KARAKTERISASI GEN ENDO- -1,4-GLUKANASE PADA
RAYAP
Coptotermes curvignathus
RUT NORMASARI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis Karakterisasi Gen Endo- -1,4-glukanase pada Rayap Coptotermes curvignathus adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada Perguruan Tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Januari 2011
Coptotermes curvignathus. Supervised by RIKA RAFFIUDIN and IMAN RUSMANA.
Termites are social insects that have a distinct task in form of caste within their colony. Workers play an important role in finding and degrading the cellulose food, due to the existence of endo- -1,4-glucanase enzyme. Hence, this study was aimed to characterize Coptotermes curvignathus endoglucanase gene. Primers were designed based on the Coptotermes formosanus (CfEG) cDNA and Nasutitermes takasagoensis (NtEG) genomic DNA. These introns of C. curvignathus all showed GT/AG rule. Exon two up to five of C. curvignathus endoglucanase were aligned with exon three up to six from NtEG; while the intron two to four of C. curvignathus endoglucanase were at the same position with that of intron three up to five on NtEG. Exon two up to five of C. curvignathus endoglucanase resulted 564 bp and BLASTN analysis revealed that C. curvignathus endoglucanase homologous to CfEG. Putative amino acid obtained from C. curvignathus endoglucanase showed its high homology with CfEG. Analysis based on Pfam revealed that C. curvignathus endoglucanase was member of Glycosyl Hydrolase Family 9 (GHF 9) family and six-hairpin glycosidase superfamily. Motifs found in endoglucanase C. curvignathus were one consensus signature of GHF 9, nucleophile, co-nucleophile, salt bridge, and N-terminal. Based on DNA and amino acid homology between CfEG and endoglukanase C. curvignathus it was expected that endoglukanase C. curvignathus has similar high cellulase activity as shown in CfEG. This will be an essential database for finding an effective cellulase enzyme for further applications.
RINGKASAN
RUT NORMASARI. Karakterisasi Gen Endo- -1,4-glukanase pada Rayap Coptotermes curvignathus. Dibimbing oleh RIKA RAFFIUDIN dan IMAN RUSMANA.
Rayap mampu bertahan hidup dengan memanfaatkan lignoselulosa yang mengandung sedikit nutrisi. Kemampuan mendegradasi selulosa dimiliki oleh rayap pekerja. Rayap merupakan serangga sosial yang memiliki pembagian tugas yang dinyatakan dalam kasta. Kasta rayap terbagi menjadi ratu, pekerja, dan prajurit. Tugas utama dari pekerja adalah mengumpulkan dan mendegradasi makanan yang berupa selulosa.
Rayap mampu mendegradasi selulosa karena adanya enzim selulase yang dihasilkan rayap dan organisme simbion. Enzim selulase yang dihasilkan simbion berasal dari protozoa (rayap tingkat rendah) dan bakteri (rayap tingkat tinggi). Enzim selulase rayap berbeda dengan enzim selulase protozoa simbion rayap. Endoglukanase rayap termasuk ke dalam Glycosyl Hydrolase Family 9 (GHF 9) dan endoglukanase simbion rayap termasuk ke dalam GHF 7.
Coptotermes curvignathus merupakan salah satu rayap subteran yang makanan utamanya berupa kayu dan bahan lain yang mengandung selulosa. C. curvignathus tersebar di Asia Tenggara dan banyak menyerang tanaman di Indonesia. Hal ini mengindikasikan C. curvignathus memiliki kemampuan yang tinggi dalam mendegradasi selulosa. Data endoglukanase rayap belum ada pada daerah tropis, hanya terbatas pada rayap daerah subtropis.
Penelitian ini bertujuan untuk mengkarakterisasi ekson dan intron gen endo-ß-1,4-glukanase C. curvignathus. Hasil penelitian ini diharapkan dapat menjadi informasi dasar bagi penelitian secara in vitro enzim selulase C. curvignathus di Indonesia.
Sampel C. curvignathus dikumpulkan dari sarang rayap yang berada di pohon pinus (Pinus mercusii) pada halaman parkir Perpustakaan IPB, Kampus IPB Darmaga, Bogor. Berdasarkan data gen endoglukanase C. formosanus (CfEG) dan N. takasagoensis (NtEG) yang sudah terkarakterisasi, pada penelitian ini dilakukan karakterisasi gen endoglukanase pada C. curvignathus. Primer disusun berdasarkan data sekuen CfEG. Karakterisasi gen endoglukanase C. curvignathus dilakukan melalui tahapan ekstraksi DNA, amplifikasi DNA, elektroforesis, pewarnaan DNA, serta proses sekuen. Elektroforesis DNA menggunakan PAGE 6% dan pewarnaan DNA dengan perak nitrat. Proses sekuen dilakukan menggunakan jasa sekuensing. Hasil sekuen diedit menggunakan program Genetyx Win versi 4.0. Analisis DNA dan asam amino terdiri dari alignment DNA dan asam amino menggunakan program Clustal X, analisis homologi menggunakan BLASTN dan BLASTP melalui situs NCBI (www.ncbi.nlm.nih.gov), serta analisis filogeni dan jarak genetik menggunakan program MEGA 4. Analisis motif protein putative meliputi consensus signature, nucleophile, co-nucleophile, salt bridge, dan N-terminal. Data endoglukanase rayap, echinodermata, cacing tanah, moluska, protozoa simbion rayap, fungi, dan bakteri digunakan dalam analisis filogeni.
menggunakan pasangan primer CfF2 dan CfR3, CfF3 dan CfR4, CfF4 dan CfR5 masing-masing sebesar 856, 1077, dan 484 pb.
Berdasarkan hasil alignment didapatkan panjang ekson 2, intron 2, ekson 3, intron 3, ekson 4, intron 4, dan ekson 5 masing-masing sebesar 160, 118, 96, 842, 183, 187, dan 125 pb. Alignment dengan NtEG menunjukkan posisi ekson dua Sedangkan panjang ekson dua dan lima gen endoglukanase C. curvignathus berbeda dengan panjang ekson tiga dan enam NtEG dan ekson dua dan lima CfEG. Perbedaan panjang juga terdapat pada masing-masing intron gen endoglukanase C. curvignathus dengan intron NtEG.
Intron dua sampai empat gen endoglukanase C. curvignathus didominasi oleh basa AT yaitu masing-masing sebesar 66.06, 58.81, dan 55.96%, serta diawali dengan basa GT dan diakhiri dengan basa AG. Kesamaan intron dua sampai empat gen endoglukanase C. curvignathus dengan intron tiga sampai lima NtEG masing-masing sebesar 8.8, 24.8, dan 46.5%.
Hasil analisis BLASTN ekson dua sampai lima gen endoglukanase C. curvignathus homolog dengan CfEG dengan nomor akses EU853671 (95%) dengan E-value 0.0. Kandungan GC ekson dua sampai lima gen endoglukanase C. curvignathus adalah 55.82%. Asam amino putative yang didapatkan sebesar 188 asam amino. Hasil analisis homologi asam amino putative gen endoglukanase C. curvignathus menggunakan BLASTP homolog (94%) dengan CfEG (BAB40696) dengan E-value sebesar 9e-92.
Gen endoglukanase C. curvignathus mengelompok dengan GHF 9 pada kelompok rayap dan terpisah dengan GHF 9 pada kelompok moluska, echinodermata, cacing tanah, dan bakteri. Gen endoglukanase C. curvignathus juga terpisah dengan GHF 7 pada kelompok fungi dan protozoa. Hasil perhitungan jarak genetik gen endoglukanase C. curvignathus lebih dekat dengan gen endoglukanase C. formosanus EG3, C. formosanus, dan C. formosanus EG4 dengan nilai jarak genetik yang paling kecil yaitu 0.055. Hubungan gen endoglukanase N. takasagoensis yang terjauh adalah dengan gen endoglukanase C. acinaciformis dengan jarak genetik terbesar yaitu 0.174.
Analisis motif protein putative gen endoglukanase C. curvignathus menunjukkan adanya satu consensus signature GHF 9, nucleophile, co-nucleophile, salt bridge, dan N-terminal. Pencarian dengan Pfam menunjukkan endoglukanase C. curvignathus masuk ke dalam gen famili GHF 9 yang termasuk ke dalam CL0059 yaitu six-hairpin glycosidase superfamily.
signature GHF 9, sedangkan pada gen endoglukanase C. curvignathus baru ditemukan satu consensus signature GHF 9 dan belum ditemukan posisi dari pusat katalitik. CfEG diekspresikan selulase pada kelenjar saliva dan usus tengah. Berdasarkan penelitian ini, untuk membuktikan ekspresi selulase pada C. curvignathus perlu dilakukan analisis mRNA pada gen endoglukanase C. curvignathus terutama pada rayap pekerja. Selain itu perlu dilakukan uji aktivitas endoglukanase untuk aplikasi selanjutnya.
Hak Cipta milik IPB, tahun 2011
Hak cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tidak merugikan kepentingan yang wajar IPB
KARAKTERISASI GEN ENDO- -1,4-GLUKANASE PADA
RAYAP
Coptotermes curvignathus
RUT NORMASARI
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Biosains Hewan
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Judul Tesis : Karakterisasi Gen Endo- -1,4-glukanase pada Rayap Coptotermes curvignathus
Nama : Rut Normasari NIM : G352080091
Disetujui
Komisi Pembimbing
Dr. Ir. Rika Raffiudin, M.Si. Dr. Ir. Iman Rusmana, M.Si.
Ketua Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana Biosains Hewan
Dr. Bambang Suryobroto Prof. Dr. Ir. Khairil A. Notodiputro, M.S.
Puji dan syukur penulis panjatkan kepada Tuhan Yesus atas segala berkat dan kasihNya sehingga karya ilmiah ini berhasil diselesaikan. Karya ilmiah ini merupakan syarat untuk memperoleh gelar Magister di Institut Pertanian Bogor. Judul yang dipilih dalam penelitian ini ialah Karakterisasi Gen Endo- -1,4-glukanase pada Rayap Coptotermes curvignathus.
Terima kasih penulis ucapkan kepada Dr. Ir. Rika Raffiudin, M.Si dan Dr. Ir. Iman Rusmana, M.Si selaku pembimbing yang telah banyak memberikan ilmu, bimbingan, masukan, dan dorongan kepada penulis. Terima kasih penulis ucapkan kepada Dr. Ir. Idham Sakti Harahap, M.Si selaku penguji yang telah memberikan saran dan masukan kepada penulis. Ucapan terima kasih penulis tujukan kepada Bapak Bambang Suryobroto, Bapak Achmad Farajallah, Bapak Tri Atmowidi, Bapak Tri Heru Widarto, Ibu Taruni Sri Prawasti, Ibu RR Dyah Perwitasari, dan Mbak Kanthi Arum W atas semua ilmu, pengalaman, bimbingan, nasihat, dan dorongan yang telah diberikan. Bapak Adi, Ibu Tini, dan Ibu Ani atas bantuannya selama di laboratorium. Teman-teman Biosains Hewan 2008, Ruth MW, Jazirotul, Dzulfaqor, Mbak Tetri, teman-teman zoologi, dan teman-teman Villga atas dukungan semangat, kebersamaan, bantuan, dan doanya.
Terima kasih juga kepada Sari, Wahyu, dan Dona atas dukungan dan persahabatan yang telah terjalin selama ini. Tak lupa kepada kakak, adek, teman-teman PA, serta Jemaat Bogor atas perhatian, dukungan, canda tawa, dan kekompakannya. Ungkapan terima kasih penulis sampaikan kepada Ayah, Ibu, dan Kakak-kakak tercinta atas dukungan, doa, cinta, dan kasih sayang yang sangat besar yang telah diberikan kepada penulis, serta pihak-pihak yang membantu dalam penyelesaian karya ilmiah ini.
Semoga karya ilmiah ini bermanfaat.
Bogor, Januari 2011
RIWAYAT HIDUP
Penulis dilahirkan di Tuban pada tanggal 2 Oktober 1984 sebagai putri dari Sukarman dan Wagiarti. Penulis merupakan putri keempat dari empat bersaudara.
Tahun 2003 penulis lulus dari SMA Negeri 1 Tuban dan pada tahun yang sama lulus seleksi masuk IPB melalui jalur Undangan Seleksi Masuk IPB (USMI). Penulis memilih Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam.
Halaman
Taksonomi dan Biologi C. curvignathus ... 5
Simbiosis Pada Saluran Pencernaan Rayap dan Metabolisme Selulosa ... 7
Struktur Ekson-Intron dan Ekspresi Gen Endoglukanase Rayap ... 9
BAHAN DAN METODE ... 12
Sekuen dan Alignment DNA dan Asam Amino ... 15
Analisis Filogeni dan Jarak Genetik ... 16
Analisis Protein Putative ... 16
HASIL ... 18
Amplifikasi DNA Gen Endoglukanase C. curvignathus ... 18
Sekuen dan Alignment DNA dan Asam Amino Gen Endoglukanase C. curvignathus ... 18
Analisis Filogeni dan Jarak Genetik Gen Endoglukanase C. curvignathus ... 24
Analisis Protein Putative Gen Endoglukanase C. curvignathus ... 26
PEMBAHASAN ... 27
Amplifikasi DNA Gen Endoglukanase C. curvignathus ... 27
Homologi DNA dan Asam Amino Putative Gen Endoglukanase C. curvignathus dan C. formosanus ... 28
Analisis Filogeni dan Jarak Genetik Gen Endoglukanase C. curvignathus ... 29
Analisis Protein Putative dan Homologi Ekspresi Gen Endoglukanase C. curvignathus dengan C. formosanus ... 30
SIMPULAN DAN SARAN ... 36
Simpulan ... 36
Saran ... 36
DAFTAR PUSTAKA ... 37
Halaman
1 Primer yang digunakan untuk mengamplifikasi DNA target gen endoglukanase C. curvignathus ... 14
2 Data endoglukanase pembanding dari GenBank ... 16
DAFTAR GAMBAR
Halaman
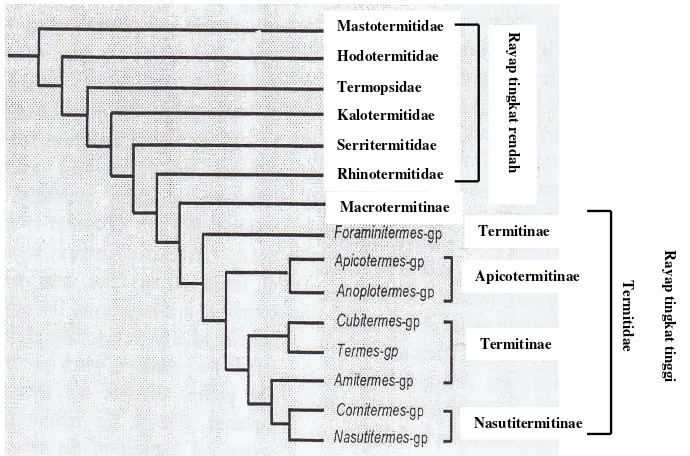
1 Filogeni rayap berdasarkan morfologi dan DNA mitokondria (16S rRNA dan NADH 5 dehydrogenease) (Kambhampati & Eggleton 2000) ... 4
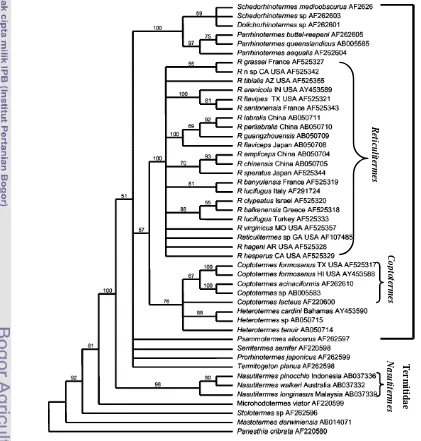
2 Filogeni genus Reticulitermes, Coptotermes, dan Nasutitermes berdasarkan gen cytochrome oxidase II (COII) (Austin et al. 2004) ... 6
3 Rayap C. curvignathus (a) pekerja dan (b) prajurit... 7
4 Morfologi saluran pencernaan rayap (Scharf & Tartar 2008) ... 8
5 Filogeni rayap berdasarkan gen endoglukanase dan pergeseran tempat
ekspresi (Tokuda et al. 2004) ... 11
6 Skema posisi primer untuk amplifikasi gen endoglukanase C. curvignathus berdasarkan CfEG (AB058670) ... 15
7 Pita DNA hasil amplifikasi gen endoglukanase C. curvignathus pada gel poliakrilamid dengan menggunakan (a) pasangan primer CfF2 dan CfR3; (b) pasangan primer CfF3 dan CfR4 (no 3-4) dan pasangan primer CfF4 dan CfR5 (no 5-6) ... 18
8 Hasil sekuen gen endoglukanase C. curvignathus dengan menggunakan pasangan primer (a) CfF2 dan CfR3, (b) CfF3 dan CfR4, dan (c) CfF4 dan CfR5 ... 19
9 Sekuen DNA gen endoglukanase C. curvignathus ... 20
10 Skema posisi ekson dan intron NtEG (AB019146), CfEG (AB058670), dan gen endoglukanase C. curvignathus (hasil penelitian ini) ... 21
11 Hasil analisis BLASTN gen endoglukanase C. curvignathus dengan CfEG (EU853671). ... 22
12 Asam amino putative gen endoglukanase C. curvignathus. ... 23
13 Hasil analisis BLASTP asam amino putative gen endoglukanase C.
curvignathus dengan gen CfEG (BAB40696) ... 24
14 Hasil rekonstruksi pohon filogeni berdasarkan asam amino putative gen endoglukanase C. curvignathus menggunakan metode Maximum Parsimony (bootstrap 1000 kali). ... 25
Halaman
1 Kromatogram hasil sekuen gen endoglukanase C. curvignathus dengan menggunakan pasangan primer CfF2 ... 44
2 Kromatogram hasil sekuen gen endoglukanase C. curvignathus dengan menggunakan pasangan primer CfR3 ... 46
3 Kromatogram hasil sekuen gen endoglukanase C. curvignathus dengan menggunakan pasangan primer CfF3 ... 48
4 Kromatogram hasil sekuen gen endoglukanase C. curvignathus dengan menggunakan pasangan primer CfR4 ... 51
5 Kromatogram hasil sekuen gen endoglukanase C. curvignathus dengan menggunakan pasangan primer CfF4 ... 53
6 Kromatogram hasil sekuen gen endoglukanase C. curvignathus dengan menggunakan pasangan primer CfR5 ... 54
7 Hasil BLASTN exon dua sampai lima gen endoglukanase C. curvignathus .. 55
8 Hasil BLASTP asam amino putative gen endoglukanase C. curvignathus .... 56
9 Alignment NtEG, CfEG, dan endoglukanase C. curvignathus... 57
10 Alignment asam amino NtEG, CfEG, dan asam amino putative gen
PENDAHULUAN
Latar Belakang
Rayap merupakan hewan unik yang teradaptasi untuk bertahan hidup dengan mendapatkan makanan yang berasal dari lignoselulosa yang mengandung sedikit nutrisi (Ohkuma 2003). Kemampuan mendegradasi selulosa dimiliki oleh rayap pekerja. Rayap merupakan serangga sosial yang memiliki pembagian tugas yang dinyatakan dalam kasta. Kasta rayap terbagi menjadi ratu, pekerja, dan prajurit. Tugas utama dari pekerja adalah mengumpulkan dan mendegradasi makanan yang berupa selulosa (Krishna 1969).
Rayap mampu mendegradasi selulosa karena adanya enzim selulase yang dihasilkan rayap (endogenous cellulose) dan organisme simbion (Nakashima et al. 2002a, 2002b; Tokuda et al. 2007). Enzim selulase yang dihasilkan simbion berasal dari protozoa (rayap tingkat rendah) dan bakteri (rayap tingkat tinggi) (Krishna 1969).
Data genomik DNA lengkap gen endo-ß-1,4-glukanase (Endoglukanase, EG, EC 3.2.1.4) pada rayap berhasil dikarakterisasi pada N. takasagoensis (Tokuda et al. 1999) dan data partial genomik pada R. speratus (Itakura et al. 2006). Data cDNA lengkap gen endoglukanase terdiri atas R. flavipes (Scharf et al. 2005), R. speratus (Tokuda et al. 1999), C. formosanus (Nakashima et al. 2002b), C. acinaciformis, dan N. walkeri (Tokuda et al. 1999). Data partial cDNA gen endoglukanase terdiri atas M. darwiniensis (Tokuda et al. 1999; Li et al. 2003), H. sjoestedti (Tokuda et al. 2004), H. japonica (Lo et al. 2000), N. koshunensis (Lo et al. 2000; Tokuda et al. 2004), S. mushae, dan O. formosanus (Tokuda et al. 2004).
diekspresikan di usus tengah (Tokuda et al. 1997, 1999, 2004; Watanabe & Tokuda 2001; Nakashima et al. 2002b). Endoglukanase simbion rayap diekspresikan pada usus belakang rayap. Hal ini mengindikasikan ada dua sistem enzim selulase yang terlibat dalam proses pencernaan rayap tingkat rendah (Tokuda et al. 2007; Nakashima et al. 2002b; Zhou et al. 2007).
Rayap memanfaatkan tanaman yang mengandung selulosa, hemiselulosa, dan lignin. Selulosa merupakan biomassa yang paling melimpah di bumi. Selulosa dapat dihidrolisis menjadi glukosa dengan menggunakan selulase. Rayap memiliki gen yang dapat menghasilkan selulase. Khamir memiliki kemampuan untuk mengubah glukosa menjadi etanol dengan cara fermentasi. Hal ini memungkinkan untuk menyisipkan gen dari rayap yang menghasilkan enzim selulase ke khamir, sehingga khamir dapat menghasilkan selulase dan mengubah glukosa yang dihasilkan dari hidrolisis selulosa menjadi etanol (Matsui et al. 2009; Scharf & Boucias 2010).
Coptotermes curvignathus merupakan salah satu rayap subteran dan tersebar di Asia Tenggara (Krishna 1969; Roonwal 1970). C. curvignathus termasuk ke dalam Famili Rhinotermitidae yang makanan utamanya berupa kayu dan bahan lain yang mengandung selulosa (Bignell & Eggleton 2000). C. curvignathus merupakan hama yang banyak menyerang tanaman di Indonesia. Hal ini mengindikasikan C. curvignathus yang memiliki kemampuan yang tinggi dalam mendegradasi selulosa.
Data endoglukanase rayap belum ada pada daerah tropis. Data yang ada terbatas pada rayap daerah subtropik yaitu C. formosanus dari Jepang dengan nomor aksesi Genbank AB058670.
Tujuan Penelitian
Tujuan penelitian ini adalah untuk mengkarakterisasi ekson dan intron gen endo-ß-1,4-glukanase C. curvignathus.
Manfaat Penelitian
TINJAUAN PUSTAKA
Klasifikasi dan Biologi Rayap
Rayap di daerah subtropik disebut dengan ’semut putih’ (white ants) karena memiliki morfologi yang mirip dengan semut. Berdasarkan hubungan evolusi (filogeni), tidak ada hubungan antara rayap dengan semut. Hubungan lebih dekat terjadi antara rayap dengan kecoa (Blattodea) (Lo et al. 2000; Inward et al. 2007).
Rayap merupakan salah satu kelompok serangga dengan jumlah keragaman yang besar. Rayap (Ordo Isoptera) terdiri atas tujuh famili, yaitu Mastotermitidae, Kalotermitidae, Termopsidae, Hodotermitidae, Rhinotermitidae, Serritermitidae, dan Termitidae. Sampai sekarang sudah tercatat 14 subfamili, 281 genus dan lebih dari 2600 spesies termasuk dalam kelompok ini (Kambhampati & Eggleton 2000). Famili Termitidae merupakan famili terbesar dalam Ordo Isoptera dan mencakup tiga perempat spesies yang diketahui dan merupakan kelompok rayap yang paling maju. Spesies rayap yang termasuk famili Termitidae dikelompokkan ke dalam rayap tingkat tinggi, sedangkan enam famili yang lain dikelompokkan ke dalam rayap tingkat rendah (Gambar 1). Pembagian kelompok ini didasarkan atas morfologi dan DNA mitokondria (16S rRNA dan NADH 5 dehydrogenease). Karakter yang digunakan sebanyak 197 karakter morfologi dan biologi dari pekerja dan prajurit dengan menekankan pada struktur usus pekerja (Kambhampati & Eggleton 2000). Gen pada mitokondria seperti 16S rRNA, NADH 5 dehydrogenease, dan cytochrome oxidase II (COII) dapat digunakan sebagai kronometer evolusi karena bukan merupakan hasil rekombinasi, berubah sesuai dengan waktu tetapi tidak terlalu cepat, ada pada semua organisme, dan memiliki fungsi yang sama (Sogin et al. 1986; Hoy 1994; Gupta 1998; Philippe et al. 2000; Freeland 2005)
Rayap merupakan serangga sosial yang memiliki pembagian tugas yang jelas yang dinyatakan dalam pembagian kasta. Berdasarkan kemampuan bereproduksi rayap dibagi menjadi dua kasta yaitu kasta reproduktif dan kasta steril (Krishna 1969; Lee & Wood 1971).
sayap pada bagian toraks. Sedangkan kasta reproduktif sekunder berfungsi menggantikan kasta reproduktif apabila raja atau ratu mati atau untuk menambah jumlah telur apabila telur yang dihasilkan oleh ratu tidak mencukupi kebutuhan koloni (Krishna 1969).
Kasta steril terdiri atas pekerja dan prajurit. Ciri dari kasta ini adalah tidak adanya sayap dan perkembangan organ seksual ditekan atau tidak berkembang. Pekerja bertanggungjawab untuk mencari makan dan memelihara telur, larva dan ratu. Larva, prajurit dan ratu tidak mampu untuk memberi makan dirinya sendiri sehingga bergantung pada makanan yang diberikan pekerja. Jumlah pekerja mencapai 90% dari seluruh anggota koloni. Rayap prajurit bertugas menjaga koloni dari serangan musuh dan juga menjaga pekerja yang mencari makan di sekitar sarang. Prajurit dibedakan dengan pekerja berdasarkan modifikasi bagian mulut dan kepala yang mengalami kitinasi yang kuat, biasanya terpigmentasi dan seringkali lebih besar daripada ukuran kepala kasta yang lain (Krishna 1969).
Secara umum makanan rayap adalah semua bahan yang mengandung selulosa. Bignell dan Eggleton (2000), membagi rayap menjadi beberapa kelompok berdasarkan jenis makanannya. Pertama, rayap pemakan tanah (
5
feeder) yang mendapatkan makanan dari mineral tanah. Material yang dicerna sangat heterogen, mengandung banyak bahan organik tanah dan silika. Rayap jenis ini ditemukan pada Apicotermitinae, Termitinae, Nasutitermitinae, dan Indotermitinae. Kedua, rayap pemakan kayu (wood-feeder) yang mendapatkan makanan dengan memakan kayu dan sampah berkayu, termasuk cabang mati yang masih menempel di pohon. Hampir semua rayap tingkat rendah adalah pemakan kayu, semua subfamili dari Termitinae kecuali Apicotermitinae. Ketiga, rayap pemakan serasah (litter-feeder). Rayap jenis ini mendapatkan makanan dari daun atau kayu-kayu kecil. Rayap ini terdapat pada Macrotermitinae, Apicotermitinae, Termitinae, dan Nasutitermitinae.
Beberapa jenis rayap yang ada di Indonesia antara lain Neotermes tectonae, Cryptotermes cyanocephalus, C. curvignathus, Schedorhinotermes javanicus, Procapritermes setiger, Pericapritermes semarangi, Macrotermes gilvus, Microtermes insperatus, Microtermes jacobseni, Nasutitermes javanicus, dan Nasutitermes matangensis (Lee & Wood 1971; Harris 1971; Teguh 2009).
Taksonomi dan Biologi C. curvignathus
Coptotermes curvignathus merupakan salah satu rayap subteran. Secara taksonomi C. curvignathus termasuk dalam Klas Insekta, Ordo Isoptera, Famili Rhinotermitidae, Subfamili Coptotermitinae, Genus Coptotermes (Ahmad 1958; Krishna 1969). Berdasarkan analisis gen cytochrome oxidase II (COII) genus Coptotermes terletak diantara genus Reticulitermes dan Nasutitermes (Gambar 2) (Austin et al. 2004).
Rayap C. curvignathus berwarna pucat (Gambar 3a). Prajurit memiliki kepala berbentuk oval, melebar, dan datar dengan fontanel menonjol yang dapat mengeluarkan cairan berwarna putih seperti susu apabila diserang. Eksudat ini berguna untuk melumpuhkan musuhnya. Mandibula berwarna merah kecoklatan dan melengkung di bagian ujungnya. Ukuran prajurit relatif besar (±4.5 mm); lebar kepala 1.34-1.52 mm; antena 14-15 ruas, ruas kedua dua kali panjangnya atau sedikit lebih panjang daripada ruas ketiga (Gambar 3b) (Ahmad 1958).
Gambar 2 Filogeni genus Reticulitermes, Coptotermes, dan Nasutitermes berdasarkan gen cytochrome oxidase II (COII) (Austin et al. 2004).
N
7
Coptotermes curvignathus terdapat di Asia Tenggara seperti Malaysia, Singapura, Indonesia (Jawa, Sumatra, Kalimantan), Thailand, Vietnam, Kamboja, Filipina, dan Cina bagian selatan. Rayap ini termasuk hama yang serius menyerang pohon karet hidup di Asia Tenggara dan menyerang kulit kayu dan bagian lain termasuk bagian dalam kayu, bahkan sering membunuh tanaman. Untuk mencapai makanannya rayap ini membuat saluran bawah tanah dan jika diperlukan mampu membentuk saluran-saluran yang ditutupi tanah yang dibuat di atas tanah (Roonwal 1970).
Simbiosis Pada Saluran Pencernaan Rayap dan Metabolisme Selulosa
Saluran pencernaan rayap terdiri atas usus depan, usus tengah, dan usus belakang (Gambar 4). Saluran pencernaan ini menempati sebagian besar dari abdomen. Usus depan terdiri atas esofagus dan tembolok yang dilengkapi dengan kelenjar saliva. Esofagus dan tembolok memanjang pada bagian posterior atau bagian tengah dari thorak. Kelenjar saliva mensekresikan endoglukanase dan enzim lain ke dalam saluran pencernaan. Usus tengah merupakan bagian yang berbentuk tubular yang mensekresikan suatu membrane peritrofik di sekeliling material makanan. Usus tengah pada rayap tingkat juga diketahui mensekresikan endoglukanase. Usus belakang merupakan tempat bagi sebagian besar simbion (Noirot & Noirot-Timothée 1969; Scharf & Tartar 2008).
Gambar 3 Rayap C. curvignathus (a) pekerja dan (b) prajurit. (a)
1 mm
1 mm
Gambar 4 Morfologi saluran pencernaan rayap (Scharf & Tartar 2008). KS=kelenjar saliva, E=esofagus, T=tembolok, UT=usus tengah, UB=usus belakang.
Rayap bersimbiosis dengan bakteri dan protozoa pada saluran pencernaannnya. Pada rayap tingkat rendah lebih banyak bersimbiosis dengan protozoa dibandingkan dengan bakteri, sebaliknya pada rayap tingkat tinggi lebih banyak bersimbiosis dengan bakteri dibandingkan dengan protozoa (Krishna 1969; Bignell 2000; Breznak 2000).
Protozoa yang bersimbiosis dengan rayap tingkat rendah berbeda pada tiap spesies. Zootermopsis angusticollis bersimbiosis dengan Tricercomitis, Hexamastix, dan Trichomitopsis. Mastotermes darwiniensis bersimbiosis dengan Mixotricha paradoxa (Breznak 2000). Coptotermes formosanus bersimbiosis dengan Pseudotrichonympha grassii, Spirotrichonympha leidy, Holomastigoides mirabile (Inoue et al. 2005; Nakashima et al. 2002b), dan Holomastigoides hartmanni (Tanaka et al. 2006). Coptotermes lacteus bersimbiosis dengan Holomastigoides mirabile (Watanabe et al. 2002). Reticulitermes speratus bersimbiosis dengan Teranympha mirabilis, Triconympha agilis (Ohtoko et al. 2000), Dinenympha exilis dan Pyrsonymphagrandis (Todaka et al. 2007).
Rayap dapat bertahan hidup dengan memanfaatkan lignoselulosa yang mengandung sedikit nutrisi (Ohkuma 2003). Rayap dapat mencerna lignoselulosa dengan menggunakan enzim selulase yang dihasilkan oleh rayap itu sendiri dan simbion (Watanabe et al. 1998; Ohkuma 2003; Scharf & Tartar 2008).
Lignoselulosa adalah suatu campuran dari tiga polimer yang dihasilkan tanaman yaitu selulosa, hemiselulosa, dan lignin. Selulosa adalah polimer glukosa yang terikat oleh ikatan -1,4 dan terikat bersama dengan hemiselulosa (Scharf & Tartar 2008).
UB UT
KS
9
Selulase merupakan enzim ekstraseluler yang terdiri atas kompleks endo-ß-1,4-glukanase (endoglukanase), kompleks ekso-ß-endo-ß-1,4-glukanase (eksoglukanase) yang terdiri dari cellobiohydrolase (EC 3.2.1.91) atau cellodextranase (EC 3.2.1.74), dan ß-glukosidase (EC 3.2.1.21). Endoglukanase aktif dalam menghancurkan kristal atau selulosa murni. Eksoglukanase menghidrolisis selulosa non-kristal atau turunan selulosa terlarut. ß-1,4-glukosidase menghidrolisis selobiosa menjadi glukosa (Wood & Garcia-Campayo 1990).
Endoglukanase rayap termasuk ke dalam GHF 9. Anggota GHF 9 rayap terdiri dari endoglukanase dari rayap tingkat tinggi (Tokuda et al. 1999, 2004; Watanabe & Tokuda 2001) dan rendah (Nakashima et al. 2002b). Anggota GHF 9 famili selulase yang secara taksonomi tersebar luas, termasuk pada tanaman, bakteri, slime mold, dan rayap. Selulase simbion diklasifikasikan menjadi beberapa famili GHF yaitu 3, 5, 7, 8, 10, 11, 26, 43, 45, 62. Selulase simbion dari C. formosanus termasuk anggota GHF 5 (Inoue et al. 2005) dan GHF 7 (Nakashima et al. 2002a). Selulase simbion dari C. lacteus (Watanabe et al. 2002) dan R. flavipes (Zhou et al. 2007) termasuk anggota GHF 7. Selulase simbion dari R. speratus termasuk anggota GHF 3, 5, 7, 8, 10, 11, 26, 43, 45, dan 62 (Ohtoko et al. 2000; Todaka et al. 2007). Selulase simbion dari M. darwiniensis termasuk anggota GHF 45 (Li et al. 2003) dan selulase bakteri simbion Nasutitermes termasuk anggota GHF 11 (Brennan et al. 2004).
Rayap tingkat rendah secara utama memakan kayu yang kaya selulosa kristalin dan amorf, hemiselulosa dan lignin. Tingkat hidrolisis hemiselulosa dan lignin yang tinggi dapat membatasi aktivitas selulolitik melalui penghambatan produk akhir. Hal ini yang menyebabkan rayap mengekspresikan berbagai macam gen selulase. Pada R. speratus dan R. flavipes dihasilkan selulase yang termasuk GHF 7, 9, dan 45 (Ohtoko et al. 2000; Zhou et al. 2007).
Struktur Ekson-Intron dan Ekspresi Gen Endoglukanase Rayap
sepuluh ekson dengan sembilan intron. Semua intron NtEG diawali dengan GT dan diakhiri dengan AG. Pada partial genomik RsEG diketahui ukuran dari tiga intron. Dua intron pada RsEG memiliki posisi yang identik dengan NtEG yaitu intron tujuh dan delapan (Tokuda et al. 1999). Itakura et al. (2006) mengkarakterisasi intron sembilan RsEG sebesar 529 pb. Situs pemotongan intron 10 berbeda dengan situs pemotongan eukariot pada umumnya (GT/AG), yaitu diawali dengan AG dan diakhiri dengan GC.
Data cDNA lengkap terdiri atas R. flavipes (Scharf et al. 2005), R. speratus (Tokuda et al. 1999), C. formosanus (Nakashima et al. 2002b), C. acinaciformis, N. walkeri (Tokuda et al. 1999). Data partial cDNA terdiri atas M. darwiniensis (Tokuda et al. 1999; Li et al. 2003), H. sjoestedti (Tokuda et al. 2004), H. japonica (Lo et al. 2000), Glyptotermes sp. (Tokuda et al. 1999), N. koshunensis (Lo et al. 2000; Tokuda et al. 2004), R. speratus (Watanabe et al. 1998; Itakura et al. 2006), S. mushae, dan O. formosanus (Tokuda et al. 2004).
Endoglukanase rayap diekspresikan pada saluran pencernaan rayap. Terdapat perbedaan ekspresi endoglukanase pada rayap tingkat rendah dan rayap tingkat tinggi. Ekspresi NtEG dan NwEG yang termasuk ke dalam rayap tingkat tinggi terjadi di usus tengah (Tokuda et al. 1999). Sedangkan pada rayap tingkat rendah terjadi perbedaan ekspresi dari tiap spesies. Ekspresi RsEG terjadi di kelenjar saliva/usus depan (Tokuda et al. 1999), CfEG diekspresikan di kelenjar saliva, usus depan, dan usus tengah, dan RfEG terdapat pada bagian kelenjar saliva, usus depan, usus tengah, dan usus belakang (Zhou et al. 2007). RfEG diekspresikan paling tinggi pada pekerja kemudian nimfa. Ekspresi EGase menurun pada laron tetapi meningkat 50% pada kasta reproduktif sekunder (Scharf et al. 2005).
11
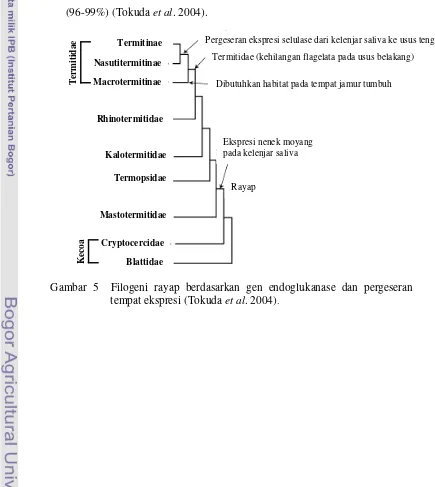
Aktivitas selulase rayap tingkat rendah dan rayap tingkat tinggi berbeda. Pada rayap tingkat rendah M. darwiniensis (Mastotermitidae), H. sjoestedti (Termopsidae), N. koshunensis (Kalotermitidae), R. speratus (Rhinotermitidae), dan C. formosanus (Rhinotermitidae), aktivitas endoglukanase tertinggi ditemukan di kelenjar saliva (45-86%). Secara keseluruhan pada rayap tingkat rendah aktivitas endoglukanase tertinggi pada Kalotermitidae dan Rhinotermitidae. Sedangkan Termitidae yang termasuk rayap tingkat tinggi yaitu O. formosanus (Macrotermitinae), N. takasagoensis (Nasutitermitinae), dan S. mushae (Termitinae) aktivitas endoglukanase tertinggi ditemukan di usus tengah (96-99%) (Tokuda et al. 2004).
Pergeseran ekspresi selulase dari kelenjar saliva ke usus tengah
Termitidae (kehilangan flagelata pada usus belakang)
Dibutuhkan habitat pada tempat jamur tumbuh
Ekspresi nenek moyang pada kelenjar saliva
Rayap
Gambar 5 Filogeni rayap berdasarkan gen endoglukanase dan pergeseran tempat ekspresi (Tokuda et al. 2004).
BAHAN DAN METODE
Waktu dan Tempat
Penelitian berlangsung dari Agustus 2009–Juli 2010. Koleksi sampel dilakukan di lingkungan kampus IPB Darmaga Kabupaten Bogor. Identifikasi, analisis molekular dan pengolahan data dilakukan di Bagian Fungsi dan Perilaku Hewan dan Laboratorium Terpadu, Departemen Biologi, FMIPA, IPB.
Bahan dan Alat
Bahan dan alat untuk ekstraksi DNA terdiri atas Cetyl Trimetyl Amonium Bromide (CTAB 0.2% (b/v) (7.5 ml 1M Tris-HCl pH 8; 3 ml 0.5 M NaEDTA pH 8; 6.135 g NaCl; 1.5 g CTAB; air hingga 75 ml)), proteinase K (5 mg/ml), PCI (Phenol : Chloroform : Isoamyl alcohol = 25 : 24 : 1), CIAA (Chloroform : Isoamyl alcohol = 24 : 1), fenol, isopropanol, etanol 70% (v/v), Tris-HCl EDTA ((TE) (10 mM Tris-HCl-EDTA pH 8; 1 mM EDTA)), nitrogen cair, grinder, pinset, scalpel, dan tabung 1.5 ml. Bahan untuk campuran PCR yaitu akuades steril, primer forward, primer reverse, dan KAPA Taq ReadyMix DNA Polymerase (bufer dengan 1.5 mM Mg2+, dNTP 0.4 mM, 0.05 U/µl KapaTaq DNA polymerase). Bahan yang digunakan untuk elektroforesis adalah medium polyacrylamide gel electrophoresis (PAGE) 6% (12 ml akuades; 4 ml acrylamide 30%; 5xTBE 4 ml; TEMED (tetramethylethylene-diamine) 15 µl; 10% APS (Amonium Peroxo Sulfat) 150 µl), bufer 1xTBE (Tris 0.5 M, asam borat 0.65 M, EDTA 0.02M), penanda 100 pb DNA ladder (Promega), dan 6x loading dye (Promega). Visualisasi DNA menggunakan CTAB (0.2 g/200 ml akuades), NH4OH (2.4 ml/200 ml akuades), AgNO3 (0.32 g/200 ml akuades), Na2CO3 (4 g/200 ml akuades), NaOH (4 g/10 ml akuades), formalin, dan asam asetat (200 µl/200 ml akuades).
Metode
Koleksi Coptotermes curvignathus
13
IPB Darmaga, Bogor. Rayap yang didapatkan dimasukkan ke dalam tabung 1.5 ml yang berisi alkohol absolut, kemudian dilakukan penggantian alkohol absolut sebanyak dua kali. Sampel yang didapatkan diidentifikasi berdasarkan Triplehorn & Johnson (2005) sampai dengan tingkat famili dan Ahmad (1958) untuk tingkat genus dan spesies.
Ekstraksi DNA
Ekstraksi DNA menggunakan metode ekstraksi CTAB 0.2% dan presipitasi alkohol (Sambrook et al. 1989) yang telah dimodifikasi. Sebelum ekstraksi, rayap dimasukkan ke dalam TE (10 mM Tris-HCl EDTA pH 8; 1 mM EDTA) untuk menghilangkan etanol dari jaringan. Penggantian TE dilakukan sebanyak tiga kali. Jaringan sumber DNA yang digunakan adalah kepala dan toraks rayap pekerja.
Kepala dan toraks rayap dipisahkan dari bagian abdomen dengan menggunakan scalpel. Tabung berisi satu kepala dan satu toraks rayap dimasukkan ke dalam nitrogen cair selama 15 menit, kemudian toraks digerus sampai hancur. Bufer CTAB 0.2% ditambahkan sebanyak 200 µl ke dalam tabung yang berisi hancuran kepala dan toraks. Proteinase K (5 mg/ml) ditambahkan sebanyak 14 µl untuk mendegradasi protein, kemudian diinkubasi pada suhu 55 0
C selama 2 jam. Setelah diinkubasi, tabung beserta isinya disentrifugasi 13000 rpm selama 10 menit. Supernatan dipindahkan ke tabung yang baru, kemudian ditambahkan larutan PCI (Phenol : Chloroform : Isoamyl alcohol = 25 : 24 : 1), sebanyak 250 µl, digoyang dengan tangan perlahan selama 5 menit lalu disentrifugasi 13000 rpm selama 5 menit. Lapisan atas yang berisi DNA dipindahkan ke tabung yang baru, kemudian ditambahkan larutan CIAA (Chloroform : Isoamyl alcohol = 24 : 1) sebanyak 200 µl, digoyang dengan tangan perlahan selama 5 menit dan disentrifugasi 13000 rpm selama 3 menit. Tahapan ini dilakukan dua kali ulangan.
dengan cara divakum selama 30 menit. Pelet DNA kemudian disuspensikan dalam TE 0.5 mM sebanyak 15-20 µl dan disimpan pada suhu –4 0C.
Amplifikasi DNA
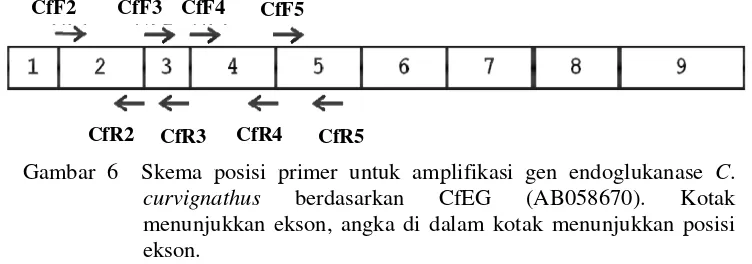
Amplifikasi DNA hasil ekstraksi dilakukan secara in vitro dengan teknik PCR (Polymerase Chain Reaction) menggunakan mesin thermocycler (ESCO SWIFT MAXI-BLC1). Segmen DNA yang diamplifikasi adalah gen endoglukanase C. curvignathus. Amplifikasi DNA target diamplifikasi dengan menggunakan beberapa pasang primer (Tabel 1, Gambar 6). Primer disusun secara manual berdasarkan CfEG nomor aksesi Genbank AB058670 dan NtEG nomor aksesi Genbank AB019146 dan verifikasi menggunakan program Primer3 (Rozen & Skaletsky 2000; Koressaar & Remm 2007). Primer disusun menggunakan template dari CfEG yang merupakan endoglukanase yang berasal dari kelenjar saliva dan perkiraan posisi intron berdasarkan NtEG.
Total pereaksi yang digunakan adalah sebanyak 20 µl, terdiri atas 7.4 µl akuades steril, 10 µl KAPA Taq ReadyMix DNA Polymerase (bufer dengan 1.5 mM Mg2+, dNTP 0.4 mM, 0.05 U/µl KapaTaq DNA polymerase), 0.8 µl primer forward 10 µM, 0.8 µl primer reverse 10 µM, dan1 µl DNA hasil ekstraksi.
Proses amplifikasi dilakukan pada kondisi suhu pra-denaturasi 94 0C selama 5 menit, dilanjutkan 30 siklus yang terdiri atas tahap denaturasi DNA 94 0C selama 1 menit, penempelan primer pada suhu 53 0C (pasangan primer CfF2 dan CfR3), 58 0C (pasangan primer CfF3 dan CfR4, CfF4 dan CfR5) selama 1 menit dan sintesis DNA ruas target pada suhu 72 0C selama 2 menit. Proses diakhiri dengan sintesis DNA akhir pada suhu 72 0C selama 7 menit.
Tabel 1 Primer yang digunakan untuk mengamplifikasi DNA target gen endoglukanase C. curvignathus
Ekson Primer Urutan basa (5’-3’) Posisi berdasarkan cDNA C.
15
Elektroforesis dan Pewarnaan DNA
Segmen DNA hasil amplifikasi dipisahkan dengan PAGE 6% menggunakan bufer 1xTBE. Pemisahan dilakukan pada tegangan 200 V selama 50 menit. Visualisasi DNA pada gel poliakrilamid menggunakan metode pewarnaan perak (Tegelstrom 1986) di dalam shaking bath.
Setelah dielektroforesis gel dicuci dengan larutan CTAB selama 8 menit. Kemudian dicuci dengan akuades sebanyak dua kali masing-masing selama 2 menit. Selanjutnya gel tersebut direndam dalam larutan NH4OH selama 8 menit. Kemudian gel direndam dalam larutan perak (0.32 g AgNO3; 0.8 ml NH4OH; 80 µl NaOH; akuades 200 ml) selama 10 menit. Setelah proses ini gel dicuci kembali dengan akuades sebanyak tiga kali, masing-masing selama 2 menit. Untuk tahap pemunculan pita DNA, gel direndam dalam larutan Na2CO3 (4 g Na2CO3; 100 µl formalin; 200 ml akuades). Tahap terakhir adalah proses perendaman gel pada larutan asam asetat 1%.
Sekuen dan Alignment DNA dan Asam Amino
Proses sekuen DNA menggunakan jasa sekuensing. Produk sekuen diedit menggunakan program Genetyx Win versi 4.0, kemudian dilakukan alignment menggunakan Clustal X (Thompson et al. 1997) dan Mega 4.0 (Tamura et al. 2007). Alignment DNA dilakukan antara sekuen gen endoglukanase C. curvignathus dengan sekuen CfEG (AB058670) dan NtEG (AB019146) dari GenBank (www.ncbi.nlm.nih.gov). Analisis homologi dilakukan melalui BLASTN untuk urutan basa dan BLASTP untuk urutan asam amino hasil translasi melalui situs NCBI (www.ncbi.nlm.nih.gov).
Gambar 6 Skema posisi primer untuk amplifikasi gen endoglukanase C. curvignathus berdasarkan CfEG (AB058670). Kotak menunjukkan ekson, angka di dalam kotak menunjukkan posisi ekson.
CfF2 CfF3 CfF4 CfF5
CfR4 CfR3
Analisis Filogeni dan Jarak Genetik
Analisis filogeni menggunakan program Mega 4.0 (Tamura et al. 2007). Analisis filogeni berdasarkan asam amino dengan menyamakan awal dan akhir asam amino terlebih dahulu berdasarkan hasil alignment. Rekonstruksi filogeni menggunakan metode Maximum Parsimony dengan bootstrap 1000 kali. Data endoglukanase bakteri, protozoa simbion rayap, fungi, rayap, echinodermata, cacing tanah, dan moluska (Tabel 2) diambil dari genbank (www.ncbi.nlm.nih.gov). Perhitungan jarak genetik dilakukan antara gen endoglukanase C. curvignathus dengan gen endoglukanase rayap lain dengan program MEGA 4.0 (Tamura et al. 2007).
Analisis Protein Putative
Analisis motif protein putative (misal: protein signature) gen endoglukanase C. curvignathus menggunakan data yang ada pada Prosite (Falquet et al. 2002, www.expasy.org/prosite), Nakashima et al. (2002b), dan Zhou et al. (2007). Analisis gen famili dari protein putative gen endoglukanase C. curvignathus menggunakan Pfam (Protein family, Bateman et al. 1999, pfam.sanger.ac.uk). Tabel 2 Data endoglukanase pembanding dari GenBank
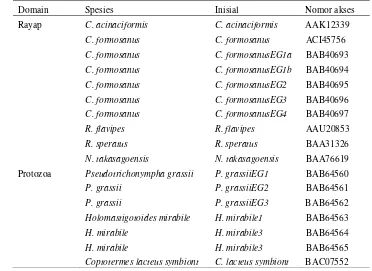
Domain Spesies Inisial Nomor akses
Rayap C. acinaciformis C. acinaciformis AAK12339
C. formosanus C. formosanus ACI45756
C. formosanus C. formosanusEG1a BAB40693
C. formosanus C. formosanusEG1b BAB40694
C. formosanus C. formosanusEG2 BAB40695
C. formosanus C. formosanusEG3 BAB40696
C. formosanus C. formosanusEG4 BAB40697
R. flavipes R. flavipes AAU20853
R. speratus R. speratus BAA31326
N. takasagoensis N. takasagoensis BAA76619
Protozoa Pseudotrichonympha grassii P. grassiiEG1 BAB64560
P. grassii P. grassiiEG2 BAB64561
P. grassii P. grassiiEG3 BAB64562
Holomastigotoides mirabile H. mirabile1 BAB64563
H. mirabile H. mirabile3 BAB64564
H. mirabile H. mirabile3 BAB64565
17
Tabel 2 Lanjutan
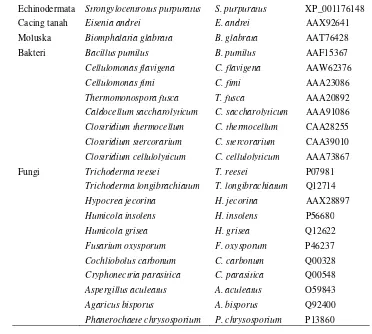
Echinodermata Strongylocentrotus purpuratus S. purpuratus XP_001176148
Cacing tanah Eisenia andrei E. andrei AAX92641
Moluska Biomphalaria glabrata B. glabrata AAT76428
Bakteri Bacillus pumilus B. pumilus AAF15367
Cellulomonas flavigena C. flavigena AAW62376
Cellulomonas fimi C. fimi AAA23086
Thermomonospora fusca T. fusca AAA20892
Caldocellum saccharolyticum C. saccharolyticum AAA91086
Clostridium thermocellum C. thermocellum CAA28255
Clostridium stercorarium C. stercorarium CAA39010
Clostridium cellulolyticum C. cellulolyticum AAA73867
Fungi Trichoderma reesei T. reesei P07981
Trichoderma longibrachiatum T. longibrachiatum Q12714
Hypocrea jecorina H. jecorina AAX28897
Humicola insolens H. insolens P56680
Humicola grisea H. grisea Q12622
Fusarium oxysporum F. oxysporum P46237
Cochliobolus carbonum C. carbonum Q00328
Cryphonectria parasitica C. parasitica Q00548
Aspergillus aculeatus A. aculeatus O59843
Agaricus bisporus A. bisporus Q92400
HASIL
Amplifikasi DNA Gen Endoglukanase C. curvignathus
Pita DNA gen endoglukanase C. curvignathus hasil amplifikasi menggunakan pasangan primer CfF2 dan CfR3 menghasilkan dua pita yaitu pita tebal berukuran sekitar 900 pasang basa (pb) (Gambar 7a) dan pita tipis yang berukuran sekitar 2000 pb. Berdasarkan panjang ekson CfEG (AB058670) yang berukuran 281 pb dan ketebalan pita DNA, target yang benar adalah pita yang berukuran 900 pb yang selanjutnya dilakukan sekuen DNA. Pasangan primer CfF3 dan CfR4 menghasilkan pita DNA sekitar 1100 pb dan pasangan primer CfF4 dan CfR5 sekitar 500 pb (Gambar 7b).
Sekuen dan Alignment DNA dan Asam Amino Gen Endoglukanase C.
curvignathus
Hasil sekuen DNA gen endoglukanase C. curvignathus dengan menggunakan pasangan primer CfF2 dan CfR3, CfF3 dan CfR4, CfF4 dan CfR5 masing-masing sebesar 856, 1077, dan 484 pb (Gambar 8, Lampiran 1-6).
a. Hasil sekuen DNA gen endoglukanase C. curvignathus dengan menggunakan pasangan primer CfF2 dan CfR3
19
b. Hasil sekuen DNA gen endoglukanase C. curvignathus dengan menggunakan pasangan primer CfF3 dan CfR4
c. Hasil sekuen DNA gen endoglukanase C. curvignathus dengan menggunakan pasangan primer CfF4 dan CfR5
Gambar 8 Hasil sekuen gen endoglukanase C. curvignathus dengan
menggunakan pasangan primer (a) CfF2 dan CfR3, (b) CfF3 dan CfR4, dan (c) CfF4 dan CfR5. Basa yang digaris bawahi menunjukkan posisi primer.
endoglukanase C. curvignathus berada pada intron tiga sampai lima pada NtEG (Gambar 10).
Panjang ekson tiga dan empat gen endoglukanase C. curvignathus sama dengan ekson empat dan lima NtEG serta ekson tiga dan empat CfEG. Sedangkan panjang ekson dua dan lima gen endoglukanase C. curvignathus berbeda dengan panjang ekson tiga dan enam NtEG dan ekson dua dan lima CfEG. Perbedaan panjang juga terdapat pada masing-masing intron gen endoglukanase C. curvignathus dengan intron NtEG (Gambar 10).
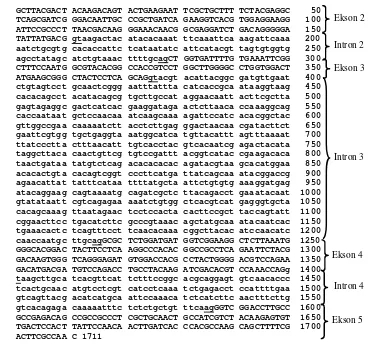
Gambar 9 Sekuen DNA gen endoglukanase C. curvignathus. Huruf besar menunjukkan urutan basa ekson dan huruf kecil menunjukkan urutan basa intron. Basa yang digaris bawahi menunjukkan awal dan akhir intron.
Gambar 10 Skema posisi ekson dan intron NtEG (AB019146), CfEG (AB058670), dan gen endoglukanase C. curvignathus (hasil penelitian ini). Kotak menunjukkan ekson dan segitiga menunjukkan intron. Angka di bawah kotak menunjukkan posisi ekson dan angka di atas segitiga menunjukkan posisi intron. Angka di dalam kotak dan segitiga menunjukkan panjang basa.
Intron dua sampai empat gen endoglukanase C. curvignathus didominasi oleh basa AT yaitu masing-masing sebesar 66.06, 58.81, dan 55.96%. Intron dua sampai empat gen endoglukanase C. curvignathus diawali dengan basa GT dan diakhiri dengan basa AG (Gambar 9). Kesamaan intron dua sampai empat gen endoglukanase C. curvignathus dengan intron tiga sampai lima NtEG masing-masing sebesar 8.8, 24.8, dan 46.5%.
Panjang ekson dua sampai lima gen endoglukanase C. curvignathus yaitu sebesar 564 pb. Hasil analisis BLASTN ekson dua sampai lima gen endoglukanase C. curvignathus homolog dengan CfEG dengan nomor akses EU853671 (95%) dengan E-value 0.0 (Gambar 11, Lampiran 7). Kandungan GC ekson dua sampai lima gen endoglukanase C. curvignathus adalah 55.82%.
Gambar 11 Hasil analisis BLASTN gen endoglukanase C. curvignathus dengan CfEG (EU853671).
C. curvignathus 1 GCTTACGACTACAAGACAGTACTGAAGAATTCGCTGCTTTTCTACGAGGCTCAGCGATCG 60
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| CfEG EU853671 82 GCTTACGACTACAAGACAGTACTGAAGAATTCGCTGCTTTTCTACGAGGCTCAGCGATCG 141
C. curvignathus 61 GGACAATTGCCCGCTGATCAGAAGGTCACGTGGAGGAAGGATTCCGCCCTTAACGACAAG 120
||| |||||||||||||||||||||||||||||||||||||||||||||||||||||||| CfEG EU853671 142 GGAAAATTGCCCGCTGATCAGAAGGTCACGTGGAGGAAGGATTCCGCCCTTAACGACAAG 201
C. curvignathus 121 GGAAACAACGGCGAGGATCTGACAGGGGGATATTATGACGCTGGTGATTTTGTGAAATTC 180
|| | || |||||||| |||||||||||||| ||||||||||||||||||||||| ||| CfEG EU853671 202 GGCCAGAAGGGCGAGGACCTGACAGGGGGATACTATGACGCTGGTGATTTTGTGAAGTTC 261
C. curvignathus 181 GGCTTTCCAATGGCGTACACGGCCACCGTCCTGGCTTGGGGCCTGGTGGACTATGAAGCG 240
||||| || ||||||||||| | ||||||||||||||||||||| || ||||| ||| | CfEG EU853671 262 GGCTTCCCTATGGCGTACACCGTCACCGTCCTGGCTTGGGGCCTTGTAGACTACGAATC- 320
C. curvignathus 241 GGC-TACTCCT-CAGCAGGCGCTCTGGATGATGGTCGGAAGGCTCTTAAATGGGGCACGG 298
||| || | || ||||||||||||||||||||||||| |||||||||||||||||||||| CfEG EU853671 321 GGCGTA-TTCTACAGCAGGCGCTCTGGATGATGGTCGCAAGGCTCTTAAATGGGGCACGG 379
C. curvignathus 299 ACTACTTCCTCAAGGCCCACACGGCCGCCTCA-GAATTCTACGGACAAGTGGGTCAGGGA 357
||||||||||||||||||||||||||||| | ||||||||||||||||||||||||||| CfEG EU853671 380 ACTACTTCCTCAAGGCCCACACGGCCGCC-AATGAATTCTACGGACAAGTGGGTCAGGGA 438
C. curvignathus 358 GATGTGGACCACGCCTACTGGGGACGTCCAGAAGACATGACGATGTCCAGACCTGCCTAC 417
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| CfEG EU853671 439 GATGTGGACCACGCCTACTGGGGACGTCCAGAAGACATGACGATGTCCAGACCTGCCTAC 498
C. curvignathus 418 AAGATCGACACGTCCAAACCAGGGTCGGACCTTGCCGCCGAGACAGCCGCCGCCCTCGCT 477
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||| CfEG EU853671 499 AAGATCGACACGTCCAAACCAGGGTCGGACCTTGCCGCCGAGACAGCCGCCGCCCTCGCT 558
C. curvignathus 478 GCAACTGCCATCGTCTACAAGAGTGTTGACTCCACTTATTCCAACAACTTGATCACCCAC 537
||||||||||||| ||||||||||| |||||||||||||||||||||||||||||||||| CfEG EU853671 559 GCAACTGCCATCGCCTACAAGAGTGCTGACTCCACTTATTCCAACAACTTGATCACCCAC 618
C. curvignathus 538 GCCAAGCAGCTTTTCGACTTCGCCAAC 564
23
Sekuen ekson dua sampai lima gen endoglukanase C. curvignathus kemudian ditranslasi menggunakan program Genetyx Win versi 4.0. Asam amino putative yang didapatkan sebesar 188 asam amino (Gambar 12). Hasil analisis homologi asam amino putative gen endoglukanase C. curvignathus menggunakan BLASTP homolog (94%) dengan CfEG (BAB40696) dengan E-value sebesar 9e-92 (Gambar 13, Lampiran 8). Perbedaan asam amino antara asam amino putative gen endoglukanase C. curvignathus dengan CfEG (BAB40696) sebanyak sembilan asam amino (Gambar 13).
Gambar 13 Hasil analisis BLASTP asam amino putative gen endoglukanase C. curvignathus dengan gen CfEG (BAB40696). Kotak menunjukkan perbedaan asam amino.
Analisis Filogeni dan Jarak Genetik Gen Endoglukanase C. curvignathus
Hasil analisis filogeni dengan menggunakan asam amino menunjukkan gen endoglukanase C. curvignathus mengelompok dengan GHF 9 pada kelompok rayap dan terpisah dengan GHF 9 pada kelompok moluska, echinodermata, cacing tanah, dan bakteri. Gen endoglukanase C. curvignathus juga terpisah dengan GHF 7 pada kelompok fungi dan protozoa (Gambar 14).
25
Tabel 3 Jarak genetik gen endoglukanase C. curvignathus dengan gen endoglukanase C. acinaciformis, C. formosanus, N. takasagoensis , R. flavipes, dan R. speratus
1 2 3 4 5 6 7 8 9 10 11
1
2 0.000
3 0.000 0.000
4 0.005 0.005 0.005
5 0.005 0.005 0.005 0.000
6 0.005 0.005 0.005 0.000 0.000
7 0.049 0.049 0.049 0.043 0.043 0.043
8 0.049 0.049 0.049 0.043 0.043 0.043 0.000
9 0.060 0.060 0.060 0.055 0.055 0.055 0.095 0.095
10 0.199 0.199 0.199 0.193 0.193 0.193 0.239 0.239 0.149
11 0.213 0.213 0.213 0.206 0.206 0.206 0.226 0.226 0.174 0.187
Keterangan: (1) C. formosanus EG1a, (2) C. formosanus EG1b, (3) C. formosanus EG2, (4) C.
formosanus EG3, (5) C. formosanus, (6) C. formosanus EG4, (7) R. flavipes, (8) R. speratus, (9)
C. curvignathus, (10) C. acinaciformis, (11) N. takasagoensis. Kotak menunjukkan jarak genetik terbesar dan terkecil.
Gambar 14 Pohon filogeni gen endoglukanase C. curvignathus berdasarkan asam amino putative menggunakan metode Maximum Parsimony (bootstrap 1000 kali).
GHF 9
Analisis Protein Putative Gen Endoglukanase C. curvignathus
Berdasarkan Prosite, Nakashima et al. (2002b), dan Zhou et al. (2007) protein putative gen endoglukanase C. curvignathus menunjukkan adanya beberapa motif yaitu satu consensus signature GHF 9, nucleophile, co-nucleophile, salt bridge, dan N-terminal (Gambar 15). Pencarian dengan Pfam menunjukkan endoglukanase C. curvignathus masuk ke dalam GHF 9 yang termasuk ke dalam CL0059 yaitu six-hairpin glycosidase superfamily.
Gambar 15 Analisis protein putative gen endoglukanase C. curvignathus. = N-terminal, __ = consensus signature GHF 9, o = nucleophile, * = co-nucleophile, # = salt bridge.
o *
CfEG1 (BAB40693) MRVFFCLLSALALCQAAYDYKTVLKNSLLFYEAQRSGKLPADKKVTWRKDSALNDKGQKG 60 CfEG3 (BAB40696) MRVFFCLLSALALCQAAYDYKTVLKNSLLFYEAQRSGKLPADQKVTWRKDSALNDKGQKG 60
C. curvignathus ---AYDYKTVLKNSLLFYEAQRSGQLPADQKVTWRKDSALNDKGNNG 44
CfEG1 (BAB40693) EDLTGGYYDAGDFVKFGFPMAYTVTVLAWGLVDYESAYSTAGALDDGRKALKWGTDYFLK 120 CfEG3 (BAB40696) EDLTGGYYDAGDFVKFGFPMAYTVTVLAWGLVDYESAYSTAGALDDGRKALKWGTDYFLK 120
C. curvignathus EDLTGGYYDAGDFVKFGFPMAYTATVLAWGLVDYEAGYSSAGALDDGRKALKWGTDYFLK 104
CfEG1 (BAB40693) AHTAANEFYGQVGQGDVDHAYWGRPEDMTMSRPAYKIDTSKPGSDLAAETAAALAATAIA 180 CfEG3 (BAB40696) AHTAANEFYGQVGQGDVDHAYWGRPEDMTMSRPAYKIDTSKPGSDLAAETAAALAATAIA 180
C. curvignathus AHTAASEFYGQVGQGDVDHAYWGRPEDMTMSRPAYKIDTSKPGSDLAAETAAALAATAIV 164
CfEG1 (BAB40693) YKSADSTYSNNLITHAKQLFDFANNYRGKYSDSITDAKNFYASGDYKDELVWAAAWLYRA 240 CfEG3 (BAB40696) YKSADSTYSNNLITHAKQLFDFANNYRGKYSDSITDAKNFYASGDYKDELVWAAAWLYRA 240
C. curvignathus YKSVDSTYSNNLITHAKQLFDFAN--- 188
PEMBAHASAN
Amplifikasi DNA Gen Endoglukanase C. curvignathus
Tiga pasang primer (CfF2-CfR3, CfF3-CfR4, CfF4-CfR5) dari sepuluh pasang kombinasi primer yang disusun (CfF2-CfR2, CfF2-CfR3, CfF2-CfR4, CfF2-CfR5, CfF3-CfR3, CfF3-CfR4, CfF3-CfR5, CfF4-CfR4, CfF4-CfR5, CfF5-CfR5) berhasil mengamplifikasi empat ekson gen endoglukanase C. curvignathus. Walaupun hanya tiga pasang primer yang berhasil mengamplifikasi gen endoglukanase C. curvignathus, sudah mencakup semua ekson target.
Tujuh kombinasi primer yang tidak berhasil mengamplifikasi gen endoglukanase C. curvignathus mungkin disebabkan karena primer yang disusun berdasarkan CfEG tidak cocok sehingga tidak menempel pada target yaitu gen endoglukanase C. curvignathus. Ketidakcocokan ini karena adanya perbedaan nukleotida primer dengan gen endoglukanase C. curvignathus yaitu pada primer CfR2 (Gambar 8a). Primer dapat bekerja dengan baik apabila sekuen nukleotida primer cocok dengan lokasi gen target yang akan saling berkomplemen (Hillis et al. 1996). Daerah yang digunakan sebagai template adalah daerah yang conserve dari gen target yang akan mendukung kecocokan dari primer (Birt & Baker 2000). Kemungkinan yang lain adalah perbedaan suhu penempelan primer yang berbeda jauh antara primer forward dan reverse (3-17 0C) serta adanya struktur hairpin dan dimer. Penentuan suhu penempelan primer adalah langkah yang paling penting di dalam proses PCR (Koressaar & Remm 2007). Struktur sekunder pada primer dapat menyebabkan masalah selama proses PCR, sehingga perlu dihindari sekuen yang saling berkomplemen di dalam primer (hairpin) atau sekuen yang saling berkomplemen antara primer forward dan reverse (primer-dimer) itu sendiri (Newton & Graham 1997; McDowell 1999; Birt 2000; Rapley 2000).
Homologi DNA dan Asam Amino Putative Gen Endoglukanase C.
curvignathus dan C. formosanus
Berdasarkan analisis homologi diketahui bahwa ekson dua sampai lima gen endoglukanase C. curvignathus yang berhasil dikarakterisasi juga terdapat pada CfEG. Persentase homologi antara gen endoglukanase C. curvignathus dengan CfEG (EU853671) sebesar 95%. Asam amino putative gen endoglukanase C. curvignathus memiliki homologi dengan asam amino CfEG. Homologi asam amino putative gen endoglukanase C. curvignathus dengan asam amino CfEG (BAB40696) sebesar 94%.
Semakin tinggi nilai homologi menunjukkan bahwa sekuen gen endoglukanase C. curvignathus terhadap CfEG semakin tepat. Dua fragmen DNA dikatakan homolog jika 70% sekuen basa atau 25% sekuen asam amino identik (panjang sekuen minimal 100 pasang basa) (Claviere & Notredame 2003).
Nilai probabilitas atau peluang yang terhitung secara statistik dalam kesamaan sekuen antara gen endoglukanase C. curvignathus dan CfEG di GenBank (www.ncbi.nlm.nih.gov) digambarkan dengan nilai Expectation value (E-value). Ekson dua sampai lima gen endoglukanase C. curvignathus memperlihatkan nilai E-value yang rendah dibandingkan CfEG yaitu 0.0. Nilai E-value yang rendah juga terlihat pada asam amino putative gen endoglukanase C. curvignathus yaitu sebesar 9e-92. Hasil BLASTP asam amino putative gen endoglukanase C. curvignathus ini signifikan. Pada analisis melalui BLAST, E-value signifikan apabila nilainya 1e-10 atau lebih kecil (Altschul et al. 1990).
29
Analisis Filogeni dan Jarak Genetik Gen Endoglukanase C. curvignathus
Analisis filogeni membandingkan 188 asam amino gen endoglukanase C. curvignathus dengan sekuen endoglukanase fungi, protozoa simbion rayap, bakteri, echinodermata, cacing tanah, dan moluska, didapatkan dua kelompok terpisah (Gambar 14). Pertama adalah kelompok GHF 9 yang terdiri atas rayap, echinodermata, cacing tanah, dan moluska. Kelompok GHF 9 terbagi menjadi dua sister group yaitu kelompok rayap dan kelompok bakteri, echinodermata, cacing tanah, dan moluska. Hal ini didukung dengan nilai bootstrap yang tinggi (100%). Kedua adalah kelompok GHF 7 yang terdiri dari fungi dan protozoa. Hal ini didukung oleh Davison & Blaxter (2005) dan Zhou et al. (2007), analisis filogeni sekuen selulase dari fungi, protozoa simbion rayap, eubakteria, dan metazoan mengelompokkan taksa yang berbeda menjadi dua kelompok yaitu GHF 9 dan GHF 7. Kelompok GHF 9 terdiri atas endoglukanase metazoa (Annelida, Arthropoda, Chordata, Echinodermata, dan Moluska) dan eubakteria, sedangkan kelompok GHF 7 terdiri atas endoglukanase fungi dan protozoa. Hal ini menunjukkan bahwa selulase protozoa simbion rayap berbeda dengan selulase pada rayap.
Gen endoglukanase C. curvignathus termasuk ke dalam kelompok GHF 9. Endoglukanase rayap termasuk ke dalam GHF9. Anggota GHF 9 rayap terdiri dari endoglukanase rayap tingkat tinggi (Tokuda et al. 1999, 2004; Watanabe & Tokuda 2001) dan rendah (Nakashima et al. 2002b). Berdasarkan analisis DNA mitokondria, rayap tingkat rendah (Mastotermitidae, Hodotermitidae, Termopsidae, Kalotermitidae, Serritermitidae, Rhinotermitidae) berada pada basal dan lebih primitif dibandingkan dengan rayap tingkat tinggi (Termitidae) yang lebih maju (Kambhampati & Eggleton 2000; Austin et al. 2004). Coptotermes curvignathus dan C. formosanus termasuk ke dalam rayap tingkat rendah, sedangkan N. takasagoensis termasuk ke dalam rayap tingkat tinggi.
genetik maka semakin dekat kekerabatan antar taksa yang dibandingkan dan semakin besar nilai jarak genetik maka semakin jauh kekerabatannya (Nei & Kumar 2000).
Coptotermes curvignathus dan C. formosanus termasuk ke dalam famili Rhinotermitidae, sedangkan N. takasagoensis termasuk ke dalam famili Termitidae. Famili yang berbeda pada rayap memiliki perbedaan perilaku, misalnya habitat dan jenis makanan. Perilaku merupakan respon hewan terhadap lingkungan dan melibatkan beberapa gen (Sokolowski 2001). Lingkungan dapat memberikan dampak terhadap keragaman genetik (Freeland 2005; Hughes et al. 2008). Habitat dan jenis makanan C. curvignathus lebih mirip dengan C. formosanus dibandingkan dengan N. takasagoensis. Enzim yang dihasilkan pada saluran pencernaan berhubungan dengan jenis makanan rayap. Salah satu enzim yang terlibat dalam pencernaan rayap adalah gen endoglukanase. Hal ini menyebabkan C. curvignathus lebih dekat dengan C. formosanus dibandingkan dengan N. takasagoensis.
Analisis Protein Putative dan Homologi Ekspresi Gen Endoglukanase C.
curvignathus dengan C. formosanus
Alignment asam amino gen endoglukanase C. curvignathus, CfEG1 dan CfEG3 menunjukkan berada pada famili yang sama. Motif protein putative gen endoglukanase C. curvignathus menunjukkan adanya satu consensus signature GHF 9, nucleophile, co-nucleophile, salt bridge, dan N-terminal. Motif ini juga terdapat pada endoglukanase rayap yang lain (Watanabe & Tokuda 2001; Nakashima et al. 2002b). Hal ini menguatkan bahwa gen endoglukanase C. curvignathus termasuk ke dalam GHF 9.
31
Pada alignment asam amino ekson satu sampai empat gen endoglukanase C. curvignathus hanya didapatkan satu consensus signature GHF 9 yaitu {G-[WY]-[YFH]-D-A-G-D-[HNY]-[VLG]-[KRM]-[FY]} dan belum ditemukan posisi donor proton dari pusat katalitik. Pada CfEG dan NtEG memiliki tiga consensus signature GHF 9 dan adanya residu donor proton dari pusat katalitik yaitu Glu 411 pada CfEG (Nakashima et al. 2002b) dan Glu412 pada NtEG (Khademi et al. 2002). Hal ini menunjukkan bahwa gen endoglukanase C. curvignathus belum terkarakterisasi secara utuh.
Gen endoglukanase C. curvignathus masuk ke dalam famili gen GHF 9 yang termasuk ke dalam CL0059 yaitu six-hairpin glycosidase superfamily. S ix-hairpin glycosidase superfamily terdiri dari 20 famili termasuk famili GHF 9. Anggota dari kelompok ini memiliki struktur yang umum yaitu terdiri dari enam hairpin yang berbentuk heliks. Hampir semua anggota six-hairpin glycosidase superfamily adalah enzim glycosyl hydrolase (Bateman et al. 1999).
Kelenjar saliva pada rayap memiliki struktur umum yang sama dengan Orthoptera dan Dictyoptera. Masing-masing kelenjar terdiri atas beberapa lobus atau acinus dihubungkan dengan saluran saliva yang terbuka pada bagian dasar labium (Noirot 1969). Kelenjar saliva terletak pada esofagus dan tembolok yang berada pada bagian toraks. Kelenjar saliva mensekresikan endoglukanase dan enzim lain ke dalam saluran pencernaan (Noirot & Noirot-Timothée 1969; Scharf & Tartar 2008). Saliva berfungsi untuk melubrikasi bagian mulut dan lebih banyak dihasilkan apabila makanan yang dicerna kering. Saliva juga mengandung enzim yang mengawali pencernaan makanan. Adanya beberapa enzim pada saliva berhubungan dengan makanan yang dicerna (Chapman 1998).
Pada rayap tingkat rendah, RsEG diekspresikan hanya di kelenjar saliva/usus depan dan CfEG diekspresikan di kelenjar saliva dan usus tengah (Lo et al. 2000; Nakashima et al. 2002b). Pada rayap tingkat tinggi, NtEG dan NwEG diekspresikan di usus tengah (Tokuda et al. 1999). Taksa rayap dari keturunan basal (Mastotermitidae, Termopsidae, Kalotermitidae, Rhinotermitidae) secara konsisten mengekspresikan endoglukanase secara spesifik di kelenjar saliva sedangkan taksa yang lebih maju (Termitidae) hanya pada usus tengah (Gambar 5). Hal ini menunjukkan bahwa ekspresi pada kelenjar saliva merupakan kondisi primitif kemudian terjadi pergeseran selama proses evolusi (Tokuda et al. 2004).
33
berbeda (Tokuda et al. 2004). Hampir semua rayap tingkat rendah adalah pemakan kayu, sedangkan pada famili Termitidae selain memakan kayu sebagian besar memakan serasah dan tanah (Bignell & Eggleton 2000). Famili Nasutitermitinae dan Termitinae merupakan subfamili dari Termitidae yang memakan tanah dan serasah dimana mengandung sedikit selulosa. Hal ini menyebabkan subfamili ini mensekresikan endoglukanase hanya pada usus tengah (Tokuda et al. 1999, 2004) karena substrat yang dicerna sudah lebih sederhana dibandingkan dengan kayu. Ekspresi endoglukanase pada Macrotermitinae yang bersimbiosis dengan fungi sama dengan ekspresi rayap yang berasosiasi dengan flagellata yaitu pada kelenjar saliva (Tokuda et al. 2004). Hal ini mengindikasikan bahwa Macrotermitinae sebagai nenek moyang dari famili Termitidae merupakan subfamili yang menjadi peralihan antara antara rayap tingkat rendah dan rayap tingkat tinggi. Sedangkan pada rayap tingkat rendah (misal: C. formosanus) yang memakan kayu memerlukan proses degradasi selulosa yang lebih panjang, sehingga endoglukanase disekresikan mulai dari kelenjar saliva, usus depan, dan usus tengah (Lo et al. 2000; Nakashima et al. 2002b).