I. PENDAHULUAN
1.1. Latar Belakang
Taman Nasional Tesso Nilo (TNTN) merupakan taman nasional yang terletak di Propinsi Riau, Indonesia. Penetapan kawasan TNTN ini tertuang dalam Surat Keputusan Menteri Kehutanan Nomor: SK.255/Menhut-II/2004 tanggal 19 Juli 2004 mengenai perubahan fungsi sebagian kawasan Hutan Produksi Terbatas di Kelompok Hutan Tesso Nilo yang terletak di Kabupaten Pelalawan dan Kabupaten Indragiri Hulu Provinsi Riau. Luas TNTN ± 38.576 hektar. Kawasan yang masuk wilayah taman nasional ini adalah kawasan bekas Hak Pengusahaan Hutan (HPH) yang terletak di Kabupaten Pelalawan dan Indragiri Hulu. Hingga kini di sekelilingnya masih terdapat kawasan HPH.
Hampir seluruh jenis satwaliar dilindungi dapat ditemukan pada daerah pada areal kawasan TNTN. Taman Nasional Tesso Nilo memiliki berbagai potensi flora dan fauna yang beragam. Terdapat 360 jenis flora yang tergolong dalam 165 marga dan 57 suku, 107 jenis burung, 23 jenis mamalia, tiga jenis primata, 50 jenis ikan, 15 jenis reptilia dan 18 jenis amfibia (Dephut 2004).
Letak dari TNTN ini dikelilingi oleh vegetasi hutan alam, hutan akasia, pemukiman dan perkebunan kelapa sawit. Berbagai macam kegiatan yang dilakukan manusia yang berdekatan dengan TNTN mengakibatkan terjadinya perubahan pada komponen ekosistem baik komponen biotik maupun komponen abiotik. Salah satunya adalah perkebunan kelapa sawit. Adanya kegiatan perkebunan dapat memberikan pengaruh terhadap keberadaan satwaliar di kawasan tersebut. Dengan berkembangannya perkebunan kelapa sawit berdampak besar terhadap rusak dan hilangnya habitat satwaliar. Pembukaan lahan untuk perkebunan kelapa sawit menyebabkan terganggunya berbagai macam spesies yang ada di dalamnya. Namun, perkebunan kelapa sawit juga berperan bagi kelangsungan hidup spesies lainnya.
terdapat di kawasan kebun kelapa sawit belum dikelola dengan baik karena kurangnya informasi dan data mengenai potensi keanekaragaman yang terkandung di dalamnya, salah satunya mengenai mamalia.
Oleh karena itu, telah dilakukan penelitian mengenai studi keanekaragaman jenis mamalia, khususnya mamalia besar yang dapat menggambarkan keberadaan spesies tersebut dan penyebarannya pada areal kawasan TNTN yang berbatasan dengan kebun kelapa sawit PT. Inti Indosawit Subur Ukui, Kabupaten Pelalawan, Propinsi Riau.
1.2. Tujuan Penelitian
Penelitian mengenai studi keanekaragaman jenis mamalia besar pada areal TNTN yang berbatasan dengan kebun kelapa sawit PT. Inti Indosawit Subur Ukui, Kabupaten Pelalawan, Propinsi Riau ini bertujuan untuk:.
1). Mengetahui keanekaragaman jenis mamalia besar di areal kawasan TNTN yang berbatasan kebun kelapa sawit PT. Inti Indosawit Subur Ukui, Kabupaten Pelalawan, Propinsi Riau.
2). Mengetahui penyebaran jenis mamalia besar di areal kawasan TNTN yang berbatasan kebun kelapa sawit PT. Inti Indosawit Subur Ukui, Kabupaten Pelalawan, Propinsi Riau.
1.3. Manfaat
II. TINJAUAN PUSTAKA
2.1. Keanekaragaman Jenis
Keanekaragaman hayati atau biological diversity (biodiversity) adalah seluruh keanekaan bentuk kehidupan di muka bumi ini beserta interaksinya (BAPPENAS 2003). Keanekaragaman hayati memliki dua komponen utama, yaitu kekayaan jenis yang merupakan jumlah jenis dari suatu areal dan kemerataan jenis yang merupakan kelimpahan relatif suatu individu pada setiap spesies (Feldhamer et al. 1999). Kedua komponen tersebut memiliki nilai perhitungan yang dikenal dengan indeks kekayaan jenis dan indeks kemerataan jenis. Ledwig dan Reynold (1988) menyatakan bahwa indeks tersebut digabungkan menjadi satu nilai yang sama dengan indeks keanekaragaman. BAPPENAS (2003) menyatakan ada tiga tingkatan yang terkait dengan keanekaragaman hayati, yaitu:
1) Keanekargaman ekosistem: keanekaan bentuk dan susunan bentang alam daratan maupun perairan, dimana makhluk atau organisme hidup berinteraksi dan membentuk keterkaitan dalam lingkungan fisiknya.
2) Keanekaragaman jenis: keanekaan jenis organisme yang menempati suatu ekosistem, di darat maupun di perairan.
3) Keanekaragaman genetik: keanekaan individu di dalam suatu jenis yang disebabkan oleh perbedaan genetik antara individu.
Keanekaragaman merupakan hal yang paling penting dalam mempelajari suatu komunitas. Keanekaragaman jenis merupakan pertanyaan yang paling mendasar dalam ekologi, baik teori maupun terapan sehingga ahli ekologi harus mengetahui cara mengukur keanekaragaman jenis dan memahami hasil pengukurannya (Odum 1971).
berdasarkan pada kelimpahan relatif suatu jenis dan tingkat dominansi (Odum 1971; Krebs 1992; Magguran 1988).
2.2. Bio-Ekologi Mamalia
Mamalia merupakan kelompok yang memiliki kelas tertinggi dalam dunia hewan. Pada umumnya mamalia dibagi ke dalam dua kelompok besar, yaitu mamalia kecil dan mamalia besar. Mamalia kecil adalah mamalia yang memiliki berat badan berkisar antara 2 g - 5 kg. Jenis-jenis mamalia kecil antara lain adalah kelelawar (Chiroptera), bajing dan tikus (Rodentia), tupai (Scadentia). Sedangkan mamalia besar adalah mamalia yang berat badannya diatas 5 kg (Bouliere 1975).
Taksonomi mamalia menurut Jasin (1992) adalah sebagai berikut: Kingdom : Animalia
Sub Kingdom : Metazoa Filum : Chordata Sub Filum : Craniata Klas : Mamalia
Selanjutnya Jasin (1992) menyebutkan bahwa kelas mamalia sendiri menjadi tiga sub kelas, yaitu:
1) Sub kelas Prototheiria, yang hanya terdiri dari satu ordo, yaitu monotremata 2) Sub kelas Allotheria (sudah punah)
3) Sub kelas Theria, yang terdiri dari 27 ordo, yaitu Marsupialia, Insektivora, Dermopyera, Chiroptera, Primata, Tillodontia (sudah punah), Taediodota (sudah punah), Edentata (Xanathra), Pholidota, Lagomorpha, Rodentia, Cetacea, Carnivora, Condylartha, Litopterma, Notoungulata, Astrapotheria, Tubulidentata, Pantodonta, Dinocerata, Pyirotheria, Proboscidea, Embrithopoda, Hyracoida, Sirenia, Prissodactyla, dan Artiodactyla.
Rodentia, Proboscidea, Prissondactyla, dan Artiodactyla. Ciri-ciri khusus mamalia sebagai berikut (Medway 1978):
1. Tubuh biasanya diliputi rambut yang lepas secara periodik, kulit banyak mengandung kelenjar keringat dan kelenjar susu.
2. Ekor umumnya panjang dan dapat digerak-gerakkan.
3. Memiliki empat anggota kaki (kecuali Anjing laut dan Singa laut tidak memiliki kaki belakang), masing-masing kaki memiliki kurang lebih lima jari yang bermacam-macam bentuknya yang disesuaikan dengan keperluan. 4. Penapasan hanya dengan paru-paru, hasil ekskresi berupa urine.
5. Suhu tubuh tetap.
6. Hewan jantan mempunyai alat kopulasi berupa penis, fertilasi terjadi didalam tubuh betina.
2.3. Penyebaran Mamalia Besar
Ada banyak untuk membedakan penyebaran fauna di Indonesia. Lekagul dan McNeely (1977) menyatakan bahwa sistem yang mendapat tanggapan luas adalah berdasarkan Sclater 1958 dan Wallace 1987, yang membagi dunia ke dalam 6 wilayah geografis fauna, yaitu: Paleartik, Oriental, Australia, Neartik, Neotropik, dan Ethopia.
Penyebaran fauna dapat diikuti dari pola sejarah geologi, sehingga dapat dikenal adanya pola penyebaran fauna khas. Akan tetapi untuk beberapa hal ada jenis tertentu yang mempunyai penyebaran luas, terutama bagi jenis-jenis burung tertentu ataupun bagi jenis organisme yang pola penyebarannya melalui pola air. Bahkan manusia mempunyai peranan penting dalam penyebaran satwaliar sejak 10.000 tahun yang lalu (Alikodra 1990).
mencairnya es, jenis-jenis beradaptasi dan berevolusi pada kondisi yang baru dan berlainan, yang kadang menghasilkan subjenis baru atau bahkan jenis baru. Semakin lama isolasi yang terjadi, semakin banyak fauna-fauna yang berbeda, seperti yang ditunjukan oleh jumlah satwa yang endemik di Sulawesi dengan lebih dari 70% jenis mamalia darat yang endemik (Zon 1979).
Penyebaran mamalia mempunyai kecendrungan untuk dibatasi oleh penghalang-penghalang fisik seperti sungai, samudera, dan gunung, serta oleh penghalang ekologis seperti batas tipe hutan dan adanya jenis saingan yang telah menyesuaikan secara optimal dengan habitatnya sekarang. Sehingga penghalang-penghalang fisik itu dapat digunakan untuk menarik batas geografis fauna sepanjang batas fisik atau ekologis (Alikodra 1990).
Fauna Sumatera sangat erat hubunganya dengan fauna yang berada di Semenanjung Malaysia dengan relatif sedikit mamalia endemik, misalnya Kelinci sumatera (Nesolagus netsheri). Sesuai dengan kondisi biogeografisnya. Pulau Kalimantan (mamalia endemik sebanyak 18 jenis) memiliki jenis-jenis satwaliar endemik yanng lebih tinggi dibandingkan Pulau Sumatera (mamalia endemik sebanyak 10 jenis) (Whitten et al. 1987 dalam Alikodra 2002).
2.4. Pergerakan dan Daerah Jelajah Satwaliar
Pergerakan adalah suatu strategi dari individu ataupun populasi untuk menyesuaikan dan memanfaatkan keaadaan lingkungannya agar dapat hidup dan berkembangbiak secara normal. Pergerakan satwaliar merupakan suatu perilaku, sehingga mempunyai pola-pola tertentu sesuai dengan jenisnya. Pergerakan ini erat hubungannya dengan sifat individu dan kondisi lingkungannya seperti ketersediaan makanan, fasilitas untuk berkembangbiak, pemangsaan, kondisi cuaca, sumber air maupun adanya pengerusakan lingkungan (Alikodra 2002).
Daerah jelajah yaitu wilayah yang dikunjungi satwaliar secara tetap karena dapat mensuplai makanan, minum, serta mempunyai fungsi sebagai tempat berlindung/bersembunyi, tempat tidur dan tempat kawin (Boeghey 1973; Pyke 1983; Van Noordwijk 1985 dalam Entebe 2005). Pengertian daerah jelajah (home
range) dibedakan dengan daerah inti satwa yang merupakan tempat untuk
melakukan kegiatan khusus, seperti tidur, bersarang kawin, dan lain-lain. Territory akan sangat dipertahankan dengan pihak yang bersangkutan (Burt 1949 dalam
Entebe 2005).
Daerah jelajah satwa dapat berubah-ubah, tergantung kepada pola pergerakan satwa dan jarak tempuhnya, biasanya daerah jelajah tersebut tidak dipertahankan, selain itu daerah jelajah satwa merupakan bagian penting dari populasi satwa, karena selain mencerminkan sifat satwa juga mencerminkan kondisi habitat dimana satwa itu berada. Luas wilayah daerah jelajah sangat tergantung dengan ukuran tubuh satwaliar baik dari golongan herbivora maupun karnivora (Mace et al. 1983).
Terdapat tiga aspek perilaku yang menyangkut kehadiran satwa dengan posisi tertentu di tempat dan pada saat pengamatan dilakukan, yaitu: organisasi sosial, pola pemanfaatan ruang dan pola pemanfaatan waktu. Ketiga aspek ini mempengaruhi keberadaan satwa yang teramati, sehingga pengamat dapat mengelola ketiga aspek ini dengan baik (Santosa 1993).
2.5. Dampak Pembalakan Terhadap Kehidupan Satwaliar
Pembalakan intensif sangat berpengaruh terhadap kerusakan struktur hutan terutama di kawasan hutan yang persentase komersialnya tinggi (Whitmore 1982
dalam Yusuf 1998). Haryanto (1987) juga berpendapat bahwa kegiatan
pembalakan dengan sistem mekanis secara intensif menyebabkan tumbuhnya jenis-jenis pionir secara dominan, seperti Macaranga spp., Mallotus spp., dan
Anthhocephalus spp.. Hal ini berarti menandakan rusaknya struktur hutan, yang berarti rusaknya habitat berbagai jenis satwaliar yang ada didalamnya.
memperkecil pengaruh negatif tersebut. Rinaldi (1985) dalam Yusuf (1998) berpendapat bahwa di Way Kambas Siamang dapat beradapatasi dengan baik pada kondisi vegetasi tanpa strata A yang didominasi oleh jenis-jenis yang berfamili non Dipterocarpaceae. Begitu juga hasil penelitian Haryanto (1987)
Hylobates muelleri dan Presbytis rubicunda yang sanggup beradaptasi dengan baik pada areal dengan strata yang lengkap.
III. METODE PENELITIAN
3.1 Lokasi dan Waktu Penelitian
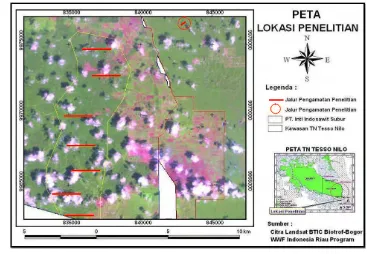
Penelitian ini dilakukan di kawasan TNTN yang berbatasan dengan perkebunan kelapa sawit PT. Inti Indosawit Subur Kecamatan Ukui, Kabupaten Pelalawan, Propinsi Riau. Pengamatan dilakukan selama lebih kurang dua bulan mulai tanggal 1 Maret sampai dengan 10 Mei 2008. Peta lokasi pengamatan ditunjukkan pada Gambar 1.
Gambar 1 Peta lokasi penelitian TNTN dan kebun kelapa sawit PT. Inti Indosawit Subur Ukui, Kabupaten Pelalawan, Propinsi Riau. (Citra Landsat BTIC Biaotrof-Bogor dan WWF Indonesia Riau Program).
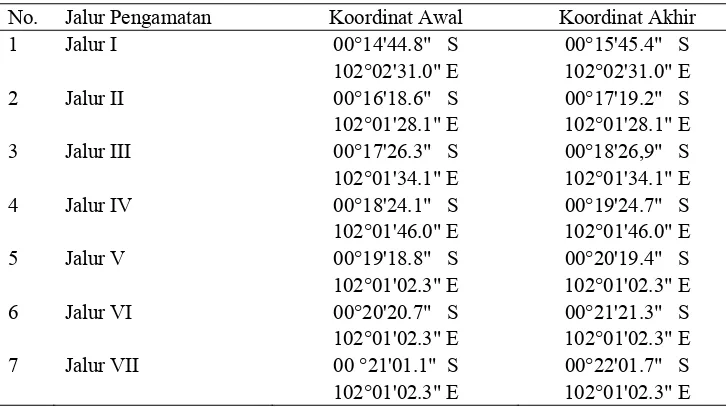
Letak koordinat jalur penelitian di areal TNTN yang berbatasan dengan kebun kelapa sawit PT. Inti Indosawit Subur Ukui, Kabupaten Pelalawan, Propinsi Riau ditunjukan pada Tabel 1 di bawah ini.
Tabel 1. Jalur penelitian di areal kawasan TNTN
No. Jalur Pengamatan Koordinat Awal Koordinat Akhir 1 Jalur I 00°14'44.8" S 00°15'45.4" S
Bahan-bahan yang digunakan dalam kegiatan penelitian ini antara lain: tali tambang (50 m), tali rafia (dua gulung), pita berwarna, buku panduan lapang Primata Indonesia, buku identifikasi tumbuhan ”Check List Tumbuhan Sumatera”, buku identifikasi jenis mamalia ”Seri Buku Panduan Lapangan Mamalia Di Kalimantan, Sabah, Serawak & Brunei Darussalam”, peta kerja, tally sheet dan obat-obatan (P3K). Sedangkan alat-alat yang digunakan dalam kegiatan penelitian ini antara lain: pengukur waktu (stopwatch), kamera digital, golok, binokuler, meteran, kompas, GPS (Global Positioning System), gunting dan alat tulis, PC. Komputer dengan perangkat lunak Arc. GIS dan Minitab 14.
3.3. Metode Pengumpulan Data
3.3.1. Inventarisasi mamalia
Data hasil perjumpaan tidak langsung berupa jejak kaki, kotoran, suara, serta bekas makanan yang dianggap sebagai satu tanda.
NRC (1981) dalam Sugardjito et.al (1997) menyatakan bahwa pengambilan data lapangan untuk primata menggunakan metode transek garis (line transect). Namun untuk penelitian ini digunakan metode transek jalur (strip transect) (Gambar 2). Jalur pengamatan adalah jalur di areal TNTN yang berbatasan dengan kebun kelapa sawit PT. Inti Indosawit Subur. Jumlah jalur yang digunakan sebanyak tujuh jalur pengamatan.
50 m
Jalur transek 2 km
50 m
Gambar 2 Bentuk transek garis pengamatan mamalia.
Pengamatan dilakukan pagi, siang, dan sore hari yaitu pukul 06.00-18.00 WIB. Pengulangan dilakukan sebanyak 3 kali untuk setiap jalur pengamatan. Panjang jalur pengamatan lebih kurang 2000 meter dengan setiap jalur memiliki lebar 100 meter. Dengan menggunakan intensitas sampling 1 % dari luas arael yang diteliti sebesar 13.500 ha, maka didapatkan total luas unit contoh yang harus diamati adalah 135 ha. Dengan total luas unit contoh tersebut dan luas setiap unit contohnya 20 ha maka jumlah jalur yang harus diamati sebanyak 7 jalur.
3.3.2. Inventarisasi vegetasi
Metode yang digunakan dalam analisis vegetasi yaitu metode garis berpetak (Soerianegara dan Indrawan 2002) (Gambar 3). Panjang jalur yang digunakan selang 100 m dengan lebar 20 m, sehingga luas setiap jalur sebesar 0,2 ha.
Soerianegara dan Indrawan (2002) menjelaskan bahwa pada tingkat pertumbuhan semai (a) digunakan ukuran dengan besar 2x2 m, tingkat pertumbuhan pancang (b) ukurannya sebesar 5x5 m, tingkat pertumbuhan tiang (c) ukurannya sebesar 10x10 m, dan tingkat pertumbuhan pohon (d) ukuran yang digunakan sebesar 20x20 m.
c
Gambar 3 Bentuk unit contoh metode garis berpetak dalam inventarisasi vegetasi.
3.3.3. Karakteristik biofisik kawasan
Data biofisik kawasan yang dikumpulkan meliputi ketinggian tempat, temperatur udara, di lokasi pengamatan. Pengukuran ketinggian tempat dilakukan sebelum penentuan unit contoh dengan menggunakan GPS Garmin 76 Csx. Penentuan temperatur udara dengan menggunakan thermometer.
3.4. Analisis Data
3.4.1. Analisis vegetasi
Analisis vegetasi yang dilakukan untuk menentukan komposisi dominasi suatu jenis pohon pada suatu komunitas. Soeranegara & Indrawan (2002) menyatakan bahwa persamaan yang digunakan dalam menentukan komposisis vegetasi adalah sebagai berikut:
Kerapatan Relatif (KR) = x100%
3.4.2. Indeks kekayaan jenis
Kekayaan jenis mamalia dihitung dengan menggunakan metode Margalef Ludwig & Reynolds (1988). Persamaan untuk menentukan jumlah kekayaan jenis adalah sebagai berikut:
Dmg = S – 1
ln (N)
Keterangan: Dmg = Indeks Margalef, N = Jumlah individu seluruh jenis, S = Jumlah jenis mamalia besar
3.4.3. Indeks keanekaragaman jenis (H’)
Ludwig & Reynolds (1988) menyatakan bahwa keanekaragaman jenis mamalia ditentukan dengan menggunakan Indeks keanekargaman Shannon-Wiener dengan rumus sebagai berikut:
H’ = - ∑ pi ln pi; dimana pi = ni/N Keterangan: H’ = Indeks Shannon-Wiener
ni = Jumlah individu setiap jenis N = Jumlah individu seluruh jenis
Ludwig & Reynolds (1988) menyatakan bahwa proporsi kelimpahan jenis mamalia dihitung dengan menggunakan indeks kemerataan, yaitu:
J’ = H’/ln S
Penentuan nilai indeks kemerataan ini berfungsi untuk mengetahui kemerataan setiap jenis mamalia dalam areal pengamatan yang ditentukan, sehingga dapat diketahui keberadaan dominansi jenis mamalia besar.
3.4.5. Kesamaan komunitas mamalia besar
Komunitas mamalia besar ditentukan dengan menggunakan indeks koefisien Jaccard digunakan untuk membandingkan diantara komunitas mamalia besar secara kualitatif (Krebs 1989) dengan memperlihatkan keberadaan mamalia besar, digabungkan antara pertemuan langsung dan tidak langsung. Persamaan yang digunakan untuk mendapatkan nilai indeks Jaccard, adalah:
JI =
Keterangan: a = Pada kedua habitat ditemukan jenis yang sama b = Mamalia besar hanya ditemukan pada habitat A c = Mamalia besar hanya ditemukan pada habitat B
3.4.6. Sebaran mamalia besar di areal TNTN
Pola sebaran spasial suatu komunitas ekologi ditentukan dengan menggunakan indeks penyebaran (Ludwig dan Reynold 1988).
ID = S2/X
Keterangan: ID = Indeks Penyebaran S2 = Ragam contoh X = Rata-rata contoh
Dalam penentuan pola sebarannya, digunakan uji Chi Square dengan persamaan sebagai berikut:
µ2 = ID (N-1)
Keterangan: N = Ukuran contoh atau jumlah Jalur
Persamaan digunakan untuk ukuran contoh kecil (N < 30), maka nilai keragaman populasi membentuk 3 pola, yaitu:
µ20,975≤ µ2≤ µ20,025, maka terjadi sebaran acak µ2> µ20,025, maka terjadi sebaran kelompok
3.4.7. Status perlindungan mamalia besar
Perlindungan terhadap jenis-jenis mamalia besar ditandai dengan status konservasi yang dimiliki setiap jenis mamalia besar. Status konservasi diberikan oleh Pemerintah RI (PP No. 7 Tahun 1999), CITES (Convention on International
Trade in Endengered Species of Wild Flora and Fauna), IUCN (International
Union for Conservation of Nature and Natural Resources).
Beberapa kategori dalam CITES, (Vulnerable = VU) rawan diterapkan pada takson yang tidak termasuk dalam kategori kritis (Critically Endangered = CR) atau genting (Endengered = EN) namun mengalami resiko kepunahan yang sangat tinggi di alam dalam waktu dekat sehingga dapat digolongkan punah in-situ (Excinct in the wild = EW). Sedangkan untuk kekhawatiran minimal (Least Concern = LC) diterapkan pada takson kategori yang cukup mendapat perhatian karena jumlah satwa yang mulai berkurang di alam. Data belum lengkap (Data Deficien = DD). Diterapkan pada takson yang kondisi biologinya mungkin telah diketahui, tetapi data persebaran dan populasinya belum lengkap sehingga analisis status kelangkaannya kurang memadai.
3.4.8. Pemanfaatan waktu aktivitas dan stratifikasi hutan
Analisis pemanfaatan stratifikasi dan waktu aktifitas, yaitu dengan memperhatikan jumlah individu setiap jenis mamalia besar dalam memanfaatkan strata hutan serta waktu untuk beraktivitas.
3.4.9. Pengaruh kebun kelapa sawit terhadap keanekargaman jenis mamalia besar
IV. KONDISI UMUM LOKASI PENELITIAN
4.1 Sejarah Kawasan
Hutan Tesso Nilo sejak awal ditetapkan sebagai hutan produksi terbatas, dimana kawasan ini sebagai daerah hutan untuk mensuplai kebutuhan bahan baku industri plywood dan produksi kayu lainnya. Hutan produksi terbatas ini dikelola oleh HPH Inhutani IV. Pada tanggal 25 Agustus 2003, Menteri Kehutanan mengeluarkan keputusan Nomor: 282/KPts-II/2003 tentang areal HPH Inhutani IV telah dicabut izinnya, dan meminta Gubernur Riau untuk melakukan langkah-langkah persiapan penunjukan kawasan hutan Tesso Nilo sebagai kawasan konservasi gajah.
Pada tanggal 19 Juli 2004, Menteri Kehutanan mengeluarkan keputusan penunjukan tesso nilo sebagai taman nasional yang berada pada areal PT. Inhutani IV melalui SK Nomor: 255/Menhut-II/2004 tentang perubahan fungsi sebagai kawasan hutan produksi terbatas di kelompok Hutan Tesso Nilo yang terletak di kabupaten Pelalawan dan Indragiri Hulu Propinsi Riau seluas ± 38.576 Ha menjadi Taman Nasional Tesso Nilo (TNTN).
4.2. Letak dan Luas
Taman Nasional Tesso Nilo yang luasnya ± 38.576 Ha, secara geografis terletak antara 01° 17'-03° 36' LS dan 101° 31'-102° 44' BT, termasuk kedalam wilayah adaministrasi pemerintahan Kabupaten Pelalawan dan Kabupaten Indragiri Hulu. Kawasan ini dibatasi oleh hutan produksi, perkebunan kelapa sawit, tanah milik dan pemukiman penduduk.
Secara adaministrasi kawasan TNTN berbatasan dengan:
1) Disebelah barat berbatasan dengan HPH Nanjak Makmur dengan vegetasi hutan sepanjang 16.460 m.
2) Disebelah utara berbatasan dengan PT. RAPP dengan vegetasi akasia sepanjang 17.264 m. Desa Lubuk Kembang Bunga dengan vegetasi semak dan sisa hutan sepanjang 3.216 m. Desa Air Hitam dengan vegetasi semak sepanjang 921 m.
PT. Inti Indosawit Subur dengan vegetasi kelapa sawit sepanjang 1.828 m. KKPA dengan vegetasi kelapa sawit dan hutan sepanjang 7.154 m.
4) Disebelah selatan berbatasan dengan PT. Putri Lindung Bulan dengan vegetasi akasia sepanjang 12.178 m. PT. Rimba Lazuardi dengan vegetasi akasia sepanjang 2.938 m. CV. Riau Jambi Sejahtera dengan vegetasi hutan sepanjang 1.075 m.
4.3. Keadaan Iklim dan Topografi
Secara umum, dataran bagian timur dari kawasan Sumatera bagian tengah digolongkan secara lembab dengan curah hujan tahunan yang berkisar antara 2000-3000 mm. Walaupun secara keseluruhan curah hujannya sangat tinggi, curah hujan rata perbulan dapat turun sampai dibawah 60 mm dengan jumlah rata-rata hari hujan pertahun bervariasi antara 120-150 hari. Taman Nasional Tesso Nillo mempunyai topografi relatif datar dan sedikit bergelombang dengan kemiringan 10°-15° dan ketinggian tempat 100-200 m dpl.
4.4. Kondisi Geologi dan Tanah
Kawasan-kawasan pada bagian barat dan timur Pekanbaru, masing-masing digolongkan sebagai dataran rendah dan rawa dataran rendah bagian timur. Kondisi litologinya dicirikan oleh bahan organik semi lapuk yang berasal dari gambut tropis zaman Kuarter dan batuan pasir Kaolinit, batuan liat serta tufa asam yang sudah mengalami proses pelapisan sedimen dari zaman Kuarter. Berdasarkan penggolongan jenis tanah oleh USDA (United State Department
Agronomics), jenis tanah yang mendominasi kawasan tersebut adalah
Tropohemist (sekarang Haplohemist) dan Paeudults.
4.5. Aksesibilitas
Kawasan hutan Tesso Nilo memiliki aksesibilitas yang sangat terbuka, hampir seluruh keliling kawasan ini memiliki jaringan jalan masuk, hal ini mempercepat penurunan kualitas hutan akibat pencurian hasil hutan dan perburuan satwa. Aksesibilitas menuju hutan Tesso Nilo antara lain:
2) Jalan Raya Lintas Timur Sumatera-Ukui-Dusun Bagan Limau 3) Jalan Raya Taluk Kuantan-Air Molek-Baserah-Simpang Inuman
4) Jalan Raya Taluk Kuantan-Air Molek-simpang lala-Desa Pontian mekar 5) Jalan Raya Taluk Kuantan-Air Molek-Simpang Klayan (simpang mangga) 6) Jalan Raya Taluk Kuantan-Air Molek-Simpang Selanjut
7) Jalan Raya Taluk Kuantan-Air Molek-Simpang Sentajo.
4.4. Potensi Flora dan Fauna
4.4.1 Flora
Flora Taman Nasional Tesso Nilo merupakan perwakilan ekosistem transisi dataran rendah dan tinggi dengan potensi keanekaragaman hayati yang tinggi. Diantaranya terdapat 360 jenis flora yang tergolong dalam 165 marga dan 57 suku dalam setiap hektarnya. Berbagai jenis flora yang dilindungi dan terancam punah terdapat juga di taman nasional ini, seperti Kayu bata (Irvingia malayana), Kempas (Koompasia malaccensis), Jelutung (Dyera costulata), Kulim (Scorodocorpus borneensis), Tembesu (Fagraea fragrans), Gaharu (Aquilaria malaccensis), Ramin (Gonystylus bancanus), Keranji (Dialium spp.), Meranti (Shorea spp.), Keruing (Dipterocarpus spp.), dan beberapa jenis Durian (Durio
spp.).
4.4.1 Fauna
Faunanya terdiri dari 107 jenis burung, 23 jenis mamalia, 3 jenis primata, 50 jenis ikan, 15 jenis reptilia, 18 jenis amfibia dan berbagai jenis serangga. Beberapa jenis satwa yang termasuk unggulan diantaranya Harimau sumatera (Panthera tigris sumatrae), Macan dahan (Neofelis nebulosa), Beruang madu (Helarcos malayanus), Rusa (Cervus unicolor), Kijang (Muntiacus muntjak), Kancil (Tragulus javanicus), Babi hutan (Sus spp.), Tapir (Tapirus indicus), dan Bajing (Callosciurus spp.), Owa (Hylobates agilis), Lutung simpai (Presbytis femoralis), dan Beruk (Macaca nemestrina).
Beberapa jenis burung, yaitu: Beo sumatera (Gracula religiosa), Kipas
(Rhipidura albicollis), Elang ular (Spilornis cheela), Alap-alap capung
(Microchierax fringillarius), Kuau (Argusianus argus), Udang pungung merah (Ceyx rufidorsa), Julang jambul hitam (Aceros corrugatus), Kangkareng hitam
(Anorrhinus malayanus), Rangkok badak (Buceros rhinoceros), Ayam hutan
(Gallus gallus), dan Betet ekor panjang (Psittacula longicauda).
Sedangkan untuk reptil dan amfibi, yaitu Ular kawat/ular hitam (Ramphotyphlops braminus), Ular kopi (Elaphe flavolineata), Ular picung air (Xenochrophis trianguligerus), Ular cabe kecil (Maticora intestinalis), Ular kobra (Ophiphagus hannah), Sanca sawah (Python reticulatus), Ular gendang/phyton darah sumatera (Python curtus), dan Buaya sinyulong (Tomistoma schlegeleii). Katak serasah berbintik (Leptobrachium hendricksoni), Kodok buduk sungai
(Bufo asper), Kodok buduk (B. melanostictus), Katak lekat (Kalophrynus
V. HASIL DAN PEMBAHASAN
5.1. Kondisi Habitat
Hutan Tesso Nilo merupakan salah satu hutan hujan dataran rendah yang tersisa di Sumatera saat ini dan merupakan daerah aliran Sungai Kampar. Hutan Tesso Nilo memiliki beberapa anak sungai dan sungai besar yang bermuara ke Sungai Kampar. Habitat yang terdapat di areal penelitian adalah habitat eks areal HPH PT. Inhutani IV. Taman Nasional Tesso Nilo dinyatakan sebagai hutan yang terkaya keanekaragaman hayati di dunia dengan ditemukannya 218 species tumbuhan Vascular di petakan 200 meter persegi oleh Center For Biodiversity Management dari Australia pada tahun 2001 (LIPI 2003 dalam Dephut 2008).
Penelitian dilakukan di areal kawasan TNTN didasarkan pada pengamatan dari tujuh Jalur yang dibuat petak contoh berbatasan dengan perkebunan kelapa sawit PT. Inti Indosawit Subur Ukui Riau. Dari hasil penelitian didapatkan data tumbuhan yang dapat diidentifikasi sebanyak 111 jenis dari 43 famili. Berdasarkan familinya, Dipterocarpaceae memiliki jumlah jenis terbanyak yaitu 25 jenis. Fabaceae sebanyak 8 jenis. Burceraceae dan Anarcadinaceae 6 jenis. Jenis-jenis tumbuhan yang teridentifikasi ditunjukkan dalam Gambar 4 dan secara lebih rinci ditunjukkan pada Lampiran 1.
5.1.1. Jalur I
Berdasarkan hasil analisis vegetasi yang dilakukan, diperoleh jenis-jenis yang dominan di kawasan ini. Pada tingkat pertumbuhan semai, didominasi oleh Nasi-nasi (Anisophyllea disticha) dengan INP sebesar 20,12%. Tingkat pertumbuhan pancang didominasi oleh Tapak-tapak (Sindora wallichii) dengan INP sebesar 22,50%. Meranti bunga (Shorea platycarpa) mendominasi pada tingkat pertumbuhan tiang dengan INP sebesar 47,09%. Tingkat pertumbuhan pohon didominasi oleh jenis Kelat (Gonystylus forbesii) dengan INP sebesar 44,84%. Jenis-jenis tumbuhan yang mendominasi pada Jalur I ditunjukkan pada Tabel 2 dan secara lengkap ditunjukkan pada Lampiran 3-6.
Tabel 2. Rekapitulasi jenis-jenis yang mendominasi pada Jalur I
Tingkat Pertumbuhan Nama Lokal Nama Ilmiah Famili INP (%)
Semai Nasi-nasi Anisophyllea disticha Rhizophoraceae 20.12
Meranti Sepat Shorea macrantha Dipterocarpaceae 18.63
Asem Triomma malaccensis Burceraceae 17.27
Pancang Tapak-tapak Sindora wallichii Fabaceae 22.50
Kayu hitam Diospyros transitoria Ebenaceae 17.50
Asem Triomma malaccensis Burceraceae 15.00
Tiang Meranti bunga Shorea platycarpa Dipterocarpaceae 47.09
Kedondong Ailanthus integrifolia Simaroubaceae 24.71
Tapak-tapak Sindora wallichii Fabaceae 17.76
Kelat Gonystylus forbesii Thymeliaceae 16.73
Pohon Kelat Gonystylus forbesii Thymeliaceae 44.84
Daru-daru Cantleya corniculata Icacinaceae 22.86
Kompas Santiria spp. Burceraceae 20.58
5.1.2. Jalur II
bunga (Shorea platycarpa) dengan INP sebesar 26,20%. Vegetasi tingkat pertumbuhan pancang didominasi oleh Jejambu (Eugenia sp.) dengan INP sebesar 22,50 %. Vegetasi tingkat pertumbuhan tiang yang mendominasi adalah Arang-arang (Eugenia sp.) dengan INP sebesar 43,17%. Vegetasi tingkat pertumbuhan pohon didominasi oleh Meranti kunyit (Shorea conica) dengan INP sebesar 32,25%. Jenis-jenis tumbuhan yang mendominasi pada Jalur II ditunjukkan pada Tabel 3 dan secara lengkap ditunjukkan pada Lampiran 7-11.
Tabel 3. Rekapitulasi jenis-jenis yang mendominasi pada Jalur II
Tingkat Pertumbuhan Nama Lokal Nama Ilmiah Famili INP (%)
Semai Meranti bunga Shorea platycarpa Dipterocarpaceae 26.20
Bengku P. xanthochymum Sapotaceae 25.40
Babi kurus Alangium ridleyi Alangiaceae 23.57
Pancang Jejambu Eugenia sp. Myrtaceae 31.57
Meranti sepat Shorea macrantha Dipterocarpaceae 21.76
Meranti rambai Shorea conica Dipterocarpaceae 15.88
Pisang mawe Dillenia sumatrana Dilleniaceae 15.88
Tiang Arang-arang Eugenia sp. Myrtaceae 43.17
Jejambu Eugenia sp. Myrtaceae 33.14
Bengku P. xanthochymum Sapotaceae 29.03
Pohon Meranti kunyit Shorea conica Dipterocarpaceae 32.25
Kulim S. bracteatus Moraceae 29.43
Balam Payena lanceolata Sapotaceae 21.80
5.1.3. Jalur III
pohon didominasi oleh Medang keladi (Talauma gigantifolia) dengan INP sebesar 44,84%. Jenis-jenis tumbuhan yang mendominasi pada Jalur III ditunjukkan pada Tabel 4 dan secara lengkap ditunjukkan pada Lampiran 12-15.
Tabel 4. Rekapitulasi jenis-jenis yang mendominasi pada Jalur III
Tingkat Pertumbuhan Nama Lokal Nama Ilmiah Famili INP (%)
Semai Kelat Gonystylus forbesii Thymeliaceae 39.51
Medang Diospyros curranii Ebenaceae 20.34
Medang keladi Talauma gigantifolia Magnoliaceae 19.78
Pancang Kelat Gonystylus forbesii Thymeliaceae 40.48
Kopi-kopi C. castanocarpus Euphorbiaceae 39.93
Medang Diospyros curranii Ebenaceae 30.68
Tiang Medang Diospyros curranii Ebenaceae 51.18
Petaling gajah H. longifolium Flacourtiaceae 41.28
Kopi-kopi C. castanocarpus Euphorbiaceae 32.11
Pohon Medang keladi Talauma gigantifolia Magnoliaceae 44.84
Balam Payena lanceolata Sapotaceae 35.13
Durian hutan Durio spp. Bombacaceae 29.81
5.1.4. Jalur IV
Tabel 5. Rekapitulasi jenis-jenis yang mendominasi pada Jalur IV
Tingkat Pertumbuhan Nama Lokal Nama Ilmiah Famili INP (%)
Semai Kelat Gonystylus forbesii Thymeliaceae 84.81
Balam Payena lanceolata Sapotaceae 26.27
Jejambu Eugenia sp. Myrtaceae 19.42
Babi kurus Alangium ridleyi Alangiaceae 17.10
Pancang Jejambu Eugenia sp. Myrtaceae 38.11
Balam Payena lanceolata Sapotaceae 35.41
Garam-garam T. feotidissima Combretaceae 25.81
Tiang Jejambu Eugenia sp. Myrtaceae 80.97
Mahang Macaranga spp. Euphorbiaceae 42.51
Balam Payena lanceolata Sapotaceae 30.31
Pohon Benau Mangifera spp. Anacardiacea 27.46
Meranti sbrg Shorea acuminata Dipterocarpaceae 23.03
Daru-daru Cantleya corniculata Icacinaceae 22.96
5.1.5. Jalur V
Tabel 6. Rekapitulasi jenis-jenis yang mendominasi pada jalur V
Tingkat Pertumbuhan Nama Lokal Nama Ilmiah Famili INP (%)
Semai Sendok-sendok E. diadenum Euphorbiaceae 40.13
Asam kumbang Adinandra dumosa Theaceae 32.31
Daru-daru Cantleya corniculata Icacinaceae 18.94
Pancang Sendok-sendok E. diadenum Euphorbiaceae 33.44
Gerunggang Cratoxylon spp. Hypericaceae 27.55
Jejambu Eugenia sp. Myrtaceae 25.23
Meranti bunga Shorea platycarpa Dipterocarpaceae 25.23
Tiang Sendok-sendok E. diadenum Euphorbiaceae 72.38
Meranti bunga Shorea platycarpa Dipterocarpaceae 32.94
Rengas Gluta renghas Anacardiacea 31.95
Jejambu Eugenia spp. Myrtaceae 29.96
Pohon Meranti sepat Shorea macrantha Dipterocarpaceae 37.37
Meranti Shorea spp. Dipterocarpaceae 33.07
Kompas Santiria spp. Burceraceae 28.86
5.1.6. Jalur VI
Berdasarkan data hasil analisis vegetasi, jenis-jenis yang dominan di kawasan ini untuk vegetasi tingkat pertumbuhan semai didominasi oleh Kulim
(Scorodocarpus bracteatus) dengan INP sebesar 74,66%. Vegetasi tingkat
pertumbuhan pancang didominasi oleh Jejambu (Eugenia sp.) dengan INP sebesar 56,14%. Vegetasi tingkat pertumbuhan tiang yang mendominasi adalah Bintangur
(Colopyhllum soulattri) dengan INP sebesar 79,58%. Vegetasi tingkat
Tabel 7. Rekapitulasi jenis-jenis yang mendominasi pada Jalur VI
Tingkat Pertumbuhan Nama Lokal Nama Ilmiah Famili INP (%)
Semai Kulim S. bracteatus Moraceae 74.66
Jengkol P. labatum Fabaceae 31.25
Gerunggang Cratoxylon spp. Hypericaceae 24.11
Pancang Jejambu Eugenia sp. Myrtaceae 56.14
Gerunggang Cratoxylon spp. Hypericaceae 48.92
Rengas Gluta rengas Anacardiacea 23.66
Bintangur Calophyllum soulattri Guttaceae 22.02
Tiang Bintangur Calophyllum soulattri Guttaceae 79.58
Jejambu Eugenia sp. Myrtaceae 70.32
Ariung D. verruscosus Dipterocarpaceae 50.65
Pohon Resak Vatica spp. Dipterocarpaceae 61.31
Mersawa Anisoptera curtisii Dipterocarpaceae 56.31
Petaling gajah H. longifolium Flacourtiaceae 29.49
5.1.7. Jalur VII
Tabel 8. Rekapitulasi jenis-jenis yang mendominasi pada jalur VII
Tingkat Pertumbuhan Nama Lokal Nama Ilmiah Famili INP (%)
Semai Gerunggang Cratoxylon spp. Hypericaceae 48.08
Jejambu Eugenia sp. Myrtaceae 43.08
Putat Barringtonia spp. Lecythidaceae 25.77
Pancang Jejambu Eugenia spp. Myrtaceae 57.98
Gerunggang Cratoxylon spp. Hypericaceae 47.98
Kulim Scorodocarpus
bracteatus
Moraceae 18.21
Tiang Jejambu Eugenia sp. Myrtaceae 80.61
Kompas Santiria spp. Burceraceae 53.26
Mahang Macaranga spp. Euphorbiaceae 37.92
Bintangur Calophyllum soulattri Guttaceae 37.29
Pohon Resak Vatica spp. Dipterocarpaceae 57.15
Bintangur Calophyllum soulattri Guttaceae 40.53
Jejambu Eugenia sp. Myrtaceae 38.57
5.2. Keanekaragaman Jenis Mamalia Besar
Tabel 9. Jenis mamalia besar yang ditemukan di lokasi penelitian TNTN berdasarkan pengamatan langsung
Jumlah individi setiap jalur
Nama lokal Nama ilmiah
I II III IV V VI VII
Keterangan: Total merupakan jumlah spesies yang ditemukan setiap jalur pengamatan
Umumnya jumlah individu yang ditemukan berdasarkan perjumpaan langsung, Hal ini dapat disebabkan jenis mamalia besar yang terdapat di TNTN telah beradaptasi dengan baik dengan kondisi habitat hutan sekunder yang telah terganggu oleh berbagai aktivitas masyarakat, seperti penebangan liar, perburuan satwaliar, dan perambahan hutan.
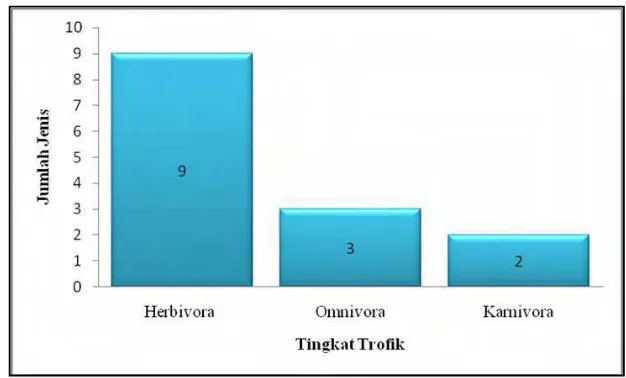
simpai, Owa ungko, Rusa sambar, Pelanduk, Kancil, dan Tapir). Jumlah jenis satwa berdasarkan tingkat trofik ditunjukkan pada Gambar 5.
Gambar 5 Jumlah jenis mamalia besar berdasarkan tingkat trofik.
Berdasarkan pengelompokkan jenis mamalia besar, menurut tingkat trofik diketahui keseimbangan ekosistem pada mamalia besar masih tergolong baik. Hal ini diketahui berdasarkan jumlah jenis herbivora yang lebih banyak dari omnivora dan karnivora yang membentuk piramida. Apabila jumlah jenis karnivora lebih banyak dari jenis herbivora, maka jejaring makanan dalam ekosistem akan menjadi tidak seimbang. Noerdjito et al. (2005) keseimbangan ekosistem telah diatur secara alami melalui mekanisme rangkaian penyedian dan keseimbangan jejaring makanan.
5.2.1. Keanekargaman jenis mamalia besar (Indeks Shannon-Wiener)
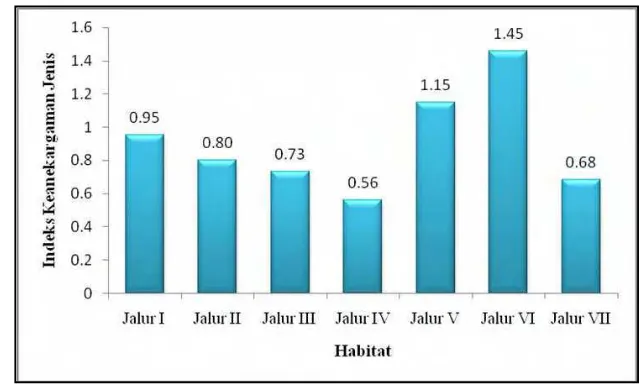
Tingkat keanekargaman jenis mamalia tertinggi di Jalur VI dengan nilai indeks Shannon Wiener sebesar 1,46. Tingkat keanekaragaman terendah terdapat di Jalur IV dengan nilai indeks Shannon Wiener sebesar 0,56. Nilai indeks keanekaragaman jenis pada setiap Jalur ditunjukkan pada Gambar 6. Secara lebih rinci, nilai indeks keanekargaman jenis ditunjukkan pada Lampiran 22.
Gambar 6 Nilai indeks Shannon Wiener di setiap Jalur.
Keanekaragaman jenis mamalia besar hampir merata pada setiap Jalur, karena habitat penelitian TNTN memiliki habitat yang hampir sama. Selain itu Zorenko dan Leontyeva (2003) menyatakan bahwa faktor luasan mempengaruhi nilai indeks yang dimiliki. Soerianegara dan Indrawan (2002) menambahkan bahwa ukuran contoh yang semakin besar menyebabkan jumlah jenis yang ditemukan bertambah.
(a) (b)
Gambar 7 Jenis mamalia besar dari ordo primata yang ditemukan (a) Owa ungko (Hylobates agilis), (b) Lutung budeng (Trachypithecus auratus).
5.2.2. Kekayaan jenis mamalia
Tingkat kekayaan jenis merupakan salah satu ukuran keanekaragaman yang dapat digunakan untuk mempelajari tingkatan suksesi. Tingkat keanekaragaman ini diukur berdasarkan jumlah jenis atau dapat ditentukan langsung dengan melihat jumlah jenisnya. Jumlah jenis mamalia besar beserta jumlah individunya yang ditemukan dalam suatu kawasan akan berpengaruh terhadap nilai indeks kekayaan jenis Margalef. Toth dan Kiss (1999) menyatakan bahwa peningkatan jumlah jenis akan menyebabkan indeks nilai Margalef semakin tinggi. Dikatakan lebih lanjut bahwa bila jumlah individu setiap jenis yang meningkat akan menyebabkan nilai indeks Margalef yang semakin menurun.
Gambar 8 Indeks kekayaan jenis di setiap Jalur.
Selama penelitian di Jalur VI ditemukan satu jenis mamalia besar yang tidak ditemukan pada lokasi lain yaitu Musang akar. Musang akar ditemukan secara langsung pada saat pagi hari dengan kondisi cuaca yang mendung. Sebagian besar satwa ini beraktivitas pada malam hari dan bersifat arboreal. Selain itu mamalia besar lainnya yang jarang ditemukan adalah Siamang, satwa ini hidup soliter dan banyak beraktivitas diatas pohon dan bergerak bebas pada kanopi pohon dan tidur di percabangan pohon yang besar.
5.2.3. Kemerataan jenis mamalia besar
Untuk mengetahui tingkat kemerataan kelimpahan individu antar suatu jenis mamalia digunakan nilai indeks kemerataan. Selain itu nilai indeks ini juga dapat digunakan sebagai indikator gejala dominansi diantara tiap jenis dalam komunitas. Pada saat setiap jenis memiliki jumlah individu yang sama-sama berlimpah akan menyebabkan nilai indeks kemerataan yang maksimum. Sebaliknya bila kelimpahan individu pada masing-masing jenis berbeda jauh, maka akan menyebabkan nilai indeks kemerataan semakin menurun. Nilai indeks kemerataan terendah yaitu di Jalur III dengan nilai indeks sebesar 0,42 dan nilai kemerataan tertinggi yaitu di Jalur VI dengan nilai indeks sebesar 0,91.
mengindikasikan bahwa tidak terdapat jenis yang mendominasi pada komunitas mamalia besar pada Jalur VI. Berdasarkan data hasil penelitian, tingkat kemerataan jenis mamalia di setiap Jalur berkisar antara nilai 0,42-0,91. Nilai indeks kemerataan jenis pada setiap Jalur penelitian ditunjukkan pada Gambar 9 dan secara lengkap ditunjukkan pada Lampiran 28.
Gambar 9 Indeks kemerataan jenis di setiap Jalur.
Pada keseluruhan habitat, didapatkan nilai indeks kemerataan jenis sebesar 0,59. Hal ini mengindikasikan bahwa masih terdapat jenis-jenis yang dominan dalam kawasan TNTN, yang diketahui dengan melimpahnya jumlah individu dan menyebar pada daerah perbatasan TNTN. Owa ungko dan Lutung budeng menyebar pada setiap daerah perbatasan TNTN dengan kebun kelapa sawit. Hal ini didukung karena kedua satwa ini memiliki ekologi yang sama yaitu ditemukan pada hutan dataran rendah.
5.2.4. Kesamaan komunitas jenis mamalia
Kesamaan komunitas dilakukan dengan membandingkan jenis mamalia besar antar Jalur. Berdasarkan indeks kesamaan komunitas, maka keberadaan mamalia besar di areal perbatasan TNTN dengan kebun kelapa sawit PT. Inti Indosawit Subur Ukui memiliki kesamaan komunitas yang cukup tinggi. Jalur yang memiliki kesamaan komunitas tinggi, diantaranya: Jalur I dan VII, Jalur VI dan VII, sedangkan kesamaan komunitas yang rendah yaitu Jalur IV dan V. Jalur yang memiliki nilai kesamaan komunitas yang lebih tinggi disebabkan adanya kemiripan kompisisi jenis yang lebih besar dibandingkan dengan Jalur lainnya. Nilai indeks kesamaan komunitas mamalia besar ditunjukkan pada Tabel 10. Tabel 10. Indeks kesamaan komunitas antar jalur
x/y I II III IV V VI VII
Tingginya kesamaan komunitas mamalia besar juga dipengaruhi dengan kondisi lingkungan yang mendukung kemampuan beradaptasi. Apabila jenis mamalia besar di TNTN dapat beradaptasi dengan kondisi lingkungan yang berbeda, baik disebabkan dengan adanya gangguan ataupun perubahan iklim akan menyebabkan komunitas mamalia besar tetap lestari. Terdapat 5 jenis mamalia besar yang sama, yaitu Lutung budeng, Owa ungko, Tapir, Beruang madu, Babi hutan, dengan perbedaan jumlah jenis sebanyak 2 di Jalur VI (7 jenis di Jalur I, 7 jenis di Jalur VII, 5 jenis di Jalur VI).
5.3. Sebaran Mamalia Besar di Areal TNTN
hidupnya (Alikodra, 1990). Sebaran populasi jenis mamalia besar di areal studi seperti ditunjukan pada Tabel 11.
Tabel 11. Sebaran jenis mamalia besar dia areal TNTN
Parameter
Mamalia besar yang ditemukan secara langsung pada areal TNTN memiliki 2 pola sebaran, yaitu mengelompok dan acak. Jenis mamalia besar yang memiliki sebaran mengelompok merupakan ordo primata yang memiliki sifat suka bergerombol seperti Monyet ekor panjang, Lutung budeng, dan Lutung simpai. Selain itu jenis mamalia besar dari ordo primata seperti Owa ungko dan Siamang memiliki pola sebaran acak, karena kedua primata ini memiliki sifat soliter. Mamalia besar yang memiliki sebaran acak seperti Babi hutan, Pelanduk, Kancil, Rusa sambar, Musang akar dan Tapir merupakan satwa yang peka terhadap keberadaan di habitatnya. Sehingga sangat sedikit ditemukan jenis tersebut di areal penelitian.
5.4. Status Konservasi Mamalia Besar
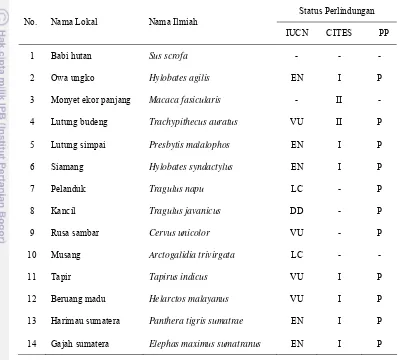
1999), terancam punah (IUCN), dan termasuk dalam kategori appendix I (CITES), seperti Owa ungko, Lutung simpai, Siamang, Harimau sumatera, dan Gajah sumatera. Status konservasi jenis mamalia besar yang ditemukan pada areal penelitian ditunjukkan pada Tabel 12.
Tabel 12. Daftar jenis mamalia dilindungi yang ditemukan di lokasi penelitian
Status Perlindungan
No. Nama Lokal Nama Ilmiah
IUCN CITES PP
1 Babi hutan Sus scrofa - - -
2 Owa ungko Hylobates agilis EN I P
3 Monyet ekor panjang Macaca fasicularis - II -
4 Lutung budeng Trachypithecus auratus VU II P
5 Lutung simpai Presbytis malalophos EN I P
6 Siamang Hylobates syndactylus EN I P
7 Pelanduk Tragulus napu LC - P
8 Kancil Tragulus javanicus DD - P
9 Rusa sambar Cervus unicolor VU - P
10 Musang Arctogalidia trivirgata LC - -
11 Tapir Tapirus indicus VU I P
12 Beruang madu Helarctos malayanus VU I P
13 Harimau sumatera Panthera tigris sumatrae EN I P
14 Gajah sumatera Elephas maximus sumatranus EN I P
5.5. Pemanfaatan Waktu Aktivitas dan Stratifikasi Hutan
Setiap jenis makhluk hidup akan selalu berinteraksi dengan lingkungan sekitarnya. Faktor lingkungan seperti faktor fisik (iklim, air, dan substrat) dan faktor biotik (vegetasi dan makanan) merupakan faktor yang mempengaruhi penyebaran mamalia besar (Vaughan 1985). Faktor ini akan menyebabkan setiap jenis satwaliar, khususnya mamalia besar memiliki kebiasan tertentu dalam memanfaatkan lingkungannya berupa hutan berdasarkan aktivitas dan stratifikasi hutan.
5.5.1. Pemanfaatan waktu aktivitas mamalia besar
Pola aktivitas setiap jenis mamalia besar yang ditemukan, berada berdasarkan waktu aktivitas yang umumnya dilakukan dan dikategorikan berdasarkan pengamatan di lapangan. Terbagi menjadi beberapa kategori, yaitu aktivitas bersuara, bergerak, makan, berkelahi dan lainnya. Aktivitas berkelahi merupakan kegiatan satwa dalam mempertahankan daerah teritorinya dari individu atau kelompok lainnya.
Satwaliar pada umumnya ada yang bersifat diurnal dan juga nokturnal. Satwa yang aktif pada pagi dan sore hari dalam keadaan terang tergolong satwa yang bersifat diurnal, sedangkan satwa yang aktif di malam hari (gelap) adalah satwa nokturnal. Selain ditinjau berdasarkan pemanfaatan waktu aktivitas satwaliar, dikategorikan juga berdasarkan pemanfaatan stratifikasi hutan yaitu sebagai satwa arboreal dan terestrial.
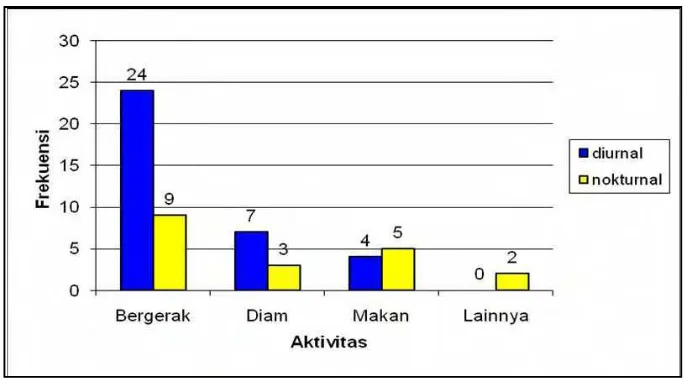
Pembagian berdasarkan aktivitas dibagi kedalam tiga bagian diantaranya, bergerak, diam, dan makan secara umum banyak dilakukan pada aktivitas diurnal. Perbedaan pada total individu jenis mamalia besar dalam pemanfaatan aktivitas diurnal dan nokturnal disajikan dalam persentase. Total individu setiap jenis yang melakukan aktivitas diurnal sebesar 64,81%, sedangkan yang memanfaatkan aktivitas nokturnal sebesar 35,19%. Pembagian bentuk aktivitas mamalia besar ditunjukkan pada Gambar 10.
Gambar 10 Pembagian bentuk aktivitas mamalia besar.
Pada waktu pagi, mamalia besar sebagian besar melakukan aktivitas untuk bergerak dan diam dibanding pada waktu sore hari. Aktivitas bergerak untuk mamalia besar diantaranya seperti melompat untuk jenis-jenis primata dan berjalan, sedangkan aktivitas diam pada mamalia besar digunakan waktunya untuk beristirahat dan berteduh.
Pada aktivitas makan, jenis mamalia besar yang ditemukan lebih banyak melakukannya pada sore hari. Hal ini dapat disebabkan oleh bentuk adaptasi jenis mamalia besar dalam menghindari adanya gangguan. Pada saat pagi hari, masyarakat banyak melakukan aktivitas kedalam hutan untuk mencari kayu bakar, hal ini yang diduga berpengaruh terhadap bentuk adaptasi jenis mamalia besar yang lebih banyak memanfaatkan waktu sore hari untuk mencari makan.
5.5.2. Pemanfaatan stratifikasi hutan mamalia besar
(2003) menyatakan bahwa perubahan ketinggian pada hutan hujan di Atlantic dapat mengubah komposisi dari komunitas pada lapisan hutan yang berbeda tanpa mengubah pola pemanfaatan habitat vertikal secara spesifik. Primack et al. (1998) menyatakan bahwa kekayaan jenis vertebrata yaitu mamalia berkorelasi dengan struktur kompleks dari hutan.
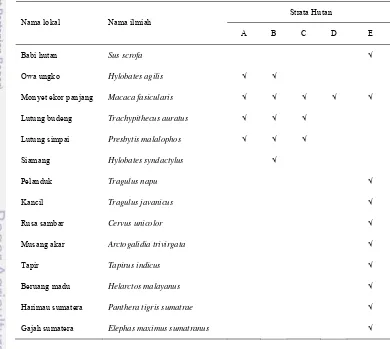
Setiap strata hutan memilki kemampuan dalam mendukung kehidupan jenis-jenis satwaliar tertentu (Alikodra 2002). Kartono et al. (2000) mengelompokkan sebaran spasial vertikal pada jenis-jenis mamalia yang ditemukan di Muara Bungo, Jambi kedalam 5 kelompok. Soerianegara dan Indrawan (2002) membagi strata hutan atas strata A (>30 m), strata B (20-30 m), strata C (4-20 m), strata D (1-4 m), dan strata E (0-1 m). Pemanfaatan strata hutan oleh masing-masing jenis mamalia besar ditunjukkan pada Tabel 13.
Tabel 13. Pemanfaatan strata hutan oleh masing-masing jenis mamalia besar Strata Hutan
Nama lokal Nama ilmiah
A B C D E
Babi hutan Sus scrofa √
Owa ungko Hylobates agilis √ √
Monyet ekor panjang Macaca fasicularis √ √ √ √ √
Lutung budeng Trachypithecus auratus √ √ √
Lutung simpai Presbytis malalophos √ √ √
Siamang Hylobates syndactylus √
Pelanduk Tragulus napu √
Kancil Tragulus javanicus √
Rusa sambar Cervus unicolor √
Musang akar Arctogalidia trivirgata √
Tapir Tapirus indicus √
Beruang madu Helarctos malayanus √
Harimau sumatera Panthera tigris sumatrae √
Dalam pembagian berdasarkan stratifikasi hutan, diketahui bahwa Monyet ekor panjang memiliki sebaran vertikal yang lebih luas jika dibandingkan mamalia besar lainnya. Monyet ekor panjang memanfaatkan setiap strata hutan yang telah dibagi hal ini disebabkan karena kebiasaan Monyet ekor panjang untuk tidak memilih sumberdaya pakan tertentu.
Mamalia besar yang memanfaatkan strata A yaitu Monyet ekor panjang, Owa ungko, Lutung budeng dan Lutung simpai. Santoso (1996) menyatakan bahwa pola aktivitas monyet ekor panjang di Pulau Tinjil yang banyak aktif pada tajuk pohon mengindikasikan bahwa ketersediaan sumberdaya pakannya sedang berlimpah pada stratifikasi atas. Lutung budeng dan Simpai lebih banyak memanfaatkan strata A dibanding jenis mamalia lainnya karena kebutuhan pakan akan daun muda atau pucuk daun. Seperti halnya pernyataan Santoso (1996), Lutung budeng juga lebih banyak memanfaatkan strata hutan A disebabkan kebutuhannya akan daun muda yang terdapat di tajuk pohon teratas.
Berdasarkan data pengamatan, rata-rata primata memanfaatkan pohon pada strata A dan strata B. Hal ini disebabkan karena pada ketinggian ini tersedia sumber pakan yang dibutuhkan oleh jenis-jenis primata seperti buah, daun, dan serangga. Selain itu, jenis-jenis primata dapat melakukan pergerakan yang lebih mudah dari strata B untuk berpindah ke strata A atau strata C.
Jenis mamalia besar selain ordo primata, merupakan jenis-jenis yang mendiami strata E atau lantai hutan. Jenis-jenis yang teramati memanfaatkan strata E adalah Babi hutan, Pelanduk, Kancil, Rusa sambar, Tapir, Beruang madu, Pelanduk, Kancil, Tapir dan Rusa sambar memanfaatkan lantai hutan dalam memenuhi kebutuhannya akan rumput dan daun sebagai sumber pakan, sedangkan Babi hutan dan Beruang madu memerlukan biota-biota dalam tanah ataupun rumput. Alikodra (2002) menyatakan bahwa variasi jenis-jenis satwaliar di lantai hutan ditentukan oleh komposisi jenis tumbuhan, kerapatan dan letak tempatnya.
5.6. Pengaruh Kebun Kelapa Sawit terhadap Keanekaragaman Jenis Mamalia Besar
sesuai untuk jenis lainnya, karena setiap jenis satwaliar menghendaki kondisi habitat yang berbeda-beda. Habitat yang baik adalah habitat yang mampu mendukung segala kebutuhan satwaliar, seperti makanan, air, dan tempat berlindung. Daya dukung habitat di tiap-tiap lokasi penelitian kurang baik, hal ini dapat dilihat dengan keanekaragaman vegetasi.
Untuk mengetahui pengaruh kebun kelapa sawit terhadap jenis mamalia yang ditemukan, maka digunakanlah analisis dengan metode regresi linear sederhana yaitu mengetahui hubungan antara jarak kebun kelapa sawit di tempat pengamatan dengan keanekaragaman mamalia di tempat pengamatan. Hasil analisis diperoleh persamaan regresi linear sederhana sebagai berikut:
y = 0,443 + 0,000098 x
Dengan y merupakan keanekaragaman mamalia dan x merupakan jarak tempat pengamatan dengan perkebunan kelapa sawit. Persamaan regresi tersebut memperlihatkan bahwa apabila jarak tempat pengamatan dengan kebun kelapa sawit bertambah sejauh 1 m, maka keanekaragaman jenis mamalia di tempat pengamatan tersebut bertambah sebesar 0,000098 kalinya. Apabila jarak tempat pengamatan dengan perkebunan kelapa sawit bernilai 0, maka keanekaragaman jenis mamalia besar di tempat pengamatan tersebut adalah sebesar 0,443.
Hasil analisis regresi linear memperlihatkan tidak terdapat hubungan yang nyata antara jarak tempat pengamatan perkebunan kelapa sawit dengan keanekaragaman mamalia di tempat pengamatan tersebut. Menurut Alikodra (2002) habitat yang sesuai bagi satu jenis satwaliar belum tentu sesuai untuk jenis lainnya. Hal ini disebabkan bahwa setiap jenis satwaliar menghendaki kondisi yang berbeda-beda. Kartono et al. (2003) menambahkan bahwa kerusakan habitat dapat menyebabkan penurunan kekayaan jenis dan penurunan tersebut akan lebih terlihat jelas pada habitat terisolasi berukuran kecil.
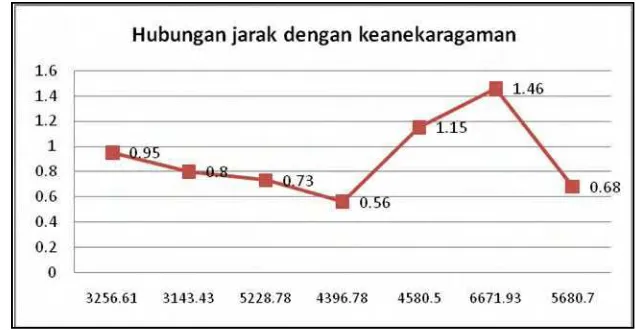
Gambar 11 Hubungan jarak pengamatan dengan keanekaragaman.
Berdasarkan Gambar 11 terlihat pada jalur pengamatan VI yang berjarak 6671,93 m dari kebun kelapa sawit memiliki keanekaragaman mamalia terbesar yaitu sebesar 1,46. Faktor yang dapat mempengaruhinya adalah kawasan hutan lebih jauh dengan pemukiman masyarakat. Intensitas masyarakat pada jalur ini lebih rendah sehingga menyebabkan kondisi hutan lebih baik.
Keanekaragaman mamalia terkecil terdapat pada jalur IV yang berjarak 4396,78 m dari kebun kelapa sawit. Faktor yang menyebabkan keanekaragaman di Jalur IV lebih kecil yaitu 0,56, karena pada jalur ini merupakan jalur yang memiliki gangguan yang tinggi sehingga sedikit ditemukannya mamalia besar. Selain itu intensitas masyarakat yang tinggi pada saat pengamatan seperti penebangan hutan dan perburuan satwaliar juga menyebabkan rendahnya keanekargaman pada jalur ini. Folke et al. (1996) menyatakan bahwa dalam pendekatan perlindungan terhadap keanekargaman hayati memasukan manusia sebagai salah satu implikasi.
5.7. Ancaman terhadap Kelestarian Jenis Mamalia
Berdasarkan hasil pengamatan di lapangan beberapa ancaman yang baik secara langsung maupun tak langsung dapat mengancam kelestarian jenis mamalia seperti penebangan liar, perburuan liar, dan perambahan. Dampak utama aktivitas ini adalah berkurang atau hilangnya beberapa jenis tumbuhan yang bernilai ekonomi tinggi. Selain itu juga berdampak pada rusaknya habitat TNTN yang merupakan daerah penting bagi jenis satwaliar, salah satunya mamalia.
5.7.1. Penebangan liar
Penebangan liar terjadi pada semua areal perbatasan TNTN dengan kebun kelapa sawit PT. Inti Indosawit Subur Ukui, Kabupaten Pelalawan, Propinsi Riau. Kegiatan penebangan dilakukan oleh masyarakat sekitar untuk memenuhi kebutuhan pokoknya, salah satunya untuk mendirikan bangunan atau rumah. Tingkat kerusakan terparah terjadi pada Jalur VI dan Jalur VII, alat yang digunakan berupa mesin gergaji (chainsaw) karena pada saat penelitian terdengar suara chainsaw. Hal ini juga didukung dengan ditemukannya potongan-potongan kayu yang sudah menjadi balok dan serbuk gergaji (Gambar 12).
Gambar 12 Bekas potongan kayu dalam kawasan TNTN.
5.7.2. Perburuan liar
terbuat dari ranting, dahan dan tali. Selain itu pada saat penelitian di lapangan bertemu dengan sekelompok orang di areal kawasan TNTN yang membawa senjata api dengan tujuan berburu.
5.7.3. Perambahan hutan
Semua areal perbatasan TNTN telah dirambah oleh masyarakat, baik masyarakat lokal, maupun masyarakat perkotaan yang bermaksud utuk membuat areal perkebunan kelapa sawit. Perambahan sangat berdampak pada keberadaan mamalia dan habitatnya. Keberadaan PT. Inti Indosawit Subur, Ukui yang berbatasan dengan TNTN semakin menjadikan alasan bagi masyarakat untuk terus membuka lahan dan dijadikan areal kebun kelapa sawit.
Untuk mengatasi kegiatan-kegiatan yang dapat mengancam keberadaan satwaliar khususnya mamalia dan habitatnya, seperti: penebangan kayu secara
illegal, perburuan liar dan perambahan hutan pihak taman nasional telah
memberikan pencegahan, salah satunya adanya pengumuman berupa papan peringatan tentang larangan yang dapat merusak kawasan hutan taman nasional (Gambar 13). Namun kenyataan di lapangan kerusakan tetap terjadi. Untuk itu perlu dilakukan kegiatan yang dapat menumbuhkan kesadaran masyarakat akan pentingnya kelestarian hutan di TNTN.
(a) (b)
VI. KESIMPULAN DAN SARAN
6.1. Kesimpulan
Berdasarkan hasil penelitian mengenai studi keanekaragaman jenis mamalia besar pada areal TNTN yang berbatasan dengan kebun kelapa sawit PT. Inti Indosawit Subur Ukui, Kabupaten Pelalawan, Propinsi Riau dapat disimpulkan sebagai berikut:
1. Jenis mamalia besar yang ditemukan di TNTN sebanyak 14 jenis dengan 11 jenis ditemukan secara langsung dan 3 jenis lainnya hanya ditemukan secara tidak langsung. Keempatbelas jenis ini termasuk kedalam 5 bangsa (ordo), yaitu Artiodactyla, Primata, Carnivora, Perissodactiyla, dan Proboscidea. 2. Jenis mamalia besar yang ditemukan di TNTN memiliki pola sebaran yang
mengelompok dan acak. Jenis yang memiliki sebaran yang mengelompok, yaitu Monyet ekor panjang, Lutung budeng, dan Lutung simpai, sedangkan jenis yang memiliki sebaran acak, yaitu Babi hutan, Owa ungko, Siamang, Pelanduk, Kancil, Rusa sambar, Musang akar, dan Tapir.
6.2. Saran
1. Perlu dilakukan pengamanan pada areal perbatasan TNTN khususnya dengan perkebunan kelapa sawit PT. Inti Indosawit Subur dengan gangguan terbesar pada jalur VI dan VII.
2. Perlu dilakukannya kegiatan inventarisasi serta berkelanjutan oleh pihak taman nasional untuk mengetahui perubahan-perubahan yang terjadi pada keanekaragaman jenis mamalia besar di TNTN.
3. Kebijakan pihak TNTN dalam penentuan zonasi kawasan perlu memperhatiakan jenis-jenis mamalia secara umum dan satwa yang dilindungi secara khususnya.
PADA AREAL KAWASAN TAMAN NASIONAL TESSO NILO
YANG BERBATASAN DENGAN KEBUN KELAPA SAWIT
PT. INTI INDOSAWIT SUBUR UKUI, KABUPATEN
PELALAWAN PROPINSI RIAU
FEBI MURYANTO
DEPARTEMEN
KONSERVASI SUMBERDAYA HUTAN DAN EKOWISATA
FAKULTAS KEHUTANAN
INSTITUT PERTANIAN BOGOR
DAFTAR PUSTAKA
Alikodra HS. 1990. Pengelolaan Satwaliar Jilid I. Departeman Pendidikan dan Kebudayaan. Direktorat Jenderal Pendidikan Tinggi. PAU Ilmu Hayat. Institut Pertanian Bogor. Hal: 31-54.
Alikodra HS. 2002. Pengelolaan Satwaliar Jilid 1. Yayasan Penerbit Fakultas Kehutanan IPB. Bogor.
BAPPENAS. 2003. Strategi dan Rencana Aksi Keanekaragaman Hayati Indonesia 2003-2020. Jakarta: BAPPENAS.
Bourliere F. 1975. Ecology of Population. Macmillan Publishing Co, Inc. New York.
Departemen Kehutanan. 2004. 50 Taman Nasional di Indonesia. Jakarta. Dephut. Effendi M. 1985. Studi Pengaruh Pembalakan Mekanis Sistem TPI terhadap
Keberadaan 10 Jenis Mamalia Hutan Hujan Tropika di Teluk Bangkurat Lampung. Jurusan Konservasi Sumberdaya Hutan. Fakultas Kehutanan. Institut Pertanian Bogor. Bogor.
Entebe RF. 2005. Penyebaran Mamalia Kecil pada Hutan Sisa (Remnant Forest) di Suaka Margasatwa Balairaja Propinsi Riau. Tesis. Institut Pertanian Bogor. Bogor.
Faldhamer GA, LC Drickamer, SH Vessey and JF Merritt. 1999. Mammalogy: Adaptation, Diversity and Ecology. Boston: McGraw-Hill.
Folke C, CS Holling and C Perrings. 1996. Biological diversity, ecosystem and the human scale. Ecological Application 6(4): 1018-1024.
Haryanto. 1987. Priliminary Study on The Habitat of Mueller’s gibbon (Hylobates meulleri) in Bukit Soeharto Protection Forest East Kalimantan dalam Studi Penelitian Hutan Tropika Basah. Fakultas Kehutanan. Institut Pertanian Bogor. Bogor.
Jasin M. 1992. Zoologi Vertebrata untuk Perguruan Tinggi. Sinar Wijaya. Surabaya.
Krebs CJ. 1989. Ecological Methodology. New York: Harper & Row, Publisher. Kurnia I, H Fadly, U Kusdinar, WG Gunawan, DW Idaman, RS Dewi, D Yandhi,
Lekagul B. and J.A. McNeely. 1997. Mammals of Thailand. Printed Under The Unsices of The Assosiaton of Wildlife. pp 278-281
Ludwig JA and JF Reynolds. 1988. Statistical Ecology: A primer on methods and computing. New York: John wilwy & Sons.
Lumme AL. 1994. Pengaruh Penebangan Terhadap Keanekaragaman Satwaliar Studi Kasus di Areal HPH di Propinsi Sulawesi Tengah. Skripsi Jurusan Konservasi Sumberdaya Hutan. Fakultas Kehutanan. Institut Pertanian Bogor. Bogor.
Magurran AE. 1998. Ecological Diversity and Its Measurement. Croom Helm. New South Wales.
Noerdjito M, I Maryanto, SN Prijono, EB Waluyo, R Ubaidillah, Mumpuni, AH Tjakrawidjaja, RM Marwoto, Heryanto, WA Noerdjito dan H Wiriadinata. 2005. Kriteria Jenis Hayati yang Harus Dilindungi Oleh dan Untuk Masyarakat Indonesia. Bogor: Pusat Penelitian Biologi-LIPI & World Agroforestry Centre-ICRAF.
Odum EP. 1997. Dasar-dasar Ekologi. Terjemahan Edisi ke-3. Yogyakarta: Gajah Mada University Press.
Payne J. CM. Francis, K Phillips dan SN Kartikasari. 2000. Panduan Lapangan: Mamalia di Kalimantan, Sabah, Serawak dan Brunei Darussalam. Jakarta: Prima Centra Indonesia. 386 Hal.
Primarck RB, J Supriatna, M Indrawan dan P Kramadibrata. 1998. Biologi Konservasi. Jakarta: Yayasan Obor Indonesia. 345 hal.
Santosa Y. 1993. Strategi Kuantitatif untuk Pendugaan Beberapa Parameter Demografi dan Kuota Pemanenan Populasi Satwaliar berdasarkan Pendekatan Ekologi Perilaku: Studi Kasus Terhadap Populasi Kera Ekor
Panjang (Macaca fasicularis) di Pulau Tinjil. Proyek Peningkatan
Penelitian dan Pengabdian pada Masyarakat. Fakultas Kehutanan. Institut Pertanian Bogor. Bogor.
Santosa N. 2002. Analisis habitat dan potensi pakan Monyet ekor panjang (Macaca fasicularis, Raffles) di Pulau Tinjil. Media konservasi 5 (1): 5-9 Soerianegara I dan A Indrawan. 2002. Ekologi Hutan Indonesia. Bogor:
Laboratorium Ekologi Hutan Fakultas Kehutanan Institut Pertanian Bogor. Suhartono T dan A Mardiastuti. 2003. Pelaksanaan Konvensi CITES di Indonesia.
Jakarta: Japan Internasional Cooperation Agency (JICA). 317 hal.
Toth F and J Kiss. 1999. Comparative analysis of epigeic spider assemblages in Northern Hungarian winter weat fields and ther adjeacent margins. The Jounal of Arachinology 27.
Vaughan TA. 1986. Mammalogy. 3rd Edition. Florida: Holt, Rinerhart and Winston, Inc.
Vieira EM and ELAM Filho. 2003. Vertical stratification of small mammals in the Atlantic rainforest of South-Eastern Brazil. Journal of Tropical Ecology 19: 501-507.
Yusuf M. 1998. Studi Keragaman dan Kelimpahan Jenis Burung Dan Mamalia Pada Beberapa Areal Bekas Tebangan dan Hutan Primer di Areal HPH PT. Narkata Rimba, Kalimantan Timur. Tesis. Institut Pertanian Bogor. Bogor. Zorenko T and T Leontyevan. 2003. Spesies diversity and distribution of
mammals in Riga. Acta Zoological Lituania 13(1).
PADA AREAL KAWASAN TAMAN NASIONAL TESSO NILO
YANG BERBATASAN DENGAN KEBUN KELAPA SAWIT
PT. INTI INDOSAWIT SUBUR UKUI, KABUPATEN
PELALAWAN PROPINSI RIAU
FEBI MURYANTO
DEPARTEMEN
KONSERVASI SUMBERDAYA HUTAN DAN EKOWISATA
FAKULTAS KEHUTANAN
INSTITUT PERTANIAN BOGOR
PADA AREAL KAWASAN TAMAN NASIONAL TESSO NILO
YANG BERBATASAN DENGAN KEBUN KELAPA SAWIT
PT. INTI INDOSAWIT SUBUR UKUI, KABUPATEN
PELALAWAN PROPINSI RIAU
Skripsi
Sebagai Salah Satu Syarat untuk Memperoleh Gelar
Sarjana Kehutanan pada Fakultas Kehutanan
Institut Pertanian Bogor
FEBI MURYANTO
DEPARTEMEN
KONSERVASI SUMBERDAYA HUTAN DAN EKOWISATA
FAKULTAS KEHUTANAN
INSTITUT PERTANIAN BOGOR
Kawasan Taman Nasional Tesso Nilo yang Berbatasan dengan Kebun Kelapa Sawit PT. Inti Indosawit Subur Ukui, Kabupaten Pelalawan, Propinsi Riau
Nama : Febi Muryanto
NRP : E 34103088
Menyetujui: Dosen Pembimbing,
Dr. Ir. A. Machmud Thohari, DEA NIP : 130 891 377
Mengetahui:
Dekan Fakultas Kehutanan IPB,
Dr. Ir. Hendrayanto, M. Agr. NIP : 131 578 788
Alhamdulillahirabbil ‘alamin. Segala Puji bagi Allah SWT atas segala rahmat dan kasih sayang-Nya, yang telah memilih kita sebagai ummat Sayyidina
Muhammad saw, dan menyelamatkan kita dari Gelapnya Kebodohan dan Kehinaan Dosa, Segala Puji bagi Allah Yang telah memberi kita Hidayah, sehingga penelitian ini dapat diselesaikan dengan Rahmat dan Kemudahan dari-Nya SWT. Sholawat serta salam semoga senantiasa tetap tercurahkan kepada Nabi Muhammad saw beserta keluarga, para sahabatnya, serta para pengikutnya yang tetap setia dan tetap istiqomah dalam mengikuti semua perjalanannya.
Skripsi ini berjudul Studi Keanekaragaman Jenis Mamalia Besar Pada Areal Kawasan Taman Nasional Tesso Nilo yang Berbatasan dengan Kebun Kelapa Sawit PT. Inti Indosawit Subur Ukui, Kabupaten Pelalawan, Propinsi Riau, disusun sebagai salah satu syarat untuk memperoleh gelar Sarjana Kehutanan pada Departemen Konservasi Sumberdaya Hutan dan Ekowisata Fakultas Kehutanan Institut Pertanian Bogor. Skripsi ini menggambarkan jenis satwaliar yang ada di Kawasan Taman Nasional Tesso Nilo, informasi mengenai perambahan dan konversi lahan yang terjadi.
Penulis menyadari keterbatasan dan kekurangan yang terdapat dalam tulisan ini karena itu masukan, kritikan dan saran dari pembaca sangat penulis harapkan untuk penyempurnaan tulisan ini. Penulis juga menyampaikan ucapan terima kasih sebesar-besarnya kepada Bapak Dr. Ir. A. Macmud Thohari, DEA sebagai dosen pembimbing yang telah memberikan masukan dan saran terhadap penulisan skripsi ini. Selanjutnya penulis juga menghaturkan terima kasih kepada pihak PT. Inti Indosawit Subur, atas berkenannya memberikan fasilitas penunjang penelitian. Akhir kata, semoga tulisan ini dapat bermanfaat serta menjadi pendorong bagi penulis untuk mengkaji dan menggali lebih dalam pengetahuan yang telah diperoleh.
Bogor, Mei 2009
Pada kesempatan ini penulis mengucapkan terima kasih, kepada:
1. Ayahanda dan Ibunda tercinta atas segala kasih sayang dan do’anya yang tidak akan pernah habis untuk dipanjatkan, kakak-kakak ku (Yudha Yudhianto, Rita Susanti dan keluarga, Tati Haryanti dan keluarga), serta seluruh keluarga. 2. Dr. Ir. A. Machmud Thohari, DEA yang telah memberikan bimbingan,
dorongan dan saran hingga selesainya penyusunan skripsi ini.
3. Prof. Dr. Ir. Cecep Kusmana, MS. selaku dosen penguji wakil dari Departemen Silvikultur dan Prof. Dr. Ir. Fauzi Febrianto, MS. selaku dosen penguji wakil dari Departemen Hasil Hutan atas saran, masukan dan wawasan yang diberikan.
4. Segenap pimpinan dan staf PT. Inti Indosawit Subur atas segala kerjasama yang diberikan.
5. Balai Taman Nasional Tesso Nilo, Polhut, dan PEH TNTN atas bantuannya selama penelitian di lapangan.
6. Ir. H. Didi Suharyadi (E 20_) dan keluarga atas segala kasih sayang yang diberikan baik moral maupun spiritual juga sehingga penulis bisa dengan baik menyelesaikan studi di KSHE IPB.
7. Staf dan Dosen-dosen Departemen Konservasi Sumberdaya Hutan dan Ekowisata.
8. Rekan-rekan Konservasi Sumberdaya Hutan dan Ekowisata’40, Manajemen Hutan’40, Teknologi Hasil Hutan’40, dan Silvikultur’40 atas kebersamaannya. 9. Keluarga Besar Asrama Mahasiswa IPB ”Sylvasari” 40 (M. Ramli, Agus R.,
Andi I, Mujinius J., Feri I., Jati S., Sandrio I., Bhakti A., Romadoni A., Sansan S., Ade W.) atas segala kebersamaan baik suka maupun duka.
10.Sahabat-sahabat ku SDN Sukatani V, SMP N 4 Cimanggis dan SMU PGRI Cibinong.
11.Rekan-rekan seperjuangan di Kutai Kertanegara.
Penulis dilahirkan di Jakarta pada hari Kamis tanggal 7 Februari 1985, putra keempat dari empat bersaudara, dari pasangan Bapak Sudiono dan Ibu Winarsih. Pendidikan formal penulis dimulai di SD Negeri Sukatani V pada tahun 1990 dan lulus pada tahun 1996 kemudian melanjutkan studi di SLTP Negeri 4 Cimanggis dan lulus pada tahun 1999. Penulis melanjutkan ke SMU PGRI Cibinong dan lulus pada tahun 2002. Pada tahun 2003 penulis diterima di Fahutan IPB Dept. KSH melalui Seleksi Penerimaan Mahasiswa Baru (SPMB).
Pada tahun 2006 penulis telah melaksanakan Praktek Pengenalan Hutan di KPH Banyumas Barat dan KPH Banyumas Timur dan Praktek Pengelolaan Hutan dikampus Getas UGM gelombang II. Pada bulan Februari-April 2007 penulis melakukan kegiatan Praktek Kerja Lapang Profesi (PKLP) di Taman Nasional Lore Lindu Propinsi Sulawesi Tengah.
Selama masa studi penulis aktif di BEM Fahutan IPB pada tahun 2004-2005 sebagai anggota Dept. Kom Info. Pada tahun yang sama di TMPLLK (Tim Mahasiswa Peduli Lingkungan Lingkar Kampus) IPB sebagai staf Lingkungan dan sebagai Sekretaris Asrama Mahasiswa IPB Sylvasari. Pada tahun 2005-2006 sebagai Ketua Komisi A di DPM Fahutan IPB dan Sebagai Wakil Ketua Asrama Mahasiswa IPB Sylvasari. Pada tahun 2006-2007 penulis diamanahkan sebagai Ketua Asrama Mahasiswa IPB Sylvasari. Pada tahun 2007-2008 penulis juga diamanahkan sebagai Koordinator Penanaman BSCA (Bumi Sengon Cahaya Alam) di Kecamatan Cipanas, Kabupaten Cianjur, Propinsi Jawa Barat. Selain itu penulis juga terpilih sebagai finalis dalam Pekan Ilmiah Mahasiswa Nasional (PIMNAS) XXI di Semarang tahun 2008.