IDENTIFIKASI KERAGAMAN GEN HORMON PERTUMBUHAN
(GH|
Msp
I) PADA SAPI FRIESIAN HOLSTEIN DI BIB LEMBANG,
BBIB SINGOSARI, DAN BET CIPELANG
SKRIPSI
DINY WIDYANINGRUM
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
RINGKASAN
DINY WIDYANINGRUM. D14070111. 2011. Identifikasi Keragaman Gen Hormon Pertumbuhan (GH|MspI) pada Sapi Friesian Holstein di BIB Lembang, BBIB Singosari, dan BET Cipelang. Skripsi. Departemen Ilmu Produksi dan Teknologi Peternakan. Fakultas Peternakan. Institut Pertanian Bogor. Pembimbing Utama : Prof. Dr. Ir. Cece Sumantri, M.Agr.Sc.
Pembimbing Anggota : Ir. Anneke Anggraeni, M.Si, Ph.D.
Upaya dalam meningkatkan produktivitas ternak dapat dilakukan melalui seleksi pada level DNA. Teknik PCR-RFLP dapat digunakan untuk mendeteksi adanya keragaman gen yang berhubungan dengan sifat ekonomis seperti sifat pertumbuhan dan produksi. Gen hormon pertumbuhan (GH) merupakan penyandi hormon pertumbuhan yang dihasilkan oleh somatotropes dalam kelenjar hipofisa bagian depan dan berperan dalam pertumbuhan jaringan, reproduksi, laktasi, serta metabolisme. Adanya keragaman gen hormon pertumbuhan diharapkan dapat menjadi informasi dasar seleksi berdasarkan penciri DNA.
Penelitian ini bertujuan untuk mengidentifikasi keragaman gen GH|MspI pada sapi Friesian Holstein (FH) dan sapi pedaging sebagai pembanding. Sampel darah yang digunakan berasal dari 89 ekor sapi FH dari BIB Lembang (17 ekor), BBIB Singosari (32 ekor), dan BET Cipelang (40 ekor); serta 37 ekor sapi pedaging (Simental, Limousin, Angus, dan Brahman) dari BET Cipelang. Amplifikasi gen GH dilakukan dengan teknik PCR, sedangkan untuk menentukan genotipe dilakukan dengan teknik PCR-RFLP menggunakan enzim restriksi MspI yang mengenali situs potong C|CGG. Analisis data yang digunakan adalah frekuensi genotipe, frekuensi alel, keseimbangan Hardy-Weinberg, dan heterozigositas.
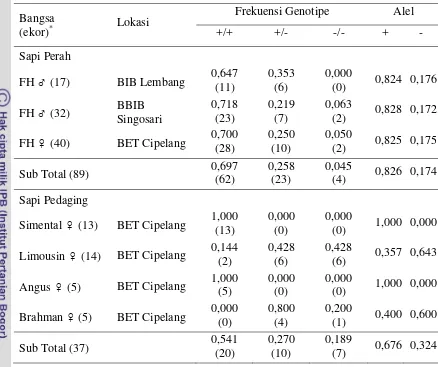
Amplifikasi gen GH menghasilkan fragmen dengan panjang 327 pb, yang terletak pada intron 3 dan ekson 4. Ada tiga genotipe yang teridentifikasi, yaitu GH|MspI (+/+), (+/-), dan (-/-); dengan dua tipe alel, yaitu GH|MspI (+) dan (-). Bangsa sapi FH di tiga lokasi memiliki frekuensi genotipe GH|MspI (+/+) lebih tinggi (0,697) dibandingkan dengan frekuensi genotipe GH|MspI (+/-) (0,258) dan GH|MspI (-/-) (0,045). Frekuensi genotipe GH|MspI (+/+) pada sapi Simental dan Angus sangat tinggi (1,000), sedangkan pada sapi Limousin dan Brahman sangat rendah (0,144 dan 0,000).
Gen GH|MspI pada sapi FH di tiga lokasi bersifat polimorfik dengan alel GH|MspI (+) tertinggi (0,826). Pada sapi Limousin dan Brahman di BET Cipelang bersifat polimorfik dengan alel GH|MspI (-) tertinggi (0,643 dan 0,600), sebaliknya pada sapi Simental dan Angus bersifat monomorfik yang seluruhnya memiliki alel GH|MspI (+) (1,000). Seluruh bangsa sapi FH berada dalam keseimbangan Hardy-Weinberg (χ2 < χ2(0,05)), namun sebaliknya pada seluruh sapi pedaging di BET
Cipelang (χ2> χ2(0,05)). Analisis heterozigositas menunjukkan tingkat keragaman gen
GH|MspI pada seluruh sapi FH adalah rendah, demikian pula pada sapi pedaging, kecuali pada sapi Brahman. Adanya keragaman gen GH|MspI dapat dijadikan pertimbangan sebagai informasi dasar seleksi ternak unggul terhadap sifat pertumbuhan pada sapi FH dan sapi pedaging.
ABSTRACT
Identification of The Growth Hormone (GH|MspI) Gene Polymorphism in Holstein Friesian Cattle in BIB Lembang, BBIB Singosari, and
BET Cipelang
Widyaningrum, D., C. Sumantri, and A. Anggraeni
Growth hormone (GH) is an anabolic hormone synthesized and secreted by somatotroph cells from the anterior lobe of the pituitary. The GH plays an important role in postnatal growth and development, tissue growth, lactation, reproduction, and metabolism. This study was aimed to identify polymorphism of the growth hormone gene (GH) in dairy cattle and beef cattle as a comparison. Holstein Friesian (HF) cattle for a total number of 89 heads from BIB Lembang (17), BBIB Singosari (32) and BET Cipelang (40); and four breeds of beef cattle for a total number of 37 heads from BET Cipelang were genotyped at intron 3 of the GH gene using PCR-RFLP method by MspI restriction enzyme. Genotyping the GH gene resulted in three genotypes, namely GH|MspI (+/+), (+/-), and (-/-), with two alleles, namely GH|MspI (+) and (-). Genetic polymorphism was detected in HF cattle and beef cattle, the exception was for Simental and Angus. The frequency of the GH|MspI (+) allele contrast to the GH|MspI (-) allele for HF cattle were 0.826 vs 0.174. GH|MspI (+/+) genotype had the highest frequency for HF (0,826), Simental, and Angus (1,000); but very limited for Limousin and Brahman (0,144 and 0,000). Chi-Square analysis showed that HF from the three locations were in Hardy-Weinberg equilibrium (χ2< χ2
(0,05)), but contrast on beef cattle (χ2 > χ2(0,05)). The value of heterozigosity
expectation (He) for HF and beef cattles were estimated between 0.000-0.800. The genetic variation of the GH|MspI gene was low in most of these cattles, the exception was for Brahman.
Keywords : Holstein Friesian, GH|MspI gene, Genetic Polymorphism, Beef Cattle, PCR-RFLP
IDENTIFIKASI KERAGAMAN GEN HORMON PERTUMBUHAN
(GH|
Msp
I) PADA SAPI FRIESIAN HOLSTEIN DI BIB LEMBANG,
BBIB SINGOSARI, DAN BET CIPELANG
DINY WIDYANINGRUM D14070111
Skripsi ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan pada
Fakultas Peternakan Institut Pertanian Bogor
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
Judul : Identifikasi Keragaman Gen Hormon Pertumbuhan (GH|MspI) pada Sapi Friesian Holstein di BIB Lembang, BBIB Singosari, dan BET Cipelang
Nama : Diny Widyaningrum NIM : D14070111
Menyetujui,
Pembimbing Utama, Pembimbing Anggota,
(Prof. Dr. Ir. Cece Sumantri, M. Agr.Sc.) (Ir. Anneke Anggraeni, M.Si., Ph.D.) NIP. 19591212 198603 1 004 NIP. 19630924 199803 2 001
Mengetahui : Ketua Departemen,
Ilmu Produksi dan Teknologi Peternakan
(Prof. Dr. Ir. Cece Sumantri, M. Agr.Sc.) NIP. 19591212 198603 1 004
RIWAYAT HIDUP
Penulis dilahirkan pada tanggal 29 Mei 1989 di Bogor, Jawa Barat. Penulis merupakan anak pertama dari dua bersaudara dari pasangan Bapak Eman Sulaeman, S.Pd, M.M. dan Ibu Nia Kania MS., SE, M.MPd. Pendidikan dasar diselesaikan pada tahun 2001 di SD Negeri Pengadilan 3, Bogor. Pendidikan lanjutan menengah pertama diselesaikan pada tahun 2004 di SLTP Negeri 2, Bogor, dan pendidikan lanjutan menengah atas diselesaikan pada tahun 2007 di SMA Negeri 6, Bogor, Jawa Barat.
Penulis diterima sebagai mahasiswa pada Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan Institut Pertanian Bogor melalui jalur Undangan Seleksi Masuk IPB (USMI) pada tahun 2007. Selama mengikuti pendidikan, penulis aktif di Himpunan Mahasiswa Ilmu Produksi dan Teknologi Peternakan (HIMAPROTER), Fakultas Peternakan IPB periode 2008-2009 sebagai staf club Ruminansia dan periode 2009-2010 sebagai Badan Pengawas Himpro; serta menjadi anggota Animal Breeding and Genetic Student Community (ABGSCi) periode 2010-2011. Penulis berkesempatan mendapatkan beasiswa BBM tahun 2009 dan PPA (Peningkatan Prestasi Akademik) tahun 2010 dan 2011. Selain itu, penulis juga berperan aktif dalam kepanitiaan di Institut Pertanian Bogor.
KATA PENGANTAR
Puji dan syukur senantiasa penulis panjatkan kepada Allah SWT atas segala rahmat dan karunia-Nya sehingga penelitian dan penulisan skripsi ini dapat diselesaikan. Shalawat dan salam senantiasa dicurahkan kepada Nabi Muhammad SAW. Skripsi yang berjudul Identifikasi Keragaman Gen Hormon Pertumbuhan (GH|MspI) pada Sapi Friesian Holstein di BIB Lembang, BBIB Singosari, dan BET Cipelang ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan, Fakultas Peternakan, Institut Pertanian Bogor.
Ternak sapi perah merupakan salah satu komoditas yang memiliki berbagai manfaat. Namun, perkembangan populasi dan produktivitas sapi perah di Indonesia dalam pemenuhan konsumsi masyarakat belum optimal. Upaya dalam meningkatkan produktivitas ternak dapat dilakukan melalui seleksi pada level DNA. Salah satunya yaitu dengan teknik PCR-RFLP yang digunakan untuk mendeteksi adanya keragaman gen yang berhubungan dengan sifat ekonomis seperti sifat pertumbuhan dan produksi.
Penelitian ini bertujuan untuk mengidentifikasi keragaman gen hormon pertumbuhan dengan PCR-RFLP pada sapi FH serta sapi pedaging sebagai pembanding agar dapat diketahui informasi dasar genetik untuk seleksi ternak-ternak unggul. Hasil dari penelitian ini diharapkan dapat bermanfaat dalam upaya meningkatkan produktivitas ternak sapi di Indonesia. Penulis menyadari sepenuhnya bahwa dalam penulisan skripsi ini masih jauh dari sempurna, namun penulis berharap semoga skripsi ini dapat bermanfaat dan memberikan sumbangan terhadap kemajuan dunia peternakan di Indonesia. Amin.
Bogor, 11 April 2011
Penulis
DAFTAR ISI
Polymerase Chain Reaction – Restriction Fragment Length Polymorphism (PCR-RFLP) ... 7
Frekuensi Genotipe dan Frekuensi Alel ... 13
Keseimbangan Hardy-Weinberg ... 14
viii
HASIL DAN PEMBAHASAN ... 16
Amplifikasi Gen Hormon Pertumbuhan (GH) ... 16
Keragaman Gen GH|MspI ... 17
Frekuensi Genotipe dan Frekuensi Alel ... 19
Keseimbangan Hardy-Weinberg ... 23
Heterozigositas ... 24
KESIMPULAN DAN SARAN ... 26
Kesimpulan ... 26
Saran ... 26
UCAPAN TERIMA KASIH ... 27
DAFTAR PUSTAKA ... 28
LAMPIRAN ... 33
DAFTAR TABEL
Nomor Halaman 1. Sampel Darah Sapi Perah dan Sapi Pedaging yang Digunakan ... 10 2. Frekuensi Genotipe dan Frekuensi Alel dari Gen GH|MspI pada
Sapi FH dan Sapi Pedaging ... 20 3. Keseimbangan Hardy-Weinberg (HW) Berdasarkan Uji χ2 ... 23 4. Nilai Heterozigositas Pengamatan (Ho) dan Nilai Heterozigositas
DAFTAR GAMBAR
Nomor Halaman 1. Rekonstruksi Struktur Gen GH ... 6 2. Visualisasi Amplifikasi PCR Fragmen Gen GH ... 16 3. Posisi Penempelan Primer, Perbedaan Fragmen Gen GH dan Situs
Pemotongan Enzim Restriksi MspI Berdasarkan Sekuen Gen GH
Sapi pada GenBank (Kode Akses : M57764) ... 17 4. Visualisasi PCR-RFLP Fragmen Gen GH|MspI ... 18 5. Keragaman Gen GH|MspI pada Sapi FH dan Sapi Pedaging ... 19
PENDAHULUAN Latar Belakang
Sapi perah merupakan salah satu sumber plasma nutfah yang berperan dalam memenuhi ketersediaan pangan. Perkembangan populasi sapi perah di Indonesia hingga saat ini belum mencapai angka optimal dalam pemenuhan pangan masyarakat. Berdasarkan data statistik peternakan, populasi sapi perah di Indonesia pada tahun 2008 sebesar 407,8 ribu ekor dan pada tahun 2009 hanya meningkat hingga mencapai 423,8 ribu ekor, dengan tingkat konsumsi susu sebesar 9,53 kg per kapita per tahun. Rataan produksi sapi perah di Indonesia masih di bawah angka 10 liter per ekor per hari. Terbatasnya populasi dan produktivitas sapi perah mengakibatkan produksi susu dari dalam negeri hanya mampu mensuplai sekitar 23 persen dari kebutuhan susu nasional, sedangkan kekurangannya masih harus impor (Direktorat Jenderal Peternakan, 2009).
Hingga saat ini, usaha pengembangan populasi dan produktivitas ternak telah dilakukan oleh berbagai institusi unit pelaksana teknis dari Direktorat Jenderal Peternakan, seperti BBIB, BIB, dan BET. Balai Inseminasi Buatan (BIB) Lembang dan Balai Besar Inseminasi Buatan (BBIB) Singosari merupakan unit pelaksana teknis yang melaksanakan replacement pejantan dan produksi bibit unggul secara berkesinambungan melalui penyediaan semen beku. Balai Embrio Ternak (BET) Cipelang merupakan unit pelaksana teknis yang melakukan produksi, penyimpanan, dan pendistribusian embrio ternak serta aplikasi transfer embrio ternak. Berbagai unit tersebut memiliki peran dan fungsi penting untuk meningkatkan kualitas bibit unggul.
2 untuk mendeteksi lokasi genetik dalam kromosom yang menyandikan atau mendeteksi adanya keragaman gen yang berhubungan dengan sifat ekonomis seperti sifat pertumbuhan dan produksi.
Sifat pertumbuhan ternak dikendalikan oleh gen-gen pengontrol pertumbuhan. Salah satu gen yang dapat mempengaruhi pertumbuhan adalah gen hormon pertumbuhan (Growth Hormone Gene). Gen hormon pertumbuhan (GH) merupakan penyandi hormon pertumbuhan yang dihasilkan oleh somatotropes, dalam kelenjar hipofisa bagian depan dan memiliki beberapa aktivitas fisiologi. Gen GH berperan penting dalam mengatur sifat-sifat pertumbuhan, reproduksi, metabolisme, laktasi, dan perkembangan kelenjar susu. Gen tersebut dapat dijadikan sebagai kandidat gen dalam program Marker Asissted Selection (MAS). Penerapan MAS memerlukan marker molekuler yang dapat diperoleh melalui teknik PCR-RFLP, PCR-SSCP, DGGE, maupun analisis sekuen (sequencing). Oleh karena itu, adanya keragaman gen hormon pertumbuhan diharapkan dapat menjadi informasi dasar seleksi berdasarkan penciri DNA untuk meningkatkan sifat pertumbuhan, produksi, serta kualitas susu sapi perah di Indonesia. Selain itu, pada sapi pedaging juga dapat menjadi informasi dasar seleksi dengan melihat tingkat keragaman gen GH yang memiliki peran berbeda, yaitu untuk pertumbuhan dan produksi karkas.
Tujuan
Penelitian ini bertujuan untuk mengidentifikasi keragaman gen hormon pertumbuhan menggunakan enzim restriksi MspI (GH|MspI) dengan metode PCR-RFLP pada sapi Friesian Holstein dari BIB Lembang, BBIB Singosari, dan BET Cipelang serta digunakan sapi pedaging dari BET Cipelang sebagai pembanding untuk mengetahui informasi keragaman gen hormon pertumbuhannya.
TINJAUAN PUSTAKA Sapi
Ternak sapi secara zoologi termasuk ke dalam kingdom Animalia, filum
Chordata, sub filum Vertebrata, kelas Mamalia, ordo Artiodactyla, famili Bovidae, genus Bos, dan spesies Bos taurus (sapi Eropa), Bos indicus (sapi bergumba), dan
Bos sondaicus (Blakely dan Bade, 1998). Spesies Bos taurus memiliki keunggulan pada tingkat pertumbuhan dan produksi yang tinggi, sedangkan spesies Bos indicus
lebih unggul dalam hal adaptasinya (resisten pada kondisi lingkungan yang kurang baik) (Gorbani et al., 2009), namun Bos indicus memiliki kelemahan yaitu membutuhkan waktu yang lebih lama untuk mencapai dewasa kelamin dengan periode kebuntingan yang lebih panjang (Parakkasi, 1999). Pengelompokan sapi juga dapat didasarkan pada tujuan produksinya, yaitu tipe sapi perah, tipe sapi pedaging, dan tipe campuran.
Sapi Perah
Bangsa sapi perah yang terdapat di dunia pada dasarnya dapat dikelompokkan menjadi dua, yaitu kelompok sapi perah sub-tropis dan kelompok sapi perah tropis. Menurut Ensminger dan Tyler (2006), bangsa-bangsa sapi perah subtropis, yaitu
Friesian Holstein, Yersey, Guernsey, Ayrshire, dan Brown Swiss. Bangsa-bangsa sapi perah tropis, yaitu Red Sindi, Sahiwal, dan PFH (Peranakan Fries Holland). Sapi Friesian Holstein (FH) menduduki populasi terbesar hampir di seluruh dunia, baik negara subtropis maupun tropis.
Sapi FH berasal dari nenek moyang sapi liar Bos taurus, Typicus primigenius
4 mencapai 4500-5500 liter per laktasi. Berat badan sapi FH jantan dapat mencapai 1000 kg dan sapi FH betina 650 kg (Chandra et al., 2009).
Peternakan sapi perah dapat dijadikan sumber penghasil susu yang efisien dan secara komersial umum ditemukan di negara-negara seperti Australia, Inggris dan Amerika. Menurut Buckle et al. (2007), seekor sapi perah yang baik akan menghasilkan sekitar 5000 liter susu per tahun (kira-kira sepuluh kali berat badannya sendiri). Di Indonesia, rataan produksi susu sapi perah mencapai 3000 kg/ekor/laktasi (Direktorat Jenderal Peternakan, 2008). Sifat produksi susu pada sapi perah adalah sifat kuantitatif yang dapat dikendalikan oleh banyak gen dan diwariskan serta dipengaruhi oleh faktor lingkungan (Noor, 2000).
Pertumbuhan, reproduksi, dan produktivitas sapi perah dapat dipengaruhi oleh pakan dan manajemen pemeliharaan. Suhu lingkungan yang optimum untuk pemeliharaan sapi perah berkisar antara 5-21 oC, dengan kisaran kelembaban 50-75% (Ensminger dan Tyler, 2006). Pada tingkat pakan tinggi, sapi Holstein dapat mencapai pubertas pada umur 262 hari, sedangkan pada tingkat pakan rendah, pubertas terjadi pada umur 504 hari atau lebih (Tomaszewska et al., 1991). Sifat reproduksi pada peternakan sapi perah rakyat di Indonesia, seperti pada sapi FH menunjukkan umur pertama beranak adalah 3,5 tahun (3-4 tahun), masa kering 45-60 hari, masa kosong 60 hari, calving interval 15-16 bulan, dan service per conception
(S/C) = 2 (Dudi et al., 2006). Sapi Pedaging
Sapi pedaging memiliki keunggulan dalam menghasilkan karkas berkualitas dan tingkat pertumbuhan yang tinggi. Beberapa bangsa sapi pedaging dari spesies
5 Pemeliharaan sapi potong untuk mempercepat kenaikan bobot badan dapat dilakukan dengan metode penggemukkan yang terdiri atas sistem penggemukkan ekstensif (pasture fattening) dan sistem penggemukkan intensif (dry lot fattening). Sapi yang digemukkan secara intensif memiliki laju pertumbuhan yang lebih tinggi daripada sapi dipelihara pada sistem ekstensif, sehingga waktu yang diperlukan untuk mencapai bobot tertentu menjadi lebih singkat. Sistem pemeliharaan yang bervariasi menyebabkan tingginya keragaman pada respon pertumbuhan sapi (Parakkasi, 1999).
Hormon Pertumbuhan
Menurut Lawrence dan Fowler (2002), pertumbuhan merupakan suatu proses deposisi, pemindahan substansi sel-sel, serta peningkatan ukuran dan jumlah sel pada tingkat dan titik yang berbeda dalam suatu waktu tertentu. Pertumbuhan dikarakterisasikan oleh peningkatan jumlah sel pada jaringan (hyperplasia) dan peningkatan ukuran sel (hypertrophy). Pertumbuhan ternak dapat dipengaruhi oleh faktor lingkungan dan faktor genetik, ataupun interaksi keduanya. Salah satu faktor genetik yang berperan dalam pertumbuhan suatu individu adalah gen GH (growth hormone) atau lebih dikenal dengan gen hormon pertumbuhan.
Hormon pertumbuhan (growth hormone) merupakan hormon peptida yang secara alami dihasilkan oleh somatotropes, subclass dari sel hipofisa acidophilic
yang terletak dalam kelenjar hipofisa bagian depan (Reis et al., 2001). Hormon pertumbuhan adalah salah satu faktor yang paling penting dalam pertumbuhan dan perkembangan sel hewan (Pierzchala et al., 2004). Hormon pertumbuhan pada ruminansia diketahui bertanggung jawab untuk galactopoiesis dan persistensi laktasi (Svennersten-Sjaunja dan Olsson, 2005), sehingga sapi perah yang dipilih untuk produksi susu tinggi diharapkan dapat melepaskan sejumlah besar GH endogen dari rata-ratanya.
Gen Hormon Pertumbuhan
6 segmen depan (leader segment) dan segmen belakang (trailer segment) yang mengapit daerah CDS. Beberapa gen pada eukaryot bersifat tidak kontinyu karena adanya ekson (pengkode protein) dan intron (space internal antara pengkode protein). Pada saat transkripsi, bagian intron hilang (splicing), sehingga proses translasi berjalan baik (Brown, 1999).
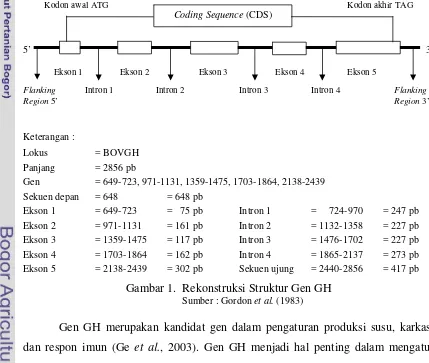
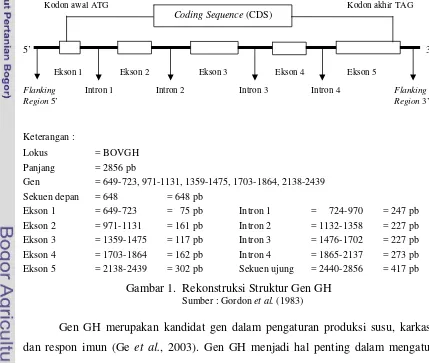
Bovine Growth Hormone (bGH) merupakan sebuah peptida tunggal dengan berat molekul 22 KDa dan disusun oleh 191 asam amino (Wallis, 1973) dengan panjang sekuen nukleotida 2856 pb (Gordon et al., 1983). Gen hormon pertumbuhan sapi Bos taurus (bovine growth hormone gene) terdiri dari lima ekson dan dipisahkan oleh empat intron (Gordon et al., 1983) dan terletak pada kromosom 19 (Hediger et al., 1990). Rekonstruksi struktur gen GH dapat digambarkan berdasarkan sekuens gen GH di GenBank (nomor akses : M57764) (Gambar 1).
Gen = 649-723, 971-1131, 1359-1475, 1703-1864, 2138-2439
Sekuen depan = 648 = 648 pb
Gambar 1. Rekonstruksi Struktur Gen GH
Sumber : Gordon et al. (1983)
Gen GH merupakan kandidat gen dalam pengaturan produksi susu, karkas, dan respon imun (Ge et al., 2003). Gen GH menjadi hal penting dalam mengatur sifat-sifat pada ternak yang bernilai ekonomi tinggi, sehingga Beauchemin et al.
(2006) menyatakan bahwa gen GH dapat dijadikan kandidat gen dalam program
7
Marker Asissted Selection pada sapi. Gen GH juga berperan sebagai pengatur utama pada pertumbuhan pasca kelahiran, perkembangan jaringan, otot, tulang, dan jaringan adiposa, pertumbuhan kelenjar mamary, laktasi, reproduksi, serta metabolisme karbohidrat, protein, dan lemak dalam tubuh (Akers, 2006). Gen GH membutuhkan receptor dalam mekanisme ekspresinya ke target jaringan. Menurut Zhou dan Jiang (2005), pada tingkatan jaringan, aksi biologis dari gen GH dimediasi oleh gen GHR.
Polymerase Chain Reaction – Restriction Fragment Length Polymorphism (PCR-RFLP)
Polymerase Chain Reaction (PCR) merupakan suatu reaksi in vitro untuk menggandakan molekul DNA pada target tertentu dengan cara mensintesis molekul DNA baru yang berkomplemen dengan molekul DNA tersebut dengan bantuan enzim polymerase dan oligonukleotida pendek sebagai primer dalam mesin
thermocycler (Muladno, 2002). RFLP adalah profil DNA berupa fragmen-fragmen DNA hasil pemotongan enzim endonuklease untuk berbagai individu. Enzim endonuklease atau enzim restruksi (RE) yang mengenali situs pemotongan empat dan enam basa umum dipakai untuk analisis keragaman genetik menggunakan pendekatan analisis RFLP (Green, 1998). Penciri molekuler DNA restriction fragment length polymorphism (RFLP) memiliki tingkat polimorfisme yang tinggi dan secara luas telah digunakan untuk mendapatkan gambaran populasi genetik dan juga untuk mengidentifikasi gen-gen yang mengkode sifat-sifat penting (Montaldo & Herrera, 1998). Analisis RFLP dapat digunakan untuk mendeteksi adanya keragaman gen yang berhubungan dengan sifat ekonomis, seperti produksi dan kualitas susu (Sumantri et al., 2004) dan kualitas karkas (Beauchemin et al., 2006)
8 Keragaman Genetik
Keragaman genetik dalam suatu populasi digunakan untuk mengetahui dan melestarikan bangsa-bangsa dalam populasi terkait dengan penciri suatu sifat khusus, serta menentukan hubungan antar subpopulasi yang terfragmentasi dalam suatu spesies (Hartl dan Clark, 1997). Keragaman genetik antara subpopulasi dapat diketahui dengan melihat persamaan dan perbedaan frekuensi alel dan genotipe di antara subpopulasi (Li et al., 2000). Suatu alel dikatakan polimorfik jika memiliki frekuensi alel sama dengan atau kurang dari 0,99 (Nei, 1987). Hukum Hardy-Weinberg menyatakan frekuensi genotipe suatu populasi yang cukup besar akan selalu dalam keadaan seimbang bila tidak ada seleksi, migrasi, mutasi, dan genetic drift; selain itu silang dalam dan silang luar juga dapat mempengaruhi frekuensi genotipe (Noor, 2008). Estimasi perhitungan keragaman genetik dalam populasi secara kuantitatif dapat diperoleh melalui dua ukuran keragaman variasi populasi, yaitu proporsi lokus polimorfisme dalam populasi dan rata-rata proporsi individu heterozigot dalam setiap lokus (Nei dan Kumar, 2000).
Pendugaan nilai heterosigositas diperoleh untuk mendapatkan keragaman genetik dalam populasi yang dapat digunakan untuk membantu program seleksi pada ternak yang akan digunakan sebagai sumber genetik pada generasi berikutnya (Marson et al., 2005). Menurut Javanmard et al. (2005), nilai heterozigositas di bawah 0,5 (50%) mengindikasikan rendahnya variasi suatu gen dalam populasi dan jika nilai Ho lebih rendah dari He maka dapat mengindikasikan adanya proses seleksi yang intensif (Machado et al., 2003; Tambasco et al., 2003). Avise (1994) menyatakan bahwa semakin tinggi derajat heterozigositas suatu populasi maka daya hidup populasi tersebut akan semakin tinggi. Seiring dengan menurunnya derajat heterozigositas akibat dari silang dalam dan fragmentasi populasi, sebagian besar alel resesif yang bersifat lethal semakin meningkat frekuensinya.
Keragaman Gen Hormon Pertumbuhan
9 dilaporkan sebelumnya pada sapi Nadji (Rastegari et al., 2010); serta sapi South Anatolian dan East Anatolian Red (Yardibi et al., 2009).
Identifikasi mutasi pada hormon pertumbuhan dapat diseleksi pada tingkat DNA (Khatami et al., 2005). Cowan et al. (1989) mendeteksi keragaman lokus gen menggunakan enzim restriksi MspI dan berdasarkan data PCR-RFLP telah diketahui bahwa gen GH memiliki keragaman yang tinggi akibat adanya mutasi. Mutasi dapat terjadi pada level DNA akibat adanya perubahan basa-basa DNA (A = Adenin, T = Timin, G = Guanin, S = Sitosin) dalam bentuk (tipe) substitusi (transisi atau transversi), delesi, insersi dan inversi (Nei, 1987). Situs pemotongan enzim restriksi
MspI berubah akibat adanya mutasi transisi dari basa C menjadi basa T (Yao et al., 1996). Mutasi transisi dapat terjadi akibat adanya substitusi antara basa Adenin dengan Guanin (Purin) atau antara basa Sitosin dengan Timin (Pirimidin) (Paolella, 1997).
Keragaman gen GH|MspI terletak pada intron 3 dari gen hormon pertumbuhan pada posisi sekuen 1547 (Zhang et al., 1993) dan panjang fragmen gen GH|MspI berdasarkan hasil yang diperoleh Zhou et al. (2005), yaitu 329 pb. Keragaman gen GH|MspI telah dilaporkan pada berbagai ternak seperti sapi Holstein Beijing yang menunjukkan adanya tiga genotipe, yaitu GH|MspI (+/+) (224 pb, 105 pb), GH|MspI (+/-) (329 pb, 224 pb, 105 pb), dan GH|MspI (-/-) (329 pb). Keragaman gen GH pada sifat produksi susu menunjukkan bahwa sapi bergenotipe GH|MspI (+/+) memiliki tingkat produksi susu dan protein susu yang lebih tinggi serta persentase lemak lebih sedikit dibandingkan sapi bergenotipe GH|MspI (+/-), dengan frekuensi alel rata-rata sebesar 0,875 untuk alel GH|MspI (+) (Zhou et al.,
2005). Menurut Thomas et al. (2006), fragmen GH|MspI pada sapi Brangus bergenotipe GH|MspI (+/-) (heterozigot) memiliki pengaruh positif terhadap pertambahan bobot badan harian dan karkas; selain itu, genotip GH|MspI (+/+) dan GH|MspI (+/-) fragmen GH|MspI berpengaruh positif pada sifat bobot badan dan kualitas daging (Unanian et al., 2000).
MATERI DAN METODE Lokasi dan Waktu
Penelitian ini dilaksanakan di Laboratorium Genetika Molekuler Ternak, Bagian Pemuliaan dan Genetika, Fakultas Peternakan, Institut Pertanian Bogor. Penelitian ini berlangsung dari bulan Agustus sampai dengan Nopember 2010.
Materi Sampel
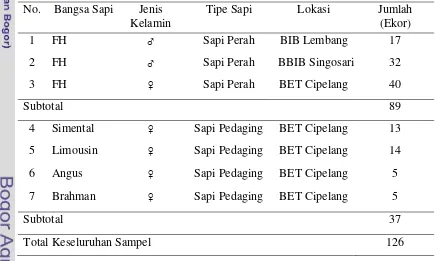
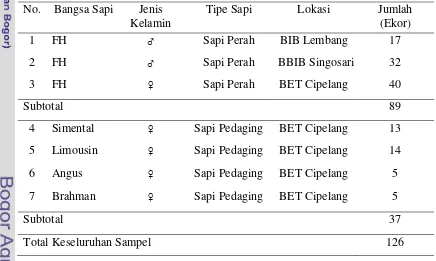
Sampel yang digunakan sebanyak 126 ekor sapi meliputi 89 ekor sapi Friesian Holstein dari BIB Lembang, BBIB Singosari, dan BET Cipelang; serta 37 ekor sapi pedaging (Simental, Limousin, Angus, dan Brahman) dari BET Cipelang sebagai pembanding (Tabel 1). Sampel-sampel tersebut berupa sampel darah yang merupakan koleksi Laboratorium Genetika Molekuler Ternak, Bagian Pemuliaan dan Genetika, Fakultas Peternakan, Institut Pertanian Bogor.
Tabel 1. Sampel Darah Sapi Perah dan Sapi Pedaging yang Digunakan No. Bangsa Sapi Jenis
Kelamin
Tipe Sapi Lokasi Jumlah (Ekor)
1 FH ♂ Sapi Perah BIB Lembang 17
2 FH ♂ Sapi Perah BBIB Singosari 32
3 FH ♀ Sapi Perah BET Cipelang 40
Subtotal 89 4 Simental ♀ Sapi Pedaging BET Cipelang 13
5 Limousin ♀ Sapi Pedaging BET Cipelang 14
6 Angus ♀ Sapi Pedaging BET Cipelang 5
7 Brahman ♀ Sapi Pedaging BET Cipelang 5
Subtotal 37
Total Keseluruhan Sampel 126
Keterangan : ♂= jantan dan ♀ = betina
Penanganan dan Pengambilan Sampel
11 Ekstraksi DNA
Bahan-bahan yang digunakan dalam ekstraksi DNA adalah sampel darah 200µl, EDTA (Ethylinediamine tetraacetic), destilation water, 40 µl SDS 10% (Sodium Dodecyl Sulfat), 10 µl enzim Proteinase K 5 mg/ml, 400 µl phenol, 400 µl CIAA, 800 µl etanol absolute, etanol 70%, 40 µl NaCl 5M, 1 x STE (5 M NaCl. 2 M Tris HCL, 0,2 M EDTA), Elution Buffer, dan 100 µl TE 80% (Tris EDTA). Peralatan yang digunakan adalah tabung eppendorf 1,5 ml, satu set mikro pipet, tip,
vortexmixer, autoclave, mikrosentrifuge, rotary mixer, inkubator, refrigerator, dan
freezer.
Primer
Primer yang digunakan dalam penelitian fragmen gen GH|MspI berdasarkan sumber Mitra et al. (1995), adalah forward : 5’ CCC ACG GGC AAG AAT GAG GC, dan reverse 5’ TGA GGA ACT GCA GGG GCC CA.
Amplifikasi Gen GH|MspI
Bahan yang digunakan dalam analisa PCR-RFLP (Polymerase Chain Reaction-Restriction Fragment Length Polymorphism) adalah sampel DNA,
destilated water, 10x buffer PCR, MgCl2, pasangan primer fragmen gen GH|MspI,
enzim Taq DNA polymerase, dNTP (deoxy Nukleotida Triposfat), dan enzim restriksi MspI serta buffernya. Alat yang digunakan adalah satu set pipet mikro, sentrifuge, mesin thermocycler, rak dan tabung eppendorf, tip pipet, dan vortex. Elektoforesis dan Genotyping (Penentuan Genotipe)
Bahan yang digunakan adalah produk PCR, agarose, loading dye, marker
12 Prosedur
Pengambilan Sampel
Sampel darah diambil melalui vena jugularis menggunakan jarum vennoject
dan tabung vaccutainer tanpa heparin. Sampel darah tersebut ditambahkan etanol absolute dengan perbandingan 1 : 2 dan disimpan pada suhu ruang.
Ekstraksi DNA
Ekstraksi DNA dilakukan dari sampel darah dengan menggunakan metode Sambrook et al. (1989), yang meliputi tahapan :
Preparasi Sampel. Sampel darah 200 µl dimasukkan ke dalam tabung 1,5 ml, kemudian ditambahkan air destilasi 1000 µl. Sampel disentrifugasi pada kecepatan 8000 rpm selama 5 menit, supernatan dibuang.
Degradasi Protein. Sampel yang telah bersih dari alkohol ditambahkan 1xSTE sebanyak 350 µl, 40 µl SDS 10% dan 10 µl proteinase K 5 mg/ml, kemudian dikocok perlahan dalam inkubasi pada suhu 55 ˚C selama dua jam.
Degradasi Bahan Organik. Larutan yang telah diinkubasi ditambahkan 400 µl
phenol, 400 µl chloroform isoamyl alcohol (24:1) dan 40 µl NaCl, kemudian dikocok perlahan pada suhu ruang selama 1 jam.
Presipitasi DNA. Larutan disentrifugasi pada kecepatan 12000 rpm selama 5 menit hingga supernatan yang mengandung DNA terpisah dari larutan fenol. Supernatan sebanyak 400 µl dipindahkan ke tabung baru, ditambahkan 40 µl NaCl 5 M dan 800 µl etanol absolute, dihomogenkan, kemudian larutan di-freezing over night. Tahapan selanjutnya, disentrifugasi pada kecepatan 12000 rpm selama 5 menit, kemudian bagian supernatan dipisahkan dan ditambahkan 800 µl EtOH 70%, dan tahap ini diulang kembali, kemudian didiamkan dalam keadaan terbuka. Tahap selanjutnya ditambahkan 100 µl TE 80% dan disimpan dalam freezer sampai akan digunakan. Amplifikasi Gen GH|MspI
Amplifikasi gen GH menggunakan metode PCR. Pereaksi amplifikasi DNA yang digunakan terdiri dari sampel DNA 1µl, destilated water 9,7 µl, primer 0,1 μl,
Taq polymerase 0,05 µl dan buffer 1,25 µl, dNTP 0,1 µl, dan MgCl2 0,25 µl.
13
thermocycler dengan kondisi suhu pradenaturasi 94 °C selama 5 menit, denaturasi
94 °C selama 45 detik, annealing 62 °C selama 45 detik dan extensi 72 °C selama 1 menit, dan extensi akhir 72 °C selama 5 menit. Produk PCR dielektroforesis menggunakan agarose 1,5% untuk mengetahui panjang amplifikasi gen GH.
Elektroforesis, Genotyping (Penentuan Genotipe), dan Penentuan Alel
Penentuan genotipe menggunakan pendekatan RFLP dengan menggunakan produk PCR 5 µl yang ditambahkan 1 µl destilationwater, buffer 0,7 µl, dan enzim
MspI 0,3 µl, kemudian diinkubasi pada suhu 37 oC selama 16 jam. Produk pemotongan DNA tersebut divisualisasikan pada gel agarose 2% dengan buffer 0,5 x TBE (Tris Borat EDTA) yang diwarnai dengan ethidium bromide, dan dijalankan menggunakan power supply electrophoresis pada tegangan 100 Volt. Hasil elektroforesis diamati dengan bantuan sinar UV trans iluminator.
Pita-pita DNA yang muncul dibandingkan dengan marker untuk diketahui panjang fragmennya dan jumlah pita DNA dari setiap sampel dibandingkan untuk menentukan genotipe pita DNA. Penentuan alel GH|MspI (+) dan GH|MspI (-) ditunjukan dengan jumlah dan ukuran besarnya fragmen yang terpotong berdasarkan sekuen gen GH (Gordon et al., 1983). Alel GH|MspI (+) memiliki titik potong MspI (C|CGG) dan menunjukan adanya dua fragmen yang masing-masing panjangnya 103 pb dan 223 pb, sedangkan alel GH|MspI (-) tidak memiliki titik potong dan hanya menunjukan satu fragmen yang panjangnya 327 pb.
Analisis Data Frekuensi Genotipe dan Frekuensi Alel
14 Frekuensi alel ) merupakan rasio relatif suatu alel terhadap keseluruhan alel pada suatu lokus dalam populasi, dengan rumus sebagai berikut :
∑
Keterangan :
= frekuensi genotipe ke-ii = frekuensi alel ke-i
nii = jumlah individu bergenotipe ii nij = jumlah individu bergenotipe ij N = jumlah individu sampel
Keseimbangan Hardy-Weinberg
Keseimbangan Hardy-Weinberg diuji dengan menggunakan perhitungan Chi-Kuadrat (Hartl dan Clark, 1997) :
Keterangan :
= uji Chi-kuadrat
O = jumlah pengamatan genotipe ke-i E = jumlah harapan genotipe ke-i
Heterozigositas
Keragaman genetik dapat diketahui melalui estimasi frekuensi heterozigositas pengamatan yang diperoleh dari masing-masing lokasi, dengan menggunakan rumus Weir (1996) sebagai berikut :
Keterangan :
Ho = heterozigositas pengamatan nij = jumlah individu heterozigot N = jumlah individu yang diamati
15 Keterangan :
He = nilai heterozigositas harapan = frekuensi alel
327 pb
M 1 2 3 4
400 pb 500 pb
200 pb 300 pb
100 pb
HASIL DAN PEMBAHASAN
Amplifikasi Gen Hormon Pertumbuhan (GH)
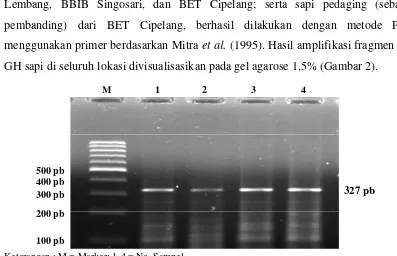
Amplifikasi gen hormon pertumbuhan pada sapi FH yang berasal dari BIB Lembang, BBIB Singosari, dan BET Cipelang; serta sapi pedaging (sebagai pembanding) dari BET Cipelang, berhasil dilakukan dengan metode PCR menggunakan primer berdasarkan Mitra et al. (1995). Hasil amplifikasi fragmen gen GH sapi di seluruh lokasi divisualisasikan pada gel agarose 1,5% (Gambar 2).
Keterangan : M = Marker; 1-4 = No. Sampel
Gambar 2. Visualisasi Amplifikasi PCR Fragmen Gen GH
Gen GH merupakan peptida tunggal dengan panjang sekuen nukleotida 2856 pb, yang terdiri dari lima ekson dan dipisahkan oleh empat intron (Gordon et al., 1983). Berdasarkan pasangan primer yang digunakan, panjang produk hasil amplifikasi fragmen gen GH adalah 327 pb, yang terletak pada intron 3 dan ekson 4. Panjang fragmen ini mendekati hasil amplifikasi Zhou et al. (2005), yaitu 329 pb.
17 Jika suhu tersebut digunakan, maka tingkat keberhasilan amplifikasi pada gen hormon pertumbuhan pada sapi ini kurang menunjukkan hasil yang optimum.
Keragaman Gen GH|MspI
Keragaman gen hormon pertumbuhan diketahui dengan menentukan alel dan genotipe pada setiap individu melalui pendekatan PCR-RFLP menggunakan enzim restriksi MspI. Enzim tersebut hanya mengenali situs pemotongan empat basa, yaitu C│CGG. Penentuan alel GH|MspI (+) dan GH|MspI (-) ditunjukkan dengan jumlah dan ukuran besarnya fragmen yang terpotong. Alel GH|MspI (+) memiliki dua fragmen dengan panjang masing-masing 104 pb dan 223 pb, sedangkan alel GH|MspI (-) hanya memiliki satu fragmen dengan panjang 327 pb. Perbedaan fragmen antara alel GH|MspI (+) dan GH|MspI (-) dapat diakibatkan oleh adanya mutasi yang menyebabkan enzim MspI mengenali situs pemotongan basa baru. Perbedaan fragmen gen GH dapat dilihat berdasarkan sekuen gen GH (dalam
GenBank, kode akses : M57764) yang terdapat pada Gambar 3 berikut, Forward
1441 cccccacggg caagaatgag gcccagcaga aatcagtgag tggcaacctc ggaccgagga
1501 gcaggggacc tccttcatcc taagtaggct gccccagctc ccgcac|cggc ctggggcggc
1561 cttctccccg aggtggcgga ggttgttgga tggcagtgga ggatgatggt gggcggtggt
1621 ggcaggaggt cctcgggcag aggccgacct tgcagggctg ccccagaccc gcggcaccca
1681 ccgaccaccc acctgccagc aggacttgga gctgcttcgc atctcactgc tcctcatcca
1741 gtcgtggctt gggcccctgc agttcctcag cagagtcttc accaacagct tggtgtttgg
Reverse
Keterangan : Alel GH|MspI (+) Mempunyai Basa C pada Posisi Basa ke-1547
Alel GH|MspI (-) Mempunyai Basa T pada Posisi Basa ke-1547
Gambar 3. Posisi Penempelan Primer, Perbedaan Fragmen Gen GH dan Situs Pemotongan Enzim Restriksi MspI Berdasarkan Sekuen Gen GH Sapi pada GenBank (Kode Akses : M57764)
Sumber : Gordon et al. (1983)
Hal ini sebanding dengan pendapat Cowan et al. (1989) yang menyatakan bahwa gen GH memiliki keragaman tinggi akibat adanya mutasi. Mutasi dapat
18 terjadi pada level DNA akibat adanya perubahan basa-basa DNA (A = Adenin, T = Timin, G = Guanin, C = Citosin) dalam bentuk substitusi (transisi atau transversi), delesi (hilang), atau insersi dan inversi (Nei, 1987). Dilihat berdasarkan perbedaan situs pemotongan basa pada masing-masing alel (Gambar 3), diduga bahwa terjadi mutasi substitusi transisi. Substitusi transisi antara basa pirimidin, yaitu C (Cytosine) menjadi T (Tymine) merubah situs pemotongan enzim restriksi MspI (Yao et al., 1996).
Keragaman gen GH|MspI sapi diketahui terletak pada intron 3 pada posisi sekuen 1547 (Zhang et al., 1993). Daerah intron yang merupakan space internal antara pengkode protein pada sekuen gen, akan hilang (splicing) saat proses transkripsi, sehingga diduga pengaruh mutasi yang terjadi pada gen GH|MspI, yaitu
silent mutation. Silent mutation atau synonimous tidak terjadi pada situs aktif protein dan tidak menyebabkan perubahan asam amino karena beberapa asam amino yang sama dikodekan oleh kodon yang berbeda (Nei, 1987 ; Paolella, 1997).
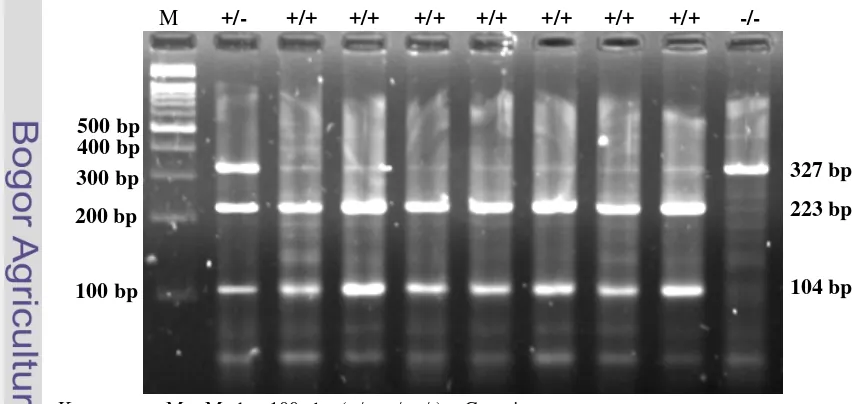
Hasil PCR-RFLP fragmen gen GH|MspI pada gel agarose 2% menunjukkan adanya pola pita beragam dengan tiga macam genotipe (Gambar 4), yaitu genotipe GH|MspI (+/+) yang terdiri dari dua pita (104 pb, 223 pb), genotipe GH|MspI (+/-) yang terdiri dari 3 pita (104 pb, 223 pb, 327 pb), dan genotipe GH|MspI (-/-) yang terdiri dari satu pita tidak terpotong (327 pb). Individu bergenotipe GH|MspI (+/+) dan GH|MspI (-/-) dikenal sebagai individu yang homozigot, sedangkan individu bergenotipe GH|MspI (+/-) dikenal sebagai individu yang heterozigot.
Keterangan : M = Marker 100 pb ; (+/+, +/-, -/-) = Genotipe
Gambar 4. Visualisasi PCR-RFLP Fragmen Gen GH|MspI
100 bp 200 bp 300 bp 400 bp 500 bp
M +/- +/+ +/+ +/+ +/+ +/+ +/+ +/+ -/-
327 bp
223 bp
19
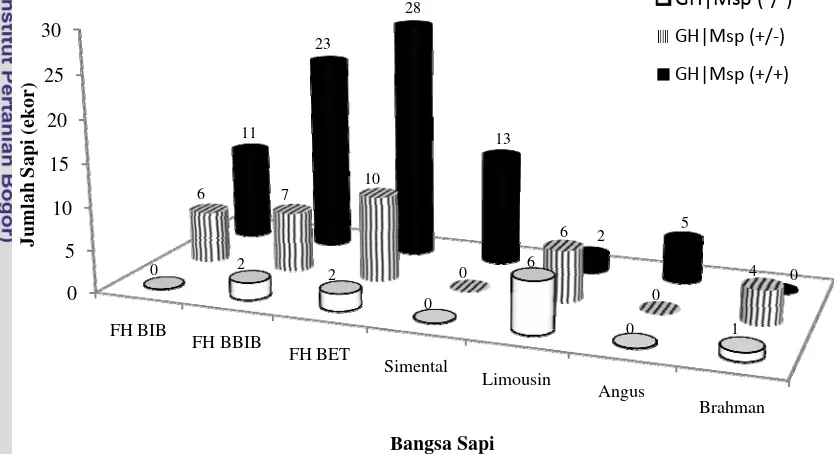
Berdasarkan hasil penelitian, ditemukan tiga macam genotipe, yaitu GH|MspI (+/+), GH|MspI (+/-), dan GH|MspI (-/-) pada sapi FH di BBIB Singosari dan BET Cipelang, sedangkan pada sapi FH di BBIB ditemukan dua macam genotipe, yaitu GH|MspI (+/+) dan GH|MspI (+/-). Hasil ini sebanding dengan penelitian Zhou et al. (2005) yang menunjukkan bahwa amplifikasi PCR-RFLP gen GH|MspI pada sapi Beijing Holstein menghasilkan tiga genotipe. Hasil penelitian untuk gen GH|MspI pada sapi pedaging di BET Cipelang, yaitu sapi Limousin juga ditemukan tiga genotipe. Pada sapi Brahman hanya ditemukan dua genotipe, yaitu GH|MspI (+/-) dan GH|MspI (-/-), sedangkan pada sapi Simental serta sapi Angus hanya ditemukan satu genotipe GH|MspI (+/+). Keragaman gen GH|MspI dapat terlihat jelas berdasarkan jumlah genotipe sapi yang diamati (Gambar 5).
Keterangan : Sapi FH = BIB Lembang (♂), BBIB Singosari (♂), dan BET Cipelang (♀); Sapi
Pedaging BET Cipelang (♀) = Simental, Limousin, Angus, dan Brahman
Gambar 5. Keragaman Gen GH|MspI pada Sapi FH dan Sapi Pedaging Frekuensi Genotipe dan Frekuensi Alel
20 Tabel 2. Frekuensi Genotipe dan Frekuensi Alel dari Gen GH|MspI pada Sapi FH
dan Sapi Pedaging
Keterangan : (...)* adalah jumlah sampel sapi
21 yang termasuk tipe perah, dapat dihubungkan dengan sifat kuantitatif seperti banyaknya produksi susu.
Perolehan hasil penelitian pada sapi FH ini menunjukkan hasil yang berbeda jika dibandingkan dengan sapi pedaging di BET Cipelang. Sapi Simental dan Angus memiliki genotipe GH|MspI (+/+) sangat tinggi dengan frekuensi genotipe sebesar 1,000. Frekuensi genotipe pada sapi Limousin dan Brahman menunjukkan hasil yang berbanding terbalik dengan frekuensi sapi FH, Simental, dan Angus. Sapi Limousin dan Brahman memiliki genotipe GH|MspI (+/+) sangat rendah dengan frekuensi genotipe masing-masing sebesar 0,144 dan 0,000. Sapi Limousin lebih banyak memiliki genotipe GH|MspI (+/-) dan GH|MspI (-/-) dengan frekuensi genotipe berimbang sebesar 0,428; sedangkan sapi Brahman memiliki genotipe GH|MspI (+/-) paling tinggi dengan frekuensi genotipe sebesar 0,800. Secara keseluruhan, bangsa sapi pedaging di BET memiliki frekuensi genotipe GH|MspI (+/+) yang lebih tinggi (0,541) dibandingkan dengan frekuensi genotipe GH|MspI (+/-) (0,270) dan GH|MspI (-/-) (0,189). Gen GH pada tipe pedaging memiliki peran yang berbeda dengan tipe perah. Tinggi atau rendahnya frekuensi genotipe GH|MspI pada tipe pedaging dapat dihubungkan dengan sifat pertumbuhan dan produksi karkas.
Genotipe dapat dihubungkan dengan sifat produksi susu maupun karkas. Menurut Zhou et al. (2005), sapi Beijing Holstein bergenotipe GH|MspI (+/+) menghasilkan produksi susu dan protein yang tinggi, dengan lemak yang lebih rendah. Hasil penelitian lain yang dilakukan pada sapi perah FH Polandia (Dybus, 2002) menunjukkan bahwa genotipe GH|MspI (+/+) memiliki produksi susu dan lemak susu yang lebih tinggi dibandingkan dengan genotipe GH|MspI (+/-) dan GH|MspI (-/-). Sapi Brangus bergenotipe GH|MspI (+/-) berpengaruh positif terhadap PBBH, karkas, dan kualitas daging (Thomas et al., 2006).
22 Sapi FH yang berasal dari BIB Lembang, BBIB Singosari, dan BET Cipelang memiliki alel GH|MspI (+) lebih tinggi dengan frekuensi alel masing-masing sebesar 0,824, 0,828, dan 0,825. Secara keseluruhan, bangsa sapi FH jantan maupun betina di tiga lokasi memiliki frekuensi alel GH|MspI (+) lebih tinggi terhadap alel GH|MspI (-) (0,826 vs 0,174). Hasil penelitian ini sebanding dengan pendapat Zhou et al. (2005) dimana frekuensi alel GH|MspI (+) pada sapi Beijing Holstein, yaitu 0,875. Dilihat berdasarkan besar frekuensi alelnya yang hampir berimbang di antara lokasi pengamatan, sapi FH dari tiga lokasi tersebut bersifat polimorfik. Hal ini sesuai dengan pendapat Nei (1987) yang menyatakan bahwa suatu alel dikatakan polimorfik atau beragam jika memiliki frekuensi alel sama dengan atau kurang dari 0,99, namun jika terjadi sebaliknya maka bersifat monomorfik atau seragam.
Terdapat perbedaan tipe dan frekuensi alel dari gen GH|MspI yang ditemukan antara sapi FH dan sapi pedaging. Sapi Limousin dan Brahman memiliki frekuensi alel GH|MspI (-) lebih tinggi terhadap alel GH|MspI (+) masing-masing sebesar 0,643 vs 0,357 untuk sapi Limousin; dan sebesar 0,600 vs 0,400 untuk sapi Brahman, sehingga kedua sapi tersebut bersifat polimorfik. Pada sapi Simental dan Angus, hanya satu tipe alel yang ditemukan, yaitu alel GH|MspI (+) dengan frekuensi sebesar 1,000, sehingga bersifat monomorfik. Hal ini dapat terjadi oleh adanya manajemen perkawinan yang tidak acak, seleksi terhadap sifat tertentu, dan tingkat silang dalam yang tinggi (Bourdon, 2000).
23 (Jakaria et al., 2009). Yao et al. (1996) berpendapat bahwa sapi Bos indicus, seperti sapi Brahman memiliki karakteristik yang lebih tinggi untuk alel GH|MspI (-).
Keseimbangan Hardy-Weinberg
Analisis Chi-Kuadrat dapat digunakan untuk mengetahui seimbang atau tidaknya frekuensi genotipe (p2, 2pq, q2) atau frekuensi alel (p dan q) pada suatu populasi ternak. Hasil analisis Chi-Kuadrat pada sapi FH dari tiga lokasi dan sapi pedaging dari BET Cipelang sebagai pembanding tertera pada Tabel 3.
Tabel 3. Keseimbangan Hardy-Weinberg (HW) Berdasarkan Uji χ2
Bangsa (ekor)* Lokasi χ2
Sapi Perah
FH ♂ (17) BIB Lembang Td
FH ♂ (32) BBIB Singosari 1,716tn
FH ♀ (40) BET Cipelang 0,720 tn
Sub Total (89) 0,919 tn
Sapi Pedaging
Simental ♀ (13) BET Cipelang Td
Limousin ♀ (14) BET Cipelang 0,062 tn
Angus ♀ (5) BET Cipelang Td
Brahman ♀ (5) BET Cipelang Td
Sub Total (37) 5,437 *
Keterangan : (...)* adalah jumlah sampel sapi; χ2 0,05(1) = 3,84; tn = tidak nyata; * = nyata ; td = tidak dapat dihitung
Analisis Chi-Kuadrat (χ2
) pada sapi FH dari BBIB Singosari dan BET Cipelang menunjukkan hasil yang tidak berbeda nyata (χ2 < χ2
(0,05)), sehingga dapat
24 nyata (χ2 > χ2(0,05)), sehingga berada dalam keadaan tidak seimbang. Nilai χ2 pada
sapi FH di BIB Lembang, Simental, Angus, dan Brahman tidak dapat dihitung. Hal ini dikarenakan frekuensi genotipe pada sapi pengamatan tersebut tidak memenuhi asumsi untuk dilakukan analisis keseimbangan Hardy-Weinberg.
Nilai Chi-Kuadrat (χ2) yang tidak berbeda nyata juga dapat dikatakan bahwa hasil perkawinan antar individu dari setiap bangsa tersebut berada pada keseimbangan. Menurut Noor (2008), suatu populasi yang cukup besar berada dalam keadaan keseimbangan Hardy-Weinberg jika frekuensi genotipe dominan dan resesif konstan dari generasi ke generasi, tidak ada seleksi, mutasi, migrasi, serta genetic drift. Seleksi merupakan salah satu faktor yang dapat mengubah keseimbangan dalam populasi secara cepat. Keseimbangan frekuensi genotipe gen GH|MspI pada sapi FH di BBIB Singosari dan BET Cipelang, serta sapi Limousin di BET Cipelang dapat menunjukkan bahwa tidak adanya seleksi secara langsung berdasarkan pada genotipe gen GH|MspI.
Heterozigositas
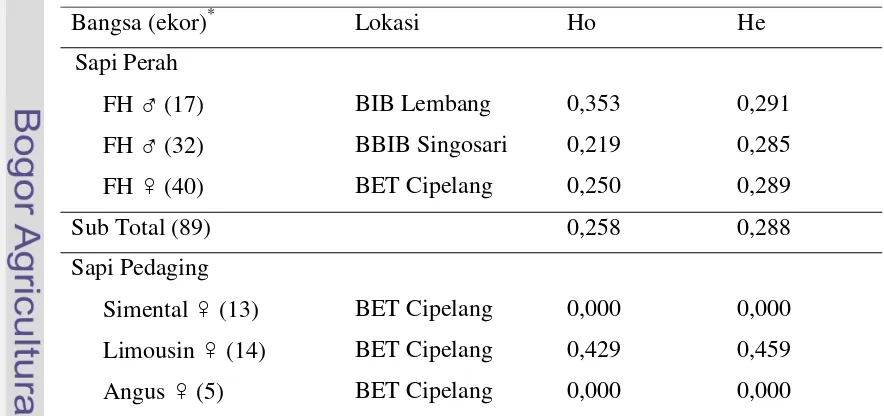
Nilai heterozigositas gen GH|MspI pada sampel sapi Friesian Holstein di tiga lokasi dan sapi pedaging di BET Cipelang diperoleh berdasarkan frekuensi alel. Hasil tersebut tertera pada Tabel 4.
Tabel 4. Nilai Heterozigositas Pengamatan (Ho) dan Nilai Heterozigositas Harapan (He) Gen GH|MspI pada Sapi FH dan Sapi Pedaging
Bangsa (ekor)* Lokasi Ho He
Sapi Perah
FH ♂ (17) BIB Lembang 0,353 0,291
FH ♂ (32) BBIB Singosari 0,219 0,285
FH ♀ (40) BET Cipelang 0,250 0,289
Sub Total (89) 0,258 0,288
Sapi Pedaging
Simental ♀ (13) BET Cipelang 0,000 0,000 Limousin ♀ (14) BET Cipelang 0,429 0,459
25 Brahman ♀ (5) BET Cipelang 0,800 0,480
Sub Total (37) 0,270 0,438
Keterangan : (...)* adalah jumlah sampel sapi
Pendugaan nilai heterozigositas diperoleh untuk mendapatkan keragaman genetik dalam populasi yang dapat digunakan untuk membantu program seleksi pada ternak yang akan digunakan sebagai sumber genetik pada generasi berikutnya (Marson et al., 2005). Berdasarkan hasil analisis, nilai heterozigositas pengamatan (Ho) pada seluruh sapi FH bernilai antara 0,219-0,353 dan pada sapi pedaging bernilai antara 0,000-0,429, kecuali pada sapi Brahman (0,800). Menurut Javanmard
et al. (2005), nilai heterozigositas di bawah 0,5 (50%) mengindikasikan rendahnya variasi suatu gen dalam populasi. Hal ini menunjukkan rendahnya tingkat keragaman gen GH|MspI pada sapi FH dari BIB Lembang, BBIB Singosari, dan BET Cipelang; serta sapi Simental, Limousin, dan Angus. Berbeda dengan sapi Brahman yang memiliki nilai Ho tertinggi, yaitu 0,800. Besarnya nilai Ho tersebut menunjukkan tingginya tingkat keragaman gen GH|MspI pada sapi Brahman. Keragaman yang tinggi dapat menunjukkan tingkat heterozigositas yang tinggi, sehingga dapat dilakukan seleksi.
Nilai heterozigositas harapan (He) diketahui untuk mengetahui perbedaan nilainya terhadap nilai heterozigositas pengamatan (Ho). Berdasarkan hasil analisis yang tertera pada Tabel 4, nilai Ho yang lebih tinggi dari He terdapat pada sapi FH di BIB Lembang, sehingga mengindikasikan bahwa tingkat heterozigositasnya tinggi. Oleh karena itu, maka seleksi dapat dilakukan agar diperoleh sifat pertumbuhan atau produksi yang seragam. Secara keseluruhan, nilai Ho dan He pada sapi FH dari tiga lokasi yang diamati tidak menunjukkan adanya perbedaan nilai yang besar, yaitu dengan rataan nilai Ho = 0,258 dan He = 0,288. Hasil yang sama juga ditunjukkan pada sapi pedaging, kecuali pada sapi Bahman yang memiliki nilai Ho lebih tinggi dari He (Ho = 0,800 dan He = 0,480). Menurut Machado et al. (2003), jika nilai Ho lebih rendah dari He maka dapat mengindikasikan adanya proses seleksi yang intensif. Hal ini menunjukkan bahwa pada seluruh sapi FH maupun sapi pedaging (kecuali sapi Brahman) diperkirakan tidak terjadi proses seleksi intensif yang secara langsung berdasarkan gen GH|MspI.
KESIMPULAN DAN SARAN Kesimpulan
Gen GH|MspI pada sapi FH di BIB Lembang, BBIB Singosari, dan BET Cipelang bersifat polimorfik dengan dua alel, yaitu alel GH|MspI (+) dan (-); serta tiga genotipe, yaitu GH|MspI (+/+), (+/-), dan (-/-). Frekuensi alel GH|MspI (+) (0,826) lebih tinggi dari alel GH|MspI (-) (0,174). Frekuensi genotipe GH|MspI (+/+) ditemukan paling tinggi pada sapi FH di tiga lokasi. Sapi FH berada dalam keseimbangan Hardy-Weinberg dan memiliki tingkat keragaman gen GH|MspI yang rendah (0,219-0,353).
Sapi pedaging di BET Cipelang memiliki genotipe yang bervariasi. Gen GH|MspI pada sapi Limousin dan Brahman bersifat polimorfik, tetapi pada sapi Simental dan Angus bersifat monomorfik dengan frekuensi alel GH|MspI (+/+) sama dengan 1,000. Sapi pedaging tidak berada dalam keseimbangan Hardy-Weinberg dan memiliki tingkat keragaman gen GH|MspI yang rendah (0,000-0,429), kecuali pada sapi Brahman (0,800).
Saran
Jumlah sampel yang lebih banyak dengan populasi dari balai-balai penelitian, industri, dan peternakan rakyat pada berbagai manajemen yang berbeda-beda akan memberikan gambaran tentang keragaman genetik yang lebih akurat.
UCAPAN TERIMA KASIH
Puji dan syukur senantiasa penulis panjatkan kepada Allah SWT atas segala rahmat dan karunia-Nya sehingga penulis dapat menyelesaikan penulisan skripsi ini. Shalawat serta salam senantiasa dicurahkan kepada Nabi Muhammad SAW. Penghargaan dan rasa syukur yang sangat tinggi penulis sampaikan kepada orang tua tercinta, Eman Sulaeman, S.Pd, M.M. dan Nia Kania MS, SE, M.MPd atas segala doa, kasih sayang, dukungan, motivasi serta bimbingannya bagi penulis; kepada adik tersayang, Naufal Rifki Kusuma, yang selalu menghibur, menyemangati, dan memberikan warna dalam kehidupan penulis; serta kepada keluarga besar Bapak dan Ibu penulis atas doanya. Semoga penulis dapat selalu memberikan kebahagian kepada keluarga tercinta.
Penulis mengucapkan banyak terima kasih kepada Prof. Dr. Ir. Cece Sumantri, M.Agr.Sc. selaku pembimbing utama dan Ir. Anneke Anggraeni, M.Si., Ph.D. selaku pembimbing anggota atas segala arahan, bimbingan, motivasi, dan ilmu pengetahuan yang bermanfaat bagi penulis; serta kepada Dr. Ir. Asnath M. Fuah. selaku dosen pembimbing akademik atas bimbingan dan arahannya selama studi. Terima kasih juga kepada Ir. Sri Darwati, M.Si., Dr. Ir. Didid Diapari, M.Si., dan Dr. Rudi Afnan, S.Pt., M.Sc.Agr., selaku dosen penguji sidang atas kritik dan sarannya yang bermanfaat.
Ucapan terima kasih penulis sampaikan kepada K’ Eryk, K’ Restu, Bpk. Ihsan, Bpk. Andi, Ibu Suri, K’ Surya, dan K’ Sutikno atas masukan dan bimbingannya. Ucapan terima kasih juga penulis sampaikan kepada Ahmad Isnaeni yang selalu memberi perhatian dan semangat penulis, kepada Annisa O.R. yang selalu menemani selama studi dan penelitian; Gilang A., Anis, Maya R., Khairul B., dan Alfian A.C. Umam yang selalu memberi warna dalam kehidupan kampus IPB tercinta; serta kepada sahabat-sahabat di Laboratorium GMT (Wike, Tiffany, Gabby, Iren, Priskilla, Ulin, Ferdy, Revy, Leni, dan lainnya), Maya G., Basriansyah, sahabat-sahabat dari IPTP 44, SMA, dan asrama, terima kasih atas doa dan kebersamaannya. Semoga skripsi ini bermanfaat untuk penulis dan dunia peternakan Indonesia. Amin.
Bogor, 11 April 2011
DAFTAR PUSTAKA
Akers, R. M. 2006. Major advances associated with hormone and growth factor regulation of mammary growth and lactation in dairy cows. J. Dairy Sci. 89 : 1222–1234.
Avise, J. C. 1994. Molekular Markers. Natural History and Evolution. Chapman and Hall, Inc., New York.
Beauchemin, V.R., Thomas, M.G., Franke, D.E., & Silver, G.A. 2006. Evaluation of DNA polymorphism involving growth hormone relative to growth and carcass characteristics in Brahman steers. Genet. Mol. Res.5 : 438-447.
Blakely, J. & D. H. Bade. 1998. Ilmu Peternakan. Terjemahan : B. Srigandono. Gadjah Mada University Press, Yogjakarta.
Bourdon, R. M. 2000. Understanding Animal Breeding. 2nd ed. Prentice Hall. Inc. Upper Saddle River, New Jersey.
Brown, T. A. 1999. Genome. Garland Science Publishing, New York.
Buckle, K.A., R.A. Edwards, G.H. Fleet, & M. Wooton. 2007. Ilmu Pangan. Terjemahan : H. Purnomo dan Adiono. Universitas Indonesia Press, Jakarta. Chandra, S., B. Sutrasno, T. H. Siwi, A. Kumalajati, H. Supriadi, A. Marsudi, &
Budiningsih. 2009. Pemeliharaan Bibit Sapi Perah. BPPTU Sapi Perah Baturaden, Purwokerto.
Cowan, C. M., M. R. Dentine, R. L. Ax, & L. A. Schuler. 1989. Restriction fragment length polymorphism associated with growth hormone and prolactin genes in Holstein bulls : evidence for a novel growth hormone allele. J. Anim. Genet. 20 : 157-165.
Direktorat Jenderal Peternakan. 2008. Statistik Peternakan 2008. Direktorat Jenderal Peternakan, Departemen Pertanian, Jakarta.
Direktorat Jenderal Peternakan. 2009. Statistik Peternakan 2009. Direktorat Jenderal Peternakan, Departemen Pertanian, Jakarta.
Dudi, D. Rahmat, & T. Dhalika. 2006. Evaluasi potensi genetik sapi perah Fries Holland (FH) di Koperasi Serba Usaha (KSU) Tandangsari Kabupaten Sumedang. J. Ilmu Ternak. 6 : 1-11.
Dybus, A. 2002. Association of growth hormone (GH) and prolactin (PRL) genes polymorphism with milk production traits in Polish Black-and-White cattle. Anim. Sci. Papers and Rep. 20 : 203-212.
Ensminger, M. E. & H. D. Tyler. 2006. Dairy Cattle Science. 4th ed. The Interstate Printers and Publisher Inc., Danville, Illinois.
French, M. H. 1996. European Breeds of Cattle Food and Agriculture. Rome, United Nations.
29 Ge, W., Davis, M. E., Hines H. C., Irvin, K. M., & R. C. M. Simmen. 2003.
Association of single nucleotide polymorphism in the growth hormone and growth hormone receptor genes with blood serum insulin-like growth factor I concentration and growth traits in Angus cattle. J. Anim. Sci. 81: 641-648. Gorbani, A., R.V. Torshizi, M. Bonyadi, & C. Amirinia. 2009. Restriction fragment
length polymorphism of bovine growth hormone gene intron 3 and its association with testis biometry traits in Iranian Holstein bull. African J. of Microbiol. Res. 3 : 809-814.
Gordon, D. F., D. P. Quick, C. R. Ewin, J. E. Donelson, & R. A. Maurer. 1983. Nukleotide sequence of the bovine growth hormone chromosomal gene. Mol. Cell. Endocrinol. 33 : 81-95.
Green, E. K. 1998. Restriction fragment length polymorphism. In : Rapley, R and J. M. Walker. Eds. Molekular Biology Methods Handbook. Human Press. Totowa, New Jersey.
Hartl, D. L. & A. G. Clark. 1997. Principle of Population Genetic. Sinaver Associates, Sunderland, MA.
Hediger, R., S. E. Johnson, W. Barendse, R. D. Drinkwater, S. S. Moore, & J. Hetzel. 1990. Assignment of the growth hormone gene locus to 19q26-qter in cattle and to 11q25-qter in sheep by in situ hybridization. J. Genom. 8 : 171-174. Jakaria, R.R. Noor, H. Martojo, D. Duryadi, & B. Tappa. 2009. Identification of
growth hormone (GH) gene MspI and AluI loci polymorphism in beef cattles. J. Anim. Prod. 1 : 42-47.
Javanmard, A., N. Asadzadeh, M. H. Banabazi, & J. Tavakolian. 2005. The allele and genotype frequencies of bovine pituitary specific transcription factor and leptin genes in Iranian cattle and buffalo populations using PCR-RFLP. Iranian J. of Biotechnol. 2 : 104-108.
Khatami, S. R., E. Lazebnvi, V. F. Maksimenko, & G.E. Sulimova. 2005. Association of polymorphism of the growth hormone and prolactin genes with milk productivity in YarosIavI and black-and-white cattle. J. Genet.
41: 229-236.
Lagziel, A., S. Denise, O. Hanotte, S. Dhara, V. Glazko, A. Broadhead, R. Davoli, V. Russo, & M. Soller. 2000. Geographic and breed distribution of an MspI PCR-RFLP in the bovine growth homone (bGH) gene. J. Anim. Genet. 31: 210–213.
Lawrence, T. L. J. & Fowler, V. R. 2002. Growth of Farm Animals. 2nd ed. CABI Publishing, New York.
Li, X., K. Li, B. Fan, Y. Gong, S. Zhao, Z. Peng, & B. Liu. 2000. The genetic diversity of seven pigs breeds in China, estimated by means of micosatellites. J. Anim. Sci. 9 : 1193-1195.
30 Marson, E. P., J. B. S. Ferraz, F. V. Meirelles, J. C. C. Balieiro, J. P. Eler, L. G. G. Figuerido, & G. B. Mourao. 2005. Genetik characterization of European-Zebu composite bovine using RFLP markers. J. Genet. Mol. Res. 4 : 496-505. Mitra, A., P. Sciilee, C. R. Balakrisiinan, & F. Pirciiner. 1995. Polymorphisms at
growth hormone and prolactine loci in Indian cattle and buffalo. J. Anim. Breed. and Genet. 112 : 71-74.
Montaldo, H. H. & C. A. M. Herrera. 1998. Use of molecular markers and major genes in the genetic improvement of livestock. J. Biotechnol. 1 : 2.
Muladno. 2002. Teknologi Rekayasa Genetik. Pustaka Wira Usaha Muda, Bogor. Nei, M. 1987. Molecular Evolutionary Genetics. Columbia University Press, New
York.
Nei, M. & S. Kumar. 2000. Molecular Evolution and Phylogenetics. Oxford University Press, New York.
Noor, R. R. 2008. Genetika Ternak. Penebar Swadaya, Jakarta.
Paolella, P. 1997. Introduction to Molecular Biology. Mc Graw Hill Companies, Inc., Boston, Massachusetts.
Parakkasi, A. 1999. Ilmu Nutrisi dan Makanan Ternak Ruminansia. Universitas Indonesia Press, Jakarta.
Phillips, C. J. 2001. Principles of Cattle Production. CABI Publishing, New York. Pierzchala, M., B. Tadeusz, & K. Jolanta. 2004. Growth rate and carcass quality in
relation to GH|MspI and GH|HaeI PCR-RFLP polymorphism in pigs. J. Anim. Sci. Papers and Rep. 22 : 57-64.
Rastegari, A., H. Roshanfekr, M. Mamouie, & S. R. Khatami. 2010. Growth hormone genotyping of Nadji cattle breed using PCR-RFLP. J. Anim. and Vet. Adv. 9 : 1265-1266.
Reis, C., D. Navas, N. Pereira, & A. Cravador. 2001. Growth hormone AluI polymorphism analysis in eight portuguese bovine breeds. Arch. Zootec.
50 : 41-48.
Sambrook, J., E. F. Fritsch, & J. F. Medrano. 1989. Molecular Cloning : A Laboratory Manual. 2nd ed. Cold Spring Harbor Laboratory Press, New York. Schmidt, G. H. & L. D. V. Vleck. 1974. Principles of Dairy Science. W. H. Freeman
and Company, San Fransisco.
Sumantri, C. A., A. Anggraeni, R. R. A. Maheswari, K. Dwiyanto, A. Fajarallah, & B. Brahmantiyo. 2004. Frekuensi gen kappa-kasein (κ-kasein) pada sapi perah FH berdasarkan produksi susu di BPTU Baturaden. Prosiding. Seminar Nasional Teknologi Peternakan dan Veteriner 2004.
31 Tambasco, D. D., C. C. P. Paz, M. Tambasco-Studart, A. P. Pereira, M. M. Alencar, A. R. Freitas, L. L. Coutinho, I. U. Packer, & L. C. A. Regitano. 2003. Candidate genes for growth traits in beef cattle crosses Bos taurus x Bos indicus. J. Anim. Breed. Genet. 120 : 51-56.
Thomas, M. G., G. A. Silver, & R. M. Enns. 2006. Relationships of DNA polymorphism in growth hormone (GH) to growth and carcass traits observed in a population of Brangus bulls with larger number of sires. Int. Plant and Animal Genome. XIV: P526. (Abstr.).
Tomaszewska, M. W., I. K. Sutama, I. G. Putu, & T. D. Chaniago. 1991. Reproduksi, Tingkah Laku, dan Produksi Ternak di Indonesia. PT. Gramedia Pustaka Utama, Jakarta.
Unanian, M. M., C. C. Borreto, A. R. De Freitas, & C. M. T. Cordeiro. 2000. Association between GH gene polymorphism and weigth traits in Nellore Novines. Rev. Bras Zootec. 29 : 1380-1386.
Vasconcellos L. P. M. K., D. T. Talhari, A. P. Pereira, L. L. Coutinho, & L. C. A. Regitano. 2003. Genetic characterization of Aberdeen Angus cattle using molecular markers. Genet. Mol. Biol. 26: 133-137.
Viljoen, G. J., L. H. Nel, & J. R. Crowther. 2005. Molecular Diagnostic PCR Handbook. Springer, Dordrecht, Netherland.
Wallis, M. 1973. The primary structure of bovine growth hormone. FEBS Lett. 35 : 11-14.
Weir, B. S. 1996. Genetic Data Analysis : Method for Discrete Population Genetic Data. 2nd ed. Sinauer Associates. Sunderland, MA.
Williamson, G. & W. J. A. Payne. 1993. An Introduction of Animal Husbandary in Tropics. Longman, London.
Yao, J., S. E. Aggerey, D. Zadworny, J. E. Hayes, & U. Kuhnlein. 1996. Sequence variations in the bovine growth hormone gene characterized by single strand conformation polymorphism (SSCP) analysis and their association with milk production traits in Holstein. J. Genet.144 : 1809–1816.
Yardibi, H., T. H. Gulhan, I. Paya, F. Kaygisiz, G. Ciftioglu, A. Mengi, & K. Oztabak. 2009. Association of growth hormone gene polymorphisms with milk production traits in South Anatolian and East Anatolian Red cattle. J. Anim. and Vet. Adv. 5 : 1040-1044.
Zakizadeh, S., G. Rahimi, S. R. Mirae-Ashtiani, A. Nejati-Javaremi, M. Moradi-Shahrbabak, P. Reinecke, M. Reissmann, A.A. Masoudi, C. Amirinia, & S.A. Mirhadi. 2006. Analysis of bovine growth hormone gene polymorphisms in three iranian native breeds and holstein cattle by RFLP-PCR. J. Biotechnol. 5 : 385-390.
32 Zhou, G. L., H. G. Jin, S. L. Guo, Q. Zhu & Y. H. Whu. 2005. Association of genetic polymorphism in GH gene with milk production traits in Beijing Holstein cows. J. Biosci. 30 : 595-598.
34 Lampiran 1. Sekuen Gen GH Sapi dari GenBank dengan Kode Akses. M57764
Sumber : Gordon et al. (1983)
LOCUS BOVGHGH 2856 bp DNA linear MAM 27-APR-1993
DEFINITION Bovine growth hormone gene, complete cds.
ACCESSION M57764 M28453
VERSION M57764.1 GI:163091
KEYWORDS growth hormone.
SOURCE Bos taurus (cattle)
ORGANISM Bos taurus </Taxonomy/Browser/wwwtax.cgi?id=9913> Eukaryota; Metazoa; Chordata; Craniata; Vertebrata; Euteleostomi; Mammalia; Eutheria; Laurasiatheria;
Cetartiodactyla; Ruminantia; Pecora; Bovidae; Bovinae; Bos. REFERENCE 1 (bases 1 to 2856)
AUTHORS Gordon,D.F., Quick,D.P., Erwin,C.R., Donelson,J.E. and Maurer,R.A.
TITLE Nucleotide sequence of the bovine growth hormone chromosomal gene
mRNA join(649..723,971..1131,1359..1475,1703..1864,2138..2439)
/gene="GH1"
/product="growth hormone"
exon 649..723
/gene="GH1" /number=1
CDS join(711..723,971..1131,1359..1475,1703..1864,2138..2338)
/gene="GH1" /codon_start=1
/product="growth hormone"
/protein_id="AAA30544.1 </protein/163092>" /db_xref="GI:163092"
35
1 gtactggggt gggttgcctt tctcttctcc aggggattta tctgacccag ggattgaacc 61 tgagtctcct gcatttgcag ctagattctt tacggctgag ccacctggga agcccattcg 121 cttctgctgc tgctgctgct gctaagttgc ttcagtcgtg tccgacctgt gcgacgccat 181 agacagcagc ccaccaggtc cccgtccctg ggattctcca ggcaagaaca ttggagtggg 241 ttgccatttc ctcctccaat gcatgaaagt gaaaagtgaa agtgaagtca ctcagttgtg 301 tccgaccctc agcgacccca tggactgcag ccttccagaa tggggtgcca ttgccttctc 361 ctcgcttctg ctacctcccc tttaaaaaga aaacctatgg ggtgggctct caagctgaga 421 ccctgtgtgc acagccctct ggctggtggc agtggagacg ggatgatgac aagcctgggg 481 gacatgaccc cagagaagga acgggaacag gatgagtgag aggaggttct aaattatcca 541 ttagcacagg ctgccagtgg tccttgcata aatgtataga gcacacaggt ggggggaaag 601 ggagagagag aagaagccag ggtataaaaa tggcccagca gggaccaatt ccaggatccc 661 aggacccagt tcaccagacg actcagggtc ctgtggacag ctcaccagct atgatggctg 721 caggtaagct cgctaaaatc ccctccattc gcgtgtccta aaggggtaat gcggggggcc 781 ctgccgatgg atgtgttcag agctttgggc tttagggctt ccgaatgtga acataggtat 841 ctacacccag acatttggcc aagtttgaaa tgttctcagt ccctggaggg aagggtaggt 901 ggggctggca ggagatcagg cgtctagctc cctggggccc tccgtcgcgg ccctcctggt 961 ctctccctag gcccccggac ctccctgctc ctggctttcg ccctgctctg cctgccctgg 1021 actcaggtgg tgggcgcctt cccagccatg tccttgtccg gcctgtttgc caacgctgtg 1081 ctccgggctc agcacctgca tcagctggct gctgacacct tcaaagagtt tgtaagctcc 1141 cgagggatgc gtcctagggg tggggaggca ggaaggggtg aatccacacc ccctccacac 1201 agtgggagga aactgaggag ttcagccgta ttttatccaa gtagggatgt ggttagggga 1261 gcagaaacgg gggtgtgtgg ggtggggagg gttccgaata aggcggggag gggaaccgcg 1321 caccagctta gacctgggtg ggtgtgttct tcccccagga gcgcacctac atcccggagg 1381 gacagagata ctccatccag aacacccagg ttgccttctg cttctctgaa accatcccgg 1441 cccccacggg caagaatgag gcccagcaga aatcagtgag tggcaacctc ggaccgagga 1501 gcaggggacc tccttcatcc taagtaggct gccccagctc ccgcaccggc ctggggcggc 1561 cttctccccg aggtggcgga ggttgttgga tggcagtgga ggatgatggt gggcggtggt 1621 ggcaggaggt cctcgggcag aggccgacct tgcagggctg ccccagaccc gcggcaccca 1681 ccgaccaccc acctgccagc aggacttgga gctgcttcgc atctcactgc tcctcatcca 1741 gtcgtggctt gggcccctgc agttcctcag cagagtcttc accaacagct tggtgtttgg 1801 cacctcggac cgtgtctatg agaagctgaa ggacctggag gaaggcatcc tggccctgat 1861 gcgggtgggg atggcgttgt gggtcccttc catgtggggg ccatgcccgc cctctcctgg 1921 cttagccagg agaatgcacg tgggcttggg gagacagatc cctgctctct ccctctttct 1981 agcagtccag ccttgaccca ggggaaacct tttccccttt tgaaacctcc ttcctcgccc 2041 ttctccaagc ctgtagggga gggtggaaaa tggagcgggc aggagggagc tgctcctgag 2101 ggcccttcgg cctctctgtc tctccctccc ttggcaggag ctggaagatg gcaccccccg 2161 ggctgggcag atcctcaagc agacctatga caaatttgac acaaacatgc gcagtgacga 2221 cgcgctgctc aagaactacg gtctgctctc ctgcttccgg aaggacctgc ataagacgga 2281 gacgtacctg agggtcatga agtgccgccg cttcggggag gccagctgtg ccttctagtt 2341 gccagccatc tgttgtttgc ccctcccccg tgccttcctt gaccctggaa ggtgccactc 2401 ccactgtcct ttcctaataa aatgaggaaa ttgcatcgca ttgtctgagt aggtgtcatt 2461 ctattctggg gggtggggtg gggcaggaca gcaaggggga ggattgggaa gacaatagca 2521 ggcatgctgg ggatgcggtg ggctctatgg gtacccaggt gctgaagaat tgacccggtt 2581 cctcctgggc cagaaagaag caggcacatc cccttctctg tgacacaccc tgtccacgcc 2641 cctggttctt agttccagcc ccactcatag gacactcata gctcaggagg gctccgcctt 2701 caatcccacc cgctaaagta cttggagcgg tctctccctc cctcatcagc ccaccaaacc 2761 aaacctagcc tccaagagtg ggaagaaatt aaagcaagat aggctattaa gtgcagaggg
IDENTIFIKASI KERAGAMAN GEN HORMON PERTUMBUHAN
(GH|
Msp
I) PADA SAPI FRIESIAN HOLSTEIN DI BIB LEMBANG,
BBIB SINGOSARI, DAN BET CIPELANG
SKRIPSI
DINY WIDYANINGRUM
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN