VERMICOMPOSTING SAMPAH DAUN SONOKELING
(Dalbergia latifolia) MENGGUNAKAN TIGA SPESIES CACING
TANAH (Pheretima sp., Eisenia fetida dan Lumbricus rubellus)
MUHAMMAD ILYAS
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis yang berjudul Vermicomposting Sampah Daun Sonokeling (Dalbergia latifolia) Menggunakan Tiga Spesies Cacing Tanah (Pheretima sp., Eisenia fetida, dan Lumbricus rubellus) adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Agustus 2009
ABSTRACT
MUHAMMAD ILYAS. Vermicomposting of Sonokeling (Dalbergia latifolia) Leaves Litter Using Three Earthworm Species (Pheretima sp., Eisenia fetida, and Lumbricus rubellus. Under direction of RIKA RAFFIUDIN and TRI HERU WIDARTO.
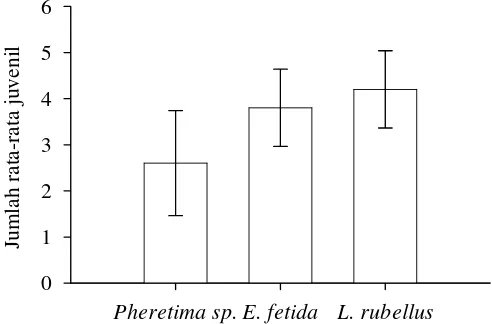
European E. fetida and L. rubellus are the most common earthworm species used in vermicomposting. Pheretima sp. is a native Asian earthworm having capability to decompose organic wastes. Therefore, the aim of the present study was to determine the effectiveness of combination of the three earthworm species in vermicomposting of sonokeling leaves litter by determining (1) their consumption rate, (2) their growth, and (3) their fecundity. Two ratios between earthworms and leaves litter were applied i.e. ratio 1:1 (R1) and ratio 2:1 (R2). The consumption rate was measured by counting the percentage of organic waste consumed by earthworms weekly. Earthworm growth was measured by weighing their biomass weekly. The number of produced cocoons were used to predict their fecundity. The results showed that combination of earthworm species affected the consumption rate significantly in both ratio (P<0.05). In the first week, earthworm biomass increased in R1 ratio, but it tended to decrease in R2 ratio. The highest number of cocoons in R1 ratio was found in L. rubellus, whereas in R2 ratio, it was found in E. fetida in the first week. Cocoon incubation period in Pheretima sp. was faster than the others. The cocoons of L. rubellus showed the highest number of hatchlings (4.2±0.8 individuals) per cocoon. Chemical composition of vermicompost among the three species of earthworms were not different significantly (P>0.05) except in potassium.
RINGKASAN
MUHAMMAD ILYAS. Vermicomposting Sampah Daun Sonokeling (Dalbergia latifolia) Menggunakan Tiga Spesies Cacing Tanah (Pheretima sp., Eisenia fetida, dan Lumbricus rubellus). Dibimbing oleh RIKA RAFFIUDIN dan TRI HERU WIDARTO.
Sampah daun berpotensi sebagai sumber nutrisi yang sangat bermanfaat dalam pertanian. Konsumsi sampah daun dapat menyuburkan tanah dan menyediakan nutrisi bagi tumbuhan. Salah satu cara pengelolaan sampah daun adalah dengan proses vermicomposting. Vermicomposting merupakan proses konsumsi bahan organik yang melibatkan kerjasama antara cacing tanah dan mikroorganisme. Hasil akhir proses ini adalah vermikompos. Pheretima sp., E. fetida, dan L. rubellus merupakan spesies cacing tanah yang berpotensi untuk mengelola sampah organik melalui proses vermicomposting.
Tujuan penelitian ini adalah untuk mengkaji efektivitas spesies cacing tanah Pheretima sp., E. fetida, dan L. rubellus dalam mengelola sampah dedaunan terhadap: (1) laju konsumsi bahan organik, (2) pertumbuhan cacing tanah, dan (3) produktivitas cacing tanah.
Penelitian dilaksanakan dalam dua tahap, yaitu tahap persiapan dan tahap percobaan. Tahap persiapan dilaksanakan dari bulan Maret 2008 sampai bulan Januari 2009. Tahap percobaan dilaksanakan pada bulan Februari-April 2009. Lokasi penelitian terletak di Bagian Fungsi Hayati dan Perilaku Hewan Departemen Biologi, FMIPA, Institut Pertanian Bogor.
Pada tahap persiapan dilakukan penyiapan populasi cacing tanah, media biak, dan wadah percobaan. Cacing tanah yang digunakan pada penelitian ini adalah spesies Pheretima sp., E. fetida dan L. rubellus yang telah berklitelum. Media biak terdiri atas kotoran sapi dan sampah daun sonokeling (Dalbergia latifolia). Kotoran sapi digunakan sebagai starter dan sumber Nitrogen bagi cacing tanah. Sebelum dijadikan sebagai media biak, kotoran sapi dikering-anginkan dan diaduk secara manual setiap hari selama 15 hari untuk menguapkan gas beracun yang ada. Daun sonokeling dihancurkan secara manual hingga mencapai ukuran partikel ± 5 mm. Ke dalam media biak ditambahkan tanah untuk membantu proses pencernaan cacing tanah. Wadah percobaan pada penelitian ini berupa wadah plastik dengan panjang, lebar, dan tinggi 35x31x12.5 cm. Seluruh wadah percobaan dilubangi di bagian bawahnya.
Percobaan ini terdiri atas dua perlakuan, yaitu perlakuan spesies cacing tanah tunggal (P, E, L) dan kombinasi (P+E, P+L, E+L, P+E+L), serta perlakuan rasio antara cacing tanah dan sampah daun 1:1 (R1) dan 2:1 (R2). Suhu, kelembaban, dan pH awal media biak di dalam wadah disesuaikan menjadi 25 oC, 65% dan 6.8 berturut-turut. Setiap minggu dilakukan pengukuran terhadap parameter biologi cacing tanah pada seluruh perlakuan.
Pertumbuhan cacing tanah diukur dengan menimbang cacing tanah di atas timbangan digital (82ADAM, d=0.001 g). Produktivitas cacing tanah diukur dengan menghitung jumlah kokon yang diproduksi di dalam seluruh wadah percobaan setiap minggu. Kokon yang diproduksi oleh masing-masing spesies cacing tanah dipisahkan dari media biak dan ditempatkan ke dalam media lain dengan suhu 25 oC. Perkembangan kokon diamati setiap hari sampai menetas untuk mengetahui masa inkubasi kokon. Selanjutnya, jumlah juvenil yang keluar dari tiap kokon dihitung.
Vermikompos yang diproduksi oleh masing-masing spesies cacing tanah pada perlakuan rasio R2 ditimbang seberat 100 gr untuk dianalisa. Kandungan C organik dan N total dianalisa dengan metode titrasi dan metode Kjeldahl. Kadar P dan K masing-masing dianalisa menggunakan Spektrofotometri dan Flamefotometer. Analisa unsur-unsur kimia vermikompos dilakukan di laboratorium tanah SEAMEO BIOTROP, Bogor.
Berdasarkan pada hasil penelitian, perlakuan spesies tunggal E. fetida dan spesies kombinasi E. fetida + L. rubellus pada rasio R1 dan R2 lebih efektif menkonsumsi sampah dedaunan dibandingkan dengan perlakuan spesies yang lain. Secara ringkas, kecepatan laju konsumsi oleh ketiga spesies cacing tanah pada masing-masing perlakuan adalah: E+L > P+E+L > E > L > P+E > P+L > P. Perbedaan laju konsumsi bahan organik pada cacing tanah disebabkan oleh densitas cacing tanah, kualitas dan kuantitas bahan makanan, serta daya konsumsi spesies spesifik. Semakin tinggi densitas cacing tanah di dalam media biak, maka semakin rendah laju konsumsi. Densitas cacing tanah yang besar menyebabkan peningkatan populasi mikroorganisme di dalam media biak dan saluran pencernaan cacing tanah selama proses vermicomposting. Laju konsumsi yang semakin rendah dikarenakan berkurangnya nutrisi yang berasal dari kotoran sapi sebagai media biak sekaligus sumber Nitrogen dan rendahnya palatabilitas daun sonokeling bagi cacing tanah.
Hasil penelitian menunjukkan bahwa pertumbuhan cacing tanah tertinggi terdapat pada perlakuan spesies tunggal E. fetida dan spesies kombinasi E. fetida + L. rubellus, sedangkan pertumbuhan terendah terdapat pada perlakuan spesies tunggal Pheretima sp dan spesies kombinasi Pheretima sp. + E. fetida pada rasio R1 dan R2. Pertumbuhan cacing tanah bergantung pada ketersediaan dan kualitas bahan makanan, densitas cacing tanah di dalam media biak, serta pola pertumbuhan spesies spesifik. Cacing tanah yang mengkonsumi bahan organik yang lebih banyak dapat tumbuh lebih cepat dibandingkan dengan cacing tanah yang mengkonsumsi sedikit bahan organik. Perbedaan laju pertumbuhan juga dikarenakan adanya interaksi diantara populasi cacing tanah akibat kombinasi spesies tersebut dalam proses vermicomposting. Cacing tanah lebih memilih bahan makanan berupa kotoran hewan daripada bahan organik tumbuhan.
jumlah bahan makanan yang tersedia di dalam media biak, tingkat konsumsi bahan makanan oleh cacing tanah, serta jenis dan kualitas bahan makanan.
Hasil analisa unsur-unsur kimiawi vermikompos Pheretima sp., E. fetida, dan L. rubellus menunjukkan bahwa rasio C:N pada vermikompos lebih rendah daripada substrat awal. Kandungan P dan K meningkat pada vermikompos masing-masing cacing tanah. Vermikompos yang dihasilkan oleh spesies Pheretima sp. mengandung kualitas yang baik. Menurunnya rasio C:N pada vermikompos disebabkan oleh aktivitas cacing tanah dalam proses vermicomposting. Penurunan rasio C:N menunjukkan tingginya humifikasi vermikompos. Peningkatan kandungan mineral P dan K pada vermikompos menunjukkan adanya peningkatan mineralisasi unsur-unsur tersebut yang disebabkan oleh aktivitas enzim dan mikroorganisme di dalam saluran pencernaan cacing tanah. Pelepasan fosfor ke dalam bentuk yang dapat diserap oleh tumbuhan diperantarai oleh fosfat yang dihasilkan di dalam saluran pencernaan cacing tanah, dan selanjutnya pelepasan fosfor dapat dilakukan oleh mikroorganisme di dalam casting setelah dikeluarkan
© Hak Cipta Milik IPB, tahun 2009
Hak Cipta dilindungi Undang-Undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber.
a. Pengutipan untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah.
b. Pengutipan tidak merugikan kepentingan yang wajar IPB.
VERMICOMPOSTING SAMPAH DAUN SONOKELING
(Dalbergia latifolia) MENGGUNAKAN TIGA SPESIES CACING
TANAH (Pheretima sp., Eisenia fetida dan Lumbricus rubellus)
MUHAMMAD ILYAS
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Mayor Biosains Hewan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Vermicomposting Sampah Daun Sonokeling (Dalbergia latifolia) Menggunakan Tiga Spesies Cacing Tanah (Pheretima sp., Eisenia fetida, dan Lumbricus rubellus)
Nama : Muhammad Ilyas NRP : G352070181
Disetujui
Komisi Pembimbing
Dr. Ir. Rika Raffiudin, M.Si Ir. Tri Heru Widarto, M.Sc
Ketua Anggota
Diketahui
Ketua Mayor Dekan Sekolah Pascasarjana
Biosains Hewan
Dr. Bambang Suryobroto Prof. Dr. Ir. Khairil A. Notodiputro, M.S
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya sehingga penulis dapat menyelesaikan tesis yang berjudul “Vermicomposting sampah daun sonokeling (Dalbergia latifolia) Menggunakan Tiga Spesies Cacing Tanah (Pheretima sp., Eisenia fetida dan Lumbricus rubellus)” dengan baik dan tepat waktu. Tesis ini merupakan salah satu syarat untuk memperoleh gelar Magister Sains pada Program Studi Biologi, Mayor Biosains Hewan, Sekolah Pascasarjana, Institut Pertanian Bogor.
Tesis ini tidak akan tersusun tanpa bantuan dari berbagai pihak. Oleh karena itu, penulis ucapkan terima kasih kepada Dr. Ir. Rika Raffiudin, M.Si dan Ir. Tri Heru Widarto, M.Sc, selaku komisi pembimbing yang telah memberikan
arahan dan bimbingan selama penelitian hingga akhir penulisan tesis; Dr. Ir. Muhadiono, M.Sc, selaku penguji luar komisi pembimbing; Bapak Rudi,
selaku peternak cacing tanah yang telah menyediakan cacing tanah sebagai objek penelitian; Hari Nugroho, S.Si, selaku Staf Peneliti di Laboratorium Entomologi, Bidang Zoologi, Puslit Biologi – LIPI yang telah membantu penulis dalam mengidentifikasi cacing tanah; Staf Laboratorium Tanah SEAMEO BIOTROP Bogor atas bantuannya menganalisa kandungan kimiawi vermikompos.
Ucapan terima kasih juga penulis sampaikan kepada Departemen Agama Republik Indonesia yang telah memberikan beasiswa pendidikan Pascasarjana di Institut Pertanian Bogor, Dr. Bambang Suryobroto, Dr. Dedi Duryadi S, DEA, Dr. Achmad Farajallah, M.Si, Dr. RR. Dyah Perwitasari, M.Sc, Dr. Tri Atmowidi, M.Si, Berry Juliandi, S.Si, M.Si, Dra. Taruni Sri Prawasti, dan seluruh staf laboratorium Zoologi yang telah memberikan ilmu dan pengalaman yang sangat bermanfaat. Tidak lupa pula penulis menyampaikan terima kasih kepada seluruh rekan mahasiswa Mayor Biosains Hewan IPB dan keluarga besar Pesantren Ar-Raudhatul Hasanah Medan atas doa, bantuan, dan dukungan yang telah diberikan. Disamping itu, ucapan terima kasih yang sangat dalam penulis sampaikan kepada kedua orang tua tercinta dan seluruh keluarga besar penulis yang telah memberikan doa, bantuan, dan dukungan selama proses pendidikan.
Akhirnya penulis menyadari bahwa tesis ini masih jauh dari kesempurnaan. Kritik dan saran sangat penulis harapkan untuk kesempurnaan tulisan ini. Semoga tesis ini dapat bermanfaat bagi semua pihak.
Bogor, Agustus 2009
RIWAYAT HIDUP
Penulis dilahirkan di Medan pada tanggal 13 Juni 1981 dari Bapak H. M. Tumin S dan Ibu Hj. Marni. Penulis merupakan anak keempat dari empat
bersaudara.
Tahun 1999 penulis lulus dari Madrasah Aliyah Pesantren Ar-Raudhatul Hasanah Medan, dan pada tahun yang sama penulis lulus seleksi masuk Universitas Negeri Medan jurusan Pendidikan Biologi melalui jalur Ujian Masuk Perguruan Tinggi Negeri. Tahun 2007 penulis melanjutkan studi di Sekolah Pascasarjana Institut Pertanian Bogor, Program Studi Biologi, Mayor Biosains Hewan melalui jalur Beasiswa Utusan Daerah yang diselenggarakan oleh Departemen Agama Republik Indonesia.
Selama mengikuti perkuliahan, penulis pernah menjadi asisten praktikum mata kuliah Fungsi Hayati Hewan pada tahun ajaran 2008/2009. Pada tahun 2008 penulis dipilih menjadi peserta terbaik dalam Workshop Pembuatan Media Pendidikan Interaktif (E-book) yang diselenggarakan oleh Himpunan Mahasiswa Ilmu Gizi, Institut Pertanian Bogor.
DAFTAR ISI
Faktor-faktor yang mempengaruhi proses vermicomposting ... 5
Taksonomi Cacing Tanah ... 7
Pengukuran Parameter Biologi Cacing Tanah ... 18
Laju konsumsi bahan organik ... 19
Pertumbuhan cacing tanah ... 19
Produktivitas cacing tanah ... 19
Analisa komposisi kimiawi vermikompos ... 20
Produktivitas Cacing Tanah ... 34
Komposisi Kimiawi Vermikompos ... 37
KESIMPULAN DAN SARAN ... 40
Kesimpulan ... 40
Saran ... 40
DAFTAR PUSTAKA ... 41
DAFTAR TABEL
Halaman
1 Kuantitas media biak dan cacing tanah yang digunakan dalam
percobaan ………..……… 18
2 Perbandingan komposisi kimiawi substrat awal dengan vermikompos yang dihasilkan oleh spesies Pheretima sp., E. fetida, dan
DAFTAR GAMBAR
Halaman
1 Klasifikasi cacing tanah berdasarkan kategori ekologi ... 8
2 Reproduksi cacing tanah (a), pembentukan kokon (b), perkembangan kokon (c), dan penetasan kokon menjadi juvenil (d) ... 11
3 Spesies cacing tanah yang digunakan dalam penelitian: Pheretima sp. (a), E. fetida (b),dan L. rubellus (c) ... 17
4 Budidaya cacing tanah dengan sistem windrow (a)
dan bedding (b) ... 17
5 Wadah percobaan (a), dan penyusunan wadah percobaan
di atas rak (b) ... 17
6 Laju konsumsi pada perlakuan spesies yang berbeda dengan rasio
antara cacing tanah dan sampah dedaunan 1:1 (R1) ... 22
7 Laju konsumsi pada perlakuan spesies yang berbeda dengan rasio
antara cacing tanah dan sampah dedaunan 2:1 (R2) ... 22
8 Pertumbuhan spesies Pheretima sp., E. fetida, dan L. rubellus pada perlakuan yang berbeda dengan rasio antara cacing tanah dan
sampah dedaunan 1:1 (R1) ... 24
9 Pertumbuhan spesies Pheretima sp., E. fetida, dan L. rubellus pada perlakuan yang berbeda dengan rasio antara cacing tanah dan
sampah dedaunan 2:1 (R2) ... 24
10 Jumlah rata-rata kokon yang diproduksi pada perlakuan spesies yang berbeda dengan rasio antara cacing tanah dan sampah dedaunan 1:1 (R1) ... 26
11 Jumlah rata-rata kokon yang diproduksi pada perlakuan spesies yang berbeda dengan rasio antara cacing tanah dan sampah dedaunan 2:1 (R2) ... 26
12 Variasi ukuran dan bentuk kokon cacing tanah: Pheretima sp. (a),
E. fetida (b), dan L. rubellus (c) ... 27
13 Jumlah total kokon yang diproduksi oleh masing-masing spesies cacing tanah dengan rasio antara cacing tanah dan sampah
14 Jumlah total kokon yang diproduksi oleh masing-masing spesies cacing tanah dengan rasio antara cacing tanah dan sampah
dedaunan 2:1 (R2) ... 27
15 Masa inkubasi rata-rata kokon dari tiga spesies cacing tanah
pada suhu 25 oC ... 28
16 Jumlah rata-rata juvenil yang diproduksi oleh masing-masing
DAFTAR LAMPIRAN
Halaman
1 Hasil analisis laju konsumsi pada perlakuan spesies tunggal
dan kombinasi dengan rasio R1 dan R2 setelah 1 minggu ... 47
2 Hasil analisis laju konsumsi pada perlakuan spesies tunggal
dan kombinasi dengan rasio R1 dan R2 setelah 4 minggu ... 51
3 Hasil analisis biomassa cacing tanah pada perlakuan spesies tunggal
dan kombinasi dengan rasio R1 dan R2 setelah 1 minggu ... 55
4 Hasil analisis biomassa cacing tanah pada perlakuan spesies tunggal dan kombinasi dengan rasio R1 dan R2 setelah 4 minggu ... 59
5 Hasil analisis produksi kokon cacing tanah pada perlakuan spesies
tunggal dan kombinasi dengan rasio R1 dan R2 setelah 1 minggu .... 63
6 Hasil analisis produksi kokon cacing tanah pada perlakuan spesies
tunggal dan kombinasi dengan rasio R1 dan R2 setelah 2 minggu .... 67
7 Hasil analisis rasio C:N vermikompos yang diproduksi oleh
cacing tanah Pheretima sp., E. fetida, dan L. rubellus ... 71
8 Hasil analisis kadar P vermikompos yang diproduksi oleh
cacing tanah Pheretima sp., E. fetida, dan L. rubellus ... 72
9 Hasil analisis kadar K vermikompos yang diproduksi oleh
cacing tanah Pheretima sp., E. fetida, dan L. rubellus ... 73
10 Data hasil pengukuran parameter biologi cacing tanah setiap
minggu ... 74
PENDAHULUAN
Latar Belakang
Sampah daun berpotensi sebagai sumber nutrisi yang sangat bermanfaat dalam pertanian. Namun, potensi ini tidak tereksploitasi. Dekomposisi sampah daun dapat menyuburkan tanah dan menyediakan nutrisi bagi tumbuhan. Sampah daun mangga (Mangifera indica) telah didekomposisi menjadi kompos oleh cacing tanah Eudrilus eugeniae (Gajalakshmi et al. 2005). Manimegala et al. (2008) menemukan bahwa sampah daun Leucaena glauca yang dicampur dengan kotoran sapi dapat mendukung pertumbuhan dan produksi kokon pada cacing tanah E. fetida. Sampah daun sonokeling (Dalbergia latifolia) merupakan sampah yang tersebar luas di lingkungan. Namun pengelolaannya menjadi sumber nutrisi yang bermanfaat bagi tanah dan tumbuhan belum dilakukan.
Pengelolaan sampah daun dapat dilakukan dengan cara pengomposan. Pengomposan adalah pembusukan sisa-sisa bahan organik secara aerob dengan cara mendegradasi zat organik menjadi CO2, H2O, NH3, zat-zat anorganik dan bahan organik yang mengandung substansi humus atau kompos (Senesi 1989). Akan tetapi, pengelolaan sampah dengan cara pengomposan tradisional membutuhkan waktu yang cukup lama. Mikroorganisme yang terlibat di dalamnya aktif pada suhu termofilik (45-65 oC) (Dominguez et al. 1997a). Cara alternatif dalam pengelolaan sampah organik adalah dengan vermicomposting.
Vermicomposting berbeda dari pengomposan tradisional dalam beberapa hal. Proses vermicomposting lebih cepat dari pada pengomposan tradisional, karena bahan-bahan organik melewati sistem pencernaan cacing tanah yang mengandung banyak aktivitas mikroorganisme yang membantu proses dekomposisi bahan organik (Dominguez et al. 1997a). Vermicomposting mengubah sampah organik menjadi kompos dalam 30 hari, menurunkan rasio C:N dan meningkatkan kandungan N total lebih tingi dari pada pengomposan tradisional (Gandhi et al. 1997; Lazcano et al. 2008).
Vermikompos yang dihasilkan dari proses vermicomposting memiliki struktur, porositas, aerasi, drainase dan kapasitas menahan kelembaban yang sangat baik (Dominguez et al. 1997a). Vermikompos mengandung banyak aktivitas, populasi dan keanekaragaman mikroorganisme. Vermikompos juga mengandung beberapa enzim seperti protease, amilase, lipase, selulase, dan kitinase (Subler et al. 1998), serta zat pengatur tumbuh seperti giberelin, sitokinin dan auksin (Tomatti et al. 1988). Subler et al. (1998) menemukan bahwa vermikompos cenderung memiliki nilai pH lebih rendah, konsentrasi nutrisi lebih tinggi, terutama nitrogen dari pada kompos alami.
Cacing tanah memiliki peranan yang penting dalam menghancurkan bahan organik sehingga dapat memperbaiki aerasi dan struktur tanah. Akibatnya lahan menjadi subur dan penyerapan nutrisi oleh tanaman menjadi baik. Keberadaan cacing tanah dapat meningkatkan populasi mikroorganisme yang bermanfaat bagi tanaman. Cacing tanah juga dapat mendekomposisi sampah organik menjadi humus (Sharma et al. 2005)
E. fetida dan L. rubellus merupakan spesies cacing tanah epigeic yang sangat toleran terhadap suhu lingkungan (Reinecke et al. 1992). Potensi E. fetida dan L. rubellus dalam mendekomposisi sampah organik telah dipelajari oleh beberapa peneliti (Albanell 1988; Reinecke et al. 1992; Delgado et al. 1995; Gunadi et al. 2003; Garg et al. 2005; Aira et al. 2006a). Spesies cacing tanah dari genus Pheretima yang mendominasi wilayah Indonesia belum diketahui potensinya dalam mengelola sampah organik.
Di alam, beberapa spesies cacing tanah yang berbeda dapat hidup pada habitat yang sama, masing-masing cacing tanah menempati relung yang berbeda dan menggunakan substrat yang berbeda sebagai bahan makanan. Oleh karena itu, pemanfaatan campuran beberapa spesies cacing tanah (spesies kombinasi) pada proses vermicomposting kemungkinan dapat mencapai stabilisasi bahan organik yang lebih tinggi daripada spesies tunggal.
Kombinasi beberapa spesies cacing tanah dapat mendekomposisi bahan organik lebih efisien (Sinha et al. 2002; Khwairakpam & Bhargava 2009). Akan tetapi, Loehr et al. (1985) menemukan bahwa kombinasi beberapa spesies tidak menunjukkan keunggulan yang signifikan dibandingkan dengan biakan spesies tunggal dalam proses vermicomposting.
Tujuan Penelitian
Berdasarkan pada latar belakang tersebut, maka tujuan penelitian ini bertujuan mengkaji efektivitas penggunaan spesies cacing tanah Pheretima sp., E. fetida, dan L. rubellus dalam mengelola sampah daun baik secara tunggal maupun kombinasi dengan rasio antara cacing tanah dan sampah daun yang berbeda terhadap: (1) laju konsumsi bahan organik, (2) pertumbuhan cacing tanah, dan (3) produktivitas cacing tanah.
Manfaat Penelitian
TINJAUAN PUSTAKA
Vermicomposting
Dominguez et al. (1997a) mendefinisikan vermicomposting sebagai proses dekomposisi bahan organik yang melibatkan kerjasama antara cacing tanah dan mikroorganisme. Mikroorganisme yang berperan dalam proses vermicomposting terutama bakteri, fungi, dan actinomycetes.
Selama proses vermicomposting, nutrisi pada tumbuhan yang penting seperti nitrogen, kalium, dan fosfor yang terdapat di dalam bahan makanan diubah melalui aktivitas mikroorganisme menjadi bentuk yang lebih mudah diserap oleh tumbuhan (Ndegwa & Thompson. 2001). Pada proses ini cacing tanah mengubah aktivitas mikroorganisme (Aira et al. 2002), sehingga laju mineralisasi bahan-bahan organik bertambah cepat (Albanell et al. 1988). Beberapa enzim yang terlibat di dalam dekomposisi bahan organik adalah dehidrogenase, protease, glukosidase, dan fosfatase (Lazcano et al. 2008). Vermicomposting menghasilkan dua manfaat utama, yaitu biomassa cacing tanah dan vermikompos (Sharma et al. 2005).
Vermikompos merupakan bahan organik seperti tanah yang memiliki struktur, porositas, aerasi, drainase, dan kapasitas menahan kelembaban yang sangat baik (Dominguez et al. 1997a). Vermikompos mengandung banyak aktivitas, populasi, dan keanekaragaman mikroorganisme. Vermikompos juga mengandung beberapa enzim seperti protease, amilase, lipase, selulase, dan kitinase (Subler et al. 1998), serta zat pengatur tumbuh seperti giberelin, sitokinin, dan auksin (Tomatti et al. 1988).
Beberapa spesies cacing tanah yang memenuhi syarat biologi dan digunakan dalam proses vermicomposting adalah: E. fetida (Albanell 1988), (Reinecke et al. 1992), (Gunadi et al. 2003), (Garg et al. 2005), (Aira et al. 2006a); E. andrei (Dominguez et al. 2000); L. rubellus (Delgado et al. 1995); L. terrestris, Eudrilus eugeniae (Banu et al. 2008); Perionyx excavatus, dan P. sansibaricus (Suthar 2007a; Suthar & Singh 2008).
Faktor-faktor yang mempengaruhi proses vermicomposting
Proses vermicomposting dalam kaitannya dengan pertumbuhan dan kelangsungan hidup cacing tanah dipengaruhi oleh beberapa faktor, misalnya: suhu, kelembaban, rasio C:N, pH, aerasi, dan makanan. Pengaruh faktor-faktor tersebut bervariasi pada setiap spesies cacing tanah.
Suhu pada substrat mempengaruhi pertumbuhan cacing tanah. Pada kisaran suhu 20-29 oC cacing tanah tumbuh dan berkembang dengan maksimal (Kaplan et al. 1980). Hou et al. (2005) menemukan bahwa laju pertumbuhan cacing tanah tercepat pada suhu 20 oC. Akan tetapi kebutuhan suhu pada masing-masing spesies cacing tanah berbeda-beda. Di awal proses vermicomposting terjadi peningkatan suhu, dan di akhir periode suhu menurun.
Rasio C:N pada substrat (media biak yang terdiri atas kotoran ternak dan sampah organik) mencerminkan mineralisasi dan stabilisasi sampah organik selama proses vermicomposting (Suthar & Singh 2008). Laju dekomposisi tertinggi terdapat pada substrat dengan rasio C:N sebesar 20 (Hou et al. 2005). Ndegwa dan Thompson (2000) menemukan bahwa cacing tanah dapat tumbuh lebih baik ketika rasio C:N substrat 25. Cacing tanah lebih memilih sampah organik yang memiliki rasio C:N lebih rendah sebagai makanannya (Moody et al. 1995). Semakin rendah rasio C:N, maka semakin tinggi laju dekomposisi sampah organik (Pramanik et al. 2007).
Substrat yang dikonsumsi cacing tanah memliki kisaran nilai pH tertentu. Substrat yang terlalu asam atau basa tidak cocok untuk kelangsungan hidup cacing tanah. Kisaran nilai pH optimum bagi cacing tanah antara 6.5 dan 8.5 (Hou et al. 2005). Pada umumnya spesies cacing tanah lebih memilih substrat yang memiliki pH sekitar 7.0 (Arrhenius 1921). Singh et al. (2005) menemukan bahwa pH substrat awal yang mendekati netral mengoptimalkan stabilisasi bahan organik dalam waktu yang singkat. Substrat awal yang memiliki pH sangat asam tidak sesuai digunakan dalam proses vermicomposting.
Proses vermicomposting dapat berlangsung dengan baik dalam kondisi aerob. Cacing tanah memerlukan oksigen untuk bernafas dan sangat sensitif terhadap kondisi anaerob. Laju respirasinya melemah jika konsentrasi oksigen di dalam substrat rendah (Edwards & Bohlen 1996). Pergerakan cacing tanah dapat menciptakan aerasi pada medianya. Untuk meningkatkan aerasi, perlu dilakukan pembalikan substrat.
Taksonomi Cacing Tanah
Cacing tanah adalah invertebrata darat yang termasuk ke dalam Filum Annelida, Kelas Clitellata, Sub Kelas Oligochaeta (Brusca & Brusca 2003). Cacing tanah terbagi ke dalam 5 famili, yaitu Moniligastridae, Megascolecidae, Eudrilidae, Glossoscolecidae, dan Lumbricidae.
Klasifikasi Cacing Tanah
Spesies cacing tanah yang berbeda memiliki sejarah hidup yang berbeda dan menempati relung ekologi yang berbeda. Lee (1985) mengelompokkan spesies cacing tanah ke dalam tiga kategori ekologi berdasarkan strategi mencari makanan dan membuat liang, yaitu spesies epigeic, endogeic, dan anecic (Gambar 1).
Cacing tanah epigeic pada dasarnya merupakan cacing tanah penghuni sampah. Cacing tanah ini hidup di dalam atau dekat permukaan sampah dan memakan sampah organik yang kasar, serta sejumlah sampah yang belum terurai. Cacing tanah epigeic membuat liang ephemeral ke dalam tanah mineral selama periode diapause. Tubuhnya berukuran kecil dan berpigmen. Laju metabolisme dan reproduksinya tinggi. Hal itu menggambarkan daya adaptasinya tinggi terhadap perubahan kondisi lingkungan pada permukaan tanah. Beberapa spesies cacing tanah yang termasuk ke dalam kategori ini adalah L. rubellus, E. fetida, E. andrei, Dendrobaena rubida, Eudrilus eugeniae, Perionyx excavatus, dan Eiseniella tetraedra (Bouche 1977; Lee 1985).
Cacing tanah jenis anecic hidup di dalam sistem liang vertikal yang lebih permanen, yang dapat meluas beberapa meter ke dalam tanah (Gambar 1). Cacing tanah jenis ini dapat di temukan di dalam liang yang dangkal atau dalam tergantung pada kondisi tanah yang baik sebagai habitatnya. Cacing tanah anecic mengeluarkan sisa pencernaannya (casting) pada permukaan tanah dan muncul di malam hari untuk memakan sampah pada permukaan tanah, kotoran, dan bahan organik lain yang diturunkan ke dalam liangnya. Laju reproduksinya relatif lambat, terbukti dari produksi kokonnya. Cacing tanah anecic memiliki peran yang sangat besar dalam dekomposisi bahan organik, siklus makanan, dan pembentukan tanah (Lavelle 1988). Lumbricus terrestris, Aporrectodea trapezoids,dan Allolobophora longa termasuk dalam kelompok kategori ini.
Biologi Cacing Tanah
Karakteristik cacing tanah adalah tubuhnya bersegmen, dan memiliki sedikit seta pada seluruh segmen tubuh. Cacing tanah dewasa memiliki klitelum yang terletak di bagian anterior tubuh. Klitelum merupakan bagian kelenjar epidermis segmen tubuh yang mengalami perkembangan, terdiri atas kelenjar epidermis yang menebal, terutama di bagian dorsal dan lateral tubuh. (Edwards & Lofty 1972). Pada umumnya klitelum berwarna lebih cerah daripada segmen yang lain. Gambar 1 Klasifikasi cacing tanah berdasarkan kategori ekologi (Lee 1985)
Epigeic
Endogeic
Anecic
Sistem reproduksi
Cacing tanah bersifat hermafrodit, tetapi fertilisasi tidak dapat terjadi pada diri sendiri. Pada umumnya individu cacing tanah dewasa melakukan reproduksi silang sebelum menghasilkan kokon (Gambar 2a). Beberapa spesies cacing tanah melakukan reproduksi pada permukaan tanah, dan beberapa spesies melakukannya di bawah tanah (Edwards & Lofty 1972).
Metode kopulasi untuk seluruh spesies cacing tanah tidak sama. Pada cacing tanah yang tergolong famili Lumbricidae, ketika akan melakukan perkawinan dua spesies cacing tanah saling berdekatan dengan mendeteksi mukus yang dikeluarkan oleh bagian ventral tubuhnya bersama-sama. Ujung kepala cacing tanah terletak pada arah yang berlawanan. Keduanya saling mendekatkan diri pada daerah pembukaan spermateka dimana daerah klitelum salah satu cacing tanah menyentuh permukaan pembukaan spermateka yang lainnya. Pada saat kopulasi, kedua cacing tanah tidak sensitif dalam merespon rangsangan luar seperti sentuhan dan cahaya. Banyak mukus yang disekresikan sehingga masing-masing cacing tanah diselubungi oleh mukus antara segmen sembilan dan sisi posterior klitelum, mukus-mukus tersebut saling melekat (Edwards & Lofty 1972).
Sebuah celah semen terbentang dari gonofor jantan sampai klitelum dan nampak seperti benang. Tiap-tiap celah semen merupakan bagian dari dinding luar tubuh yang melekuk ke dalam akibat dari terbentuknya rangkaian pori-pori oleh kontraksi otot yang terbentang pada lapisan otot longitudinal. Kontraksi otot membawa cairan sperma dari gonofor jantan menuju daerah klitelum. Cairan sperma berkumpul di daerah klitelum, dan akhirnya memasuki spermateka cacing tanah lawannya (Edwards & Lofty 1972).
cairan sperma disalurkan ke dalam sepasang spermateka berikutnya sampai seluruhnya terisi.
Setelah kopulasi berlangsung selama satu jam, cacing tanah terpisah, dan masing-masing klitelum mengeluarkan getah mukus yang akhirnya mengeras di sekeliling permukaan luarnya. Ketika getah mukus mengeras, cacing tanah bergerak ke arah belakang, kemudian membuat selubung di sekeliling kepalanya, dan ketika cacing tanah terpisah sempurna, ujung selubung menutup untuk membentuk kokon (Gambar 2b). Kokon mengandung cairan albumin yang diproduksi oleh kelenjar klitelum, ovum, dan spermatozoa yang disalurkan ke dalamnya ketika melewati pembukaan spermateka. Kokon terus dibentuk sampai cairan sperma yang tersedia habis. Fertilisasi terjadi secara eksternal tubuh cacing tanah, di dalam kokon (Edwards & Lofty 1972).
Produksi kokon dapat dipengaruhi oleh beberapa faktor. Keadaan tanah dan jenis cacing tanah yang berbeda menyebabkan perbedaan jumlah kokon yang diproduksi. Cacing tanah memproduksi lebih sedikit kokon pada kondisi tanah yang terlalu kering dan terlalu basah (Gerard 1967). Jenis makanan yang dikonsumsi cacing tanah dewasa juga dapat mempengaruhi produksi kokon. Produksi kokon bergantung pada spesies cacing tanah dan kondisi lingkungan (Evans & Guild 1948). Jumlah kokon yang diproduksi cacing tanah epigeic lebih banyak daripada kokon cacing tanah anecic (Bhattacharjee & Chaudhuri 2002).
Sistem pencernaan
Sistem pencernaan cacing tanah terdiri atas rongga mulut, faring, esofagus, tembolok, lambung, dan usus. Cacing tanah memperoleh makanan dari bahan organik berupa organ tumbuhan, protozoa, rotifera, nematoda, bakteri, fungi, dan sisa-sisa pembusukan hewan (Edwards & Lofty 1972).
Bagian tubuh cacing tanah dikelompokkan ke dalam tiga daerah berdasarkan pada segmen tubuhnya (Gansen 1962). Daerah yang pertama adalah daerah penerimaan, yang terdapat pada segmen 1-14. Daerah ini terdiri atas mulut yang peka, esofagus, dan kelenjar faring tak berpembuluh yang mensekresi getah asam yang mengandung enzim amilase.
Daerah yang kedua adalah daerah sekresi, yang terdapat pada segmen 15-44. Daerah ini terdiri atas tembolok, yang menuju ke lambung dan usus. Cacing tanah mensekresi dua enzim protease dan satu enzim amilase terutama dari “sel piala”
b
c
Juvenil
d a
Kepala Klitelum
Kepala Klitelum
Liang jantan Reproduksi
cacing tanah
Perpindahan sperma
Klitelum
dinding usus yang mensekresi banyak getah. Beberapa enzim yang berbeda berasal dari daerah ini untuk spesies lain, seperti lipase dan protease (Arthur 1965) serta selulase dan kitinase (Tracey 1951). Makanan yang telah dicerna melintasi aliran darah di sepanjang epithelium usus dan dialirkan ke berbagai bagian tubuh dan jaringan untuk digunakan dalam proses metabolisme dan sebagai cadangan makanan.
Daerah yang ketiga terdapat pada segmen 44 sampai ke anus, disebut daerah absorpsi. Pada daerah ini, bahan-bahan makanan yang tidak tercerna di dalam usus diselubungi oleh membran peritrofik yang melapisi usus. Ketika bahan-bahan makanan diekskresi, membran tersebut akan membungkus casting (sisa pencernaan/kotoran cacing) (Gansen 1962).
Sistem ekskresi
Organ ekskresi yang terpenting pada cacing tanah adalah nefridia. Organ tersebut mengekstraksi bahan-bahan limbah dari cairan selom dan mengeluarkannya ke luar tubuh melalui nefridiofor sebagai urin yang mengandung amonia dan urea. Cairan selom yang mengandung material eksresi melintas melalui nefrostom dan dilanjutkan ke tabung nefridia oleh gerakan silia. (Edwards & Lofty 1972).
Nefridia berperan sebagai penyaring diferensial. Disebut demikian karena terdapat lebih banyak urea dan amonia, tetapi lebih sedikit kreatinin dan protein di dalam urin yang dihasilkan daripada di dalam cairan selom (Ramsay 1949). Nefridia memiliki tiga fungsi pada proses ekskresi, yaitu filtrasi, reabsorbsi, dan transformasi kimiawi (Bahl 1947).
dinding usus, dan selanjutnya masuk ke dalam usus untuk diekskresikan bersama feses (Edwards & Lofty 1972).
Sistem saraf
Sistem saraf cacing tanah terdiri atas sebuah ganglion serebral dorsal, sepasang konektif sirkumentrik dan satu atau lebih tali saraf longitudinal. Ganglion serebral dorsal menyuplai saraf bagian anterior tubuh dan saraf prostomial. Pengontrolan pergerakan tubuh cacing tanah diatur oleh ganglion subenterik. Sedangkan konektif sirkumenterik dan tali saraf longitudinal mengontrol saraf sensoris dan motoris keseluruhan dinding tubuh serta organ di setiap tubuh (Edwards & Lofty 1972).
Organ sensoris cacing tanah terdiri atas dua macam, yaitu organ fotoreseptor dan organ sensoris epitel. Di dalam setiap sel organ fotoreseptor terdapat organel optik yang berbentuk lonjong atau memanjang. Permukaan luar organel ini terdiri atas retina, dan permukaan dalam terdiri atas substansi hialin yang transparan. Organ sensoris epitel merupakan kumpulan dari 35-45 sel-sel yang memanjang dan besar di bagian dasarnya. Ujung distalnya berakhir pada penonjolan rambut-rambut sensori sepanjang daerah kutikula yang tipis. Cacing tanah tidak memiliki mata, tetapi spesies ini memiliki sel-sel sensori yang strukturnya seperti lensa di daerah epidermis dan dermis, terutama pada prostomium. Bagian tengah dan prosterior tubuhnya kurang sensitif terhadap cahaya (Laverack 1963).
Cacing tanah sangat respon terhadap rangsangan cahaya, terutama jika tiba-tiba terpapar cahaya setelah berdiam lama dalam kondisi gelap (Laverack 1963). Cacing tanah Lumbricus bersifat fotopositif terhadap sumber cahaya yang sangat lemah, dan fotonegatif terhadap sumber cahaya yang kuat. Akan tetapi cacing tanah tidak terlalu bereaksi terhadap peningkatan intensitas cahaya yang tiba-tiba jika telah teradaptasi dalam waktu yang lama dalam kondisi cahaya yang kuat. Hal itu disebabkan oleh saturasi reseptor cahaya (Hess 1924). Anggota spesies dari genus Pheretima seluruhnya bersifat fotonegatif terhadap intensitas cahaya (Howell 1939).
dalam kehidupan cacing tanah. Kemoreseptor dapat mendeteksi dan mengumpulkan bahan makanan. Kemoreseptor juga dapat digunakan untuk memberi informasi tentang kondisi lingkungan. Disamping itu, kemoreseptor berperan dalam proses perkawinan dengan mendeteksi getah mukus yang dihasilkan oleh cacing tanah yang lain (Smith 1902).
Distribusi Geografi Cacing Tanah
Distribusi cacing tanah sangat luas di seluruh dunia. Akan tetapi pada daerah gurun, kutub, pegunungan, dan daerah dengan sedikit tanah dan vegetasi cacing tanah jarang ditemukan. Beberapa spesies cacing tanah yang terdistribusi secara luas dikenal dengan istilah perigrin (kosmopolitan), sedangkan spesies yang hanya terdapat pada satu daerah tertentu dikenal dengan istilah endemik (Edwards & Lofty 1972).
Spesies perigrin dari famili Lumbricidae menyebar ke beberapa wilayah, terutama di Eropa. Wilayah-wilayah lain yang merupakan daerah penyebaran spesies cacing tanah dari famili ini adalah Chili, Selandia Baru, beberapa daerah di Amerika Serikat, Afrika Barat, India bagian barat laut, dan Australia. Penyebaran Eisenia terdapat di wilayah Eropa, Siberia, Rusia Selatan, Israel, dan Amerika Utara. Sedangkan Lumbricus tersebar ke wilayah Eropa, Siberia, Islandia, dan Amerika Utara. Beberapa spesies perigrin dari genus-genus tersebut terdistribusi di seluruh dunia (Edwards & Lofty 1972; Reynolds 1994).
METODE
Waktu dan Lokasi
Penelitian ini dilakukan dalam dua tahap, yaitu tahap persiapan dan tahap percobaan. Tahap persiapan dilaksanakan dari bulan Maret 2008 sampai bulan Januari 2009. Tahap percobaan dilaksanakan pada bulan Februari-April 2009. Lokasi penelitian terletak di Bagian Fungsi Hayati dan Perilaku Hewan Departemen Biologi, FMIPA, Institut Pertanian Bogor.
Tahap Persiapan
Pada tahap ini dilakukan penyiapan populasi cacing tanah, media biak, dan wadah percobaan. Cacing tanah yang digunakan pada penelitian adalah spesies Pheretima sp., E. fetida dan L. rubellus yang telah berklitelum (Gambar 3a-c). Identifikasi spesies cacing tanah dilakukan di Laboratorium Entomologi, Bidang Zoologi Puslit Biologi – LIPI, Cibinong. Spesies Pheretima sp. diperoleh dari dalam kotoran sapi yang berasal dari kandang Fakultas Peternakan IPB, sedangkan E. fetida dan L. rubellus diperoleh dari Bapak Rudi Rochmat pada peternakan “Kelompok Cacing Tanah Mandiri” di Desa Warna Sari, Kecamatan Pengalengan, Kabupaten Bandung. Ketiga populasi cacing tanah dibudidaya di Bagian Fungsi Hayati dan Perilaku Hewan Departemen Biologi IPB selama 10 bulan. Budidaya bertujuan memperbanyak populasi cacing tanah sehingga dapat dijadikan sebagai stok pada tahap percobaan. Cacing tanah yang dibudidaya pada tahap persiapan sebanyak ± 1 kg untuk spesies E. fetida dan L. rubellus, serta ± 1.5 kg untuk spesies Pheretima sp. Budidaya cacing tanah dilakukan dengan sistem windrow, yaitu sistem budidaya cacing tanah dalam barisan yang disusun sejajar (Gambar 4a), dan bin, yaitu sistem budidaya cacing tanah yang menggunakan wadah (Gambar 4b)
media biak, kotoran sapi dikering-anginkan dan diaduk secara manual setiap hari selama 15 hari untuk menguapkan gas beracun (Garg et al. 2005).
Daun pohon sonokeling (D. latifolia) diperoleh dari sekitar Kampus IPB. Daun dihancurkan secara manual hingga mencapai ukuran partikel ± 5 mm. Ke dalam media biak ditambahkan tanah untuk membantu proses pencernaan cacing tanah. Tanah diperoleh dari kebun yang terletak di sekitar Bagian Fungsi Hayati dan Perilaku Hewan Departemen Biologi IPB. Selama masa budidaya, media biak diaduk dan disiram dengan air secukupnya untuk mempertahankan suhu (25 oC) dan kelembaban media (65%). Pengadukan dan penyiraman media biak dilakukan dengan selang waktu tiga hari.
Wadah percobaan pada penelitian ini berupa wadah plastik dengan panjang, lebar, dan tinggi 35x31x12.5 cm (Gambar 5a). Seluruh wadah percobaan dilubangi di bagian bawahnya untuk menciptakan aerasi di dalam wadah.
Tahap Percobaan
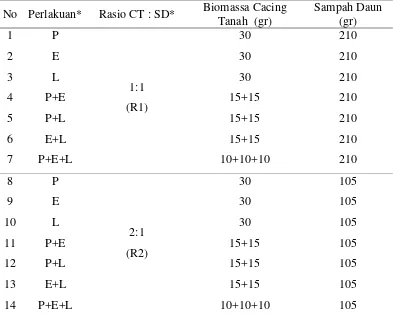
Seluruh bahan pada media biak dimasukkan ke dalam masing-masing wadah percobaan. Selanjutnya populasi cacing tanah dimasukkan ke dalam wadah yang telah berisi media biak. Kuantitas media biak dan cacing tanah yang digunakan dalam percobaan ini tertera pada Tabel 1. Seluruh wadah percobaan disusun bertingkat di atas rak (Gambar 5b).
Percobaan ini terdiri atas dua perlakuan, yaitu: 1) perlakuan spesies cacing tanah tunggal dan kombinasi, dan 2) perlakuan rasio antara cacing tanah dan sampah daun. Perlakuan spesies cacing tanah tunggal meliputi P, E, dan L, sedangkan spesies cacing tanah kombinasi meliputi P+E, P+L, E+L, dan P+E+L. Perlakuan rasio cacing tanah dan sampah daun terdiri atas 1:1 (selanjutnya disebut rasio R1), dan 2:1 (selanjutnya disebut rasio R2) (Tabel 1).
Gambar 5 Wadah percobaan (a), dan penyusunan wadah percobaan di atas rak (b).
a b
Gambar 4 Budidaya cacing tanah dengan sistem windrow (a) dan bin (b).
Bin
b a
Gambar 3 Spesies cacing tanah yang digunakan dalam penelitian: Pheretima sp. (a), E. fetida (b),dan L. rubellus (c).
Tabel 1 Kuantitas media biak dan cacing tanah yang digunakan dalam percobaan *Perlakuan; P: Pheretima sp., E: E. fetida, L: L. rubellus. CT : SD: Cacing Tanah:Sampah Daun.
Ke dalam wadah percobaan diletakkan bedding berupa daun pisang untuk mempertahankan kelembaban media. Bedding diletakkan di bagian atas bahan-bahan media biak secara terpisah dengan menggunakan kain jala.
Suhu, kelembaban, dan pH awal media biak disesuaikan menjadi 25 oC, 65% dan 6.8 berturut-turut. Suhu diukur menggunakan thermometer, sedangkan kelembaban dan pH diukur dengan soil tester. Setiap minggu dilakukan pengukuran terhadap parameter biologi penelitian pada seluruh perlakuan.
Pengukuran Parameter Biologi Cacing Tanah
0 t
Laju konsumsi bahan organik diukur dengan menimbang seluruh bahan makanan yang terdapat pada media biak menggunakan timbangan digital (82ADAM, d=0.001 g). Persentase laju konsumsi dihitung dengan persamaan:
Keterangan: B0 : berat awal bahan makanan
Bt : berat bahan makanan pada waktu pengamatan ke-t
Bahan pada media yang telah ditimbang dimasukkan kembali ke dalam wadah percobaan. Kemudian dilakukan penambahan sampah daun sonokeling setiap minggu ke dalam media biak sebanyak 210 gr untuk rasio R1 dan 105 gr untuk rasio R2. Kuantitas sampah daun yang ditambahkan pada rasio R1 dan R2 diperoleh berdasarkan pengetahuan umum yang menyatakan bahwa cacing tanah dapat mengkonsumsi bahan makanan sebesar bobot tubuhnya dalam waktu 24 jam. Di lain pihak, Haimi dan Huhta (1986) menemukan bahwa cacing tanah dapat mengkonsumsi bahan makanan setengah dari bobot tubuhnya per hari pada kondisi lingkungan yang sesuai. Sampah daun diletakkan secara homogen ke dalam media biak.
Pertumbuhan cacing tanah
Populasi cacing tanah yang telah dipisahkan secara hand sorting ditimbang sebagai biomassa basah untuk mengetahui pertumbuhannya (Suthar & Singh 2008). Pengukuran biomassa cacing tanah dilakukan mengggunakan timbangan digital (82ADAM, d=0.001 g). Kemudian cacing tanah dimasukkan kembali ke dalam wadah percobaan.
Produktivitas cacing tanah
Perkembangan kokon diamati setiap hari sampai menetas untuk mengetahui masa inkubasi kokon. Selanjutnya, jumlah juvenil yang keluar dari tiap kokon dihitung (Bhattacharjee & Chaudhuri 2002).
Analisa komposisi kimiawi vermikompos
Pada minggu keempat, bahan-bahan organik yang telah terdekomposisi selama masa percobaan disaring untuk memperoleh hasil akhir vermicomposting berupa vermikompos. Vermikompos yang diproduksi oleh masing-masing spesies cacing tanah pada perlakuan rasio R2 ditimbang seberat 100 gr untuk dianalisa.
Komposisi kimiawi vermikompos yang dianalisa terdiri atas karbon organik, nitrogen total, fosfor dan kalium. Kandungan C organik dan N total dianalisa dengan metode titrasi dan metode Kjeldahl (Walkey & Black 1934). Nilai rasio C:N diperoleh dengan membagi nilai kandungan C organik dengan N total. Kadar P dan K masing-masing dianalisa menggunakan Spektrofotometri dan Flamefotometer (Jackson 1973). Analisa unsur-unsur kimia vermikompos dilakukan di laboratorium tanah SEAMEO BIOTROP, Bogor.
Analisa Data
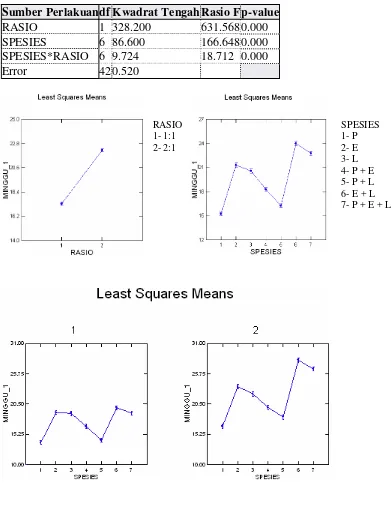
Rancangan percobaan yang digunakan pada penelitian ini adalah Rancangan Acak Lengkap (RAL) pola faktorial (7 x 2) dengan 4 ulangan. Sebagai faktor pertama adalah spesies cacing tanah tunggal dan kombinasi: P, E, L, P+E, P+L, E+L dan P+E+L. Faktor kedua adalah rasio cacing tanah:sampah: 1:1 dan 2:1, sehingga total percobaan adalah 56 unit percobaan.
HASIL
Laju konsumsi bahan organik
Hasil penelitian menunjukkan bahwa setelah 1 minggu laju konsumsi tertinggi pada perlakuan spesies tunggal dengan rasio R1 terdapat pada perlakuan E (19.05%), sedangkan laju konsumsi terendah terdapat pada perlakuan P (13.82%). Pada perlakuan spesies kombinasi, laju konsumsi tertinggi terdapat pada perlakuan E+L (19.84%), sedangkan laju konsumsi terendah terdapat pada perlakuan P+L (14.24%) (Gambar 6). Namun, setelah 4 minggu laju konsumsi tertinggi terdapat pada perlakuan spesies kombinasi P+E+L (36.32%), sedangkan laju konsumsi terendah terdapat pada perlakuan spesies tunggal P (31.56%) (Lampiran 1 & 2).
Laju konsumsi dengan rasio R2 menunjukkan pola grafik yang hampir sama dengan rasio R1, akan tetapi persentasenya lebih tinggi. Setelah 1 minggu laju konsumsi tertinggi pada perlakuan spesies tunggal terdapat pada perlakuan E (23.54%), sedangkan laju konsumsi terendah terdapat pada perlakuan P (16.64%). Pada perlakuan spesies kombinasi, laju konsumsi tertinggi terdapat pada perlakuan E+L (28.13%), sedangkan laju konsumsi terendah terdapat pada perlakuan P+L (18.24%) (Gambar 7). Setelah 4 minggu, laju konsumsi tertinggi terdapat pada perlakuan spesies kombinasi E+L (41.08%), sedangkan laju konsumsi terendah terdapat pada perlakuan spesies tunggal P (33.57) (Lampiran 1 & 2).
Waktu (minggu)
Gambar 6 Laju konsumsi pada perlakuan spesies yang berbeda dengan rasio antara cacing tanah dan sampah dedaunan 1:1 (R1).
Pertumbuhan Cacing Tanah
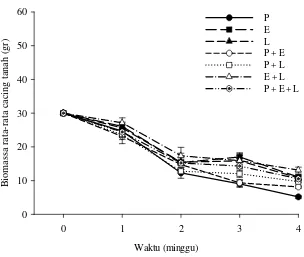
Secara umum pertumbuhan cacing tanah pada rasio R1 cenderung menurun (Gambar 8). Setelah 1 minggu pertumbuhan cacing tanah tertinggi pada perlakuan spesies tunggal terdapat pada perlakuan E dengan biomassa rata-rata maksimum 46.8±1.2 gr. Pertumbuhan terendah ditunjukkan pada perlakuan P dengan biomassa rata-rata 29.0±1.1 gr. Pada perlakuan spesies kombinasi, biomassa tertinggi terdapat pada perlakuan E+L (39.7±3.8 gr), sedangkan biomassa terendah terdapat pada perlakuan P+E (33.7±4.8 gr). Gambar 8 menunjukkan biomassa cacing tanah pada seluruh perlakuan bertambah setelah satu minggu, kecuali pada perlakuan spesies tunggal P yang menunjukkan penurunan biomassa (29.0±1.1 gr). Setelah 4 minggu, biomassa tertinggi terdapat pada perlakuan spesies kombinasi E+L (19.6±1.4 gr), sedangkan biomassa terendah terdapat pada perlakuan spesies tunggal P (9.9±0.3 gr) (Lampiran 3 & 4).
Seluruh cacing tanah pada rasio R2 mengalami penurunan biomassa sejak minggu pertama (Gambar 9). Pada perlakuan spesies tunggal, biomassa tertinggi setelah 1 minggu terdapat pada perlakuan E (26.1±1.0 gr), sedangkan biomassa terendah terdapat pada perlakuan P (24.7±1.8 gr). Pada perlakuan spesies kombinasi, biomassa tertinggi terdapat pada perlakuan E+L (27.1±1.5 gr), sedangkan biomassa terendah terdapat pada perlakuan P+E (23.1±2.1 gr). Setelah 4 minggu, biomassa tertinggi terdapat pada perlakuan spesies kombinasi E+L (13.1±0.9 gr), sedangkan biomassa terendah terdapat pada perlakuan spesies tunggal P (5.1±0.5 gr) (Lampiran 3 & 4).
Waktu (minggu)
Gambar 8 Pertumbuhan spesies Pheretima sp., E. fetida, dan L. rubellus pada perlakuan yang berbeda dengan rasio antara cacing tanah dan sampah dedaunan 1:1 (R1).
Produktivitas Cacing Tanah
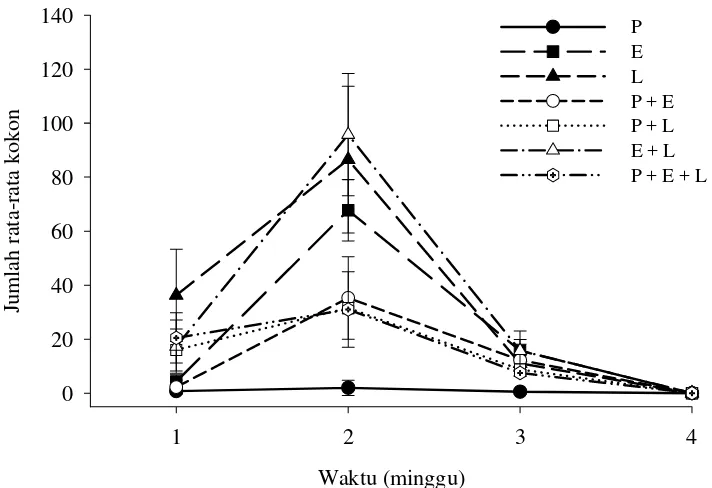
Hasil penelitian ini menunjukkan bahwa pada perlakuan spesies tunggal dengan rasio R1 jumlah rata-rata produksi kokon tertinggi terdapat pada perlakuan L (36.25±17.1 kokon), sedangkan produksi kokon terendah terdapat pada perlakuan P (0.75±0.1 kokon) setelah 1 minggu (Gambar 10). Pada perlakuan spesies kombinasi, jumlah rata-rata produksi kokon tertinggi terdapat pada perlakuan P+E+L (20.5±5.7 kokon), sedangkan produksi kokon terendah terdapat pada perlakuan (2.25±1.5 kokon) (Lampiran 5). Jumlah rata-rata kokon maksimum terdapat pada perlakuan spesies kombinasi E+L (95.75±17.3 kokon) setelah 2 minggu, sedangkan jumlah rata-rata kokon minimum terdapat pada perlakuan P (2±2.83 kokon) (Lampiran 6). Pada minggu ketiga jumlah kokon yang diproduksi oleh seluruh cacing tanah berkurang dari minggu sebelumnya, sedangkan di akhir percobaan tidak ditemukan kokon yang diproduksi pada seluruh perlakuan spesies tunggal dan spesies kombinasi.
Jumlah rata-rata kokon tertinggi dengan rasio R2 pada perlakuan spesies tunggal terdapat pada perlakuan E (34±13.7 kokon) setelah 1 minggu, sedangkan jumlah rata-rata kokon terendah terdapat pada perlakuan P (2.25±1.3 kokon) (Gambar 11; Lampiran 5). Pada rasio R2, seluruh perlakuan tidak menunjukkan peningkatan produksi kokon pada minggu kedua (Lampiran6). Produksi kokon berhenti pada perlakuan P dan P+E pada minggu ketiga, dan di akhir masa percobaan tidak ditemukan lagi kokon pada seluruh perlakuan.
Kokon-kokon yang diproduksi oleh cacing tanah pada seluruh perlakuan dipisahkan berdasarkan spesiesnya. Kokon yang diproduksi oleh Pheretima sp., E. fetida, dan L. rubellus memiliki ukuran dan bentuk yang bervariasi (Gambar 12 a-c). Kokon E. fetida berukuran lebih besar dibandingkan dengan kokon L. rubellus dan Pheretima sp., yaitu 4.85 mm x 2.82 mm, 3.50 mm x 2.46 mm, dan 3.90 mm x 1.85 mm berturut-turut (ukuran: panjang x lebar).
Waktu (minggu)
Gambar 10 Jumlah rata-rata kokon yang diproduksi pada perlakuan spesies yang berbeda dengan rasio antara cacing tanah dan sampah dedaunan 1:1 (R1).
Gambar 11 Jumlah rata-rata kokon yang diproduksi pada perlakuan spesies yang berbeda dengan rasio antara cacing tanah dan sampah dedaunan 2:1 (R2).
0
Gambar 14 Jumlah total kokon yang diproduksi oleh masing-masing spesies cacing tanah dengan rasio antara cacing tanah dan sampah dedaunan 2:1 (R2).
Gambar 13 Jumlah total kokon yang diproduksi oleh masing-masing spesies cacing tanah dengan rasio antara cacing tanah dan sampah dedaunan 1:1 (R1).
Gambar 12 Variasi ukuran dan bentuk kokon cacing tanah: Pheretima sp. (a), E. fetida (b), dan L. rubellus (c).
Gambar 16 Jumlah rata-rata juvenil yang diproduksi oleh masing-masing kokon cacing tanah pada suhu 25 oC.
Pheretima sp. E. fetida L. rubellus
Jum
Pheretima sp. E. fetida L. rubellus
Mas
Gambar 15 Masa inkubasi rata-rata kokon dari tiga spesies cacing tanah pada suhu 25 oC.
Setelah diinkubasi di dalam wadah lain dengan suhu 25 oC, kokon masing-masing spesies cacing tanah menetas pada waktu yang berbeda. Masa inkubasi kokon cacing tanah berkisar 14-21 hari. Kokon Pheretima sp. lebih cepat menetas daripada kokon E. fetida dan L. rubellus. Secara berurutan, rata-rata waktu yang dibutuhkan kokon untuk menetas adalah 15.6±1.1, 18.6±2.9, dan 19±3.0 hari (Gambar 15).
Komposisi Kimiawi Vermikompos
Selama 4 minggu masa percobaan, proses vermicomposting menghasilkan vermikompos. Komposisi kimiawi vermikompos berbeda dari substrat awal (Tabel 2). Namun hasil ini hanya menunjukkan komposisi kimiawi vermikompos pada spesies tunggal dengan rasio R2.
Berdasarkan pada hasil analisa, rasio C:N vermikompos lebih rendah dibandingkan dengan substrat awal (P=0.108), sedangkan kadar P dan K vermikompos lebih tinggi dibandingkan dengan substrat yang belum terdekomposisi. Kadar P pada vermikompos yang diproduksi oleh Pheretima sp. berbeda nyata dari substrat awal (P=0.03). Peningkatan kadar K yang nyata terdapat pada vermikompos yang diproduksi oleh Pheretima sp. dan L. rubellus (P<0.05) (Lampiran 12, 13, 14).
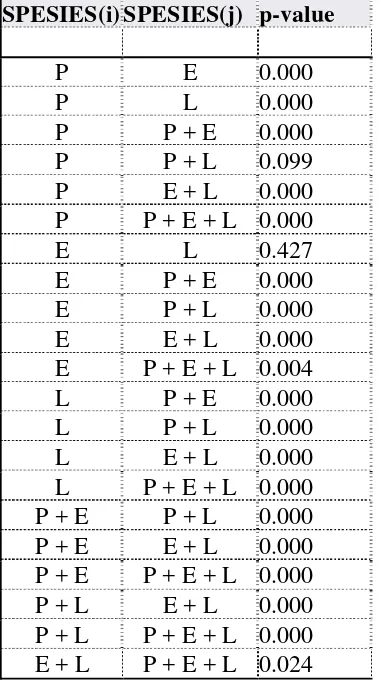
Proses vermicomposting menggunakan spesies cacing tanah yang berbeda pada umumnya menghasilkan vermikompos dengan komposisi kimiawi yang berbeda. Hasil analisis dengan ANOVA menunjukkan bahwa rasio C:N, kadar P dan K pada vermikompos yang dihasilkan oleh ketiga spesies cacing tanah tidak berbeda nyata (P>0.05).
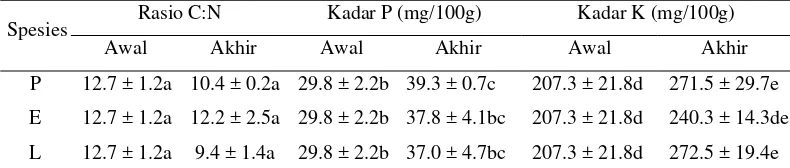
Tabel 2 Perbandingan komposisi kimiawi substrat awal dengan vermikompos yang dihasilkan oleh spesies Pheretima sp., E. fetida, dan L. rubellus
Huruf yang sama di belakang angka dalam baris atau kolom yang sama menunjukkan tidak berbeda nyata dengan uji Tukey pada tingkat kepercayaan 95%.
Rasio C:N Kadar P (mg/100g) Kadar K (mg/100g) Spesies
Awal Akhir Awal Akhir Awal Akhir
P 12.7 ± 1.2a 10.4 ± 0.2a 29.8 ± 2.2b 39.3 ± 0.7c 207.3 ± 21.8d 271.5 ± 29.7e
E 12.7 ± 1.2a 12.2 ± 2.5a 29.8 ± 2.2b 37.8 ± 4.1bc 207.3 ± 21.8d 240.3 ± 14.3de
PEMBAHASAN
Laju konsumsi bahan organik
Berdasarkan pada hasil penelitian, laju konsumsi pada rasio R2 lebih tinggi dibandingkan dengan rasio R1 (Gambar 6 & 7). Secara ringkas, laju konsumsi oleh ketiga spesies cacing tanah pada perlakuan spesies tunggal adalah E > L > P, sedangkan laju konsumsi pada perlakuan kombinasi adalah: E+L > P+E+L > P+E > P+L. Perbedaan laju konsumsi ini disebabkan oleh perbedaan densitas cacing tanah di dalam wadah. Semakin besar densitas cacing tanah, maka semakin cepat aktivitas konsumsi bahan organik. Densitas cacing tanah yang besar meningkatkan laju konsumsi pada permulaan. Ketika makanan berkurang aktivitas cacing tanah semakin rendah. Hasil ini sama seperti penelitian Sinha et al. (2002). Densitas cacing tanah yang besar menyebabkan peningkatan populasi mikroorganisme di dalam media biak dan saluran pencernaan cacing tanah selama proses vermicomposting (Banu et al. 2008). Sebagai dekomposer, cacing tanah dapat meningkatkan pertumbuhan mikroorganisme. Oleh karena itu, kehadiran cacing tanah dapat menyebabkan volume bahan organik yang dikonsumsi lebih banyak, sehingga pada gilirannya meningkatkan persentase laju konsumsi (Stachell et al 1984).
Selain itu, mikroorganisme menghasilkan beberapa enzim di dalam saluran pencernaan cacing tanah. Enzim-enzim tersebut terdiri atas, protease, selulase, kitinase, amilase dan lipase (Edwards & Lofty 1972). Enzim-enzim tersebut menguraikan komponen-komponen makanan yang masuk ke dalam saluran pencernaan untuk digunakan oleh jaringan tubuh cacing tanah. Parle (1963) menemukan bahwa enzim pencernaan seperti selulase lebih banyak terdapat di bagian tengah saluran pencernaan, sedangkan kitinase di bagian belakang.
Peningkatan biomassa fungi di dalam saluran pencernaan cacing tanah juga ditemukan oleh Aira et al. (2006b). Actinomycetes dan bakteri lebih banyak terdapat di bagian belakang saluran pencernaan dibandingkan dengan bagian tengah dan depan. Jumlah populasi bakteri yang terdapat di seluruh bagian saluran pencernaan cacing tanah lebih banyak daripada actinomycetes (Parle 1963).
Jenis dan jumlah mikroorganisme yang berperan di dalam sistem pencernaan setiap spesies cacing tanah berbeda-beda. Marialigeti (1979) menemukan sejumlah bakteri Vibro sp. di dalam saluran pencernaan cacing tanah Eisenia lucens, dan Aeromonas hydrophilia di dalam saluran pencernaan Pheretima sp. Jumlah mikroorganisme yang terdapat di dalam saluran pencernaan cacing tanah yang hidup di permukaan tanah lebih banyak dibandingkan dengan cacing tanah yang hidup pada tanah yang lebih dalam. Kozlovskaya dan Zhdannikova (1961) menemukan bahwa jumlah total bakteri yang terdapat di dalam saluran pencernaan cacing tanah L. rubellus 10 kali lebih banyak daripada yang terdapat di dalam saluran pencernaan cacing tanah Octolasium lacteum.
Faktor lain yang dapat mempengaruhi laju konsumsi adalah jenis dan kualitas bahan makanan yang juga merupakan media biak. Pada penelitian ini, media biak dan bahan makanan yang digunakan adalah kotoran sapi dan sampah daun sonokeling (D. latifolia). Hasil penelitian menunjukkan bahwa laju konsumsi semakin rendah setelah 2 minggu (Gambar 6 & 7). Laju konsumsi yang semakin rendah dikarenakan berkurangnya nutrisi yang berasal dari kotoran sapi sebagai media biak sekaligus sumber Nitrogen dan rendahnya palatabilitas daun sonokeling bagi cacing tanah. Sehubungan dengan hal ini, Sinha et al. (2002) menyatakan bahwa cacing tanah mengkonsumsi sedikit bahan makanannya jika tidak tersedia substrat utama seperti kotoran ternak. Selain itu, cacing tanah memiliki palatabilitas yang rendah terhadap sampah dedaunan (Satchell & Lowe 1967).
dikeluarkan sebagai vermicast. Vermicast tercampur dengan getah mukus dinding saluran pencernaan cacing tanah dan mikrob, selanjutnya diubah menjadi vermikompos (Edwards & Lofty 1972).
Spesies cacing tanah yang digunakan juga dapat mempengaruhi laju konsumsi. Spesies cacing tanah yang digunakan pada penelitian ini adalah Pheretima sp., E. fetida, dan L. rubellus. Berdasarkan pada pengamatan, perlakuan spesies tunggal E. fetida mengkonsumsisi bahan makanannya lebih tinggi dibandingkan dengan spesies tunggal L. rubellus dan Pheretima sp. (Gambar 6 & 7), sedangkan pada perlakuan spesies kombinasi, perlakuan E. fetida + L. rubellus menunjukkan laju konsumsi yang lebih tinggi daripada perlakuan kombinasi lainnya. Hal ini dikarenakan karakteristik spesifik spesies cacing tanah yang berbeda-beda. E. fetida dan L. rubellus merupakan spesies cacing tanah epigeic yang pada dasarnya merupakan cacing tanah pemakan sampah dan kotoran pada permukaan tanah (Lee 1985), sehingga daya konsumsinya lebih tinggi daripada Pheretima sp. yang belum teridentifikasi status ekologinya. E. fetida dan L. rubellus merupakan spesies cacing tanah yang paling umum digunakan dalam proses vermicomposting, karena keduanya telah terbukti mendekomposisi sampah organik dengan cepat (Edwards & Lofty 1972).
Pertumbuhan Cacing Tanah
penelitian Barley (1959). Seluruh spesies cacing tanah lebih menyukai bahan makan berupa kotoran hewan atau daun rumput-rumputan yang berair daripada daun pepohonan (Guild 1955). Jenis dan jumlah bahan makanan yang tersedia pada media biak tidak hanya mempengaruhi pertumbuhan cacing tanah tetapi juga produktivitasnya (Edwards & Lofty 1972).
Di lain pihak, pada rasio antara cacing tanah dan sampah dedaunan 2:1 (R2), biomassa cacing tanah pada seluruh perlakuan berkurang sejak minggu pertama (Gambar 9). Fakta ini mengindikasikan bahwa densitas cacing tanah dan ketersediaan bahan makanan di dalam media biak mempengaruhi pertumbuhan cacing tanah. Densitas cacing tanah yang tinggi dan ketersediaan substrat yang tidak mencukupi kebutuhan dapat menurunkan laju pertumbuhan cacing tanah (Nauhauser et al. 1980; Dominguez et al. 1997b; Garg et al. 2005).
Pertumbuhan cacing tanah dengan rasio R2 lebih rendah dibandingkan dengan rasio R1 (Gambar 8 & 9). Hal ini disebabkan oleh tingginya densitas cacing tanah pada rasio R2. Densitas cacing tanah yang tinggi pada media biak menyebabkan tingginya kompetisi makanan (Dominguez et al. 1997b), sehingga cacing tanah kekurangan sumber makanan. Reinecke et al. (1992) juga menemukan rendahnya pertumbuhan cacing tanah karena densitasnya yang tinggi. Biomassa cacing tanah berkurang karena berkurangnya bahan makanan yang diterima oleh setiap individu cacing tanah.
Pertumbuhan cacing tanah bergantung pada kualitas makanannya. Kualitas bahan organik yang dikonsumsi cacing tanah merupakan salah satu faktor yang menentukan laju pertumbuhan dan reproduksi (Dominguez et al. 2000; Suthar 2007b). Cacing tanah yang mengkonsumsi bahan makanan dengan rasio C:N rendah, lebih banyak menggunakan energinya untuk pertumbuhan daripada reproduksi (Aira et al. 2006a). Oleh karena itu, kualitas bahan makanan yang digunakan dalam proses vermicomposting sangat mempengaruhi biomassa cacing tanah (Suthar 2007a).
individu spesies cacing tanah. Hasil ini memperkuat penelitian Suthar dan Singh (2008) yang menemukan perbedaan pertumbuhan cacing tanah Perionyx excavatus dan P. sansibaricus (Megascolecidae) yang mengkonsumsi bahan organik dari sampah rumah tangga. Perbedaan pertumbuhan cacing tanah disebabkan oleh perbedaan spesiesnya (Garg et al. 2005).
Selain itu, pertumbuhan yang berbeda pada spesies cacing tanah dapat disebabkan oleh interaksi diantara populasi cacing tanah akibat kombinasi spesies tersebut dalam proses vermicomposting. Berdasarkan pada hasil pengamatan, setiap perlakuan kombinasi yang terdiri atas spesies E. fetida dan L. rubellus menunjukkan pola pertumbuhan yang lebih tinggi dibandingkan dengan perlakuan kombinasi yang berisi Pheretima sp. Pada percobaan skala laboratorium,interaksi inter-spesifik dan intra-spesifik diantara populasi cacing tanah L. terrestris, L. rubellus, Aporrectodea longa, dan Allolobophora chlorotica berpengaruh negatif terhadap laju pertumbuhan dan produktivitas. Keberadaan satu spesies dapat menghambat pertumbuhan dan produktivitas spesies yang lain (Lowe & Butt 2002). Hal itu terjadi karena adanya kompetisi antara spesies epigeic dan aneic dalam memperoleh sumber makanan.
Produktivitas Cacing Tanah
Pada umumnya produktivitas cacing tanah dapat digambarkan dengan laju produksi kokon dan jumlah juvenil yang muncul dari masing-masing kokon. Hasil penelitian ini menunjukkan bahwa produksi kokon tertinggi terjadi pada minggu kedua pada perlakuan spesies kombinasi E. fetida + L. rubellus (Gambar 10). Hal ini mungkin disebabkan oleh ketersediaan bahan makanan yang cukup dari minggu pertama, dan laju konsumsi yang tinggi dari minggu pertama sampai minggu kedua. Oleh karena itu cacing tanah memiliki energi yang banyak untuk memproduksi kokon. Produksi kokon dipengaruhi oleh jumlah bahan makanan yang tersedia di dalam media biak (Edwards & Lofty 1972).
digunakan untuk memproduksi kokon (Garg et al. 2005). Rendahnya produksi kokon pada rasio R2 disebabkan oleh pengaruh densitas cacing tanah pada media biak. Pada rasio R2, densitas cacing tanah tinggi, sehingga jumlah makanan yang dikonsumsi lebih sedikit, akibatnya pertumbuhan menjadi rendah dan produksi kokon berkurang. Densitas cacing tanah yang tinggi menyebabkan adanya kompetisi makanan, sehingga cacing tanah kekurangan energi untuk memproduksi kokon. (Dominguez et al. 1997b).
Pada minggu keempat, produksi kokon berhenti pada semua perlakuan (Gambar 10 & 11). Hal ini disebabkan oleh berkurangnya sumber nutrisi berupa Nitrogen bagi cacing tanah yang berasal dari kotoran sapi. Fakta ini membuktikan bahwa ketersediaan makanan pada media biak sangat mempengaruhi produksi kokon. Ketika kokon tidak diproduksi lagi, energi cacing tanah dimanfaatkan untuk pertumbuhan jaringan tubuh (Chaudhari & Bhattacharjee 2002).
Berdasarkan pada pengamatan, kokon yang diproduksi oleh ketiga spesies cacing tanah memiliki ukuran dan bentuk yang bervariasi. Kokon E. fetida lebih besar daripada kokon L. rubellus dan Pheretima sp. (Gambar 12 a-c). Chaudhari dan Bhattacharjee (2002) yang meneliti beberapa spesies cacing tanah (Eutyphoeus gammiei, Lampito mauritii, Pontoscolex corethurus, Polypheretima elongata, Drawida napalensis, Perionyx excavatus, dan Dischogaster modiglianii), menemukan bahwa ukuran dan bentuk kokon bervariasi diantara spesies cacing tanah tersebut. Senapati dan Sahu (1993) menemukan hubungan yang positif antara ukuran tubuh cacing tanah dewasa dengan ukuran kokon yang diproduksi. Penemuan ini sesuai dengan hasil pengamatan. Spesies E. fetida berukuran lebih panjang (± 9 cm) dan menghasilkan kokon lebih besar (4.85 mm x 2.82 mm), dibandingkan dengan ukuran tubuh spesies L. rubellus (± 5 cm) yang menghasilkan kokon sebesar 3.50 mm x 2.46 mm. Walaupun demikian, ukuran kokon tidak selalu berhubungan dengan ukuran tubuh cacing tanah dewasa (Edwards & Bohlen 1996).
kisaran suhu 24-32 oC dibandingkan dengan cacing tanah anecic (Bhattacharjee & Chaudhuri 2002). Namun produksi kokon tidak dipengaruhi oleh ukuran tubuh cacing tanah. Senapati dan Sahu (1993) menemukan bahwa ukuran tubuh cacing tanah memperlihatkan hubungan negatif terhadap produksi kokon.
Masa inkubasi kokon berbeda-beda pada masing-masing spesies cacing tanah. Masa inkubasi rata-rata kokon Pheretima sp. lebih cepat dibandingkan dengan masa inkubasi kokon E. fetida dan L. rubellus (Gambar 15). Walaupun demikian, masa inkubasi masing-masing kokon cacing tanah pada penelitian ini tidak berbeda nyata (P=0.09). Masa inkubasi kokon dapat dipengaruhi oleh suhu lingkungan. Pada penelitian ini, kokon diinkubasi pada suhu 25 oC. Masing-masing kokon menetas pada kisaran 14-21 hari. Suhu yang lebih tinggi daripada 25 oC menurunkan masa inkubasi rata-rata kokon cacing tanah epigeic (Reinecke et al. 1992). Peningkatan suhu menyebabkan peningkatan masa inkubasi pada cacing tanah endogeic dan penurunan masa inkubasi pada cacing tanah epigeic. Pada cacing tanah anecic, masa inkubasi kokonnya panjang (110 hari). Pada cacing tanah endogeic, masa inkubasi kokon pendek (15 hari), sedangkan pada cacing tanah epigeic masa inkubasi kokon sangat pendek (13-14 hari) (Bhattacharjee & Chaudhuri 2002).
Komposisi Kimiawi Vermikompos
Karakteristik vermikompos bergantung pada karakteristik awal bahan mentahnya. Bahan mentah vermikompos pada penelitian ini adalah kotoran sapi dan sampah daun sonokeling dengan rasio C:N 12.7±1.2, kadar P 29.8±2.2 mg/100g, dan kadar K 207.3±21.8 mg/100g (Tabel 2).
Berdasarkan pada hasil analisa, rasio C:N vermikompos lebih rendah daripada substrat awal (Tabel 2), namun perbedaan ini tidak nyata (P=0.10). Hal ini mungkin disebabkan oleh masa vermicomposting yang relatif singkat, sehingga vermikompos yang dihasilkan belum cukup matang. Beberapa peneliti juga menemukan penurunan tingkat rasio C:N vermikompos (Atiyeh et al. 2000; Tripathi & Bhardwaj 2004; Suthar & Singh 2008; Khwairakpam & Bhargava 2009). Rasio C:N pada substrat menggambarkan mineralisasi dan stabilisasi sampah organik selama proses vermicomposting (Suthar & Singh 2008). Rasio C:N menunjukkan tingkat dekomposisi sampah organik. Unsur C dilepaskan sebagai CO2 selama biooksidasi, sedangkan N dilepaskan pada tingkat yang lebih rendah. Semakin banyak bahan organik yang terdekomposisi, semakin rendah rasio C:N (Pramanik et al. 2007; Lazcano et al. 2008). Beberapa penelitian menunjukkan bahwa rasio C:N merupakan salah satu indikator yang paling sering digunakan untuk mengetahui kematangan vermikompos (Tripathi & Bhardwaj 2004; Loh et al. 2005).
Menurunnya rasio C:N pada vermikompos disebabkan oleh aktivitas cacing tanah dalam proses vermicomposting. Penurunan rasio C:N menunjukkan tingginya humifikasi vermikompos (Albanell et al. 1988). Hilangnya karbon sebagai karbon dioksida pada proses respirasi dan produksi mukus serta kotoran bernitrogen dapat menurunkan rasio C:N (Suthar 2007b). Pramanik et al. (2007) menemukan bahwa kotoran sapi dapat menurunkan rasio C:N vermikompos.