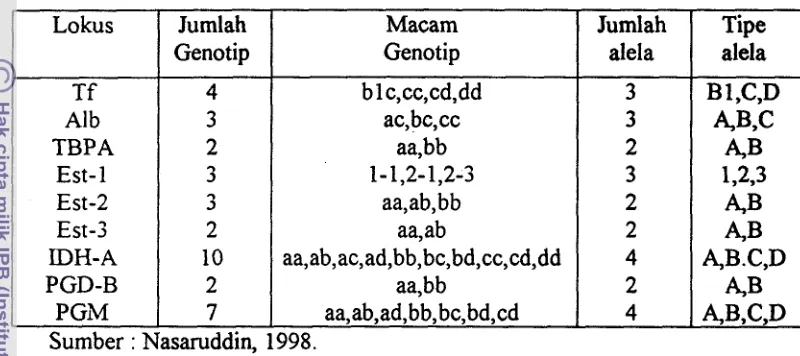
cancrivora yang terdapat pada beberapa daerah di Jawa Tengah dilakukan oleh Nasaruddin (1998) dengan melakukan pengamatan terhadap 92 individu diperoleh gambarandistribusi lokus untuk protein dan enzim seperti yang terlihat pada Tabel 1. Dari 18 lokus yang diperiksa ditemukan 9 lokus bersifat polimorf yaitu transferin (Tf), albumin(Alb), thyroxin binding prealbumin (TBPA), esterase (Est: 1,2,3), isositrat
&
dehidrogenase A(IDH-A), fosfoglukonat dehidrogenase B (PGD-B) dan fosfoglukomutase(PGM). Berdasarkan distribusi alela tersebut dapat
ditentukan keragaman genetik katak sawah dari masing-masing daerah dengan menggunakan nilai proporsi lokus polimorf (Ppoly) dan rataan heterozigositas (H). Dari keenam daerah yang diteliti diperoleh nilai Ppoly sebagai berikut :
Jepara (27,8%), Purwokerto (33,3%), Kebumen (44,4%), Yogyakarta (38,9%), Rawapening (44,4%) dad Boyolali (33,3%). Berdasarkan nilai Ppoly tergambar
bahwa umumnya populasi kodok sawah yang tersebar di Jawa Tengah memiliki
(12,5%), Jepara (15,4%), ~Boyolali (16,7%), Rawapening (19,1%), Kebumen
[image:131.534.80.480.161.339.2](19,3%) dan Yogyakarta (19,7%).
Tabel 1. Komposisi Genotip dari Sembilan Lokus Polimorf Kodok Sawah
di Jawa Tengah Lokus
T f A1 b TBPA Est- 1 Est-2 Est-3 IDH-A PGD-B PGlV
Keragaman genetik untuk spesies yang sama juga dilakukan oleh Sugiri et Jumlah Genotip 4 3 2 3 3 2 10 2 7 Macam Genotip blc,cc,cd,dd ac, bc,cc aa,bb 1-1,2-1,2-3 aa,ab,bb aa,ab aa,ab,ac,ad,bb,bc,bd,cc,cd,dd aa,bb aa,ab,ad,bb, bc,bd,cd
al., (1997) dengan menggunaan 16 protein diperoleh 5 lokus yang bersifat Sumber : Nasaruddin,
polimorf antara lain : Isositrat dehydrogenase mitokondrion (Icdm), Isositrat
dehydrogenase sitoplasmia (Isds), "Cell Esterase" (CEst 1; 2) dan esterase B
(ESB) seperti terlihat pada Tabel 2. 998. Jumlah alela 3 3 2 3 2 2 4 2 4
Tabel 2. Komposisi Genotip dari Lima Lokus Polimorf Kodok Sawah di Jawa Barat
Tipe alela
B 1 ,C,D A,B,C 4 B 1,2,3 4 B &I3 A,B.C,D A 3 AB,C,D LoLxs Icdm Icds ESB Cest 1
Tipe alela
4 B , C AB,C A,B.C,D AB,C,D 4 B , C Cest 2
Berdasarkan kelima lokus polimorf yang didapatkan dari analisis Jum 1ah Genotip 4 3 4 10
elektroforesis kodok sawah pada beberapa lokasi di Jawa Barat, nilai proporsi Sumber: Sugiri et nl.(l997)
4
lokus polimorf yang berbeda yaitu Bekasi, Ciamis, Cianjur, Kerawang, Macam
Geno t ip
aa,ab,bb,bc aa,bb,cc aa,bb,cc,dd aa,ab,ac,ad,bb,bc,bd,cc,cd,dd Jumlah alela 3 3 4 3
Pandeglang dan Sumedang (66,7%), Bogor dan Sukabumi (SO%), Majalengka dan Pati (83,3%). Nilai rataan heterozigositas tertinggi terdapat di Majalengka (36,9%) dan terendah di Sukabumi (22,3%).
Kim et al.(1976) telah mempublikasikan data keragaman genetik tiga spesies dari genus Bipes. Dengan menggunakan 21
-
22 lokus genetik yang dianalisis diperoleh persentase polimorfisme (P) dan rataan persentase lokus heterozigositas seperti tersaji pada Tabel 3.Tabel 3. T i n g h t Variabilitas Tiga spesies dari Genus Bipes
Spesies
r--
B. tridacylus B. caimlzo~ latus
I
Sumber : I
Heterozigositas
20 22 13,60 3,20
5 2 1 0,OO 0900
28 2 1 9,SO 0,20
Ukuran Sampel
Berdasarkan persentase polimorfisme dan rataan persentase heterozigositas
pada Tabel 3, diperoleh jarak genetik @) di antara ketiga spesies sebagai berikut
: B. tridcctylzcs dengan B. biporus sebesar 0,609, B.tri&ctylus dengan B. ca~mlic~rlatlrs sebesar 1,012 dan B. biporirs dengan B. canalicuIatzts sebesar 0,6 16.
Jumlah Lokus
Prosentase Polimorfisme
Rataan %
IIL METODE PENELITIAN
A. Tempat d m Waktu Penelitian
Penelitian ini telah dilaksanakan di daerah Timor Barat Propinsi Nusa
Tenggara Timur (NTT) dengan lokasi pengambilan sampel tersebar di tiga kabupatenlkota yaitu Kupang, Timor Tengah Selatan (TTS) clan Belu seperti
. -
yang terlihat pada Gambar 3.
Penelitian berlangsung dari bulan Maret 2001 sampai dengan bulan
Nopember 2001. Kegiatan penelitian terdiri dari tiga tahapan yaitu 1) kegiatan
lapansan yang meliputi pengamatan pendahuluan; 2) pengambilan sampel, pengukuran morfometri dan kualitas air habitat (suhu, pH, Oz terlarut dan COz
bebas); 3) pemeriksaan laboratorium meliputi analisis kromosom dan pola pita protein (dengan elektroforesis) dilakukan di laboratorium Pusat Studi Ilmu Hayati
IPB .
B. Bahan dan Alat Penelitian
Bahan yang digunakan adalah kodok genus Rana, sampel air yang
diperoleh dari perairan lokasi penelitian, gel akrilamid, alkohol 70%, buEer, dan formalin 3%. Alat yang dipergunakan terdiri atas pH meter, tennometer,
peralatan titrase, kaliper, timbangan elektrik, pewarna, "power supply", satu unit elektroforesis vertikal, refrigator, cawan dan mortir, labu didih, Erlemeyer, gelas
C. Metode Kerja
1. Pengamatan Pendahuluan
Sebelum melakukan observasi dan penangkapan katak pada masing- masing lokasi penelitian, terlebih dahulu dilakukan pengamatan pendahuluan yang meliputi kegiatan :
a. Mengetahui keberadaan katak genus Rana, jika tidak ditemukan salah s a t -
spesies genus Rana di suatu lokasi, maka lokasi tersebut tidak diteliti.
b. Jika di suatu lokasi ditemukan spesies dari genus Rana maka pengukuran Lualitas air habitat yang pertama (jam 06.00 ).
c. Pengamatan habitat secara umum pada siang hari.
Untuk mengetahui keberadaan kodok genus Rana, pada malam hari dilakukan survei sambil menangkap katak jika ditemui di lokasi.
2. Pengambilan Contoh
Pengambilan contoh dilakukan pada malam hari, menggunakan sorok (jarins bertangkai) atau langsung menggunakan tangan dan lampu sorot.
Penan@apan dilakukan dengan menyusuri daerah persawahan dan sungai pada ketiga populasi lokasi penelitian. Hasil tangkapan ditaruh dalam bubu dengan
Spesimen yang telah diidentifikasi, diukur bagian tubuh dan diambil
sampel darah, hati dan otot. Selanjutnya spesimen diawetkan dengan formalin 3% selama tiga jam lalu dipindahkan ke alkohol60%.
Sampel air, diambil dari sawah tempat spesimen diarnbil (genangan air
yang bersih) dan aliran air dari lokasi katak tersebut berada. Sampel air ditempatkan dalam botol pengukur (50 cc) yang telah diberi label berdasarkan stasiun pengambilan contoh yang telah ditetapkan sebelumnya.
3. Kualitas Air
Pengambilan sampel air pada masing-masing tempat dilakukan pada pukul
06.00 pagi hari dengan pengukuran pada masing-masing lokasi dilakukan sebanyak dua kali. Pengukuran kualitas air terhadap empat parameter sifat fisik, kimiatvi air antara lain suhu, pH, 0 2 terlarut, COz bebas dengan prosedur seperti terlihat pada Lampiran 1.
4. hiorfologi
Spesimen dipilah berdasarkan daerah asal penangkapan. Sebelum dilakukan
Gambar 2. Karakter-karakter morfologi Genus Rana yang diukur
[image:136.532.80.475.51.538.2]1
Kererangan :
0
Lokasi Penelitian [image:137.523.23.459.37.642.2]5. Kariotip
Pemeriksaan kariotip dilakukan menurut metode Satmoko (1970) dengan
sedikit modifikasi. Dalam pengamatan kariotip digunakan 2
-
3 ekor katak dari masing-masing spesies yang berhasil ditangkap di lokasi penelitian, dibuatsediaan kromosom dan diperiksa di bawah mikroskop. Selanjutnya dihitung
jumlah, tipe dan susunan kromosom dari setiap pengamatan.
Sebelum dilakukan pembuakn sediaan kromosom, maka disiapkan larutan-
larutan sebagai berikut :
a. Larutan kolkisin yang dosisnya tergantung pada berat badan spesimen.
Untuk metoda injeksi diperlukan kolkisin 0,0125 gram yang dilarutkan
dalam larutan fisiologis NaCl 0,8%.
b. Larutan hipotonis KC1 0,075 M dibuat dari KC1
SM
dengan melarutkan 2,795 gram KC1SM
di dalam akuades sampai menjadi 500 ml.c. Larutan fiksasi Carnoy adalah campuran alkohol glasial (etanoVmetano1)
dan asam asetat glasial dengan perbandingan 3: 1
d. Pewarna Giemsa lo%, dibuat dencgan melarutkan 1 bagian Giemsa dalam 9
bagian akuades.
Pengamatan kariotip dilakukan melalui langkah-langkah sebagai berikut :
a. Kodok sampel disuntik dengan larutan kolkisin 0,0125 gram kemudian
dibiarkan lepas dalam baskom selama 1- 2 jam.
b. Hewan dimatikan dengan eter dan dengan menggunakan gunting kecil
diambil sum-sum tulang pahanya, kemudian diletakkan dalam cawan petri.
c. Selanjutnya sum-sum tulang dimasukkan dalam larutan hipotonis KC1 0,075
jaringan clan dimasukan dalam tabung reaksi yang selanjutnya disentrifbs
(1 200 rpm) selama 10 menit.
d. Jaringan difiksasi dengan memasukkannya ke dalam botol-botol sampel
yang telah berisi larutan Carnoy sebanyak 3 cc, diaduk selama 15 menit.
Teduhkan selama 5 menit, kemudian disentrifbs dengan kecepatan 1200
rpm selama 10 menit dan larutannya diganti dengan larutan Carnoy yang
baru. Tahapan ini dilakukan sebanyak dua kali berturut-turut.
e. Buang supernatan dan tambahkan 0,5 cc cairan fiksasi, dicampur dengan
baik dan merata.
f Siapkan 3
-
5 gelas obyek bersih (yang sudah direndam dalam larutan etanol 50% selama 24 jam) untuk setiap sampel.g. Kemudian dengan menggunakan pipet suspensi sel diisap dan teteskan di
atas gelas obyek yang telah direndam dalam etanol 50%. Saat meneteskan
jarak pipet dengan gelas obyek minimal 10 cm, ini dilakukan pada beberapa
gelas obyek yang lain.
-
h. Gelas obyek dipanaskan di atas api alkohol hingga kering. Biarkan preparat
iernalaman, kemudian keesokan harinya diwarnai dengan larutan Giemsa
10% selama 10 menit.
i. Setelah itu bilas dengan air kran yang bersih dan bilas lagi dengan akuades
2
-
3 kali dan biarkan kering di udara. Apabila telah kering dapat diamati dibawah mikroskop dengan pembesaran 10 x 40 dan sampel kromosom yang
6. Pemeriksarn Polr protein
Preparasi dilakukan terhadap darah untuk mendapatkan proteinnya. Sebelum
pembedahan, katak dibius dengan kloroform selama kurang lebih 15 menit.
Darah diambil melalui ventrikel jantung dengan siring berheparin (Heparin
Sodium 5000 unit/ml, sebanyak 0,02 ml) sebagai antikoogulan. Plasma dan sel-
sel darah dipisahkan dengan sentrifbs dengan kecepatan 3000 rpm selama 10
menit.
Sel-sel darah yang terpisah selanjutnya dicuci dengan larutan Ringer (larutan
fisiologis) 0,69% sebanyak tiga kali, mengikuti prosedur Nishioka, Ohtani dan
Sumida (1980). Plasma dan sel-sel darah selanjutnya disimpan dalam lemari
pendingin dengan temperatur
-
2 0 ' ~ sampai pemeriksaan dilakukan. Sedangkan otot dan hati digerus dengan menggunakan tabung penggerus kaca dalam larutanbuffer 0,01 M Tris-HCL pH 8,O yang mengandung 0,001 M EDTA. Homogenat
disentrifbs dengan kecepatan 3000 rpm selama 5 menit pada sentrifbs
berpendingin. Supernatan disimpan dalam lemari pendingin dengan temperatur
-
2 0 ' ~ sampai dikerjakan lebih lanjut.
Dalam penelitian ini, analisis protein yang dilahkan dengan
mengygunakan teknik elektroforesis gel akrilamid dengan langkah-langkah
pelaksanaan analisis sebagai berikut :
(a) Keping gelas dan keping metal digabungkan dengan "plastik spacers" di
bagian lateral yang pendek, lalu diklem atau dijepit dan ditaruh berdiri di
(b) Penyumbat agarose dipanaskan supaya encer, lalu dipipet secukupnya
untuk menyumbat dasar keping dan sisi, biarkan sampai mengental (polimerasi).
(c) Larutan "running gel" dipipet ke keping (antara metal dan gelas) secara
merata, ditambah sedikit isobutanol di atas gel untuk mencegah oksigen menghambat reaksi, lalu dibiarkan agar terjadi polimerisasi. Setelah polimerisasi isobutanol dicuci dengan akuades d m dikeringkan dengan
kertas saring.
(d) Diletakkan sisir antara gelas dan metal di atas "running gel". Kemudian
larutan "stacking gel" dipipetkan ke keping, diusahakan agar tidak terbentuk gelembung udara dan dibiarkan sampai terjadi polimerisasi. (e) Selanjutnya sisir dilepaskan dengan hati-hati, lalu gel dan keping diklem
(dijepit) dalam "mini gel holder', gelas pada sisi luamya. Kemudian kamar atas dan kamar bawah (ruang di belakang) diisi dengan "running buffer"
(f) Masukkan "Marker" 5-10 pl ke dalam salah satu sumur yang merupakan protein standar, kemudian pada sumur lainnya diisi dengan protein sampel.
(g) "Water Cooling Houses" diletakkan pada inlet d m outlet dari "gel holdery'. Dua buah kabel disambungkan ke "power supply" sesuai dengan
tempatnya (kutub positif berwarna merah dan kutub negatif benvarna
sumur dengan tetap mernperhatikan kestabilan jarum penunjuk "power supply".
(h) "Power supply" dimatikan dan kedua kabel penghubung dilepaskan, dan dengan seksama kedua gel dilepaskan dari keping dengan menggunakan skapel dan dimasukkan ke dalam bak pewarna selama 3 0 menit.
(i) Lakukan pencucian dalam campuran 7% asam asetat dengan 50% metanol selama kira-kira 15 menit. Untuk peyimpanan gel dilalcukan dalam larutan 7% asam asetat (tanpa metanol), lalu difoto.
D. Analisis Data 1. Kualitas Air
Analisis kualitas air dari ke tiga lokasi penelitian meliputi empat parameter sifat fisik kimiawi yang meliputi suhu, pH, 02 terlarut, COz bebas dm seperti yang disajikan dalam Tabel 4. Pengukuran suhu (air dan lingkungan), pH, kandungan oksigen terlarut, karbondioksida bebas dilakukan di lapangan. Dalam
penelitian juga disajikan kondisi iklim habitat katak genus Rana sebagai data
pendukung berupa jumlah curah hujan tahunan maupun jenis flora yang ada pada habitatnya.
Tabel 4. Sifat Fisik Kimiawi Air yang Diukur, Metode serta Alat Ukur Kualitas Air
S u h u pH
0 2 terlarut C02 bebas
Untuk mendapatkan gambaran tentang keragarnan morfologi genus Rana
berdasarkan letak geografis pada ketiga lokasi penelitian, maka dilakukan analisis variasi morfologi dengan mendeskripsikan ciri-ciri sekunder kedua spesies
kodok. Data 15 karakter morfometri kodok genus Rana dianalisis dengan program komputer Minitab. Uji statistik didahului dengan menghitung range,
rataan hitung dan standar deviasi. Kemudian dilanjuthn dengan uji t untuk membandingkan perbandingan ukuran tubuh dan anggota tubuh kedua spesies
pada populasi Kupang maupun Rana cancrivora pada ketiga populasi. Pengujian dimorfisme seksual dilakukan dengan menghitung ratio perbedaan antara ciri-ciri
betina dan jantan dan dilanjutkan dengan uji t. Hasil uji t berguna bagi kepentingan aplikasi yaitu jika ter