RESISTENSI PISANG AMPYANG
(
Musa acuminata,
AAA, subgrup non-Cavendish)
TERHADAP
Fusarium oxysporum
f.sp.
cubense
HASIL
MUTASI INDUKSI DAN SELEKSI
IN VITRO
RENI INDRAYANTI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
i
RESISTENSI PISANG AMPYANG
(
Musa acuminata,
AAA, subgrup non-Cavendish)
TERHADAP
Fusarium oxysporum
f.sp.
cubense
HASIL
MUTASI INDUKSI DAN SELEKSI
IN VITRO
RENI INDRAYANTI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ii
Dengan ini saya menyatakan bahwa disertasi Resistensi Pisang Ampyang
(Musa acuminata, AAA, subgrup non-Cavendish) terhadap Fusarium oxysporum
f.sp. cubense Hasil Mutasi Induksi dan Seleksi In vitro adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Desember 2011
Reni Indrayanti
iii
banana cv. Ampyang (Musa acuminata, AAA, subgroup non-Cavendish) through induced mutation and in vitro selection. Under supervised by SUDARSONO, NURHAJATI ANSORI MATTJIK, ASEP SETIAWAN.
Fusarium wilt caused by Fusarium oxysporum Schlecht f.sp. cubense (EF Smith) Snyd & Hans (Foc) is one of the most important disease infecting root of banana and plantain (Musa spp.). This disease reported has become a primary constrain in banana production and lost of some banana germplasm, including in banana cv. Ampyang. Chemical control of this disease is economically impractical, and to widen banana genetic diversity is also a difficult task because most of edible banana are either triploid, male sterility and partenocarp. Therefore conventional breeding approach for resistance to Fusarium wilt for this crop is difficult. The objectives of this research were (1) to increase phenotypic diversity of banana cv. Ampyang through induced mutation; (2) to obtain banana cv. Ampyang resistant to Fusarium wilt. Explants of in vitro grown shoots were exposed to gamma irradiation at 0, 20, 25, 30, 35, 40, 45, 50 Gy to determine its the radiosensitivity. The CurveExpert ver. 1.4 analysis results indicated that lethal doses of irradiation reducing 20% to 50% of shoot growth (LD20-50) were 51.07-64.54 Gy. After 10
months of proliferation and regeneration periods, all regenerated plantlets from irradiated explants produced less numbers of roots, and some of regenerated plantlets, showed significantly less plantlet fresh weight and height, and longer leaves. Glasshouse evaluation indicated that banana plant regenerated from gamma irradiated explants to exhibit lower plant height, shorter leaf, and leaf lenght by leaf width ratio than from non-irradiated ones. Population of banana cv. Ampyang gamma irradiated at 30, 40, 45 and 50 Gy showed wider variation in qualitative characters than that treated with 0 Gy. Field evaluation showed that a number of variants (12 clones) regenerated from gamma irradiated 25, 30, 45, 50 Gy and non-irradiated one identifying as a positive variant. This charachterize were based on agronomyc charachter and quality of fruit. Improvement program to banana cv. Ampyang resistant to Fusarium wilt requiring some previous study in virulences and effectiveness Foc of Banyuwangi isolate as an selective agent. This research showed that Foc ofBanyuwangi isolate were virulent to banana cv. Ampyang, and culture filtrate of Foc concentration 40-60% (v/v) was an effective concentration to obtain banana shoots insensitive to culture filtrate of Foc. This experiment also showed that inoculation methods by dipping banana roots for 2 hours into Foc
conidial suspensión 2.5 x 107 conidial mL-1 is the most effective methods to identifying banana cv. Ampyang resistant to Fusarium wilt. Severalbanana clones resistance to Fusarium wilt was obtain. Induced mutation followed by in vitro
selection procedures found that 48 clones (40.7%) indicated resistant to
Fusarium wilt, most of them originated from gamma irradiated explants of 30 and 50 Gy. Glasshouse evaluation of banana variant regenerated from induced mutation without in vitro selection produced 5 clones (13.8%) and evaluation of banana variant in endemic Fusarium wilt found that 5 clones (4.9%) indicated resistant to Fusarium wilt.
iv
subgrup non-Cavendish) terhadap Fusarium oxysporum f.sp. cubense hasil Mutasi Induksi dan Seleksi In vitro. Dibimbing oleh SUDARSONO, NURHAJATI ANSORI MATTJIK, ASEP SETIAWAN
Pisang dan plantain (Musa spp) merupakan komoditas utama untuk ketahanan pangan, walaupun dalam kondisi lingkungan yang buruk tanaman pisang dapat berperan sebagai tanaman pangan yang bermanfaat. Pisang terdiri dari banyak species dan kultivar, diantaranya pisang cv. Ampyang (Musa acuminata, AAA, subgrup non-Cavendish). Pisang cv. Ampyang merupakan pisang meja (dessert banana) yang sudah sulit dijumpai di pasar tradisional maupun modern, diduga pisang ini sudah jarang dibudayakan oleh petani lokal. Keberadaan pisang cv. Ampyang yang sulit dijumpai ini diduga karena pisang cv. Ampyang rentan terhadap penyakit layu Fusarium yang disebabkan oleh cendawan Fusarium oxysporum f.sp. cubense (Foc). Tujuan umum penelitian ini adalah untuk (1) meningkatkan keragaman tanaman pisang (Musa acuminata, AAA) cv. Ampyang melalui teknik mutasi induksi dengan iradiasi gamma secara in vitro; (2) mendapatkan klon-klon dari kultivar pisang (Musa acuminata, AAA) cv. Ampyang yang resistensi terhadap layu Fusarium yang disebabkan oleh cendawan Fusarium oxysporum f.sp. cubense.
Percobaan tahap awal dilakukan untuk menentukan radiosensitivitas tunas pisang cv Ampyang (Musa acuminata, AAA) terhadap perlakuan iradiasi gamma yang ditentukan berdasarkan dosis yang mereduksi pertumbuhan tanaman sebesar 20-50% (LD20-50), dan LD20-50 berada dikisaran 51.07-64.54 Gy dengan dosis
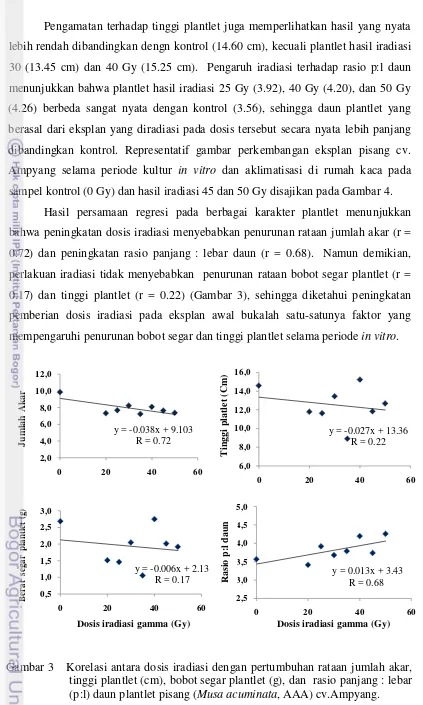
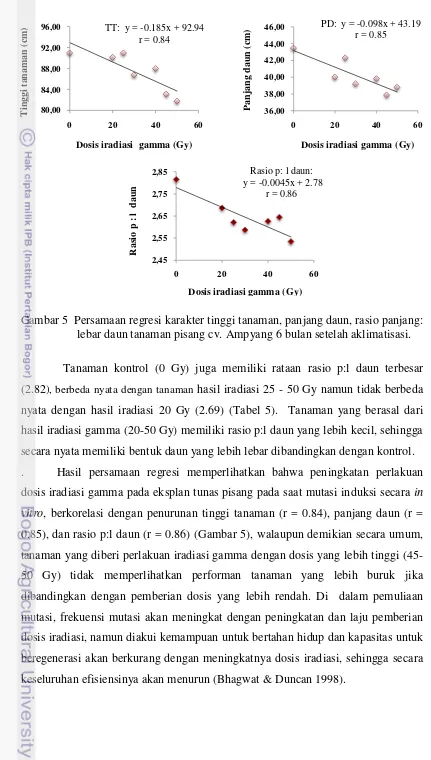
optimum untuk pertumbuhan tunas pisang aseptis berada pada dosis 30 Gy. Secara fenotipik tunas yang diradiasi cenderung menghasilkan plantlet dengan keragaman karakter kuantitatif (jumlah akar, tinggi plantlet, bobot segar plantlet) yang lebih rendah, dan bentuk daun yang lebih panjang dari tanaman kontrol. Evaluasi di rumah kaca menunjukkan, tanaman yang diradiasi dengan dosis iradiasi yang lebih tinggi, menghasilkan rataan tinggi tanaman dan panjang daun yang makin rendah,. Tanaman yang diradiasi memiliki bentuk daun yang cenderung lebih lebar dari tanaman kontrol. Tanaman yang diradiasi dengan dosis diatas 30, 40, 45 dan 50 Gy) menghasilkan jenis varian karakter kualitatif yang lebih banyak dibandingkan dengan dosis yang lebih rendah. Evaluasi keragaman terhadap densitas stomata memperlihatkan bahwa tanaman kontrol dan tanaman hasil iradiasi 45 Gy memiliki densitas stomata yang nyata lebih tinggi dari tanaman 20, 25 dan 30 Gy.
v
menghasilkan buah dengan karakteristik lebih rendah dari tanaman lainnya. Tanaman kontrol (0 Gy), 30 dan 50 Gy memperlihatkan konsistennya dalam kemampuan tumbuh dan menghasilkan buah dengan kuantitas dan kualitas yang cukup baik. Beberapa tanaman teridentifikasi memiliki bentuk buah yang unik, sedangkan tanaman yang tidak diradiasi pada umumnya memiliki bentuk buah yang lebih besar. Tanaman-tanaman dengan karakter yang lebih unggul dari perlakuan lainnya, memberi dugaan bahwa beberapa sel-sel atau jaringan yang terkena iradiasi dan tidak mengalami kematian, merupakan mutan hasil iradiasi gamma yang positif, sedangkan tanaman yang tidak diradiasi (0 Gy) diduga karena faktor subkultur berulang.
Usaha perolehan tanaman pisang cv. Ampyang dengan karakter resisten terhadap layu Fusarium pada percobaan ini dilakukan secara in vitro dan ex vitro. Pendekatan pertama melalui melalui teknik mutasi induksi yang dilanjutkan dengan seleksi in vitro. Pendekatan kedua teknik mutasi induksi tanpa tahapan seleksi in vitro. Bibit varian yang diperoleh langsung diinfeksi dengan konidia Foc, atau di evaluasi sifat ketahanannya di lahan endemik layu Fusarium.
Evaluasi di rumah kaca membutuhkan pengujian virulensi dan patogenitas mycelium F. oxysporum f.sp. cubense (Foc) isolat Banyuwangi (Bw), dan efektivitas teknik inokulasi dan kerapatan suspensi konidia Foc yang mampu menginfeksi tanaman pisang cv. Ampyang. Hal ini dilakukan karena Foc isolat Bw belum diketahui virulensinya terhadap pisang meja. Pengujian virulensi Foc
isolat Bw terhadap tunas pisang Ampyang aseptis dengan metode kultur ganda-1 (dual culture method), membuktikan bahwa Foc isolat Bw virulen terhadap pisang cv. Ampyang. Persentase kematian plantlet sebesar 100% dengan gejala awal perubahan warna merah-kecoklatan yang dimulai dari batang dan daun bagian bawah, yang merupakan ciri tanaman terinfeksi Foc. Evaluasi dan efektivitas metode uji untuk ketahanan pisang cv. Ampyang terhadap layu Fusariumdi rumah kacamemperlihatkan bahwa tanaman yang diinokulasi dengan Foc kerapatan 2.5 x 107 konidia mL-1 melalui metode perendaman akar, lebih efektif dalam menimbulkan kerusakan dibandingkan dengan metode injeksi batang. Hal ini diperlihatkan dengan gejala kelayuan dan kerusakan bibit yang nyata lebih besar, serta jumlah daun, panjang akar, dan ketebalan daun yang lebih rendah, sehingga memberi gambaran bahwa bibit tanaman yang diinokulasi mengalami penghambatan pertumbuhan akibat terhambatnya aliran air pada jaringan vaskuler tanaman akibat infeksi Foc.
vi
tunas varian yang diseleksi ternyata diperoleh pada media selektif mengandung FK
Foc 60% (v/v). Sejumlah tunas (1695 tunas) yang mampu bertahan hidup dalam kondisi selektif dengan konsentrasi tinggi ini dikategorikan sebagai tunas-tunas insensitif FK Foc 60%, walaupun pada konsentrasi ini belum dapat dibedakan tunas varian yang tahan atau rentan. Tunas-tunas insensitif FK Foc hasil seleksi in
vitro ini secara teoritis merupakan varian yang mampu menginduksi atau
mengaktifkan gen-gen ketahanannya terhadap layu Fusarium, atau menginduksi enzim-enzim yang berperan dalam mendetoksifikasi toksin cendawan.
Regenerasi tunas-tunas insensitif FK Foc dalam media perakaran mengandung 6-benzyladenin 2.25 mg L-1 dan Indole-3-acetic acid 0.175 mg L-1 selama 2 bulan, menghasilkan plantlet-plantlet yang secara fenotipik terlihat sehat dan segar, mampu membentuk akar dan tunas baru. Persentase kemampuan hidup terbesar dihasilkan oleh plantlet yang berasal dari hasil iradiasi 30 Gy (56.2%). Hasil ini tidak jauh berbeda dengan kemampuan tumbuh tunas pisang cv. Ampyang berdasarkan kurva reduksi pertumbuhan pada siklus vegetatif pertama, yaitu kemampuan bertahan hidup yang lebih besar pada tunas pisang yang diradiasi dengan dosis 30 Gy. Aklimatisasi plantlet hasil seleksi in vitro pada percobaan ini secara tidak langsung telah menyeleksi tanaman-tanaman varian yang rentan terhadap penyakit layu Fusarium. Aklimatisasi sebanyak 499 plantlet insensitif FK
Foc menghasilkan 118 (23.7%) bibit varian tanaman pisang cv. Ampyang yang mampu beradaptasi dan bertahan hidup 60 hari setelah aklimatisasi.
Evaluasi ketahanan bibit varian yang diperoleh dari hasil mutasi induksi yang dilanjutkan dengan seleksi in vitro secara bertingkat, diperoleh 48 (40.7%) bibit tanaman dari 118 bibit yang diinfeksi dengan konidia Foc isolat Medan VCG 01213/16 Tropical Race 4, atau Foc isolat Banyuwangi dengan kerapatan 2.5 x 107 konidia mL-1 memiliki karakteristik tahan layu Fusarium. . Bibit varian dengan karakteristik tahan layu Fusarium hasil induksi mutasi di rumah kaca diperoleh 5 klon tanaman (13.8%) dari 36 bibit yang dievaluasi, dan di lapangan diperoleh 5 klon anakan (4.9%) dari 102 tanaman yang dievaluasi, yang mampu bertahan hidup sampai 24 bulan setelah tanam pada lahan endemik Foc. Bibit varian yang mengindikasikan resisten layu Fusarium tersebut banyak dijumpai dari varian yang berasal dari hasil iradiasi 30 dan 50 Gy. Hasil percobaan ini memberikan kesimpulan bahwa peluang untuk mendapatkan bibit varian tanaman resisten terhadap layu Fusarium lebih besar diperoleh melalui teknik mutasi induksi secara
in vitro yang dilanjutkan dengan seleksi in vitro secara bertingkat, walaupun prosedur ini relatif membutuhkan waktu pekerjaan yang lebih lama
Bibit pisang cv. Ampyang yang tahan terhadap layu Fusarium hasil seleksi
in vitro ini berpotensi digunakan sebagai populasi untuk di evaluasi kembali sifat ketahanannya di lahan yang terserang cendawan F. oxysporum f.sp. cubense, untuk memastikan sifat ketahanannya dan mengidentifikasi varian tahan layu Fusarium
vii
© Hak cipta milik Institut Pertanian Bogor, tahun 2012 Hak cipta dilindungi
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebut sumber.
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tujuan suatu masalah. b. Pengutipan tidak merugikan kepentingan yang wajar IPB
viii
RESISTENSI PISANG AMPYANG
(
Musa acuminata,
AAA, subgrup non-Cavendish)
TERHADAP
Fusarium oxysporum
f.sp.
cubense
HASIL
MUTASI INDUKSI DAN SELEKSI
IN VITRO
RENI INDRAYANTI
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Agronomi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ix
Penguji pada Ujian Tertutup: Dr. Ir. Endang Nurhayati MSc. Dr. Sintho W. Ardie SP, MSi
x
Mutasi Induksi dan Seleksi in vitro
Nama Mahasiswa : Reni Indrayanti
NRP : A161060031
Program Studi : Agronomi
Disetujui: Komisi Pembimbing
Ketua
Prof. Dr. Ir. Sudarsono MSc
Prof. Dr. Ir. Nurhajati A. Ansori M. MS
Anggota I Anggota II
Dr. Ir. Asep Setiawan MS
Diketahui
Ketua Program Studi Agronomi Dekan Sekolah Pascasarjana
Prof. Dr. Ir. Munif Ghulamahdi MS Dr. Ir. Dahrul Syah MScAgr
xi
Puji syukur penulis panjatkan kepada Allah SWT atas segala rizkiNya sehingga disertasi ini berhasil diselesaikan. Penelitian yang dilaksanakan sejak September 2007 hingga Juni 2011 berjudul: Resistensi Pisang Ampyang (Musa acuminata, AAA, subgrup non-Cavendish) terhadap Fusarium oxysporum f.sp.
cubense hasil Mutasi Induksi dan Seleksi In vitro.
Terima kasih penulis sampaikan kepada Bapak Prof. Dr. Ir. Sudarsono MSc selaku ketua komisi pembimbing, Ibu Prof. Dr. Ir. Nurhajati Mattjik MSi dan Bapak Dr. Ir. Asep Setiawan MSc selaku anggota komisi pembimbing yang telah banyak memberi saran dan bimbingan sejak persiapan, pelaksanaan penelitian hingga selesainya disertasi ini. Ir. Y. Supriati MS dari Balai Besar Bioteknologi dan Sumber Daya Genetik Bogor, Dr. Ir. RD Purwati MSc dari Balitas Malang, Ir. A. Sutanto MSc dari Balitbu Solok. Staf dan teknisi pada Laboratorium Biologi Molekuler Tanaman IPB dan Laboratorium Ekofisologi IPB, analis pada Pusat Penelitian Sumberdaya Hayati dan Bioteknologi LPPM IPB, serta staf pada Laboratorium Kultur Jaringan Tanaman UNJ. Kepada Rektor Universitas Negeri Jakarta atas dukungan biaya studi, dan Direktur Penelitian dan Pengabdian kepada Masyarakat (DP2M) Dikti Kementerian Pendidikan Nasional yang telah memberikan dukungan biaya penelitian.
Terima kasih penulis sampaikan kepada rekan-rekan sahabat di Program Studi Agronomi dan Fitopatologi FAPERTA IPB angkatan 2006, kepada rekan-rekan di Laboratorum Biologi Molekuler Tanaman yang telah bersahabat dan berdiskusi selama penulis studi di IPB, kepada seluruh rekan dari jurusan Biologi FMIPA UNJ atas persahabatan dan dukungannya. Ungkapan terima kasih yang mendalam kepada suami Ir. Masyudin MM, dan anak-anakku M. Aditya Yudiantama, Afina Rahmah Widyanafi dan Hijran Luqman Harisha, serta seluruh keluarga yang telah memberikan doa dan kasih sayangnya. Kepada semua pihak yang telah memberikan bantuan selama penelitian dan penyusunan disertasi, penulis dengan segala kerendahan hati menyampaikan terima kasih semoga Allah SWT membalas dengan sebaik-baik balasanNya.
Bogor, Desember 2011
xii
Penulis dilahirkan di Jakarta pada tanggal 23 Oktober 1962 sebagai anak ke empat dari orang tua Sukardiono Windudipuro (alm) dan Sumartinah Tomopuspito (almh). Pendidikan sarjana ditempuh di Program Studi Biologi, Fakultas Pendidikan Matematika dan Ilmu Pengetahuan Alam, IKIP Jakarta lulus pada tahun 1984. Pendidikan Pascasarjana ditempuh pada tahun 1991 pada Program Studi Biologi, sub program studi Botani pada Program Pascasarjana IPB dan lulus pada tahun 1994. Kesempatan melanjutkan pendidikan Doktor dilaksanakan pada Program Studi Agronomi - Pemuliaan dan Bioteknologi Tanaman pada perguruan tinggi yang sama pada tahun 2006. Beasiswa pendidikan pascasarjana diperoleh melalui BPPS Dikti – Departemen Pendidikan Nasional. Penulis bekerja sebagai Staf Pengajar di Jurusan Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam pada Universitas Negeri Jakarta.
xiii
Halaman
DAFTAR TABEL ... xv
DAFTAR GAMBAR ... xviii
DAFTAR LAMPIRAN ..... xxii
BAB I PENDAHULUAN Latar Belakang ... 1
Pendekatan Masalah ... 4
Tujuan Umum Penelitian ... 5
Manfaat Penelitian ... Hipotesis ... 5 5 Ruang Lingkup Penelitian ... 6
BAB II TINJAUAN PUSTAKA Tanaman Pisang (Musa spp) ... 9
Cendawan F. oxysporum f.sp. cubense Penyebab Layu Fusarium ... 10
Kultur InVitro dan Variasi Somaklonal ... 15
Seleksi in vitro untuk Resistensi Tanaman Pisang terhadap Layu Fusarium ... 18
BAB III RADIOSENSITIVITAS PISANG cv. AMPYANG DAN POTENSI PENGGUNAAN IRADIASI GAMMA UNTUK INDUKSI VARIAN Abstrak ………...…… 19
Abstract ………...… 20
Pendahuluan ………... 20
Bahan dan Metode ………. 22
Hasil dan Pembahasan ……… 23
Simpulan ……… 31
Daftar Pustaka ……… 31
BAB IV EVALUASI KERAGAMAN FENOTIPIK PISANG cv. AMPYANG (Musa acuminata, AAA) HASIL MUTASI INDUKSI DENGAN IRADIASI GAMMA Abstrak ………... 35
Abstract ……….. 36
Pendahuluan ………... 36
Bahan dan Metode ………. 38
Hasil dan Pembahasan ……… 39
Simpulan ……… 62
xiv
PISANG cv. AMPYANG TERHADAP LAYU FUSARIUM
Abstrak ………...…… 65
Abstract ………...…... 66
Pendahuluan ………... 66
Bahan dan Metode ………. 68
Hasil dan Pembahasan ……… 70
Simpulan ……… 77
Daftar Pustaka ……… 77
BAB VI SELEKSI IN VITRO UNTUK MENDAPATKAN PLANTLET PISANG cv. AMPYANG HASIL IRADIASI GAMMA INSENSITIF FILTRAT KULTUR F. oxysporum f.sp. cubense Abstrak ………... 79
Abstract ……….. 80
Pendahuluan ………... 80
Bahan dan Metode ………. 82
Hasil dan Pembahasan ……… 84
Simpulan ……… 100
Daftar Pustaka ……… 100
BAB VII KETAHANAN VARIAN PISANG (Musa acuminata, AAA) cv. AMPYANG TERHADAP LAYU FUSARIUM Abstrak ………... 105
Abstract ……….. 106
Pendahuluan ………... 106
Bahan dan Metode ………. 108
Hasil dan Pembahasan ………. 110
Simpulan ……… 134
Daftar Pustaka ……… 135
BAB VIII PEMBAHASAN UMUM 139 BAB IX SIMPULAN DAN SARAN Simpulan ……… 149
Saran ……….. 151
DAFTAR PUSTAKA ……… 153
xv
Halaman
1. Rataan jumlah tunas pisang cv. Ampyang sebelum diradiasi (M0V0) dan
diproliferasi selama enam minggu (M1V1).
………
24
2. Rataan jumlah daun dan jumlah akar plantlet pisang cv. Ampyang hasil iradiasi gamma dan tanaman kontrol setelah diproliferasi dan diregenerasikan selama 10 bulan secara in vitro. ………..
27
3. Rataan bobot segar plantlet, tinggi plantlet, rasio panjang:lebar daun plantlet pisang cv. Ampyang hasil iradiasi gamma dan tanaman kontrol setelah diproliferasi dan diregenerasikan selama 10 bulan secara in vitro.
27
4. Karakter tinggi tanaman dan lingkar batang semu pisang cv. Ampyang hasil perlakuan iradiasi gamma usia 6 bulan setelah aklimatisasi. ...
40
5. Karakter panjang daun, lebar daun dan rasio panjang:lebar daun pisang cv. Ampyang hasil perlakuan iradiasi gamma pada usia 6 bulan
setelah aklimatisasi. ... .
40
6. Jenis varian pada tanaman pisang cv. Ampyang yang berasal dari hasil iradiasi gamma dan regenerasi secara in vitro berdasarkan karakter kuantitatif tanaman pada usia 6 bulan setelah aklimatisasi. ...
43
7. Jumlah tanaman dan persentase varian tanaman pisang cv. Ampyang hasil iradiasi gamma usia 6 bulan setelah aklimatisasi di rumah kaca.
44
8 Uji F rataan densitas stomata pada epidermis bawah daun ke-2 dan ke-3 tanaman pisang cv. Ampyang pada usia 8 bulan setelah aklimatisasi. ...
49
9. Rataan densitas stomata daun ke-3 pisang cv. Ampyang hasil iradiasi gamma usia 8 bulan setelah aklimatisasi. ...
49
10. Rataan tinggi tanaman dan lingkar batang semu pisang cv. Ampyang hasil iradiasi gamma dan regenerasi secara in vitro, pada usia 11 bulan setelah tanam di lapangan. ……….
51
11. Rataan panjang daun (cm), lebar daun (cm), dan rasio panjang : lebar daun pisang cv. Ampyang hasil iradiasi gamma dan regenerasi secara in vitro, pada usia 11 bulan di lapangan . ...
52
12. Rataan rasio panjang dan lebar daun, jumlah anakan pisang cv. Ampyang hasil iradiasi gamma dan regenerasi secara in vitro, pada usia 11 bulan setelah tanam di lapangan.
xvi
tanam di lapangan.. ………....……… 14. Pengamatan karakter kuantitatif buah pisang cv. Ampyang yang berasal dari hasil iradiasi gamma dan regenerasi secara in vitro saat panen usia 14-18 bulan setelah tanam di lapangan. ...
55
15. Hasil analisis proksimat buah pisang cv. Ampyang hasil perlakuan iradiasi gamma yang ditumbuhkan di lapangan. ...
59
16. Respon kerusakan tunas pisang cv. Ampyang (Musa acuminata, AAA) terhadap Foc isolat Banyuwangi. ...
71
17. Rataan skor gejala kelayuan bibit pisang dengan dua metode inokulasi dengan kerapatan suspensi konidia Foc 2.5 x 106 dan 2.5 x 107 kon mL-1, usia 30 hari setelah inokulasi. ...
72
18. Rataan skor gejala kelayuan bibit, jumlah daun, tinggi tanaman, panjang akar, ketebalan daun pisang bibit pisang hasil inokulasi dengan kerapatan suspensi konidia Foc 2.5 x 107 kon mL-1, usia 60 hari setelah inokulasi. ...
73
19. Skoring tingkat kerusakan dan persentase kematian tunas pisang cv. Ampyang pada media selektif mengandung filrat kultur Foc isolat Banyuwangi 2 bulan setelah inokulasi.. ...
87
20. Rataan jumlah tunas pisang cv. Ampyang pada media tanpa FK Foc
(0%) dan media selektif dengan FK Foc 30% usia 6 minggu setelah subkultur. ...
89
21. Rataan jumlah tunas pisang cv. Ampyang pada media selektif dengan FK Foc 30% dan FK Foc 40% , usia 6 minggu setelah subkultur. ....
91
22. Rataan jumlah tunas pisang cv. Ampyang pada media selektif dengan FK Foc 40% dan FK Foc 50%, usia 6 minggu setelah setelah subkultur.
93
23. Rataan jumlah tunas pisang cv. Ampyang pada media selektif dengan FK Foc 50% dan FK Foc 60% , usia 6 minggu setelah subkultur. …...
94
24. Jumlah total tunas varian pisang cv. Ampyang hasil iradiasi gamma yang tumbuh setelah dilakukan seleksi secara bertingkat dari media selektif mengandung FK Foc 30% ke media selektif mengandung FK
Foc 40%, 50% dan 60%. ...
95
26. Jumlah dan persentase tanaman varian pisang cv. Ampyang insensitif FK Foc 60% yang mampu bertahan hidup sampai usia 60 hari setelah aklimatisasi. ...
xvii
bulan setelah diinfeksi konidia Foc isolat Medan di rumah kaca ... 28. Identifikasi tanaman varian pisang cv. Ampyang usia 2 bulan setelah
infeksi. Persentase tanaman yang bertahan hidup, dan yang mengalami
recovery, usia 5 bulan setelah diinfeksi konidia Foc isolat Medan. ....
114
29 Ketahanan varian pisang cv. Ampyang hasil iradiasi gamma yang dilanjutkan dengan seleksi in vitro terhadap layu Fusarium, usia 2 bulan setelah diinfeksi konidia Foc isolat Banyuwangi di rumah kaca. ...
116
30. Identifikasi tanaman varian pisang cv. Ampyang, usia 2 bulan setelah infeksi. Persentase tanaman yang bertahan hidup, dan yang mengalami
recovery, usia 5 bulan setelah diinfeksi konidia Foc isolat Banyuwangi. 117
31. Ketahanan varian tanaman pisang cv. Ampyang hasil mutasi induksi dengan iradiasi gamma tanpa tahapan seleksi in vitro terhadap infeksi cendawan Foc isolat Banyuwangi usia 2 bulan setelah inokulasi di rumah kaca. ...
122
32. Rataan jumlah daun, tinggi tanaman (cm) dan panjang akar (cm) pada tanaman pisang cv. Ampyang usia 60 hari setelah recovery. ...
123
33. Tinggi tanaman dan lingkar batang semu varian tanaman pisang cv. Ampyang hasil iradiasi gamma usia 12 bulan setelah ditumbuhkan pada lahan endemik layu Fusarium di lapangan. ...
125
34. Jumlah anakan, jumlah pelepah, rasio panjang : lebar daun varian tanaman pisang cv. Ampyang hasil iradiasi gamma usia 12 bulan setelah ditumbuhkan pada lahan endemik layu Fusarium di lapangan..
125
35. Karakter kuantitatif buah pisang cv. Ampyang hasil iradiasi gamma pada lahan terserang penyakit layu Fusarium di lapangan. ...
131
36 Hasil analisis proksimat buah pisang cv. Ampyang hasil iradiasi gamma yang ditumbuhkan pada lahan terserang layu Fusarium. ...
132
37 Jumlah dan persentase tanaman varian pisang cv. Ampyang yang dapat bertahan hidup pada usia 12 dan 21 bulan setelah tanam pada lahan endemik penyakit layu Fusarium di lapangan ...
xviii
Halaman
1. Bagan alir strategis penelitian: (A) Induksi mutasi untuk meningkatkan keragaman dan ketahanan tanaman pisang cv. Ampyang; (b) Seleksi in vitro untuk mendapatkan klon pisang cv. Ampyang (Musa acuminata, AAA) resisten terhadap penyakit layu Fusarium. ...
7
2. Penentuan LD50 berdasarkan penghambatan proliferasi tunas aksilar
dari eksplan pisang cv. Ampyang yang diberi perlakuan iradiasi gamma. ....
24
3. Korelasi antara dosis iradiasi dengan pertumbuhan rataan jumlah akar, tinggi plantlet (cm), bobot segar plantlet (g), dan rasio panjang:lebar (p:l) daun plantlet pisang (Musa acuminata, AAA) cv.Ampyang. …….
28
4. Proliferasi tunas aksilar, regenerasi plantlet dan pertumbuhan bibit pisang cv. Ampyang, yang berasal dari eksplan hasil perlakuan iradiasi 0, 45 dan 50 Gy: (a) Eksplan tunas sebelum diradiasi, (b) Proliferasi tunas, (c) Regenerasi plantlet dan (d) Pertumbuhan bibit pisang paska aklimatisasi usia 2 bulan. ...
29
5. Persamaan regresi karakter tinggi tanaman, panjang daun, rasio panjang: lebar daun tanaman pisang cv. Ampyang 6 bulan setelah aklimatisasi ....
41
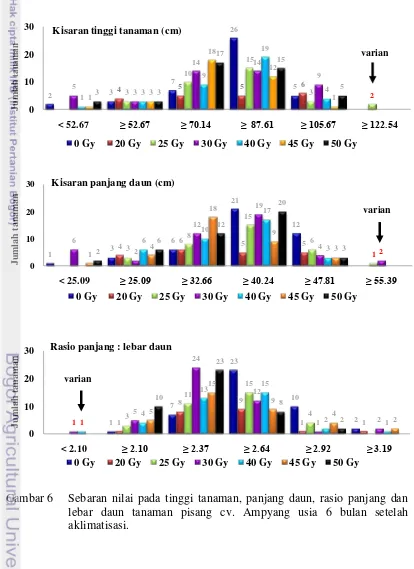
6. Sebaran nilai pada tinggi tanaman, panjang daun dan rasio panjang: lebar daun tanaman pisang cv. Ampyang usia 6 bulan setelah aklimatisasi ...
42
7. Representasi varian tanaman hasil iradiasi usia 6-8 bulan berdasarkan karakter kuantitatif dan kualitatif: (a) tinggi tanaman tertinggi dan terendah, (b) daun berkerut, (c) daun bergaris hijau tua-muda, (d) daun variegata, (e) susunan daun berbentuk melingkar (roset), (f) tepi daun menggulung, (g) daun tegak dengan ujung daun lancip, (h) bentuk daun tidak beraturan dan robek ...
45
8. Varian tanaman hasil iradiasi usia 6-8 bulan berdasarkan karakter kualitatif: (a) pelepah daun berhadapan, (b) pelepah daun menumpuk, (c) pelepah tersusun seperti kipas (d) pelepah daun berwarna merah ...
46
9. Keberadaan bercak pada daun; (a) daun tanpa bercak, (b) bercak sedikit dan (c) bercak banyak.. ...
46
11. Stomata daun pisang cv. Ampyang diamati pada usia 8 bulan setelah diaklimatisasi. (a) stomata daun tanpa diradiasi (0 Gy) dan (b) hasil iradiasi 20 Gy dengan perbesaran 40x, (c) stomata daun
xix
tanaman standar (normal), (b-d) tanaman dengan tinggi dan kualitas buah rendah dan kate, (e-f) variasi bentuk dan warna brachtea pada tanaman pisang dengan kuantitas buah 96-100 buah per tandan dan (g-i) kuantitas buah 38-57 buah per tandan pada usia 11-14 bulan setelah tanam. …... 13. Keragaman fenotipik buah yang dihasilkan dari tanaman yang berasal
dari hasil iradiasi gamma dan regenerasi secara in vitro (M1V6) yang
dikategerikan sebagai varian positif: (a-c) usia 11-12 bln setelah tanam; (d-h) saat panen usia 14-18 bln setelah tanam; (i) buah klon anakan hasil iradiasi 50 Gy usia panen 24 bln setelah tanam. ………...………
58
14. Karakteristik buah pisang cv. Ampyang setelah masak fisiologis: (a-b) Warna kecoklatan pada kulit buah dan warna orange tua daging buah pisang yang berasal dari hasil iradiasi gamma 25 Gy; (b) hasil iradiasi 45 Gy; (d-e) sisir buah pisang, buah dibelah membujur dan melintang yang berasal dari tanaman 0 Gy. .……….
60
15. Pertumbuhan dan perkembangan varian tanaman pisang cv. Ampyang yang berasal dari eksplan hasil iradiasi [45 Gy (4) 17] pada usia: (a) 2 bulan, (b) 4 bulan setelah aklimatisasi, (c-d) usia 18 bulan di lapangan, (e-f) karakteristrik bentuk buah sebelum dan setelah panen. ...
61
16. Plantlet pisang cv. Ampyang usia 2 bulan yang diinfeksi dengan mycelia Foc dengan teknik inokulasi (a) metode kultur ganda-1 (MKG-1) dan (b) metode kultur ganda-2 (MKG-2). ...
69
17. Gejala kelayuan tunas pisang terhadap Foc isolat Bw dengan teknik inokulasi (a) metode kultur ganda-1 dan (b) metode kultur ganda-2. ...
71
18. Gejala kelayuan bibit pisang hasil inokulsi Foc dengan: (a) metode injeksi batang semu kerapatan Foc 2.5 x 106 kon mL-1 dan (b) 2.5 x 107 kon mL-1 (c) metode perendaman akar kerapatan Foc 2.5 x 106 kon mL-1 dan (d) 2.5 x 107 kon mL-1, usia 60 hari setelah inokulasi. ...
74
19. Warna merah marone pada batang semu pisang hasil iokulasi Foc
dengan (a-b) metode injeksi batang dan (c) metode perendaman akar dengan kerapatan Foc 2.5 x 107 kon mL-1, 60 hari setelah inokulasi. ...
74
20. Potongan melintang daun tanaman pisang hasil inokulasi dengan suspensi konidia cendawan Foc kerapatan 2.5 x 107 konidia mL-1 melalui (a) metode perendaman akar dan (b) injeksi batang semu pada perbesaran 40x. ...
75
21 Fusarium oxysporum f.sp. cubense (Foc) isolat Banyuwangi: (a) Pertumbuhan cendawan dalam media PDA (b) filtrat kultur (FK)
xx
mengandung berbagai konsentrasi filtrat kultur Foc pada usia 2 bulan setelah tanam. ... 23. Respon pertumbuhan tunas pisang cv. Ampyang hasil iradiasi gamma
dalam media selektif mengandung filtrat kultur (FK) Foc 30% usia 6 minggu setelah subkultur. ...
89
24. Respon pertumbuhan tunas pisang cv. Ampyang hasil iradiasi gamma dalam media selektif mengandung filtrat kultur (FK) Foc 40%, usia 6 minggu setelah subkultur. ………
91
25. Respon pertumbuhan tunas pisang cv. Ampyang hasil iradiasi gamma dalam media selektif mengandung filtrat kultur (FK) Foc 50%, usia 6 minggu setelah subkultur. ...
92
26. Respon pertumbuhan tunas pisang cv. Ampyang hasil iradiasi gamma dalam media selektif mengandung filtrat kultur (FK) Foc 60%, usia 6 minggu setelah subkultur. ...
94
27. Persamaan regresi jumlah total tunas pisang yang tumbuh dalam media selektif mengandung FK Foc 30% - 40% -50% dan 60%. ……….…..
96
28. Plantlet insensitif FK Foc 60 % yang berasal dari hasil iradiasi: (a) 20 Gy, (b) 35 Gy dan (c) 50 Gy setelah 2 bulan ditumbuhkan pada media regenerasi secara in vitro. ...
98
29. Gejala kelayuan bibit varian tanaman pisang hasil seleksi in vitro yang berasal dari hasil iradiasi: (a-b) 50 Gy dan 25 Gy pada usia 30 hari setelah aklimatisasi, (c) pertumbuhan koloni cendawan Foc dalam media tanah, (d) bibit tanaman insensitif FK Foc usia 45 hari, (e) pengamatan mikroskopis potongan melintang akar tanaman insensitif FK Foc 60% pada usia 60 hari dengan perbesaran 40x. ...
101
30. Respon varian tanaman pisang cv. Ampyang hasil seleksi in vitro yang di infeksi konidia Foc: (a-e) representasi gambar skoring gejala kelayuan bibit pisang pada skor 0-4, (f-k) representasi gambar skoring nekrosis bonggol pada skor 0-5. ...
111
31 Respon ketahanan bibit varian insensitif Foc hasil seleksi in vitro
setelah 2 bulan diinfeksi dengan konidia F. oxysporum f.sp. cubense
isolat Medan di rumah kaca. Gambar horizontal (a) Gejala kelayuan bibit dan nekrosis bonggol (insert), (b) gejala kelayuan bibit dan nekrosis bonggol pisang cv. Ampyang. ………
113
32. Respon ketahanan bibit varian insensitif Foc hasil seleksi in vitro
setelah 2 bulan diinfeksi dengan konidia F. oxysporum f.sp. cubense
xxi
Fusarium pada usia 5 bulan setelah infeksi dengan (a-d) cendawan Foc
isolat Medan dan (e-g) Foc isolat Banyuwangi. ... 34. Potongan melintang akar pisang cv. Ampyang: (a) akar pisang sehat, (b) perubahan warna menjadi kecoklatan akibat kerusakan pada jaringan epidermis, xylem dan bakal akar cabang yang terinfeksi Foc dilihat dengan mikrokop cahaya, (c) warna merah marone pada jaringan xylem akar yang terinfeksi Foc pada akar tanaman yang sama dilihat dengan mikroskop fluorescent, perbesaran 40x. ...
122
35. Tanaman pisang yang berasal dari hasil iradiasi (a-b) 20 Gy, (c-d) 40 Gy, (e) 45 Gy dan (f) 50 Gy yang mengalami recovery setelah diinfeksi melalui metode perendam akar dalam suspensi konidia Foc
kerapatan 2.5 x 107 kon. mL-1 usia 1-2 bulan setelah recovery. ...
123
36. Representatif tanaman varian pisang cv. Ampyang pada lahan endemik layu Fusarium: (a) tanaman usia 3 bulan dengan varian bentuk daun menggulung, (b) pelepah menumpuk, (c) tanaman
recovery, (d) tanaman sehat, (e-f) tanaman yang menunjukkan gejala awal terinfeksi Foc pada usia 3 bulan dan 11 bulan setelah tanam, (g) tanaman berbuah pada usia 20 bulan setelah tanam dan gejala kalayuan pada anakan terserang layu Fusarium (insert). ...
127
37 (a) batang tidak terinfeksi Foc, (b) warna kecoklatan pada batang terinfeksi Foc, (c) pertumbuhan koloni cendawan Foc pada pelepah membusuk. Pengamatan secara mikroskopis dengan perbesaran 100x potongan membujur dan melintang, (d-e) pelepah pisang terinfeksi
Foc, (f-g) pelepah pisang sehat, (h-j) pertumbuhan koloni Foc hasil isolasi dari pelepah terinfeksi Foc tampak depan dan hasil isolasi dari tangkai buah tampak belakang (reversed colony). ...
129
38. Keragaman fenotipik buah pisang cv. Ampyang yang tumbuh pada lahan endemik layu Fusarium (a) buah dipanen setelah masak, (b-d) buah di panen setelah pohon tumbang, (e-g) karakteristik buah abnormal (h-i) buah yang cepat membusuk setelah dipanen. ...
130
39 Klon anakan yang berasal dari tanaman varian pisang cv. Amyang hasil iradiasi gamma yang di tanaman pada lahan endemik layu
Fusarium: (a) tanaman anakan yang mengalami recovery dan
tanaman kimera, (b) tunas baru tumbuh pada tanaman kimera, (c-d) klon anakan usia 4 bulan setelah dipindahkan dari lapang. ...
xxii
Halaman
1. Sebaran nilai dan jumlah tanaman pada karakter jumlah anakan, tinggi tanaman dan lingkar batang semu tanaman pisang cv. Ampyang pada pada usia 11 bulan setelah tanaman di lapangan. ...
165
2. Sebaran nilai dan jumlah tanaman pada karakter panjang daun, lebar daun tanaman dan rasio panjang dan lebar daun tanaman pisang cv. Ampyang pada usia 11 bulan setelah tanaman di lapangan. ...
166
3. Pengamatan keasaman (pH) tanah dan suhu tanah pada lahan endemik layu Fusarium ...
BAB I
PENDAHULUAN
Latar Belakang
Pisang dan plantain (Musa spp) merupakan tanaman pangan yang ditumbuhkan dan dikonsumsi oleh lebih dari 100 negara di daerah tropis dan subtropis (INIBAP 2000). Di Indonesia pisang dan plantain merupakan komoditas buah tropika yang perlu dikembangkan selain manggis, nenas dan pepaya yang dicanangkan oleh Kemetrian Riset dan Teknologi berdasarkan pertimbangan bahwa
komoditas buah tropika tersebut berorientasi kerakyatan, yang mampu menjadi
leverage factor bagi peningkatan kesejahteraan petani, kecil kemungkinan diimpor
secara segar, kualitas produk masih perlu ditingkatkan untuk memenuhi standar
konsumen, dapat diterima luas di pasar domestik, dan memiliki potensi di pasar dunia.
Kultivar-kultivar pisang yang dikonsumsi secara alami berkembang dari species Musa acuminata (genom A) dan Musa balbisiana (genom B). Kultivar diploid (AA dan AB), triploid (AAA, AAB, ABB) dan tetraploid (AAAA, AAAB, AABB atau ABBB), berasal dari hibrida di antara kedua species tersebut, dan di antara 9 subspecies Musa acuminata (Moore et al. 2001; Ploetz et al. 2003; 2007). Kedua species alami dan hibrida kompleks ini menghasilkan kombinasi jenis pisang yang dikonsumsi saat ini. Kultivar pisang dikelompokkan ke dalam dua tipe, yaitu jenis pisang meja (dessert type) yang dikonsumsi tanpa dimasak terlebih dahulu, dan jenis pisang olahan (cooking type) yang dimasak terlebih dahulu sebelum dikonsumsi (Valmayor et al. 2000; Pillay et al. 2004; Heslop-Harisson & Schwarzacher 2007). Produksi pisang meja atau pisang manis diperkirakan mencakup 43% dari total produksi pisang di seluruh dunia, sedangkan pisang olahan diperkirakan mencakup 57% (Valmayor et al. 2000).
tanaman pisang di Sumatra, Kalimantan, Sulawesi dan Papua sebesar 20.000.000 ha (Nasir et al. 2003). Pada saat ini penurunan produktivitas dan kualitas pisang disebagian besar kebun pekarangan, perkebunan tradisional dan perkebunan komersial di berbagai daerah di Jawa, Sumatra dan Sulawesi sebagian besar dikarenakan adanya penyakit layu Fusarium yang disebabkan oleh cendawan Fusarium oxysporum f.sp. cubense (Foc).
Penyakit layu Fusarium pada pisang disebabkan oleh cendawan tular tanah Fusarium oxysporum Schlecht. f.sp. cubense (E. F. Smith) Snyder and Hansen
(Foc), dikenal juga dengan nama Panama disease, atau ’penyakit layu kuning’. Patogen tumbuhan ini menyerang jaringan vaskuler, dan sangat berperan dalam sejumlah kerusakan pada tanaman pisang dan plantain. Pengendalian yang efektif dan berkelanjutan merupakan salah satu topik yang paling penting dalam bidang pertanian saat ini. Cendawan Foc pertama kali dilaporkan di Indonesia pada tahun 1916 dari perkebunan pisang di Pulau Jawa (Nasir et al. 1999), dan penyebaran penyakit ini berkembang secara eksponensial 32.2% setelah 4 tahun (Lee et al. 2001). Hilangnya eksport komoditas pada kultivar-kultivar komersial, seperti pada pisang Cavendish di Sumatera Selatan dapat mencapai 70%, di Sumatra Utara lebih dari 1000 ha perkebunan komersial pisang rusak dalam waktu tiga tahun, dan sebagian besar kerusakan pada kebun pekarangan, perkebunan tradisional dan perkebunan komersial tersebut disebabkan oleh serangan cendawan Fusarium oxysporum f.sp. cubense. Di propinsi Riau, 3000 ha perkebunan Cavendish seharga
US$ 2.5 juta, di Halmahera 3000 ha dan Lampung 1700 ha telah diserang oleh Foc ras 4 (Nasir et al. 2003), secara umum penyakit layu Fusarium dimasa depan dapat menjadi epidemik di perkebunan-perkebunan di Sumatra dan di Indonesia (Nasir et al. 1999; Moore et al. 2000).
penyakit ini, transfer gen-gen resisten ke dalam varietas yang rentan dengan persilangan konvensional sangat sulit karena sifat triploid kultivar pisang dan produksi biji yang rendah (Matsumoto et al. 1999; De Ascensao & Dubery 2000; Hwang & Ko 2004). Teknik persilangan konvensional juga diketahui memiliki tingkat keberhasilan yang rendah (Companioni et al. 2003; 2006) dan membutuhkan waktu yang lama, sehingga prospek pengendalian penyakit pisang sangat bergantung pada tersedianya kultivar-kultivar inang yang resisten (Moore et al. 2001; Hwang & Ko 2004).
Pisang dan plantain (Musa spp) diperbanyak secara vegetatif melalui suckers atau potongan rhizome, dan sebagian besar pisang yang dikonsumsi
bersifat triploid, steril, dan partenokarpi serta membutuhkan waktu generasi yang panjang dalam siklus vegetatifnya, sehingga metode pemuliaan secara konvensional sulit dilakukan (Musoke et al. 1999; Valmayor et al. 2000; Hwang & Ko 2004, Suprasana et al. 2008). Pada banyak tanaman yang diperbanyak secara vegetatif, induksi mutasi yang dikombinasikan dengan teknik kultur in vitro merupakan suatu metode yang efektif untuk peningkatan keragaman suatu tanaman (Novak & Brunner 1992; Roux 2004), sangat efektif untuk peningkatkan sumber genetik alami, dan secara signifikan mampu mendukung pengembangan kultivar-kultivar tanaman buah (IAEA 2009). Agen mutagenik seperti iradiasi dan beberapa bahan kimia seperti EMS dapat digunakan untuk mutasi induksi dan menghasilkan variasi genetik dimana mutan-mutan yang diinginkan kemungkinan dapat diseleksi (Novak & Brunner 1992; IAEA 2009).
Pendekatan Masalah
Penyakit layu Fusarium yang disebabkan oleh cendawan Fusarium oxysporum f.sp. cubense (Foc) menyerang sebagian besar pertanian pisang dan
plantain (Musa spp.) di Indonesia. Pengendalian patogen secara kimiawi maupun
hayati sulit dilakukan, karena cendawan ini dapat bertahan hidup secara saprofit dalam tanah sebagai mycelium atau sebagai chlamydospora tanpa adanya inang, serta mampu bertahan hidup pada tanah dalam jangka waktu yang lama (Agrios 2005), sehingga penyediaan kultivar-kultivar pisang yang resisten terhadap layu Fusarium merupakan suatu alternatif yang penting untuk dilakukan.
Usaha penyediaan kultivar yang resisten terhadap penyakit layu Fusarium memiliki kendala berupa sulitnya diperoleh keragaman genetik pada kultivar-kultivar pisang, karena tanaman ini diperbanyak secara vegetatif (klonal). Peningkatan keragaman pada tanaman akan dilakukan melalui pendekatan secara in vitro, melalui induksi tunas pisang aseptis dengan sinar gamma (γ) agar diperoleh varian somaklonal serta melalui teknik subkultur berulang untuk meningkatkan kemungkinan perolehan varian dan kestabilan karakter varian. Peningkatan variasi somaklonal perlu dilakukan untuk menjamin ketersediaan plasma nutfah yang akan digunakan untuk seleksi in vitro ketahanan tanaman terhadap penyakit. Identifikasi fenotipik varian tanaman hasil induksi mutasi dengan sinar gamma di rumah kaca dan di lapangan dilakukan untuk memperoleh tanaman pisang dengan keragaman genetik yang tinggi, sehingga dari percobaan ini diperoleh tanaman pisang (Musa acuminta, AAA) cv. Ampyang yang memiliki karakter agronomis yang diinginkan.
Sistem seleksi secara in vitro untuk resistensi terhadap layu Fusarium telah dideskripsikan untuk tanaman pisang dan plantain, namun tanaman pisang terdiri dari banyak kultivar dengan genotipe yang berbeda, dan kemungkinan besar memiliki karakteristik ketahanan yang berbeda. Demikian pula cendawan Foc isolat Banyuwangi merupakan isolat yang menyerang abaka (Musa textilis Nee), sehingga pengujian awal untuk mengetahui patogenitas dan virulensi Foc isolat Banyuwangi terhadap pisang cv. Ampyang secara in vitro dan ex vitro, serta pengujian efektivitas Foc isolat ini sebagai agen penyeleksi tetap diperlukan.
tanaman terhadap layu Fusarium di rumah kaca, sehingga terhindar dari penarikan kesimpulan yang salah. Identifikasi resistensi klon-klon varian yang berasal dari hasil mutasi induksi yang dilanjutkan dengan seleksi in vitro dilakukan di rumah kaca, dan identifikasi resistensi klon varian yang berasal dari hasil mutasi induksi tanpa tahapan seleksi in vitro dilakukan melalui pendekatan secara agronomis di rumah kaca dan di lapangan. Serangkaian percobaan ini diharapkan dapat memberikan kontribusi yang paling relevan dalam penyediaan benih pisang yang resisten atau meningkat resistensinya terhadap penyakit layu Fusarium.
Tujuan Umum Penelitian:
1. Meningkatkan keragaman tanaman pisang (Musa acuminata, AAA) cv. Ampyang melalui teknik mutasi induksi dengan iradiasi gamma secara in vitro. 2. Mendapatkan klon-klon dari kultivar pisang (Musa acuminata, AAA) cv.
Ampyang yang resistensi terhadap layu Fusarium yang disebabkan oleh cendawan Fusarium oxysporum f.sp. cubense.
Manfaat Penelitian
1. Penentuan kisaranan dosis letal (LD20-50) pisang cv. Ampyang (Musa acuminata,
AAA, subgrup non-Cavendish) yang dapat menjadi acuan untuk menginduksi mutasi pada pisang kultivar lainnya dengan genom AAA.
2. Diperoleh parameter untuk mengidentifikasi keragaman fenotipik tanaman secara kuantitatif dan kualitatif, dan prosedur baku untuk mendapatkan tanaman pisang yang resisten terhadap penyakit layu Fusarium.
3. Perolehan klon-klon yang dapat digunakan sebagai plasma nutfah tanaman pisang cv. Ampyang resisten layu Fusarium dan potensi pengembangan tanaman pisang secara klonal.
Hipotesis.
1. Teknik mutasi induksi melalui pemberian berbagai dosis iradiasi gamma dapat meningkatkan keragaman tanaman pisang cv. Ampyang.
Ruang Lingkup Penelitian
Tujian akhir dari penelitian ini adalah mendapatkan klon tanaman pisang cv. Ampyang dengan keragaman yang tinggi dan resisten penyakit layu Fusarium. Untuk mencapai tujuan tersebut dilakukan serangkaian percobaan yang meliputi:
I. RADIOSENSITIVITAS PISANG cv. AMPYANG DAN POTENSI PENGGUNAAN IRADIASI GAMMA UNTUK INDUKSI VARIAN
Percobaan 1. Radiosensitivitas tunas pisang terhadap iradiasi gamma.
Percobaan 2. Regenerasi tunas dan identifikasi fenotipik plantlet pisang hasil iradiasi gamma.
II. EVALUASI KERAGAMAN FENOTIPIK PADA PISANG cv.
AMPYANG HASIL MUTASI INDUKSI DENGAN IRADIASI GAMMA
Percobaan 3. Evaluasi di rumah kaca terhadap keragaman fenotipik tanaman pisang.
Percobaan 4. Evaluasi di lapangan terhadap keragaman fenotipik tanaman pisang.
III. VIRULENSI F. oxysporum f.sp. cubense ISOLAT BANYUWANGI UNTUK IDENTIFIKASI KETAHANAN PISANG cv. AMPYANG
TERHADAP LAYU FUSARIUM
Percobaan 5. Uji virulensi Foc isolat Banyuwangi dengan metode kultur ganda secara in vitro.
Percobaan 6. Evaluasi virulensi Foc isolat Banyuwangi dan efektivitas metode uji ketahanan pisang cv. Ampyang terhadap layu Fusarium.
IV. SELEKSI IN VITRO UNTUK MENDAPATKAN PLANTLET PISANG
cv. AMPYANG HASIL IRADIASI GAMMA INSENSITIF FILTRAT KULTUR F. oxysporum f.sp. cubense
Percobaan 7. Efektivitas filtrat kultur (FK) Foc isolat Banyuwangi sebagai agen penyeleksi.
Percobaan 8. Seleksi in vitro secara bertingkat tunas pisang cv. Ampyang dengan FK Foc
Percobaan 9. Regenerasi dan aklimatisasi plantlet pisang insensitif FK Foc.
V. KETAHANAN VARIAN PISANG (Musa acuminata, AAA) cv.
AMPYANG TERHADAP LAYU FUSARIUM
Percobaan 10. Evaluasi ketahanan varian pisang cv. Ampyang hasil iradiasi gamma yang dilanjutkan dengan seleksi in vitro terhadap layu Fusarium di rumah kaca.
Percobaan 11. Evaluasi ketahanan varian pisang cv. Ampyang hasil iradiasi gamma terhadap layu Fusarium di rumah kaca.
Diagram Alir Strategi Penelitian.
A. Mutasi induksi untuk meningkatkan keragaman dan ketahanan tanaman
3. Eva lua si ke ra g a m a n fe no tip ik ta n. d i rum a h ka c a
P ro li fe ra si tu n a s (8 b ln ))
11. Eva lua si ke ta ha na n va ria n p isa ng ha sil ira d ia si d i rum a h ka c a
klo n va ria n p isa ng (3- 4 b ln)
Pla ntle t p isa ng in v itro Pe rb a ny aka n tuna s (2 b ln)
MT: MS + 4.50 m g L- 1BA P, 0.22 TDZ,
0.175 m g L- 1IA A
12. Eva lua si ke ta ha na n va ria n
t’ha d a p la yuFusa rium d i la p a ng a n
M1V1
2. Re g e ne ra si & id e ntifika si fe no tip ik va ria n
M1V2… M1V6- 8
LD20- 50
6. Eva lua si virule nsi Fo c iso la t Bw & e fe ktivita s m e to d e uji ke ta ha na n
p isa ng t’ha d a p la yu fusa rium
I. Ra d io se nsitivita s p isa ng thd p sina rγ
0, 20, 25, 30, 35, 40, 45, 50 G y
4. Eva lua si ke ra g a m a n fe no tip ik ta na m a n d i la p a ng a n
O ut p ut: Klo n p isang c v. A m p y ang tahan p e ny akit lay u Fusarium
B. Seleksi in vitro untuk mendapatkan klon pisang resisten layu Fusarium
Inkub a si c e nd a wa nFo c d a la m m e d ia PDB →shaker
Fo c iso la t Bw
d lm m e d ia PDA Filtra t KulturFo c
Ste rilisasi 121oC – 20’
7. Efe ktivita s FK Fo c sb g
a g e n p e nye le ksi.
8. Se le ksiin v itro se c a ra b e rting ka t
tuna s p isa ng d e ng a n FK Fo c
Ko nse ntrasi FK y g e fe ktif
9. Re g e ne ra si d a n A klim a tisa si
Pla nle t inse nsitifFo c
O ut p ut: Klo n p isang c v. A m p y ang tahan p e ny akit lay u Fusarium
Klo n inse nsitifFo c
Va ria n tuna s p isa ngin vitro
(M1V6- 8)
5. Uji virule nsiFo c iso la t Bw
p d p isa ng c v. A m p ya ng
Tunas pisang in vitro
FK Fo c 30%
FK Fo c 40%
FK Fo c 50%
FK Fo c 60%
10. Eva lua si ke ta ha na n va ria n ha sil se le ksi in vitro thd p
[image:30.595.93.516.67.769.2]la yuFusa rium d i rum a h ka c a
BAB II
TINJAUAN PUSTAKA
Tanaman Pisang (Musa spp.)
Taksonomi
Pisang dan plantain (Musa spp) dan pisang ornamental merupakan tanaman yang termasuk famili Musaceae, ordo Zingiberales. Berasal dari Asia Tenggara dan daerah tropis dan sub tropis termasuk Afika dan Papua New Guinea (Ploetz et al. 2007). Menurut Valmayor et al. (2000) klon-klon dari pisang diploid di
kultivasi di daerah lembab di Asia Tenggara, dan perkembangan kultivar triploid tanpa biji yang kokoh merupakan hasil dari restitusi kromosom, dan/ atau persilangan dari edible diploid dengan M. acuminata liar (Daniells et al 2001).
Kultivar-kultivar pisang secara alami berkembang dari species Musa acuminata (genom A) dan Musa balbisiana (genom B). Kultivar diploid (grup AA
dan AB), triploid (grup AAA, AAB, ABB) dan tetraploid (grup AAAB) berasal dari hibrida di antara subspecies M. acuminata yang memiliki 9 subspecies, dan di antara M. acuminata dan M. balbisiana (Moore et al. 2001; Ploetz et al. 2003). Kedua species alami dan hibrida kompleks ini menghasilkan kombinasi jenis-jenis pisang yang dikonsumsi saat ini. Kondisi ini menyebabkan identifikasi pisang menjadi sulit, sehingga para pemulia sepakat mengadopsi penamaan tanaman melalui tiga kategori sistem penamaan yaitu species, grup genom dan kultivar (Valmayor et al. 2000; Ploetz et al. 2007). Penamaan pisang umumnya digunakan untuk subgrup spesifik jenis pisang meja (dessert banana) dan plantain untuk pisang olahan (cooking banana). Pisang Ampyang merupakan jenis pisang meja yang termasuk grup genom AAA, subgrup non-Cavendish (Valmayor et al. 2000)
Karakteristik dan Manfaat
Pisang dapat berbuah tanpa melalui proses fertilisasi yang dikenal sebagai partenokarpi dan menghasilkan bonggol di bagian basal (basal suckers) yang dapat
partenokarpi dari M. acuminata sampai saat ini belum dikarakterisasi. Buah yang merupakan produk utama pisang memiliki komposisi zat gizi/100 gram bahan yang cukup tinggi, yaitu air 75.00 gr; Energi 88.00 K; Karbohidrat 23.00 gr, Protein 1.20 gr; Lemak 0.20 gr; Ca 8.00 mg; P 28.00 mg; Fe 0,60 mg; Vitamin A 439.00 SB, B-1 0.04 mg dan C 78.00 mg.
Cendawan F. oxysporum f.sp. cubense Penyebab Layu Fusarium
Biologi Fusarium oxysporum
Fusarium spp termasuk dalam subklas Deuteromycetes, penamaan Fusarium oxysporum Schlecht sebagai yang dikemukakan oleh Snyder and
Hansen pada tahun 1940 (Kistler 2001), merupakan cendawan penyebab layu Fusarium yang tersebar luas menyerang berbagai tanaman. Cendawan ini
memproduksi tiga tipe spora aseksual yaitu mikrokonidia, makrokonidia dan chlamidospora yang dorman dan immobil. Chlamidospora terdapat pada jaringan yang membusuk, sampai akhirnya berkecambah melalui nutrisi yang dilepaskan oleh akar tanaman (Agrios 2005), ekskresi dari akar non-inang, atau kontak dengan bagian akar segar tanaman yang tidak terkoloni oleh cendawan Fusarium oxysporum.
Bentuk patogenik Fusarium oxysporum yang menyerang tanaman dibagi menjadi formae speciales berdasarkan tanaman inang yang diserang, selanjutnya dibagi kembali ke dalam ’ras’ berdasarkan virulensinya terhadap beberapa kultivar inang (Agrios 2005). Penyakit layu Fusarium diketahui menjadi faktor pembatas dalam produktivitas pada beberapa tanaman pangan dan hortikultura di antaranya pada tanaman pisang (Musa spp) (F. oxysporum f.sp. cubense), melon (Cucumis spp) (F. oxysporum f.sp. melonis), bawang merah (Allium spp) (F. oxysporum f.sp. cepae), tomat (Lycopersicon spp) (F. oxysporum f.sp. lycopersici), krisan (Chrysanthemum spp) (F. oxysporum f.sp. chrysantemi) dan
carnation (Dianthus spp) (F. oxysporum f.sp. dianthi) (Agrios 2005).
Perkembangan dan Penyebaran Cendawan F. oxysporum f.sp. cubense.
layu Fusarium atau panama disesase, merupakan penyakit yang disebabkan oleh Fusarium oxysporum Schlecht. f.sp. cubense (E.F. Smith) Snyder and Hansen
(Foc), berasal dari Asia Tenggara, namun pertama kali dilaporkan pada tahun 1876 di Australia (Ploettz & Pegg 1997). Di Indonesia penyakit ini berkembang pada tahun 1916 di pulau Jawa. Cendawan ini menyerang akar tanaman pisang dan plantain yang tersebar luas, sangat destruktif dan merupakan penyakit tanaman
yang sangat merugikan yang menyerang berbagai perkebunan dan industri pisang di Indonesia (Nasir et al. 2003), walaupun informasi yang akurat dan pengaruhnya secara ekonomi masih sangat terbatas (Hermanto et al. 2011). Besarnya kerusakan dan hilangnya produksi tanaman pisang bervariasi bergantung pada lokasi, kultivar, lama kultivasi, dan ada atau tidaknya ras 4 (Moore el al 2001; Masdek et al 2003)
Cendawan F. oxysporum f.sp. cubense ini dikenal dalam empat ras, yang dibedakan berdasarkan suseptibilitas inang. Ras 1 menyerang pisang varietas ‘Gros Michel’, ‘Lady Finger’ (AAB), dan ‘Silk’ (AAB). Ras 2 menyerang antara lain varietas ‘Bluggoe’ (ABB) dan ras 4 menyerang pisang Cavendish (AAA) dan pisang lain yang diserang ras 1 dan 2. Ras 3 menyerang Heliconia spp yang memiliki kekerabatan dekat dengan pisang, tetapi tidak dianggap sebagai patogen terhadap tanaman pisang. Ras 4 dikelompokkan lagi menjadi ‘sub-tropikal’ dan ‘tropikal’ strains. Di Indonesia dari 261 isolat yang telah dikoleksi di Sumatra dan di Jawa sejak tahun 1990 sampai 2002, 70% adalah ras 4 (Masdek et al. 2003), dan ras 4 ‘tropikal’ ini dilaporkan paling virulen dan tersebar di Halmahera, Irian Jaya, Jawa, Sumatra dan Sulawesi (Ploetz & Churchill 2011).
Penyebaran Foc dan Gejala Kelayuan pada Tanaman Pisang
Gejala eksternal pertama yang ditimbulkan penyakit ini adalah warna kuning pada daun bagian bawah yang sangat jelas terlihat, sehingga penyakit ini dinamakan pula yellow leaf disease. Pada pisang Cavendish gejala ini umumnya terjadi 5 bulan setelah penanaman (Hwang & Ko, 2004). Daun-daun yang menguning dimulai dari daerah tepi daun dan berkembang ke arah tulang daun, kemudian daun berkembang menjadi titik-titik hitam, petiole akan menjadi coklat dan melengkung, yang diikuti oleh pengeringan daun. Lembaran daun-daun yang telah mati seringkali akan mengelilingi batang-semu pisang. Proses ini dimulai dari daun tua sampai daun-daun muda (Nasir et al. 1999; Daly & Walduck 2006).
Gejala pertama secara internal terlihat jelas di jaringan xylem akar dan rhizoma, berupa titik-titik atau garis-garis berwarna merah kecoklatan sampai berwarna merah maroon. Pada tahapan awal infeksi secara khusus terjadi perubahan warna kuning dengan pola melingkar dibagian tengah pada potongan melintang rhizoma. Gejala berkembang ke dalam batang semu pisang membentuk garis-garis kecoklatan sampai bagian ujung dari batang (Daly & Walduck 2006), selanjutnya daun menjadi layu dan menguning, pucuk-pucuk tanaman menjadi pucuk sukulen, yang pada akhirnya seluruh tanaman akan mengalami kematian (Hwang & Ko 2004; Agrios 2005).
Metabolit dan Toksin pada Cendawan Fusarium
Toksin utama yang diproduksi berbagai species cendawan Fusarium dan Gibberella adalah asam fusarat (5-n-butylpicolinic acid). Asam fusarat merupakan
toksin non-spesisfik inang (Goodman et al. 1986; Huang 2001; Agrios 2005), karena bersifat non-spesifik inang maka asam fusarat dapat berpengaruh baik terhadap inang atau non-inang (Goodman et al. 1986). Asam fusarat merupakan suatu myco-toksin, antibiotik, insektisida dan fitotoksin (Huang, 2001), berperan dalam perubahan permiabilitas membran sel, mengakibatkan terganggunya keseimbangan ion-ion anorganik dalam tanaman yang terinfeksi patogen dan mengakibatkan kebocoran elektrolit dalam sel (Goodman et al. 1986).
polygalacturonase) berperan aktif dalam perkembangan penyakit pada jaringan
vaskuler (Agrios 2005). Pembongkaran substansi pektat akan membebaskan fragmen pektat yang masuk melewati aliran vaskuler yang akan meningkatkan viskositas dan mereduksi laju aliran air (Goodman et al. 1986).
Mekanisme Resistensi pada Tanaman
Tanaman tingkat tinggi dan organisme lain akan menggunakan mekanisme pertahanan dengan cakupan yang luas untuk melindungi dirinya menghadapi invasi patogen. Kemampuan setiap tanaman untuk menimbulkan suatu reaksi pertahanan diketahui dimediasi melalui suatu proses inisiasi pengenalan antara tanaman dan patogen, yang melibatkan deteksi beberapa molekul signal tertentu yang unik dari patogen melalui molekul menyerupai reseptor (receptor-like molecules) dalam tanaman, dan mekanisme cascade biokimiawi yang mengawali ekspresi resistensi tanaman (Ryals et al. 1994; Huang 2001; Roncero et al. 2003).
Sel tanaman memiliki kemampuan mempertahankan diri dari patogen melalui mekasime dengan cakupan yang luas, baik secara kimiawi maupun fisik. Induksi resistensi dapat berupa induksi lokal yang merespon molekul signal dengan mobilitas terbatas atau induksi sistemik yang merespon molekul sinyal dengan mobilitas tinggi, konstitutif (fitoantisipin) atau respon yang diiduksi (fitoaleksin) (Mansfield 2000; van Loon 2000). Resistensi pada tanaman terjadi melalui jalur pensinyal yang diinduksi oleh infeksi patogen dan patogen avirulen (systemic acquired resistance), bakteri non-patogen (induced systemic resistance) dan
senyawa kimia tertentu. Resistensi melalui jalur SAR (systemic acquired resistance) menyebabkan hipersensitif respon berupa nekrosis jaringan. Ekspresi
SAR bergantung pada akumulasi asam salisilat (SA), dan asosiasi induksi protein yang berperan dalam patogenesis (patogenesis-related proteins) dengan aktivitas anti-patogen, sebaliknya rhizobacteria yang memediasi ISR (induced systemic resistance) tidak membutuhkan SA, dapat terjadi tanpa produksi PR protein, dan
bergantung pada signal etilen dan asam jasmonat (JA) (van Loon 2000).
Respon Tanaman Pisang terhadapInfeksiFoc.
sekitar akar pisang yang terinfeksi. Lignifikasi dinding sel akar pisang ini merupakan salah satu barier yang paling efektif melawan invasi patogen. Sintesis enzim-enzim yang terlibat dalam penguatan dinding sel akar pisang juga terjadi, seperti phenylalanine ammonia lyase, cinnamyl alcohol dehydrogenase, peroxidase, dan pholyphenol oxidase (De Ascensao & Dubery 2000; 2003).
Enzim yang berperan dalam fortifikasi dinding sel ini akan meningkat selama adanya ekspresi hipersensitif respon (Huang 2001), yaitu suatu respon yang terjadi pada setiap tanaman, dan berperan utama dalam pertahanan tanaman terhadap infeksi patogen, karena sifatnya yang sangat resisten terhadap degradasi enzim mikroba. Selanjutnya terjadi deposisi kalose, terbentuk gums, tylose, dan gel pada jaringan xylem akar pisang, yang kemungkinan merupakan hasil fotoasimilat dari tanaman untuk mengisolasi patogen (Agrios 2005), namun apabila terjadi infeksi yang berkelanjutan, akan terjadi akumulasi senyawa tersebut pada pembuluh trachea dan tracheid jaringan xilem, yang berakibat terganggunya transportasi air dan garam mineral dari akar ke daun. Akumulasi gel yang berlebihan juga terjadi dari perubahan enzim kompleks pada lamela tengah-dinding primer sel parenkim xilem, baik pada pit membran atau dibagian lempeng perforasi. Pembentukan gel dan degradasi lamela tengah dari pit membran ini, selanjutnya merangsang pembentukan tylose yang akan mereduksi transport air dan garam mineral di jaringan vaskuler sehingga terjadi kelayuan yang diikuti oleh pencoklatan jaringan karena adanya oksidasi senyawa fenolik (Goodman et al. 1986).
Mekanisme resistensi tanaman pisang juga terjadi melalui pembentukan fitoaleksin, dan pembentukan senyawa protein antimikroba, beberapa fitoaleksin yang telah berhasil diisolasi pada tanaman pisang yang diinfeksi oleh Fusarium oxysporum f.sp. cubense antara lain musalenones C, D, E, F pada rhizoma,
irenolone I pada daun dan buah (Luis et al. 1996),
2-hydroxy-9-phenyl-phenalene-1-one (hydroxyanigofuron) pada rhizoma (Luis et al. 1996; Binks et al. 1997;
Borges et al. 2003) dari Musa acuminata, AAA, kultivar Grande Naine (subgrup Cavendish). Senyawa anti-mikroba yang telah berhasil diisolasi adalah Banana thaumatin-like protein (Ban-TLP) pada buah pisang yang masak (Barre et al.
Kultur In vitro dan Variasi Somaklonal
Kultur in vitro Tanaman Pisang
Kultur in vitro atau kultur jaringan pada saat ini merupakan suatu metode yang sudah banyak dilakukan untuk tujuan perbanyakan tanaman dan merupakan alat penting bagi studi dasar maupun aplikasi yang berorientasi komersial, termasuk teknik-teknik dan metode yang sesuai untuk penelitian dalam banyak disiplin ilmu botani dan beberapa objek percobaan. Pada tanaman pisang, pertumbuhan jaringan yang terorganisir secara in vitro dibatasi pada kultur embrio dan kultur pucuk, sampai saat ini, lebih dari 1.000 aksesi yang berbeda dari kultur pucuk (shoot-tips culture) telah diinisiasi secara in vitro, dimultiplikasi dan dipelihara pada kondisi temperatur rendah (16 ± 1oC) (Strosse et al. 2004).
Multiplikasi pucuk tanaman pisang secara in vitro pada umumnya menggunakan pucuk meristem pada media Murashige dan Skoog (MS) dengan penambahn BA (6-benzyladenin) 2.25 mg L-1 dan IAA (Indole-3-acetic acid) 0.175 mg L-1 (Strosse et al. 2004), atau BA 4.5 mg L-1 (Mariska et al. 2006). Pemberian asam askorbat atau asam sitrat dengan kisaran 10-150 mg L-1 pada medium dasar dapat mengurangi jaringan menjadi hitam, sedangkan untuk pemeliharaan dan menginduksi perakaran konsentrasi BA diturunkan menjadi 0.225 mg L-1 (Strosse et al. 2004). Konsentrasi BA dapat ditingkatkan 10 kalinya menjadi 22.5 mg L-1 dan 0.175 mg L-1 IAA apabila produksi proliferasi meristem yang tinggi dibutuhkan.
Laju multiplikasi tunas pisang bergantung pada baik pada sitokinin maupun genotip tanaman. Secara umum pucuk-pucuk kultivar yang hanya memiliki genom A akan memproduksi 2-4 pucuk baru, sedangkan kultivar yang memiliki satu atau dua genom B akan memproduksi kluster-kluster pucuk yang banyak dan kuncup pada setiap siklus subkultur. Pucuk aksilar dan adventif baru dapat tumbuh langsung dari eksplan ujung pucuk sekitar 6-12 minggu setelah inisiasi kultur, bergantung ukuran eksplan yang diinisiasi. Kluster pucuk dapat dipisahkan, dan di subkultur kembali dengan interval 4-6 minggu (Strosse et al. 2004).
meristem, pada medium semi padat mengandung 1 mgL-1 2,4-D (2.4-dichlorophenoxyacetic acid) dan 0.22 mgL-1 zeatin. Respon embriogenik bergantung tidak hanya pada genotipe pisang tetapi juga pada galur terseleksi dan percobaan yang dilakukan (Strosse et al. 2004).
Faktor-faktor dalam Induksi Variasi Somaklonal.
Variasi somaklonal merupakan variasi genetik yang muncul di antara individual sel anakan pada tanaman yang diregenerasi melalui kultur jaringan dan sel (Collin & Edwards 1999; Hartman et al. 2002), seringkali didefinisikan sebagai keragaman genetik dari tanaman yang berasal dari sel somatik. Variasi somaklonal yang diperoleh melalui teknik in vitro merupakan pendekatan yang cepat dan dapat dipercaya untuk peningkatan dan perbaikan tanaman. Beberapa klon pisang yang memiliki kualitas superior diproduksikan melalui seleksi terhadap berbagai pendekatan, termasuk resisten terhadap penyakit (Ho 1999; Hwang & Ko 2004).
Laju terbentuknya varian somaklonal dapat ditingkatkan melalui induksi mutasi secara fisik maupun kimiawi. Berbagai varian somaklon dalam kultur in vitro memberi peluang untuk mengidentifikasi varian-varian tertentu yang mempunyai sifat unggul. Berbagai variasi genetik telah digunakan untuk resistensi tanaman terhadap patogen, toleran terhadap stress abiotik dan biotik, resistensi herbisida, dan perolehan banyak variasi lainnya. Faktor-faktor lain yang dapat mempengaruhi frekuensi munculnya variasi somaklonal dalam kultur in vitro pisang, adalah genotip tanaman induk, jumlah siklus sub-kultur dan umur jaringan dalam kultur in vitro, konsentrasi berbagai komponen media pertumbuhan, tipe regenerasi, tipe dan konsentrasi zat pangatur tumbuh (Nwauzoma et al. 2002), dan penggunaan kondisi selektif dalam media in vitro. Menurut Hartman et al. (2002), variabilitas somaklonal dapat dihasilkan karena (a) variasi dengan karakter tertentu memang ada di antara populasi sel, (b) sistem perbanyakan tanaman secara in vitro melalui pembentukan kalus akan menghasilkan variasi atau (c) variasi dapat diinduksi melalui agen mutagenik secara fisik maupun kimiawi.
Induksi variasi somaklonal dengan mutagen
pisang merupakan tanaman yang diperbanyak secara vegetatif sehingga memiliki keterbatasan dalam perolehan variasi genetik. Meskipun mutasi secara spontan dapat memberi kontribusi terhadap diversitas genetik pada Musa dan secara nyata meningkatkan variasi, namun frekwensinya relatif kecil. Penggunaan kultur in vitro untuk menginduksi mutasi pada Musa spp agar diperoleh variasi somaklon dapat menjadi suatu metode alternatif, bila beberapa tahapan dalam proses induksi mutasi dapat di optimalkan yaitu : (1) penggunaan bahan tanaman sebagai eksplan; (2) determinasi dosis mutagenik yang optimal untuk mutagen fisik dan kimia; (3) proses pemisahan chimera; (4) aplikasi metode skrining awal untuk seleksi mutan-mutan yang berguna untuk peningkatan karakter tanaman (Roux 2004).
Peningkatan variasi somaklonal melalui pemberian mutagen fisik, mutagen kimia dan subkultur berulang telah dilakukan oleh banyak peneliti. Beberapa laporan penelitian mengemukakan bahwa pemberian dosis letal (LD) yang tepat juga merupakan faktor penentu untuk keberhasilan perolehan varian-varian somaklon pada pisang (Musoke et al. 1999; Mak et al. 2004). Penggunaan iradiasi gamma (γ) untuk pengembangan tanaman buah-buahan telah banyak digunakan di antaranya untuk mendapatkan tanaman pisang resisten penyakit layu Fusarium (Smith et al. 1995; Sutarto et al. 1998; Musoke et al.1999; Roux et al 2004; Mak et al. 2004; Hwang & Ko 2004; Hutami et al. 2006; Smith et al. 2006; ); induksi
embriogenesis dan multiplikasi tunas jeruk limau madu dan limau langkat (Citrus suhuiensis) (Noor et al. 2009), untuk mendapatkan tanaman mangga (Mangifera
indica L.) resisten Antracnose (Litz 2009), dan tanaman pepaya (Carica papaya L.)
subkultur ke- lima dan ke sembilan pada media Murashige dan Skoog dengan 2.5 mg L-1 BAP yang disubkultur ke media 4.0 mg L-1 BAP (Rodrigues 2004).
Seleksi in vitro untuk Resistensi Pisang terhadap Layu Fusarium
Seleksi in vitro untuk ketahanan terhadap penyakit pada tanaman buah merupakan suatu fenomena yang melibatkan interaksi antara inang-patogen dan pengaruh aditif faktor abiotik yang mendukung perkembangan suatu penyakit (Jayasankar & Gray 2005; Chandra et al. 2010). Pada saat ini pengembangan strategis pemuliaan tanaman untuk ketahanan terhadap penyakit dilakukan melalui metode kultur jaringan melalui teknik seleksi varian somaklonal terhadap patogen spesifik atau fitotoksin. Pada jenis pisang meja dan pisang olahan, pengujian adanya varian somaklonal dengan karakteristik resisten terhadap F. oxysporum f.sp. cubense (Foc) dapat dilakukan melalui seleksi in vitro dengan menggunakan agen
penyeleksi berupa elisitor dari Foc (De Ascensao & Dubery 2000, 2003; Aguielar et al. 2