POLIMORFISME GEN β
-KASEIN DAN LAKTOFERIN
SERTA HUBUNGANNYA DENGAN PROTEIN
SUSU PADA SAPI FRIESIAN-HOLSTEIN
ELMY MARIANA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS
DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis yang berjudul Polimorfisme Gen
Β-Kasein dan Laktoferin serta Hubungannya dengan Protein Susu pada Sapi Friesian-Holstein adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar Pustaka di bagian akhir tesis ini.
Bogor, Desember 2010
ABSTRACT
ELMY MARIANA. Association of The β-Casein and Lactoferrin Genes Polymhorphisms with Milk Protein in Friesian Holstein Cows. Under direction of CECE SUMANTRI, ANNEKE ANGGRAENI, RARAH R. A. MAHESWARI
The purposes of this study were to identify the polymorphism of the β-casein
and the lactoferrin genes and the association of genetic variants of the β-casein and the lactoferrin genes on protein yield, milk yield and SCC in Holstein-Friesian (HF) cows. The study was conducted on 282 heads of HF lactating cows coming from two managements of: 1) intensive at dairy breeding station of BPPT-SP Cikole, and 2) semi intensive of small dairy farmers in two villages of the KPSBU Lembang, Lembang District. Investigation on variant genotypes of the β-casein and the lactoferrin genes used PCR-RFLP method. Genotyping the β-casein gene with MspI restriction enzyme to all DNA samples produced only the BB genotype (100%), meaning this gene was monomorphic. While genotyping the lactoferrin gene with EcoRI restriction enzyme produced two genotypes, i.e. AA (65%) and AB (35%) genotypes. The AB genotype of the lactoferrin gene tended to produce higher milk protein than AA genotype, though the differences were statistically not significant (P>0.05). Both the AA and AB genotypes had inconsistency in controlling milk fat and milk yield. Further, the AB lactoferrin cows had a higher frequency in the classification of low somatic cells count (0.75), while the AA cows were at a higher frequency in the classification of high somatic cells count (0.60). It was concluded
that the β-casein gene studied was monomorphic, so the effect in controlling milk protein had still not been known, while the AB genotype of the lactoferrin gene had a consistency in producing higher milk protein and lower somatic cells count compared to the AA genotype.
RINGKASAN
ELMY MARIANA. Polimorfisme Gen β-Kasein dan Laktoferin serta Hubungan-nya dengan Protein Susu pada Sapi Friesian-Holstein. Dibimbing oleh CECE SUMANTRI, ANNEKE ANGGRAENI, RARAH R. A. MAHESWARI
Perbaikan genetik sapi perah domestik dapat dilakukan antara lain melalui seleksi keunggulan pada gen yang diprediksi berasosiasi kuat dengan sifat produksi, kualitas susu dan ketahanan terhadap penyakit, khususnya seleksi untuk gen-gen yang mengontrol protein susu dan resistensi terhadap penyakit. β-kasein merupakan salah satu jenis dari fraksi kasein yang berpengaruh terhadap bentuk dan kestabilan butiran susu dan memiliki kontribusi sebesar 28.4% dari total protein susu, sedangkan laktoferin merupakan salah satu fraksi whey yang merupakan protein antimikrobia dalam susu yang berfungsi sebagai imunitas untuk melawan berbagai macam penyakit. Pemeriksaan dua atau lebih gen-gen protein susu (kasein dan whey protein) secara bersamaan bermanfaat untuk memperhitungkan pengaruh interaksi yang mungkin terjadi antara dua atau lebih gen haplotipe. Penelitian ini bertujuan untuk mengidentifikasi polimorfisme gen β-kasein dan laktoferin dan mengetahui konsistensi hubungan antara genotipe gen β-kasein dan laktoferin baik secara masing-masing maupun interaksi kedua gen dengan protein susu dan somatic cell count(SCC) pada sapi Friesian-Holstein (FH) laktasi pada dua kondisi pemeliharaan berbeda, yaitu intensif di stasiun bibit sapi perah BPPT-SP Cikole dan semi intensif di peternakan rakyat di bawah binaan KPSBU Lembang.
Data produksi dan kualitas susu merupakan data sekunder yang telah diambil pada bulan Agustus 2008. Data produksi susu merupakan produksi satu hari yang diperoleh dengan menjumlahkan produksi pagi dan sore dari sapi laktasi dalam kisaran lama laktasi antara 1 sampai 6 bulan dan periode laktasi antara 1 sampai 5. Data kualitas susu yang diamati adalah kadar protein, lemak, BK, BKTL dan BJ. Pengujian setiap komponen kualitas susu segar mengikuti standar uji pada Dewan Standarisasi Nasional (1998). Sampel yang digunakan sebanyak 282 ekor, untuk BPPT Cikole yang mewakili kondisi pemeliharaan intensif sebanyak 88 ekor, KPSBU Lembang yang mewakili kondisi pemeliharaan semi intensif sebanyak 194 ekor yaitu dari peternakan rakyat desa Pasir Kemis 96 ekor dan desa Cilumber sebanyak 98 ekor. DNA yang digunakan untuk identifikasi polimorfisme gen β -kasein dan laktoferin merupakan koleksi dari Laboratorium Pemuliaan dan Genetika Ternak Departemen Ilmu Produksi dan Teknologi Peternakan, Institut Pertanian Bogor. Isolasi DNA dilakukan dari darah dan sel somatik susu. Isolasi DNA menggunakan metode ekstraksi phenol-chloroform (Sambrook et al. 1989) dengan modifikasi untuk sampel darah yang disimpan dalam alkohol. Isolasi DNA dari sampel susu menggunakan metode ekstraksi DNA dari sel somatik (Arnould et al. 2009). Identifikasi polimorfisme gen β-kasein dan laktoferin dilakukan melalui metode PCR-RFLP. Keragaman gen ditentukan dengan analisis frekuensi alel,
frekuensi genotipe dan nilai heterozigositas. Keseimbangan genotipe gen β-kasein
Amplifikasi gen β-kasein dan laktoferin menghasilkan ruas gen target
sepanjang 233 pb untuk gen β-kasein dan 301 pb untuk gen laktoferin. Penentuan genotipe (genotyping) gen β-kasein dan laktoferin menggunakan metode Restriction Fragment Length Polymorphism (RFLP) dengan enzim pemotong MspI untuk gen β -kasein dan EcoRI untuk gen laktoferin. Genotyping gen β-kasein lokus MspI pada exon 7 menghasilkan hanya satu varian genotipe yaitu BB dengan panjang fragmen 233 pb (100%), menunjukkan bahwa gen β-kasein lokus MspI bersifat seragam (monomorfic). Genotyping gen laktoferin lokus EcoRI pada intron 6 menghasilkan dua varian genotipe yaitu AA dengan panjang fragmen 301 pb (0.655%) dan AB dengan panjang fragmen 100, 201 dan 301 pb (0.345%).
Rataan nilai heterozigositas gen laktoferin pada dua kondisi pemeliharaan adalah 0.2856. Rataan nilai heterozigositas gen laktoferin tergolong rendah, hal ini mengindikasikan rendahnya variasi gen laktoferin dalam populasi. Semakin tinggi nilai heterozigositas dalam populasi maka semakin besar peluang untuk melakukan seleksi pada populasi tersebut. Pengujian keseimbangan genotipe menggunakan proporsi Hardy-Weinberg menghasilkan frekuensi genotipe diantara lokus gen
laktoferin sangat berbeda nyata (χ2 =12.2277, df = 2, P=0.0005) yang artinya dalam populasi tidak terjadi keseimbangan Hardy-Weinberg. Hal ini disebabkan pada populasi sapi perah terjadi seleksi dan tidak terjadi sistem perkawinan random mating yang menjadi salah satu syarat terpenuhinya hukum keseimbangan Hardy-Weinberg. Proses perkawinan ternak perah menggunakan aplikasi kawin buatan (Inseminasi buatan) dengan sperma dari pejantan unggul tertentu.
Hubungan antara genotipe laktoferin dengan kualitas susu sapi FH menunjukkan bahwa sapi dengan genotipe AB secara konsisten cenderung menghasilkan kadar protein susu lebih tinggi dari genotipe AA meskipun perbedaannya tidak nyata. Hal ini disebabkan sifat kadar protein susu merupakan sifat yang dikendalikan oleh polygenes (banyak gen). Genotipe AA secara konsisten cenderung menghasilkan kadar BKTL lebih tinggi dari sapi dengan genotipe AB. Genotipe AA maupun AB tidak memiliki konsistensi dalam mengontrol kadar lemak dan produksi susu.
Hubungan antara genotipe laktoferin dengan SCC menunjukkan bahwa sapi dengan genotipe AB mempunyai frekuensi yang tinggi pada klasifikasi total sel somatik susu rendah (0.75%) dan genotipe AA pada klasifikasi total sel somatik susu tinggi (0.60%). Varian genotipe AB pada gen laktoferin memiliki kecenderungan menghasilkan angka sel somatik yang lebih rendah jika dibandingkan dengan varian AA.
Genβ-kasein lokus MspIbersifat monomorfik sehingga hubungannya dengan protein susu tidak diketahui, sedangkan genotipe AB gen laktoferin memiliki konsistensi dalam menghasilkan kadar protein yang lebih tinggi dan nilai SCC yang lebih rendah jika dibandingkan dengan genotipe AA.
© Hak Cipta milik IPB, tahun 2010
Hak Cipta dilindungi Undang-Undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya.
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah.
b. Pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
POLIMORFISME GEN β
-KASEIN DAN LAKTOFERIN
SERTA HUBUNGANNYA DENGAN PROTEIN
SUSU PADA SAPI FRIESIAN-HOLSTEIN
ELMY MARIANA
Tesis
sebagai salah satu syarat untuk memperoleh gelar
Magister Sains pada
Program Studi Ilmu Produksi dan Teknologi Peternakan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
HALAMAN PENGESAHAN
Judul Tesis : Polimorfisme Gen β-Kasein dan Laktoferin serta Hubungannya dengan Protein Susu pada Sapi Friesian-Holstein
Nama : Elmy Mariana
NIM : D151080061
Disetujui Komisi Pembimbing
Prof. Dr. Ir. Cece Sumantri. M.Agr.Sc. Ketua
Ir. Anneke Anggraeni, M.Si, Ph.D Dr. Ir. Rarah R. A. Maheswari, DEA Anggota Anggota
Mengetahui
Ketua Mayor Dekan Sekolah Pascasarjana IPB
Ilmu Produksi dan Teknologi Peternakan
Dr. Ir. Rarah R. A. Maheswari, DEA Prof. Dr. Ir. Khairil A. Notodiputro, M.S
PRAKATA
Alhamdulillah puji syukur penulis panjatkan kehadirat Allah SWT atas segala nikmat dan karunia-Nya yang telah diberikan sehingga penulis mendapat kemudahan dalam menyusun dan menyelesaikan tesis ini. Tesis ini merupakan salah satu syarat untuk memperoleh gelar Magister Sains pada Program Studi Ilmu Produksi dan Teknologi Peternakan, Institut Pertanian Bogor. Tesis ini berjudul Polimorfisme Gen
β-Kasein dan Laktoferin serta Hubungannya dengan Protein Susu pada Sapi Friesian-Holstein. ”
Penulis mengucapkaan terima kasih kepada Prof. Dr. Ir. Cece Sumantri, M. Agr. Sc., Ir. Anneke Anggraeni, M. Si Phd dan Dr. Ir. Rarah Ratih Adji Maheswari, DEA. Selaku pembimbing, serta Dr. Jakaria, S. Pt, M. Si yang telah banyak member saran. Disamping itu, penghargaan penulis sampaikan kepada seluruh staf laboratorium Genetika Molekuler Departemen Ilmu Produksi dan Teknologi Peternakan yang telah banyak membantu selama penulis melaksanakan penelitian.
Ucapan terima kasih juga penulis sampaikan kepada kedua orang tua dan seluruh keluarga yang telah mengiringi setiap langkah penulis dengan doa dan restu yang tidak terputus. Penghargaan penulis sampaikan kepada suami (Amran M. Yusuf Adji) dan putra (Aji Rahmatullah Amran) yang selalu memberi dukungan, doa dan cinta kepada penulis. Terima kasih juga penulis ucapkan kepada semua pihak yang tidak dapat disebutkan satu persatu atas bantuan dan dukungannya. Penulis berharap semoga karya ilmiah ini dapat bermanfaat bagi upaya pengembangan keilmuan dan pembangunan peternakan di Indonesia.
Bogor, Desember 2010
RIWAYAT HIDUP
Penulis lahir di Tulungagung pada tanggal 9 September 1979 dari ayah Sumani dan ibu Istikomah. Penulis merupakan anak bungsu dari 3 bersaudara. Pendidikan sarjana ditempuh di jurusan Teknologi Hasil Ternak, Fakultas Peternakan Universitas Gadjah mada, lulus pada tahun 2003. Selama mengikuti perkuliahan penulis menjadi asisten pada jurusan Teknologi Hasil Ternak. Pada tahun 2008 penulis mendapatkan kesempatan untuk melanjutkan studi pada Program Pascasarjana Institut Pertanian Bogor. Beasiswa pendidikan pascasarjana diperoleh dari Departemen Pendidikan Republik Indonesia melalui program BPPS.
Penulis bekerja sebagai staf pengajar pada program studi Produksi Ternak, Jurusan Peternakan, Fakultas Pertanian Universitas Syiah Kuala sejak tahun 2006. Mata kuliah yang menjadi tanggung jawap penulis adalah Dasar Ternak Perah dan Ilmu Produksi Ternak Perah.
DAFTAR ISI
Penentuan Genotipe (Genotyping) ... 19
Pengukuran Produksi Susu ... 19
Analisis Kualitas Susu ... 20
Penghitungan Jumlah Sel Somatik dalam Susu... 20
Rancangan dan Analisis Data ... 21
Frekuensi Alel dan Genotipe ... 21
Derajat Heterozigositas ... 21
Proporsi Hardy-Weinberg ... 21
Pengaruh Manajemen Pemeliharaan terhadap Produksi dan Kualitas Susu ... 22
Pengaruh Genotipe β-Kasein dan Laktoferin terhadap Protein dan Kualitas Susu... 22
Kualitas Susu ... 27
Kadar Protein Susu ... 28
Kadar Lemak Susu... 30
Amplifikasi Gen β-Kasein... 32
Identifikasi Keragaman Gen β-Kasein... 33
Amplifikasi Gen Laktoferin ... 35
Identifikasi Keragaman Gen Laktoferin ... 36
Frekuensi Alel dan Frekuensi Genotip Gen Laktoferin ... 37
Derajat Heterozigositas ... 38
Proporsi Hardy-Weinberg ... 39
Pengaruh Genotipe laktoferin pada Protein dan Kualitas Susu... 40
Kadar Protein Susu ... 42
Parameter Kualitas Susu yang lain ... 43
Pengaruh Genotipe Gen Laktoferin terhadap Produksi Susu... 44
Pengaruh Genotipe Gen Laktoferin terhadap Jumlah Sel Somatik (SCC) ... 45
KESIMPULAN DAN SARAN Kesimpulan ... 48
Saran ... 48
DAFTAR PUSTAKA ... 49
DAFTAR TABEL
Halaman
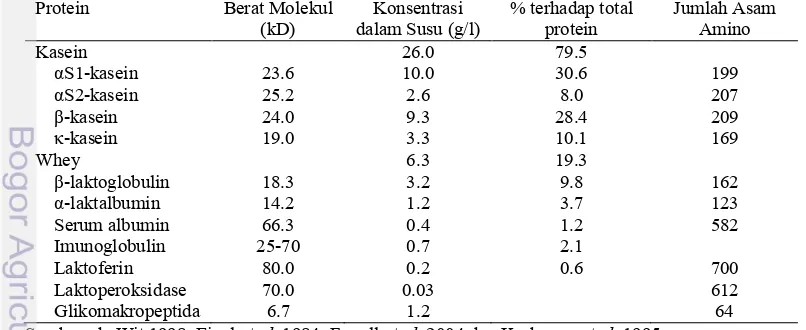
1 Karakteristik protein susu sapi ... 8
2 Perubahan sekuens asam amino pada variasi β-kasein... 11
3 Variasi gen β-kasein pada berbagai bangsa sapi FH ... 12
4 Informasi sekuen primer yang digunakan dalam penelitian ... 19
5 Nilai rataan produksi susu harian yang dikelompokkan berdasarkan bulan laktasi pada kondisi pemeliharaan yang berbeda... 24
6 Nilai rataan produksi susu harian yang dikelompokkan berdasarkan periode laktasi pada kondisi pemeliharaan yang berbeda... 26
7 Nilai rataan kualitas susu harian sapi FH pada kondisi pemeliharaan yang berbeda... 27
8 Nilai rataan protein susu harian yang dikelompokkan berdasarkan bulan laktasi pada kondisi pemeliharaan yang berbeda... 29
9 Nilai rataan protein susu harian yang dikelompokkan berdasarkan periode laktasi pada kondisi pemeliharaan yang berbeda... 29
10 Nilai rataan lemak susu harian yang dikelompokkan berdasarkan bulan laktasi pada kondisi pemeliharaan yang berbeda... 31
11 Nilai rataan lemak susu harian yang dikelompokkan berdasarkan periode laktasi pada kondisi pemeliharaan yang berbeda... 32
12 Frekuensi genotipe dan frekuensialel gen β-kasein lokus MspI pada sapi FH berdasarkan kondisi pemeliharaan yang berbeda ... 35
13 Frekuensi genotipe dan frekuensi alel dari gen laktoferin sapi FH pada kondisi pemeliharaan yang berbeda ... 38
14 Nilai heterozigositas (ĥ) gen laktoferin lokus EcoRIsapi FH pada kondisi pemeliharaan yang berbeda... 38
15 Hasil pengujian proporsi Hardy-Weinberg berdasarkan tes Chi-Square gen laktoferin lokus EcoRIpada kondisi pemeliharaan yang berbeda. ... 39
16 Pengaruh varian genotipe laktoferin terhadap sifat kualitas susu sapi FH berdasarkan kondisi pemeliharaan... 41
17 Pengaruh varian genotipe laktoferin terhadap produksi susu sapi FH berdasarkan kondisi pemeliharaan... 44
tinggi dan rendah pada sapi FH. ... 46
DAFTAR GAMBAR
Halaman 1 Diagram alir pelaksanaan penelitian... 32 Produksi susu selama masa laktasi dengan tingkat persistensi yang berbeda ... 4
3 Konsentrasi protein, lemak dan laktosa dalam susu selama masa laktasi... 7
4 Struktur lokus kasein pada sapi ... 9
5 Struktur gen β-kasein ... 10
6 Struktur gen laktoferin ... 14
7 Posisi penempelan primer (cetak tebal) pada sekuen gen β-kasein, C bergaris bawah menunjukkan mutasi pada posisi basa ke 8267 ... 32
8 Visualisasi hasil amplifikasi ruas gen β-kasein pada gel agarose 1,5% ... 33
9 Perbedaan sekuen gen β-kasein nomor akses gen bank AJ29330 dan X14711 yang disebabkan mutasi titik C – G... 33
10 Hasil elektroforesis gen β-kasein lokus MspIpada gel agarose 2%... 34
11 Posisi penempelan primer (cetak tebal) pada sekuen gen laktoferin... 36
12 Visualisasi hasil amplifikasi ruas gen laktoferin pada gel agarose 1,5% ... 36
13 Perbedaan sekuen gen laktoferin nomor akses gen bank AH000852S03 dan EU192148 yang disebabkan mutasi titik C – T ... 36
DAFTAR LAMPIRAN
Halaman
1 Informasi gen β-kasein pada sapi perah... 57
2 Informasi gen laktoferin pada sapi perah... 60
3 Metode isolasi DNA yang dimodifikasi…... 62
4 Metode isolasi DNA dari sampel sel somatik susu ... 62
5 Analisis kadar protein dengan metode titrasi formol (Standar Nasional Indonesia, 1992) ... 63
6 Analisis kadar lemak (Standar Nasional Indonesia, 1992) ... 64
7 Analisis berat jenis (Badan Standarisasi Nasional, 1992)... 64
8 Persamaan regresi antara bulan laktasi dengan parameter produksi dan kualitas susu…. ... 64
9 Persamaan regresi antara periode laktasi dengan parameter produksi dan kualitas susu…. ... 65
10 Uji sidik ragam parameter produksi dan kualitas susu berdasarkan genotipe laktoferin pada kondisi pemeliharaan intensif ... 65
11 Uji sidik ragam parameter produksi dan kualitas susu berdasarkan genotipe laktoferin pada kondisi pemeliharaan semi intensif ... 66
12 Uji sidik ragam parameter produksi dan kualitas susu berdasarkan genotipe laktoferin pada gabungan dua kondisi pemeliharaan ... 67
PENDAHULUAN
Latar Belakang
Perkembangan industri susu nasional sudah berjalan lebih dari dua dekade. Produksi susu olahan nasional pada tahun 2010 mencapai 5.600 ton per hari, namun untuk dapat berproduksi secara optimal industri pengolahan susu saat ini masih harus mengimpor 76,55% bahan baku yang dibutuhkannya karena produksi susu segar dalam negeri baru bisa memasok tidak lebih dari 23,45% dari permintaan nasional. Produksi susu dalam negeri perlu ditingkatkan sehingga secara bertahap dapat memenuhi kebutuhan bahan baku industri susu dalam negeri. Usaha untuk meningkatkan produksi susu dilakukan melalui beberapa cara antara lain: peningkatan populasi sapi perah, perbaikan manajemen pemeliharaan, perbaikan pakan, pengendalian penyakit dan seleksi bibit unggul.
Arah seleksi ternak perah adalah untuk menghasilkan sapi-sapi dengan sifat unggul produksi susu yang tinggi, kualitas susu yang baik dan ketahanan terhadap penyakit. Seleksi keunggulan genetik melalui identifikasi gen yang diprediksi berasosiasi kuat dengan kualitas susu dan ketahanan terhadap penyakit akan sangat mendukung bagi program perbaikan genetik sapi perah domestik.
Protein merupakan komponen susu yang sangat penting dan memiliki nilai ekonomi yang tinggi. Beberapa industri pengolahan susu saat ini mulai menginginkan untuk memproduksi susu dengan kadar protein dan kualitas yang lebih baik. Hal ini disebabkan perbaikan tingkat sosial masyarakat sehingga tren permintaan konsumen atas susu berprotein tinggi dan berkualitas saat ini semakin meningkat. Selain itu bertambahnya jumlah balita dan anak sekolah berdampak langsung pada meningkatnya kebutuhan susu berprotein tinggi. Mengantisipasi permintaan pasar yang berkembang lebih kearah produksi susu berkadar protein tinggi, diperlukan sapi perah yang berkemampuan memproduksi susu dengan kadar protein yang tinggi.
78-82%) yang terdiri atas empat lokus saling berdekatan (haplotipe) dengan runutan genom alfa s1-kasein, beta-kasein, alfa s2-kasein dan kapa-kasein (αs1-, β-, αs2- dan
κ-kasein) dengan panjang 250 kb pada kromosom 6/BTA 6q31. Keterkaitan lokasi yang sangat dekat antara keempat mayor gen kasein dan diwariskan lebih sebagai klaster, menjadikan kasein potensial untuk dieksplorasi sebagai marka pembantu seleksi sifat protein susu. Whey selain dikontrol oleh laktoferin pada kromosom 22 (22q24), juga dikontrol oleh tiga lokus lain yaituα-laktalbumin pada kromosom 5
(5q21), β-laktoglobulin pada kromosom 11 (11q28) dan lisozim pada kromosom 5 (5q22). β-kasein merupakan salah satu jenis dari fraksi kasein yang berpengaruh terhadap bentuk dan kestabilan butiran susu dan memiliki kontribusi sebesar 28.4% dari total protein susu. Laktoferin merupakan salah satu fraksi whey yang merupakan protein antimikrobia dalam susu yang berfungsi sebagai imunitas untuk melawan berbagai macam penyakit.
Banyak studi mempertimbangkan pengaruh gen kasein dan gen whey terhadap kualitas susu, terutama untuk kadar protein susu. Hasil menunjukan pengaruh nyata varian genetik gen s1-, -, s2-, -kasein, -laktalbumin, -laktoglobulin dan laktoferin terhadap produksi, kualitas dan komposisi susu sapi perah. Pemeriksaan dua atau lebih gen-gen protein susu (kasein dan whey protein) secara bersamaan bermanfaat untuk memperhitungkan pengaruh interaksi yang mungkin terjadi antara dua atau lebih gen haplotipe. Kegiatan penelitian ini bertujuan untuk mengetahui konsistensi genotipe gen β-kasein dan laktoferin serta interaksi kedua gen dalam mengkontrol kadar protein, kualitas susu dan SCC pada sapi FH laktasi pada dua kondisi pemeliharaan berbeda, yaitu intensif di stasiun bibit sapi perah BPPT-SP Cikole dan semi intensif di peternakan rakyat di bawah binaan KPSBU Lembang.
Tujuan
yaitu intensif di stasiun bibit sapi perah BPPT-SP Cikole dan semi intensif di peternakan rakyat di bawah binaan KPSBU Lembang.
Manfaat
Informasi yang diperoleh dari penelitian diharapkan dapat digunakan untuk mengetahui konsistensi genotipe gen β-kasein dan laktoferin dan interaksi kedua gen dalam mengontrol kadar protein dan SCC pada sapi FH domestik yang selanjutnya dapat digunakan sebagai informasi dasar pemanfaatan gen-gen tersebut sebagai marka seleksi kualitas susu.
Kerangka Pemikiran
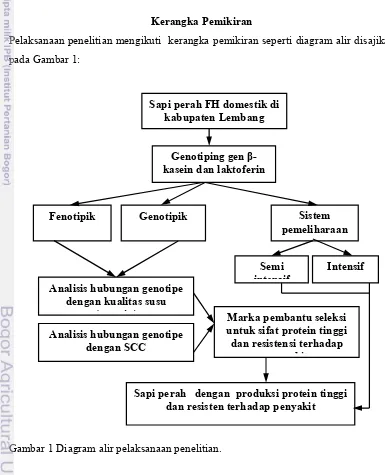
Pelaksanaan penelitian mengikuti kerangka pemikiran seperti diagram alir disajikan pada Gambar 1:
Gambar 1 Diagram alir pelaksanaan penelitian.
Fenotipik Sistem
pemeliharaan Genotipik
Analisis hubungan genotipe dengan kualitas susu
(protein)
Analisis hubungan genotipe dengan SCC
Intensif Semi
intensif
Marka pembantu seleksi untuk sifat protein tinggi dan resistensi terhadap
penyakit
Sapi perah dengan produksi protein tinggi dan resisten terhadap penyakit
Sapi perah FH domestik di kabupaten Lembang
TINJAUAN PUSTAKA
Susu
Susu adalah sekresi kelenjar ambing dari hewan mammalia tidak ditambah atau dikurangi suatu zat apa pun ke dalamnya dan diperoleh dari pemerahan ternak yang sehat (Sudono, 1985). Pada umumnya susu terdiri atas tiga komponen utama, yaitu protein, lemak dan laktosa (Schmidt et al. 1988) ditambah air, vitamin dan mineral (Sudono, 1985).
Sekresi susu terjadi selama masa laktasi yaitu selang antara masa beranak dan masa kering selama sepuluh bulan (Phillips 2002; Tyler dan Ensminger 1993). Setelah melahirkan, selama lima hari pertama sapi akan menghasilkan kolostrum. Pada awal laktasi produksi susu terus meningkat dengan cepat dan puncak produksi akan dicapai pada hari ke 30 sampai 60 (De Ross et al. 2004). Setelah puncak produksi dicapai selanjutnya produksi susu cenderung menurun sampai masa kering. Kemampuan untuk mempertahankan puncak laktasi secara terus menerus dalam waktu yang lama (persistensi) akan menyebabkan seekor sapi memiliki total produksi yang tinggi (Phillips 2002; Tyler dan Ensminger 2006). Gambar 2 memperlihatkan variasi produksi susu selama masa laktasi dengan tingkat persistensi yang berbeda.
Gambar 2. Produksi susu selama masa laktasi dengan tingkat persistensi yang berbeda (Tyler dan Ensminger 2006)
Persistensi sedang
Persistensi tinggi
Sapi perah yang selama masa laktasinya mempunyai produksi susu yang rendah, puncak produksi dicapai lebih awal dan penurunan produksinya lebih cepat daripada sapi yang selama masa laktasinya mempunyai produksi susu yang tinggi ( Mukhtar 2006). Bertujuan untuk meningkatkan produksi susu selama laktasi, seleksi dilakukan selain dengan memilih sapi-sapi yang mempunyai produksi susu tertinggi juga mempunyai persistensi yang bagus (Mukhtar 2006).
Komposisi Susu
Komposisi susu terdiri atas: protein, lemak, karbohidrat, mineral, vitamin dan air. Komponen penyusun susu masing-masing individu sangat bervariasi tergantung spesies hewan (Boland 2000, Phillips 2002, Schmidt et al. 1988). Perbedaan tersebut dapat terjadi akibat pengaruh spesies, bangsa, kondisi kesehatan, kondisi nutrisi, tingkat laktasi dan umur yang berbeda (Fox 2003; Fox dan Mc Sweeney 1998; Wong et al. 1988).
Protein. Protein dalam susu terdiri atas kasein dan whey. Kasein terdiri atas empat jenis polipeptida, yaitu αs1-, β-,αs2-dan κ-kasein (Eigel, et al. 1984; Wong et al. 1988). Whey terdiri atas β-1aktoglobulin, α-laktalbumin, serum albumin, glikomakropeptida dan protein antimikrobia yang berupa laktoferin, laktoperoksidase dan lisozim (Edwards 2009; Eigel, et al. 1984; Wong et al. 1988). Kandungan protein susu relatif tetap selama laktasi. Protein susu yang berupa kasein, β -1actoglobulin danα-laktalbumin disintesis di dalam kelenjar ambing yang dikontrol oleh gen, sedangkan sisanya (5%) di absorbsi dari darah (Fox 1989; Fox 2003; Tyler dan Ensminger 2006).
dan cadangan lemak tubuh (Palmquist 2006; Tyler dan Ensminger 1993). Sumber pembentukan lemak susu ada tiga yaitu glukosa, triasilgliserol dari bahan makanan atau yang dibentuk oleh bakteri rumen dan asam lemak yang disintesis oleh kelenjar ambing (Tyler dan Ensminger 1993).
Karbohidrat. Menurut Huppertz dan Kelly (2009), Fox (2003) dan Wong et al.(1988) karbohidrat utama dari susu adalah laktosa yang terdapat dalam bentuk alfa dan beta. Laktosa terlarut didalam susu sehingga mempengaruhi stabilitas titik beku, titik didih dan tekanan osmosis dari susu. Kadar laktosa dalam susu adalah 4.8-5.1% (Huppertz dan Kelly 2009; Phillips 2001; Tyler dan Ensminger 1993). Kadar laktosa relatif tetap, namun produksi laktosa meningkat sejalan dengan peningkatan produksi susu. Fluktuasi kadar laktosa terjadi sesuai dengan dinamika produksi susu selama laktasi (Phillips 2001).
Mineral dan vitamin. Mineral utama yang terdapat dalam susu adalah kalsium, fosfor, potasium, magnesium dan sodium. Hanya 25% kalsium, 20% magnesium dan 44% fosfor terdapat dalam bentuk yang tidak larut, sedangkan mineral-mineral lainnya semuanya dalam bentuk larut. Kalsium dan magnesium dalam bentuk yang tidak larut bersenyawa dengan kaseinat, fosfat dan sitrat. Hal inilah yang memungkinkan air susu dapat mengandung kalsium dalam konsentrasi yang besar serta pada saat yang sama dapat mempertahankan tekanan osmosis secara normal dengan darah. Kemampuan bekerja sebagai bufer dari susu disebabkan oleh adanya sitrat, fosfat, bikarbonat dan protein (Fox 2003; Walstra et al. 2006 ).
Vitamin yang terdapat dalam susu adalah vitamin A, B2 dan B12. Vitamin dalam susu diserap dari darah secara langsung sehingga peningkatan status vitamin dalam darah akan mempengaruhi konsentrasi vitamin dalam susu. Vitamin A terlarut dalam lemak sehingga kadar vitamin A dalam darah dipengaruhi oleh kadar lemak susu (Phillips 2001; Tyler dan Ensminger 1993).
Air. Air yang terkandung didalam susu bervariasi antara 32-89%, dengan kandungan rata-rata 87%. Air berguna sebagai medium dispersi untuk total solid. Naik atau turunnya bahan padat total akan mengubah persentase air (Boland 2000; Fox 2003).
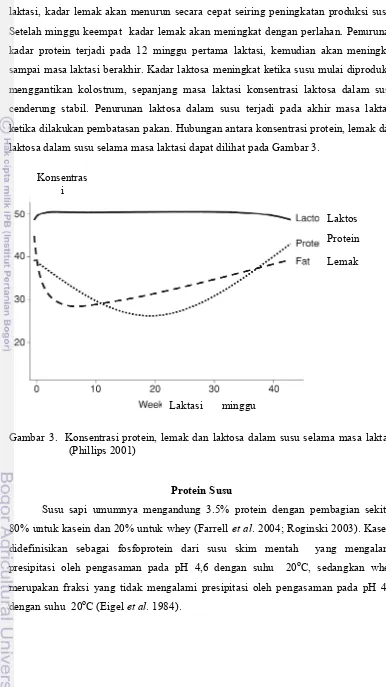
laktasi, kadar lemak akan menurun secara cepat seiring peningkatan produksi susu. Setelah minggu keempat kadar lemak akan meningkat dengan perlahan. Penurunan kadar protein terjadi pada 12 minggu pertama laktasi, kemudian akan meningkat sampai masa laktasi berakhir. Kadar laktosa meningkat ketika susu mulai diproduksi menggantikan kolostrum, sepanjang masa laktasi konsentrasi laktosa dalam susu cenderung stabil. Penurunan laktosa dalam susu terjadi pada akhir masa laktasi ketika dilakukan pembatasan pakan. Hubungan antara konsentrasi protein, lemak dan laktosa dalam susu selama masa laktasi dapat dilihat pada Gambar 3.
Gambar 3. Konsentrasi protein, lemak dan laktosa dalam susu selama masa laktasi (Phillips 2001)
Protein Susu
Susu sapi umumnya mengandung 3.5% protein dengan pembagian sekitar 80% untuk kasein dan 20% untuk whey (Farrell et al. 2004; Roginski 2003). Kasein didefinisikan sebagai fosfoprotein dari susu skim mentah yang mengalami presipitasi oleh pengasaman pada pH 4,6 dengan suhu 20oC, sedangkan whey merupakan fraksi yang tidak mengalami presipitasi oleh pengasaman pada pH 4,6 dengan suhu 20oC (Eigel et al. 1984).
Laktos Protein
Lemak
Laktasi minggu Konsentras
Kasein terdiri atas empat jenis polipeptida, yaitu αs1- kasein (39–46% dari total kasein), β-kasein (25–35% dari total kasein), αs2-kasein (8–11% dari total kasein) dan κ-kasein (8–15% dari total kasein) (Eigel et al. 1984; Roginski 2003). Whey terdiri atas β-1aktoglobulin, α-laktalbumin, serum albumin, glikomakropeptida dan protein antimikrobia yang berupa immunoglobulin, laktoferin, laktoperoksidase dan lisozim (Adlerova 2009; de Wit 1998; Eigel et al. 1984; Wong et al. 1988).
Kasein merupakan protein utama dalam susu dan termasuk dalam keluarga dari fosfoprotein yang disintesis oleh kelenjar ambing. Sintesis protein tersebut merupakan respon dari hormon laktogenik dan disekresikan dalam bentuk butiran (Farrell et al. 2004). Kasein dibedakan berdasarkan kecepatan perambatan pada elektroforesis menggunakan gel poliakrilamida atau gel starch alkali yang mengandung urea dengan atau tanpa mercaptoethanol (Eigel et al. 1984).
Whey merupakan fraksi yang terlarut pada pH 4.6. Kandungan protein utama pada whey adalah β-laktoglobulin, α-laktalbumin, serum albumin dan protein antimikrobia dengan konsentrasi secara berurutan adalah 50, 20, 10 dan 20 % dari total whey (Edwards et al. 2009). Sebagian besar protein pada whey merupakan protein globular dengan struktur sekunder dan tersier sehingga mudah terdenaturasi pada pemanasan dengan suhu sekitar 60°C (Madureira et al, 2007; Permyakova et al. 2000).
Sekitar 95% komponen protein susu disintesis dari asam amino dan 5% lainnya diserap dari darah. Komponen yang diserap dari darah yaitu serum albumin dan immunoglobulin (Edwards et al. 2009). Protein antimikrobia dalam susu berupa immunoglobulin, laktoferin, lisozim, laktoperoksidase dan glikomakropeptida yang merupakan komplek protein yang berfungsi sebagai imunitas untuk melawan berbagai macam penyakit (Adlerova 2009; Connely 2001). Konsentrasi protein antimikrobia dalam kolostrum sangat tinggi yaitu mencapai 100 g/l dan secara cepat turun menjadi 1 g/l selama kurang dari satu minggu setelah kelahiran (Roginski 2003; Walstra dan Jenness 1984).
Gen β-Kasein
Menurut Jann et al. (2004), Mercier dan Vilolte (1993), Rijnkels (2002) dan Rijnkels et al. (1997) β-kasein bersama keluarga gen kasein lainnya bertempat pada lokus yang sama yaitu di kromosom 6 (6q31). Kasein terdiri atas empat lokus saling berdekatan (haplotipe), dengan runutan genom s1-, -, s2-, dan -kasein dengan panjang 250 pb. Jarak antara -kasein dengan s1- kasein dan s2-kasein secara berurutan adalah 19.6 kb dan 70kb. Struktur lokus kasein pada sapi disajikan pada Gambar 4.
Gambar 4 Struktur lokus kasein pada sapi (Rijnkels 2002).
β-kasein merupakan salah satu jenis dari fraksi kasein yang berpengaruh
1984; Yahyaoui et al. 2003). β-kasein bersifat paling hidrofobik dan mengandung residu prolin paling banyak jika dibandingkan dengan keluarga kasein yang lainnya. Pada suhu rendah sebagian fraksi β-kasein akan terlarut sehingga meningkatkan viskositas susu (Eigel et al. 1984; Farrell et al. 2004; Roginski 2003).
Menurut Rijnkels et al. (1997) β-kasein mengandung 209 asam amino dengan berat molekul 24.000 Da. Gen β-kasein memiliki panjang 8498 pb dan terdiri atas 9 exon dan 8 intron. Struktur gen β-kasein disajikan pada Gambar 5. Perbandingan panjang sekuens exon dengan intron β-kasein adalah 1: 6.8. Panjang total exon adalah 1090 pb dan panjang total intron adalah 7408 pb.
Gambar 5 Struktur gen β-kasein (disusun berdasarkan sekuens gen β-kasein nomor akses gen bankX14711).
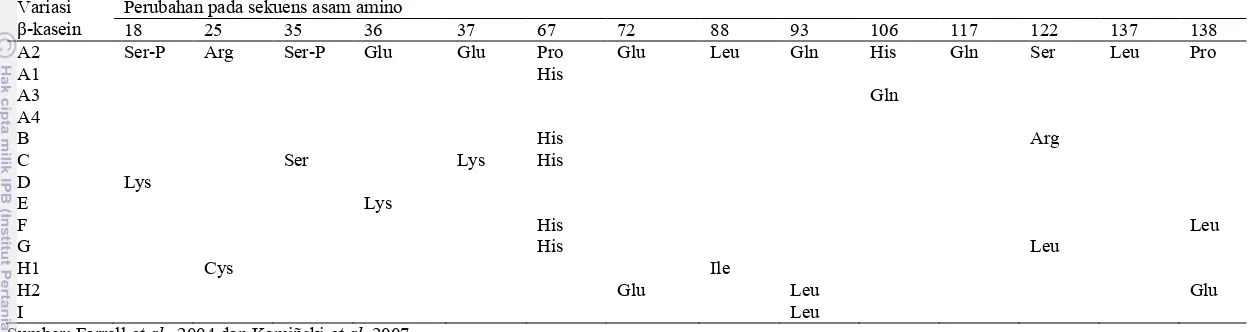
KeragamanGen β-Kasein
Keragamangen β-kasein disebabkan oleh perbedaan susunan sekuens asam amino penyusun protein. Perbedaan susunan sekuens β-kasein terdapat pada asam amino ke-18, 25, 35, 36, 37, 67, 72, 88, 93, 106, 117, 122, 137 dan 138. Berdasarkan perbedaan sekuens asam aminonya terdapat 13 variasi alel β-kasein. Seluruh alel β-kasein telah diketahui sekuens asam aminonya kecuali untuk alel A4 (Farrell et al.2004 dan Kamiñski et al.2007). Perbedaan sekuens asam amino pada variasi β-kasein disajikan pada Tabel 2.
E1 E2 E3E4 E5E6 E7 E8 E9
Keterangan: untuk exon dan untuk intron
Panjang Panjang
Exon 1 44 pb Intron 1 1934 pb
Exon 2 63 pb Intron 2 724 pb
Exon 3 27 pb Intron 3 112 pb
Exon 4 27 pb Intron 4 1895 pb
Exon 5 24 pb Intron 5 92 pb
Exon 6 42 pb Intron 6 1320 pb
Exon 7 498 pb Intron 7 601 pb
Exon 8 42 pb Intron 8 730 pb
Exon 9 323 pb
Tabel 2 Perubahan sekuensasam amino pada variasi β–kasein.
Variasi
β-kasein
Perubahan pada sekuens asam amino
18 25 35 36 37 67 72 88 93 106 117 122 137 138
A2 Ser-P Arg Ser-P Glu Glu Pro Glu Leu Gln His Gln Ser Leu Pro
A1 His
A3 Gln
A4
B His Arg
C Ser Lys His
D Lys
E Lys
F His Leu
G His Leu
H1 Cys Ile
H2 Glu Leu Glu
I Leu
57
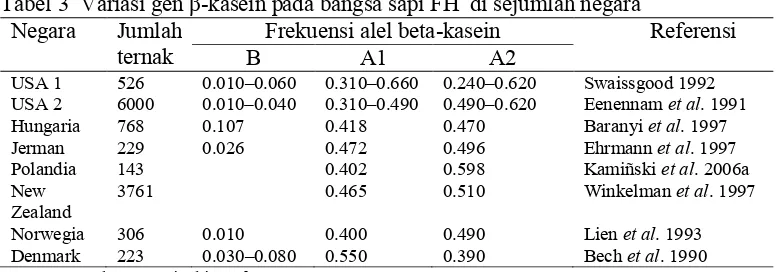
Tabel 3 Variasi gen β-kasein pada bangsa sapi FH di sejumlah negara Negara Jumlah
ternak
Frekuensi alel beta-kasein Referensi
B A1 A2
USA 1 526 0.010–0.060 0.310–0.660 0.240–0.620 Swaissgood 1992 USA 2 6000 0.010–0.040 0.310–0.490 0.490–0.620 Eenennam et al. 1991 Hungaria 768 0.107 0.418 0.470 Baranyi et al. 1997 Jerman 229 0.026 0.472 0.496 Ehrmann et al. 1997 Polandia 143 0.402 0.598 Kamiñski et al. 2006a New
Zealand
3761 0.465 0.510 Winkelman et al. 1997
Norwegia 306 0.010 0.400 0.490 Lien et al. 1993 Denmark 223 0.030–0.080 0.550 0.390 Bech et al. 1990
Sumber: Kamiñski et al. 2007.
58
bahwa alel A2 berhubungan dengan kadar protein serta produksi susu yang tinggi dan kadar lemak yang rendah. Hal ini berbeda dengan hasil penelitian Kučerováet al. (2006) yang menyatakan bahwa genotipe A1A1 menghasilkan produksi susu paling tinggi dan genotipe A2A3 menghasilkan kadar protein dan kadar lemak yang tertinggi.
Gen Laktoferin
Laktoferin merupakan glikoprotein pengikat zat besi rantai tunggal anggota kelompok gen transferin. Laktoferin terdapat dalam susu dan juga sekresi kelenjar eksokrin lainnya seperti semen, air liur dan cairan servik. Laktoferin disintesis oleh sel granulosit dan kelenjar epitel ambing sebagai respon terhadap infeksi seperti mastitis (Arnould et al 2009; Brock 2002; Gifford 2005). Konsentrasi laktoferin dalam kolostrum sangat tinggi yaitu mencapai 100 g/l dan secara cepat turun menjadi 1 g/l selama kurang dari satu minggu setelah kelahiran. Konsentrasi laktoferin pada susu sapi berkisar antara 0.02 sampai 0.35 g/l, tergantung pada periode laktasi (Roginski 2003, Walstra dan Jenness 1984), total sel somatik, konsentrasi lisosim dalam susu, musim dan umur ternak (Cheng et al. 2008; Hagiwara et al. 2003).
Connely (2001) Baker dan Baker (2005) dan Molenaar et al. (1996) menyatakan bahwa laktoferin merupakan protein multi fungsi yang berperan membantu penyerapan zat besi di usus, pertumbuhan sel usus, melindungi dari serangan mikroba penyebab infeksi dan sebagai sistem kekebalan tubuh. Laktoferin mengatur proses inflamatori, respon sistem immun dan pertumbuhan sel (Baker dan Baker 2005; Schanbacheret al. 1993, Connely 2001).
59
dalam pemusnahan fagosit dan imunitas (Kutila et al. 2003; Molenaar et al. 1996; Pan 2007).
Laktoferin memiliki berat 82.600 Da dan secara spesifik dapat mengikat dua atom feri per molekulnya (Baker dan Baker 2005). Sekuens laktoferin yang bersifat antimikroba pada manusia terdiri dari 18 residu asam amino yang dibentuk dari ikatan disulfida antara residu sistein 20 dan 37 (Dionysius dan Milne 1997), sedangkan pada sapi 19 dan 36 (Schanbacher et al. 1993; Teng 2002). Sekuens lengkap mRNA laktoferin telah dilakukan oleh Schanbacher et al. (1993) dan didapatkan homologi yang tinggi antara laktoferin dengan transferin. Namun terdapat perbedaan wilayah deduksi sekuens asam amino antara laktoferin sapi dibandingkan dengan laktoferin manusia dan transferin, hal ini menunjukkan adanya fungsi yang berbeda di antara ketiganya.
Keterangan: untuk exon dan untuk intron
Panjang Panjang
Exon 1 81 pb Intron 1 32 pb
Exon 2 163 pb Intron 2 49 pb
Exon 3 108 pb Intron 3 292 pb
Exon 4 182 pb Intron 4 478 pb
Exon 5 147 pb Intron 5 941 pb
Exon 6 55 pb Intron 6 1486 pb
Exon 7 178 pb Intron 7 447 pb
Exon 8 174 pb Intron 8 526 pb
Exon 9 154 pb Intron 9 527 pb
Exon 10 90 pb Intron 10 1092 pb
Exon 11 47 pb Intron 11 630 pb
Exon 12 155 pb Intron 12 12 pb
Exon 13 141 pb Intron 13 918 pb
Exon 14 68 pb Intron 14 495 pb
Exon 15 183 pb Intron 15 79 pb
Exon 16 189 pb Intron 16 60 pb
Exon 17 224 pb
Panjang exon 2339 pb Panjang intron 8064 pb
Gambar 6 Struktur gen laktoferin (disusun berdasarkan sekuens gen laktoferin nomor akses gen bankAB052164).
60
Gen laktoferin terletak pada kromosom 22 (22q24). Struktur gen laktoferin terdiri atas 17 exon dan 16 intron (Seyfert et al. 1994; Seyfert et al. 1997). Total basa pada gen laktoferin sapi adalah 13647 pb. Panjang total exon, total intron dan daerah promotor gen laktoferin secara berurutan adalah 2339 pb, 8064 pb dan 1122 bp (Schanbacher et al. 1993; Seyfert et al. 1994; Seyfert et al. 1997). Struktur gen laktoferin disajikan pada Gambar 6.
Keragaman Gen Laktoferrin
Struktur gen laktoferin diidentifikasi oleh Seyfert et al. (1994) yaitu mempunyai 17 exon dan 16 intron dengan panjang total 13647 pasangan basa (pb). Lee et al. (1997) melaporkan adanya polimorfisme pada gen laktoferin pada kambing asli Korea yang disebabkan oleh adanya mutasi di 15 titik cDNA. Mutasi tersebut mengakibatkan adanya variasi sebanyak lima asam amino di bagian N-lobe dan satu asam amino dibagian C-lobe. Mutasi ini berhubungan langsung dengan kadar laktoferin dan kemampuannya sebagai antibakteri.
Menurut Arnould et al. (2009) terdapat polimorfisme pada exon 2, exon, 4, exon 8, exon 9, exon 11, exon 15, intron 6, intron 8 dan daerah promoter yang berkaitan secara nyata dengan produksi susu, kadar laktoferin, kadar protein, kadar lemak dan skor sel somatik. Arnould et al. (2009) melaporkan adanya korelasi negatif antara jumlah sel somatik dengan konsentrasi laktoferin dalam susu. Menurut Arnouldet al. 2009, Pawlik et al.(2009), Renaville (2009) serta Rupp dan Boichard (2003) kadar laktoferin sangat nyata dipengaruhi oleh genetik.
Menurut Sender (2006), Wojdak-Maksymiec (2006) dan Zhao (2008) gen laktoferin memiliki tiga varian genotipe yaitu genotipe AA, AB dan BB. Menurut Sender (2006) frekuensi genotipe laktoferin varian AA paling tinggi (0.644) diikuti oleh AB (0.296) dan BB (0.04). Hal serupa dinyatakan oleh Zhao yang melaporkan bahwa pada ternak yang sehat frekuensi genotipe AA yang paling tinggi. Sedangkan Wojdak-Maksymiec (2006) melaporkan bahwa ternak dengan genotipe AB memiliki frekuensi tertinggi (0.597) diikuti oleh genotipe AA (0.379) dan BB (0.024).
61
62
MATERI DAN METODE
Waktu dan Tempat
Penelitian ini dilaksanakan selama sembilan bulan, yaitu dari bulan Oktober 2009 sampai dengan Juni 2010. Penelitian ini dilaksanakan di Laboratorium Genetika Molekuler, Bagian Pemuliaan dan Genetika Ternak, Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, Institut Pertanian Bogor.
Materi
Sapi Perah FH
Sapi perah yang digunakan untuk penelitian adalah sapi FH laktasi dengan kisaran hari laktasi 1 sampai 6 bulan dan periode laktasi 1 sampai 5 pada daerah sentra produksi susu di Kabupaten Lembang Jawa Barat yang telah diketahui data produksi dan kualitas susunya. Studi dilakukan pada dua kondisi pemeliharaan berbeda yaitu pada pemeliharaan secara intensif dan semi intensif.
Pada pemeliharaan ternak perah secara intensif usaha dilakukan dengan organisasi operasional yang modern dan skala usaha yang besar, dijalankan secara teratur di suatu tempat dalam jangka waktu tertentu dan bertujuan komersial yaitu menghasilkan susu dan hasil sampingan yang lainnya. Pemeliharaan dengan manajemen intensif diwakili oleh Unit Pelaksana Teknis Daerah (UPTD) Balai Pembibitan dan Pengembangan Sapi Perah (PBPT-SP) Cikole. Penerapan aspek teknis pemeliharaan sapi perah yang meliputi breeding dan reproduksi, makanan ternak, pengelolaan, kandang dan peralatan serta kesehatan ternak pada PBPT-SP Cikole secara umum telah terkontrol dan dilakukan sesuai Good Farming Practice. Gambaran pelaksanaan pemeliharaan ternak pada PBPT-SP Cikole dapat dilihat pada Lampiran 13.
63
peternakan rakyat di bawah binaan KPSBU Lembang secara umum baru mencapai 80%. Penerapan aspek teknis dari yang tertinggi hingga terendah berturut-turut adalah aspek kesehatan hewan (85.5%), diikuti aspek makanan ternak (84.38%), aspek pengelolaan (78.16%), aspek kandang dan peralatan (76.13%) serta aspek breedingdan reproduksi (72.35%). Gambaran pelaksanaan pemeliharaan ternak pada peternakan rakyat di bawah binaan KPSBU Lembang dapat dilihat pada Lampiran 13.
Sampel Darah
Sampel darah yang digunakan dalam penelitian ini adalah darah yang berasal dari sapi FH laktasi yang telah diketahui data produksi dan kualitas susunya. Sampel yang digunakan sebanyak 282 ekor dengan perincian dari PBPT Cikole sebanyak 88 ekor, KPSBU Desa Pasir Kemis sebanyak 96 ekor dan KPSBU Desa Cilumber sebanyak 98 ekor. Sampel darah diambil dari bagian vena juguralis dengan menggunakan jarum vaccutainer no. 21 G dan disimpan dalam tabung vaccutainer 10 ml dengan alkohol absolut. Perbandingan antara volume alkohol dengan darah adalah 1:1.
Metode
Isolasi DNA
DNA yang digunakan untuk identifikasi polimorfisme gen β-kasein dan laktoferin merupakan koleksi dari Laboratorium Pemuliaan dan Genetika Ternak, Departemen Ilmu Produksi dan Teknologi Peternakan, Institut Pertanian Bogor. Isolasi DNA dilakukan dari darah dan sel somatik susu. Isolasi DNA dari sampel darah menggunakan metode ekstraksi phenol-chloroform (Sambrook et al. 1989) yang telah dimodifikasi untuk sampel darah yang disimpan dalam alkohol. Isolasi DNA dari sampel susu menggunakan metode ekstraksi DNA dari sel somatik (Arnouldet al. 2009).
64
Reaksi PCR dilakukan pada mesin thermocycler dengan enzim taq polymerase dan buffernya. Informasi mengenai primer yang digunakan untuk gen laktoferin dan β-kasein disajikan pada Tabel 4.
Pereaksi yang digunakan untuk amplifikasi kedua ruas gen target adalah 2 µl sampel DNA cetakan, primer 1 pmol, dNTPs 200 µM, MgCl21 mM, dan 0.5 unit taq polymerase(Real Taq) serta bufernya dalam larutan total 25 µl. Amplifikasi in vitro dengan mesin thermal cyclerdilakukan dengan kondisi denaturasi awal pada suhu 94 ºC selama 5 menit, 35 siklus yang terdiri atas denaturasi pada suhu 94 ºC selama 45 detik, annealing pada suhu 59 ºC selama 1 menit untuk gen β-kasein dan suhu 60 ºC selama 45 detik untuk laktoferin. Elongasi pada suhu 72 ºC selama 1 menit dan elongasi akhir pada suhu 72 ºC selama 5 menit.
Tabel 4 Informasi sekuen primer yang digunakan dalam penelitian.
Gen
CSN2 233 Exon 7 F:CCAGACACAGTCTCTAGTCTATCCC Medrano dan Sharrow 1991 R:CAACATCAGTGAGAGTCAGGCTCCG
Penentuan Genotipe (Genotyping)
Penentuan genotipe untuk gen β-kasein dan gen laktoferin menggunakan metode restriction fragment length polymorphism (RFLP). Hasil genotyping divisualisasikan pada gel agarose 2% dengan bufer 0,5x TBE (tris borat EDTA) yang dijalankan pada tegangan 100 volt selama 40 menit yang diwarnai dengan etidium bromida di atas UV trans ilmuninator. Enzim restriksi yang digunakan untuk gen
target β-kasein adalah MspI dengan situs pemotong C↓CGG dan untuk gen target laktoferin adalah EcoRI dengan situs pemotong G↓AATTC.
Pengukuran Produksi Susu
65
Analisis Kualitas Susu
Data kualitas susu yang terdiri atas kadar protein, berat jenis, kadar lemak, kadar bahan kering dan kadar bahan kering tanpa lemak yang merupakan data sekunder (data telah diambil pada bulan Agustus 2008). Prosedur pengujian kualitas susu yang dilakukan sesuai standart uji pada Dewan Standarisasi Nasional (1998) yaitu:
Analisis kadar protein. Prosedur pengujian kadar protein menggunakan cara titrasi formol, p yaitu banyaknya NaOH yang terpakai untuk titrasi sampel (susu) dan q yaitu banyaknya NaOH yang terpakai untuk titrasi blanko. Kadar protein dihitung dengan rumus berikut:
% protein = (p – q) ml x 1.7 (faktor formol)
Analisis kadar lemak. Pengukuran kadar lemak susu menggunakan metode Gerber.
Analisis berat jenis. Pengukuran berat jenis dilakukan dengan alat laktodensimeter, kemudian dilakukan penyetaraan pada suhu 27.5°C. Penyebab utama bervariasinya berat jenis ini adalah kandungan lemak susu. Berat jenis dari skim, krim dan susu segar dapat dihitung dengan rumus berikut:
Keterangan: ρ = berat jenis mF = kadar lemak
Bahan kering dan bahan kering tanpa lemak. Dihitung setelah kadar lemak dan berat jenis diperoleh dengan rumus:
Bahan kering tanpa lemak = Bahan kering – Kadar lemak
Penghitungan Jumlah Sel Somatik Susu
66
membentuk suatu lapisan tipis dan merata. Sampel dibiarkan kering udara selama 5 sampai 10 menit selanjutnya difiksasi dengan api Bunsen. Pewarnaan Breed dilakukan dengan merendam gelas objek dalam eter alkohol selama 2 menit. Sampel diwarnai dengan larutan Methylen Loeffler biru selama 1-2 menit, kemudian dimasukkan dalam larutan alkohol 96 % untuk menghilangkan sisa zat warna yang tidak melekat. Jumlah sel somatis dihitung dengan bantuan mikroskop pada pembesaran 100 kali.
Rancangan dan Analisis Data
Frekuensi Alel dan Genotipe
Frekuensi alel dari masing-masing lokus dapat diperkirakan dengan penghitungan jumlah gen pada populasi. Menurut Nei (1987), jika niiadalah individu yang bergenotipe AiAi, nij adalah individu yang bergenotipe AiAj, dan jumlah total sampel adalah n, maka frekuensi alel Ai (χi) dihitung dengan persamaan sebagai berikut:
nii nij
ni 2 /2
Frekuensi genotipe dapat diperkirakan dengan menghitung perbandingan jumlah genotipe pada populasi. Menggunakan asumsi sebelumnya, maka frekuensi genotipe AiAi(χii) dapat dihitung dengan persamaan sebagai berikut:
n nii ii
Keterangan : Xi = frekuensi alel ke-i
nij = jumlah individu yang bergenotipe ii nii = jumlah individu yang bergenotipe ij n = jumlah sampel
Derajat Heterozigositas
Derajat heterozigositas (ĥ)dihitung berdasarkan frekuensi alel pada tiap lokus DNA dengan rumus Nei (1987) :
ĥ = 2n (1 -ΣXi2) / (2n – 1) Keterangan : Xi = frekuensi alel
67
Proporsi Hardy-Weinberg
Proporsi Hardy-Weinberg dihitung berdasarkan rumus sebagai berikut:
O = jumlah pengamatan genotipe ke-i E = jumlah harapan genotipe ke-i
Penentuan keragaman genetik dilakukan dengan analisis frekuensi alel, frekuensi genotipe dan nilai heterozigositas. Penentuan keseimbangan genotipe dilakukan dengan menghitung Proporsi Hardy-Weinberg. Analisis keragaman dan keseimbangan genetik gen β-kasein dan laktoferin diuji dengan software Tools for Population Genetics Analyses (TFPGA) (Miller 1997).
Pengaruh Manajemen Pemeliharaan terhadap Produksi dan Kualitas Susu
Hubungan antara manajemen pemeliharaan dengan produksi dan kualitas susu sapi perah FH dianalisis dengan pendekatan General Linear Model. Data produksi dan kualitas susu dikelompokkan berdasarkan manajemen pemeliharaan, periode dan bulan laktasi yang berbeda. Pengujian dilakukan berdasarkan perbedaan
kelompok dan jika respon menunjukkan perbedaan (α=0,05) dilakukan pengujian
lanjut dengan metode uji beda Tukey. Model matematis dirumuskan sebagai berikut (Mattjik dan Sumertawijaya (2006):
Yij= μ + αi+ eij
Keterangan : Yij = nilai pengamatan
μ = komponen aditif dari rataan
αi = pengaruh aditif perlakuan eij = pengaruh galat perlakuan
Pengaruh Genotipe β-Kasein dan Laktoferin terhadap Protein dan Kualitas Susu
Data produksi dan kualitas susu sebelum dianalisis distandarisasi terhadap hari laktasi (1 sampai ≥ 6 bulan) dan periode laktasi (1sampai ≥ 5). Standarisasi menggunakan rumus :
Peubah terstandarisasi
68
x = peubah yang akan distandarisai
Analisis hubungan antara genotipe β-kasein dan laktoferin dengan produksi susu, kadar protein dan kualitas susu (pH, kadar lemak, bahan kering dan berat kering tanpa lemak) dilakukan dengan metode analisis General Linear Model(GLM) dengan menggunakan software SAS Ver 9,1. Pengaruh keragaman genetik serta korelasi antara dua gen terhadap produksi susu dan kualitasnya dihitung berdasarkan model linear dari Mattjik dan Sumertawijaya (2006) dan jika respon menunjukkan
perbedaan (α=0,05) dilakukan pengujian lanjut dengan metode uji Tukey. Model matematis dirumuskan sebagai berikut:
Yijk= μ + αi+βj+ (αβ)ij+ eijk
Keterangan : Yij = nilai pengamatan
μ = komponen aditif dari rataan
αi = pengaruh aditif gen β-kasein
βj = pengaruh aditif gen laktoferin
69
HASIL DAN PEMBAHASAN
Produksi Susu
Pemeriksaan produksi susu dari sapi FH dilakukan untuk setiap kondisi pemeliharaan yang berbeda. Pemeriksaan terpisah antara lokasi BPPT-SP Cikole dengan KPSBU Lembang bertujuan untuk meniadakan pengaruh perbedaan manajemen pemeliharaan. Data produksi susu merupakan data dari hasil pemerahan susu selama satu hari pada kisaran bulan laktasi pertama sampai bulan laktasi keenam pada periode laktasi kesatu sampai kelima.
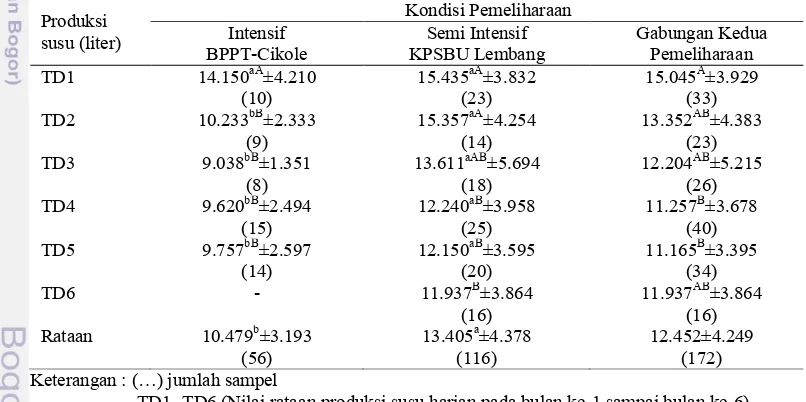
Jumlah catatan harian atau test day untuk seluruh laktasi adalah 272 data. Karakteristik data catatan harian produksi susu setiap kondisi pemeliharaan yang dikelompokkan berdasarkan bulan laktasi disajikan pada Tabel 5.
Tabel 5. Nilai rataan produksi susu harian yang dikelompokkan berdasarkan bulan laktasi pada kondisi pemeliharaan yang berbeda.
Produksi TD1 14.150aA±4.210 15.435aA±3.832 15.045A±3.929
(10) (23) (33)
TD2 10.233bB±2.333 15.357aA±4.254 13.352AB±4.383
(9) (14) (23)
TD3 9.038bB±1.351 13.611aAB±5.694 12.204AB±5.215
(8) (18) (26)
TD4 9.620bB±2.494 12.240aB±3.958 11.257B±3.678
(15) (25) (40)
TD5 9.757bB±2.597 12.150aB±3.595 11.165B±3.395
(14) (20) (34)
TD6 - 11.937B±3.864 11.937AB±3.864
(16) (16)
Rataan 10.479b±3.193 13.405a±4.378 12.452±4.249
(56) (116) (172)
Keterangan : (…) jumlah sampel
TD1- TD6 (Nilai rataan produksi susu harian pada bulan ke-1 sampai bulan ke-6). Superskip huruf kecil yang berbeda pada baris yang sama menyatakan berbeda pada tingkat kepercayaan 99 % atau 95%.
Superskip huruf kapital yang berbeda pada kolom yang sama menyatakan berbeda pada tingkat kepercayaan 99 % atau 95%.
70
bulan maupun nilai rataan keseluruhan. Nilai rataan produksi susu harian pada peternakan dengan kondisi pemeliharaan semi intensif lebih tinggi karena peternak memberikan pakan dengan intensitas yang lebih tinggi jika dibandingkan dengan pada peternakan dengan pemeliharaan intensif. Pada peternakan rakyat pemberian pakan hijauan dilakukankan tiga kali sehari atau lebih dengan pemberian secara ad libitum. Menurut Jenkins dan McGuired (2006) serta Tyler dan Ensminger (2006) frekuensi pemberian pakan yang lebih dari dua kali akan dapat meningkatkan konsumsi bahan kering pakan, kadar lemak susu dan produksi susu. Pada peternakan rakyat, peternak memberikan pakan hijauan dalam jumlah cukup sampai berlebih dan diberikan tiga kali sehari, sedangkan pada peternakan dengan pemeliharaan secara intensif pemberian pakan hijauan hanya dilakukan dua kali sehari dengan proporsi 31.15% dari total bahan kering pakan. Proporsi bahan kering hijauan sesuai kebutuhan untuk sapi perah laktasi minimal adalah 40% dari total bahan kering pakan (Jenkins dan McGuired 2006; Phillips 2001; SDCN 2001). Kecukupan pakan sapi perah secara kuantitas dan kualitas menentukan optimalitas produksi susu (Mukhtar 2006; Tyler dan Ensminger 2006; Phillips 2001).
Produksi susu tertinggi dicapai pada bulan laktasi pertama baik pada kondisi pemeliharaan intensif, pemeliharaan semi intensif maupun gabungan kedua kondisi pemeliharaan. Secara umum terlihat bahwa puncak laktasi terjadi pada bulan pertama, lalu pada bulan berikutnya akan mengalami penurunan sampai pada akhir masa pengukuran. Hasil penelitian tersebut menunjukkan bahwa pola produksi susu mencapai puncak pada bulan pertama, hal ini sejalan dengan pernyataan De Ross et al. (2004) bahwa setelah beranak produksi susu mulai naik dan mencapai puncak produksi pada minggu ke 3 sampai ke 6, setelah itu mengalami penurunan secara perlahan sampai pada masa akhir laktasi. Hamidah (1987) dan Indrijani (2001) yang yang melakukan penelitian pada sapi perah di wilayah Kabupaten Bandung melaporkan hasil yang berbeda yaitu produksi susu sapi perah mencapai puncak produksi pada 45 sampai 60 hari setelah sapi beranak. Menurut Phillips (2001) penurunan produksi susu seiring dengan lama laktasi terjadi karena adanya penurunan fungsi kelenjar ambing.
71
berbeda nyata (P<0.01). Nilai rataan produksi susu setiap periode laktasi menunjukkan adanya peningkatan dari laktasi kesatu sampai laktasi ketiga baik pada kondisi pemeliharaan intensif maupun kondisi pemeliharaan semi intensif. Keadaan tersebut sesuai dengan pendapat Caccamo et al. (2008), De Ross et al. (2004) dan Phillips (2001) yang melaporkan bahwa kapasitas produksi susu sapi perah akan meningkat terus menerus dan mencapai puncak produksi pada periode laktasi ketiga atau keempat kemudian turun berlahan-lahan. Peningkatan produksi susu pada empat periode laktasi pertama disebabkan adanya pertumbuhan kelenjar susu seiring proses kedewasaan dan pertumbuhan tubuh, sehingga akan menyebabkan peningkatan jumlah hormon prolaktin yang disekresikan.
Tabel 6. Nilai rataan produksi susu harian yang dikelompokkan berdasarkan periode laktasi pada kondisi pemeliharaan yang berbeda.
Produksi Laktasi ke-1 9.465bB±2.114 13.954aA±3.409 10.331C±2.977
(46) (11) (57)
Laktasi ke-2 - 11.708A±2.746 11.708BC±2.746
(24) (24)
Laktasi ke-3 15.786aA±3.684 14.954aA±4.258 15.100A±4.131
(7) (33) (40)
Laktasi ke-4 13.633aA±1.415 12.886aA±5.013 12.945ABC±4.821
(3) (35) (38)
Laktasi ke-5 - 13.538A±5.149 13. 538AB±5.149
(13) (13)
Rataan 10.478b±3.193 13.405a±4.378 12.452±4.249
(56) (116) (172)
Keterangan : (…) jumlah sampel
Superskip huruf kecil yang berbeda pada baris yang sama menyatakan berbeda pada tingkat kepercayaan 99 %.
Superskip huruf kapital yang berbeda pada kolom yang sama menyatakan berbeda pada tingkat kepercayaan 99 %.
72
persistensi produksi susu berpengaruh terhadap total produksi susu (Lin dan Togashi 2005). Sapi perah dengan persistensi yang tinggi maka akan menghasilkan produksi susu yang lebih tinggi jika dibandingkan dengan sapi perah yang memiliki persistensi yang rendah (Togashi dan Lin 2003; Togashi dan Lin 2004).
Secara keseluruhan produksi susu pada setiap periode menunjukkan hasil yang berbeda nyata (P<0.01), menunjukkan bahwa periode laktasi memiliki pengaruh yang sangat besar terhadap produksi susu harian pada setiap periode laktasi. Hal ini sejalan dengan hasil penelitian Hamidah (1987) dan Indrijani (2001) yang menyatakan bahwa periode laktasi memberikan pengaruh yang besar pada produksi susu. Diduga terjadinya fluktuasi produksi susu disebabkan perbedaan tata laksana pemeliharaan dan perubahan mutu genetik ternak sebagai akibat dari adanya proses seleksi pada sapi perah betina dan juga karena pengaruh perbedaan periode laktasi yang dapat menyumbangkan keragaman dalam produksi susu setiap tahunnya.
Kualitas Susu
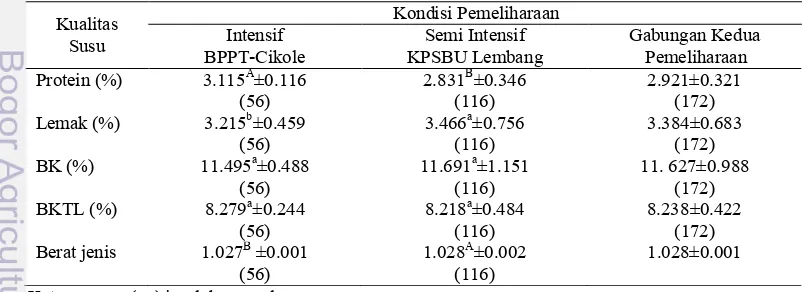
Pemeriksaan parameter kualitas susu dari sapi FH dilakukan untuk setiap lokasi. Pemeriksaan terpisah antara lokasi BPPT-SP Cikole dengan KPSBU Lembang bertujuan untuk meniadakan pengaruh perbedaan manajemen pemeliharaan (Tabel 7). Data kualitas susu diperoleh dari pengujian kualitas hasil pemerahan selama satu hari pada kisaran bulan laktasi pertama sampai bulan laktasi keenam pada periode laktasi kesatu sampai kelima.
Tabel 7. Nilai rataan kualitas susu harian sapi FH pada kondisi pemeliharaan yang Protein (%) 3.115A±0.116 2.831B±0.346 2.921±0.321
(56) (116) (172)
Lemak (%) 3.215b±0.459 3.466a±0.756 3.384±0.683
(56) (116) (172)
BK (%) 11.495a±0.488 11.691a±1.151 11. 627±0.988
(56) (116) (172)
BKTL (%) 8.279a±0.244 8.218a±0.484 8.238±0.422
(56) (116) (172)
Berat jenis 1.027B±0.001 1.028A±0.002 1.028±0.001
(56) (116)
Keterangan: (…) jumlah sampel
73
Superskip huruf kecil yang berbeda pada baris yang sama menyatakan berbeda pada tingkat kepercayaan 95 %.
Kualitas susu pada kondisi pemeliharaan yang berbeda (Tabel 7) menunjukkan bahwa nilai rataan kadar protein, lemak dan BKTL pada kedua kondisi pemeliharaan berada pada skala normal berdasarkan syarat mutu susu segar menurut Dewan Standarisasi Nasional (1998) yaitu kadar protein 2.7%, lemak 3% dan BKTL 8%, sedangkan rataan BJ susu baik pada kondisi pemeliharaan intensif maupun semi intensif di bawah standar Dewan Standarisasi Nasional (1998) yaitu 1.028. Hal ini menunjukkan bahwa secara umum susu yang dihasilkan dari peternakan baik pada kondisi pemeliharaan intensif maupun pada kondisi pemeliharaan semi intensif memiliki kualitas yang baik.
Pengelompokkan parameter kualitas susu berdasarkan kondisi pemeliharaan memberikan hasil yang berbeda nyata pada protein dan berat jenis, berbeda pada lemak, sedangkan pada dan BKTL memberikan hasil yang sama. Pada kondisi pemeliharaan intensif, kadar protein lebih tinggi 2.8% jika dibandingkan dengan kondisi pemeliharaan semi intensif dan sebaliknya dengan kadar lemak. Kadar lemak pada pemeliharaan semi intensif lebih tinggi 2.5% jika dibandingkan dengan kondisi pemeliharaan intensif. Kondisi tersebut disebabkan karena rendahnya proporsi hijauan pada total berat kering pakan pada kondisi pemeliharaan intensif. Proporsi hijauan dalam pakan yang rendah akan menyebabkan penurunan kadar lemak dan produksi susu, sebaliknya akan menyebabkan peningkatan protein susu (Jenkins dan McGuired 2006; Phillips 2001; SDCN 2001).
Kadar Protein Susu
74
tinggi 8.8% jika dibandingkan dengan yang disarankan SDCN (2001). Jenkins dan McGuired (2006) serta Phillips (2001) menyatakan bahwa pengurangaan proporsi hijauan pada komposisi pakan sapi perah akan menyebabkan peningkatan kadar protein susu. Perbedaan kadar protein susu pada kedua kondisi pemeliharaan diduga juga disebabkan perbedaan genetik ternak pada kedua kondisi pemeliharaan.
Tabel 8. Nilai rataan protein susu harian yang dikelompokkan berdasarkan bulan laktasi pada kondisi pemeliharaan yang berbeda.
Protein TD1 3.075aA±0.128 2.727bA±0.281 2.832B±0.292
(10) (23) (33)
TD2 3.135aA±0.151 2.753bA±0.177 2.903AB±0.251
(9) (14) (23)
TD3 3.103aA±0.055 2.766bA±0.404 2.870B±0.370
(8) (18) (26)
TD4 3.126aA±0.098 2.839bA±0.356 2.946AB±0.318
(15) (25) (40)
TD5 3.129aA±0.127 3.0154aA±0.425 3.062A±0.337
(14) (20) (34)
TD6 - 2.882A±0.287 2.882AB±0.287
(16) (16)
Rataan 3.115a±0.116 2.831b±0.346 2.921±0.321
(56) (116) (172)
Keterangan : TD1- TD6 (Nilai rataan protein susu harian pada bulan ke-1 sampai bulan ke-6)
Superskip huruf kecil yang berbeda pada baris yang sama menyatakan berbeda pada tingkat kepercayaan 99 %.
Superskip huruf kapital yang berbeda pada kolom yang sama menyatakan berbeda pada tingkat kepercayaan 99 %.
Tabel 9. Nilai rataan protein susu harian yang dikelompokkan berdasarkan periode laktasi pada kondisi pemeliharaan yang berbeda.
Protein (%) Laktasi ke-1 3.143aA±0.086 2.701bA±0.279 3.058A±0.226
(46) (11) (57)
Laktasi ke-2 - 2.813A±0.316 2.813B±0.316
(24) (24)
Laktasi ke-3 3.024aAB±0.155 2.937aA±0.424 2.952AB±0.390
(7) (33) (40)
Laktasi ke-4 2.905aB±0.090 2.772aA±0.256 2.783B±0.249
(3) (35) (38)
Laktasi ke-5 - 2.867A±0.409 2.867AB±0.409
(13) (13)
Rataan 3.115a±0.116 2.831b±0.346 2.921±0.321
(56) (116) (172)
Keterangan : (…) jumlah sampel
75
Superskip huruf kapital yang berbeda pada kolom yang sama menyatakan berbeda pada tingkat kepercayaan 99 %.
Hasil pengelompokkan protein susu harian berdasarkan bulan dan periode laktasi (Tabel 8 dan 9) menunjukkan bahwa nilai rataan protein susu setiap bulan laktasi maupun setiap periode laktasi berbeda nyata (P<0.01). Nilai rataan protein susu harian berdasarkan bulan laktasi terendah terjadi pada bulan laktasi pertama. Rataan protein susu pada laktasi bulan pertama memiliki nilai paling rendah karena pada bulan pertama produksi susu mencapai puncaknya. Caccamo et al. (2008) melaporkan bahwa protein susu berdasarkan pengukuran harian mengikuti pola yang teratur pada setiap laktasi. Pada puncak laktasi, kandungan protein susu akan mencapai nilai terendah (Phillip 2001) dan akan meningkat pada minggu ke-12 (Phillip 2001; Tyler dan Ensminger 2006). Lebih lanjut Jenkins dan McGuired (2006) menyatakan bahwa pada puncak laktasi terjadi peningkatan kebutuhan energi
sehingga jika tidak diimbangi dengan pemberian pakan dengan kualitas dan kuantitas yang baik akan menyebabkan penurunan kandungan protein dan lemak dalam susu.
Pengelompokkan protein susu harian berdasarkan periode laktasi (Tabel 9) menunjukkan bahwa nilai rataan protein susu setiap periode laktasi berbeda nyata (P<0.01). Nilai rataan protein susu tertinggi diperoleh pada periode laktasi kesatu. Caccamo et al. (2008) dan De Ross et al. (2004) melaporkan bahwa pada tiga periode laktasi yang pertama protein susu akan mengalami peningkatan yang signifikan. Hasil penelitian ini menunjukkan bahwa nilai rataan protein susu setiap periode laktasi tidak
menunjukkan trend peningkatan. Keadaan tersebut kemungkinan disebabkan karena dukungan manajemen dan pakan yang kurang memadai.
Kadar Lemak Susu
76
disebabkan proporsi hijauan pakan pada kondisi pemelihaaraan semi intensif lebih tinggi jika dibandingkan pada kondisi pemelihaaraan intensif. Pada sistem pemeliharaan semi intensif, peternak memberikan pakan hijauan secara berlebihan (ad libitum) sebanyak tiga kali sehari. Phillip (2001) serta Tyler dan Ensminger (2006) menyatakan bahwa tingginya proporsi hijauan dalam pakan ternak perah menyebabkan peningkatan kadar lemak susu yang dihasilkan.
Tabel 10. Nilai rataan lemak susu harian yang dikelompokkan berdasarkan bulan laktasi pada kondisi pemeliharaan yang berbeda.
Lemak (%) TD1 3.416aA±0.536 3.266aA±0.792 3.312A±0.719
(10) (23) (33)
TD2 3.216aA±0.419 3.141aA±0.839 3.170A±0.694
(9) (14) (23)
TD3 3.241aA±0.639 3.443aA±0.735 3.381A±0.701
(8) (18) (26)
TD4 3.165bA±0.491 3.655aA±0.759 3.471A±0.706
(15) (25) (40)
TD5 3.109bA±0.241 3.676aA±0.759 3.443A±0.660
(14) (20) (34)
TD6 - 3.508A±0.560 3.508A±0.560
(16) (16)
Rataan 3.215b±0.459 3.466a±0.756 3.384±0.683
(56) (116) (172)
Keterangan : (…) jumlah sampel
TD1- TD6 (Nilai rataan produksi susu harian pada bulan ke-1 sampai bulan ke-6) Superskip huruf kecil yang berbeda pada baris yang sama menyatakan berbeda pada tingkat kepercayaan 95 %.
Superskip huruf kapital yang berbeda pada kolom yang sama menyatakan berbeda pada tingkat kepercayaan 95 %.
77
Tabel 11. Nilai rataan lemak susu harian yang dikelompokkan berdasarkan periode laktasi pada kondisi pemeliharaan yang berbeda.
Lemak (%) Laktasi ke-1 3.205bA±0.459 3.618aA±0.662 3.285A±0.524
(46) (11) (57)
Laktasi ke-2 - 3.447A±0.813 3.447A±0.813
(24) (24)
Laktasi ke-3 3.193aA±0.563 3.369aA±0.724 3.339A±0.695
(7) (33) (40)
Laktasi ke-4 3.410aA±0.163 3.460aA±0.785 3.456A±0.754
(3) (35) (38)
Laktasi ke-5 - 3.636A±0.795 3.636A±0.795
(13) (13)
Rataan 3.215b±0.459 3.466a±0.756 3.384±0.683
(56) (116) (172)
Keterangan : (…) jumlah sampel
Superskip huruf kecil yang berbeda pada baris yang sama menyatakan berbeda pada tingkat kepercayaan 95 %.
Superskip huruf kapital yang berbeda pada kolom yang sama menyatakan berbeda pada tingkat kepercayaan 95 %.
Amplifikasi Gen β-Kasein
Amplifikasi ruas gen β-kasein dilakukan pada mesin thermal cycler(ESCO) dengan suhu annealing 59oC selama 45 menit. Panjang gen β-kasein yang teramplifikasi dalam penelitian adalah 233 pb sesuai panjang ruas DNA yang diapit oleh primer pada sekuen gen β-kasein (nomor akses gen bankX14711 dan AJ29330). Posisi penempelan pasangan primer pada sekuen gen β-kasein ekson 7 disajikan pada Gambar 7. Hasil amplifikasi ruas gen β-kasein yang divisualisasikan pada gel