KAJIAN PENANDA GENETIK GEN CYTOCHROME B
DAN DAERAH D-LOOP PADA
Tarsius sp.
OLEH :
RINI WIDAYANTI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ABSTRACT
RINI WIDAYANTI. The Study of Genetic Marker on Cytochrome B gene and D-loop region of Tarsius sp.. Under the direction of DEDY DURYADI SOLIHIN, DONDIN SAJUTHIand R. R. DYAH PERWITASARI.
Tarsius is an endemic species of Indonesia that is threatened. Conservation of this species would yield better results if its genetic make up and diversity determined. The objective of this research was to study the specific genetic marker on Cyt b gene and D-loop region of Tarsius sp. Sequencing of PCR product using primer H15149 on Cyt b gene yielded base sequence of 276 nts (coding 92 amino acids), while sequencing using primer DLTARPROF on D-loop resulted in base sequence of 270 nts. Results of Cyt b and D-loop fragments sequencing were put on multiple alignment with other primates from Genbank with the aid of software Genetyc-Win Version 3.0 and Clustal W, and were analyzed using MEGA program version 3.1.Fourteen different amino acid sites were found. Tarsius dianae had 12 amino acid sites (amino acid no. 2, 6, 9, 22, 23, 29, 39, 41, 42, 45, 55, and 85), T. spectrum had 7 amino acid sites (amino acid no. 2, 6, 9, 41, 45, 55 and 85) and T. bancanus had 2 amino acid sites (amino acid no. 23 and 45) which can be used as genetic marker. Five unique amino acids were found on T. dianae, such as amino acid site: 6 (valine), 22 (alanin), 29 (alanin), 39 (serin) and site 42 (valine). The genetic distance based on nucleotide Cyt b calculated using Kimura 2-parameter model indicated that in the smallest value of 0.7%, biggest 22.3% and average 13.1%, while for D-loop, the smallest genetic distance is 0% and biggest 11.8% and average 2.3%. The phylogenetic tree using neighbor Joining Method based on the sequence of nucleotide and amino acid Cyt b reveded differentiation among
ABSTRAK
RINI WIDAYANTI. Kajian Penanda Genetik Pada Gen Cytochrome B dan Daerah D-loopTarsius sp.. Dibimbing oleh DEDY DURYADI SOLIHIN, DONDIN SAJUTHI, R.R. DYAH PERWITASARI
Tarsius merupakan salah satu satwa endemik Indonesia yang keberadaannya mulai memprihatinkan. Konservasi sebagai salah satu cara untuk pelestarian satwa ini akan lebih terarah dan berhasil guna apabila karakteristik dan keragaman sumber genetiknya diketahui dengan pasti. Tujuan dari penelitian ini adalah mengkaji penanda genetik spesifik pada gen Cyt b dan daerah D-loopTarsius sp.
Penentuan runutan hasil PCR menggunakan primer H15149 pada gen
Cyt b didapatkan runutan basa sebesar 276 nt (menyandi 92 asam amino), sedangkan hasil pengurutan menggunakan primer DLTARPROF pada daerah D-loop didapatkan runutan basa sebesar 270 nt. Fragmen Cyt b dan D-loop hasil pengurutan disejajarkan berganda dengan primata lain dari data Genbank
dengan bantuan perangkat lunak Genetyx-Win versi 3.0 dan Clustal W, kemudian dianalisis dengan menggunakan program MEGA versi 3.1. Dari hasil analisis diperoleh 14 situs asam amino yang berbeda. Tarsius dianae memiliki 12 situs asam amino (asam amino ke 2, 6, 9, 22, 23, 29, 39, 41, 42, 45, 55, 85),
T. spectrum memiliki 7 situs asam amino (asam amino ke 2, 6, 9, 41, 45, 55 dan 85) dan T. bancanus memiliki 2 situs asam amino (ke 23 dan 45) yang dapat digunakan sebagai penanda genetik. Lima asam amino unik ditemukan pada T. dianae, yaitu pada situs asam amino ke 6 (valina), ke 22 (alanina), ke 29 (alanina), ke 39 (serina) dan ke 42 (valina).
Jarak genetik berdasar nukleotida Cyt b yang dihitung menggunakan model 2 parameter Kimura ditemukan nilai paling kecil sebesar 0,7%, nilai paling besar 22,3% dan rata-rata 13,1%, sedangkan pada D-loop jarak genetik paling kecil adalah 0% dan nilai paling besar 11,8% dengan rata-rata 2,3%. Filogram menggunakan metode neighbor joining berdasar hasil penentuan runutan nukleotida dan asam amino Cyt b tersebut dapat dijadikan pembeda masing-masing spesies Tarsius, tetapi filogram berdasar runutan nukleotida pada daerah
D-loop tidak dapat digunakan untuk membedakan spesies Tarsius tersebut.
KAJIAN PENANDA GENETIK GEN CYTOCHROME B
DAN DAERAH D-LOOP PADA Tarsius sp.
OLEH :
RINI WIDAYANTI
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Primatologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Disertasi : Kajian Penanda Genetik Gen Cytochrome B dan Daerah D-loop pada Tarsius sp.
Nama : Rini Widayanti NRP : P30600001 Program Studi : Primatologi
Menyetujui,
1. Komisi Pembimbing
Dr. Ir. Dedy Duryadi Solihin, DEA
Prof. drh. Dondin Sajuthi, MST, Ph.D Dr. Ir. R.R. Dyah Perwitasari, MSc
Mengetahui,
2. Ketua Program Studi Primatologi 3. Dekan Sekolah Pascasarjana
Dr. Ir. Sri Supraptini Mansjoer Prof. Dr.Ir. Syafrida Manuwoto, MSc
SURAT PERNYATAAN
Saya menyatakan dengan sebenar-benarnya bahwa segala pernyataan dalam disertasi saya yang berjudul:
“KAJIAN PENANDA GENETIK GEN CYTOCHROME B DAN DAERAH
D-LOOP PADA Tarsius sp .”
Merupakan gagasan atau hasil penelitian disertasi saya sendiri dengan bimbingan Komisi Pembimbing, kecuali yang dengan jelas ditunjukkan rujukannya. Disertasi ini belum pernah diajukan untuk memperoleh gelar pada program sejenis di perguruan tinggi lain. Semua data dan informasi yang digunakan telah dinyatakan secara jelas dan dapat diperiksa kebenarannya.
Bogor, 26 Januari 2006
RIWAYAT HIDUP
Penulis dilahirkan di Magelang pada tanggal 25 Januari 1963 dari ayah Djuwari (alm) dan ibu Hidajati. Penulis merupakan anak ketiga dari lima bersaudara.
Pendidikan Sekolah Dasar diselesaikan pada tahun 1974 di SD Cacaban III Magelang, Sekolah Menengah Pertama lulus tahun 1977 di SMP Negeri I Magelang dan Sekolah Menengah Atas di SMA Negeri I Magelang lulus pada tahun 1981.
Sarjana Kedokteran Hewan Universitas Gadjah Mada Yogyakarta diselesaikan pada tahun 1986, selanjutnya profesi Dokter Hewan diperoleh pada tahun 1987. Pada tahun 1999 penulis menyelesaikan S2 untuk program studi Sain Veteriner di Sekolah Pascasarjana Universitas Gadjah Mada Yogyakarta. Penulis pada tahun 2000 mendapat kesempatan untuk mengikuti program S3 pada program studi Primatologi di Sekolah Pascasarjana Institut Pertanian Bogor. Beasiswa pendidikan Pascasarjana diperoleh dari Direktorat Jenderal Pendidikan Tinggi Republik Indonesia.
PRAKATA
Puji syukur ke hadirat Allah SWT penulis panjatkan atas segala karunia-Nya sehingga berhasil menyelesaikan disertasi yang berjudul “ Kajian Penanda Genetik Gen Cytochrome B dan Daerah D-loop Pada Tarsius sp.”. Disertasi
ini diajukan sebagai salah satu syarat untuk memperoleh gelar doktor pada Sekolah Pascasarjana di Institut Pertanian Bogor.
Terima kasih penulis ucapkan kepada Dr. Ir Dedy Duryadi Solihin , DEA, selaku ketua komisi pembimbing serta Prof. drh. Dondin Sajuthi, MST, Ph.D dan Dr. Ir. R.R. Dyah Perwitasari. M.Sc. selaku anggota komisi pembimbing atas waktu, bimbingan, perhatian dan nasehat yang telah diberikan kepada penulis sejak awal rencana penelitian hingga selesainya penulisan disertasi ini.
Terima kasih penulis sampaikan juga kepada Dr. Ir. Achmad Farajallah M.Si selaku penguji luar komisi pada sidang tertutup dan kepada Prof. Dr. Ir. H. Abdullah Syam, M.Sc dan Ir. Muladno, M.Sc., Ph.D selaku penguji luar komisi pada sidang terbuka atas masukan dan koreksinya, sehingga lebih sempurnanya disertasi ini.
Terima kasih juga penulis sampaikan kepada Dr. Ir. Sri Supraptini Mansjoer selaku Ketua Program Studi Primatologi dan staf pengajar Program Studi Primatologi, Dr. drh. Joko Pamungkas, M.Sc selaku Kepala Pusat Studi Satwa Primata, LPPM-IPB dan Dr. Ir Komang G Wiryawan selaku Kepala Pusat Studi Ilmu Hayati, PAU-IPB yang telah memberi ijin penulis melakukan penelitian beserta fasilitasnya di laboratorium Biologi Molekuler, PSIH-PAU, IPB. Dalam kesempatan ini pula, penulis mengucapkan terima kasih kepada bapak Drs. Yulius Duma, MSi, Dr. Saroyo dan Dr. Hengki Johannis Kiroh yang telah membantu dalam memperoleh sampel Tarsius. Penulis berterima kasih pula kepada rekan-rekan PRM, teman seperjuangan di lab (mbak Niken, dik Evi, Ari, dan Chule, bu Amin, bu Donata, pak Is, pak Asep, pak Jack, pak Sinaga, pak Heri) atas dukungan dan kerjasamanya.
Untuk mas Prapto, Galih, Wida dan Tika, ibu dan saudaraku terima kasih atas dukungan dan doanya.
DAFTAR ISI
DAFTAR GAMBAR ………..
DAFTAR TABEL ………...………..
DAFTAR LAMPIRAN ………...……..
PENDAHULUAN ………..………..
Latar Belakang ……… Ruang Lingkup Penelitian ………. Tujuan Penelitian ……… Manfaat Penelitian ………. Hipotesis ……….. TINJAUAN PUSTAKA ………..…….…
Tarsius sp. ………. Klasifikasi ………... Morfologi ……… Karakteristik dan Anatomi T. spectrum dan T. dianae ……... Ekologi ……… Penyebaran ……… Usaha Konservasi ………...……….. DNA Mitokondria (mt-DNA) ……….. Cytochrome b ……….………. D-loop ………..……….. BAHAN DAN METODE ………..
Tempat dan Waktu Penelitian ……….. Rancangan Penelitian ……… Koleksi Contoh Darah dan Otot ………... Isolasi DNA Total ……… Amplifikasi DNA dengan PCR ……….. Penentuan Runutan Nukleotida ……….……….. Analisis Data ………... HASIL DAN PEMBAHASAN ………....
DNA Total ……….…….………….……. Amplifikasi Gen Cyt b dan D-loop ……….……….. Amplifikasi Gen Cyt b ………....…..………….…….. Amplifikasi daerah D-loop ………..……….
Penentuan Runutan Nukleotida Gen Cyt b Parsial …..………... Keragaman Runutan Asam Amino ……….... Keragaman Runutan Nukleotida ……… Hubungan Kekerabatan Tarsius sp. Berdasar Runutan
Nukleotida dan Asam Amino Cyt b Parsial ……….…… Penentuan Runutan Nukleotida D-loop Parsial ……… Keragaman Runutan Nukleotida ………... Hubungan Kekerabatan Tarsius sp. Berdasar Runutan Nukleotida D-loop Parsial ……….……… Keragaman Runutan Nukleotida Gen Cyt b dan D-loop ….….… Hipotesis Terjadinya Keragaman Tarsius sp. di Sulawesi ………. Konsistensi Hubungan Kekerabatan Tarsius sp. ...……… Strategi Konservasi untuk Tarsius sp. ……… SIMPULAN DAN SARAN ………..…...
Simpulan ……….. Saran ……… DAFTAR PUSTAKA ………...………
29 29 35
40 42 42
KAJIAN PENANDA GENETIK GEN CYTOCHROME B
DAN DAERAH D-LOOP PADA
Tarsius sp.
OLEH :
RINI WIDAYANTI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ABSTRACT
RINI WIDAYANTI. The Study of Genetic Marker on Cytochrome B gene and D-loop region of Tarsius sp.. Under the direction of DEDY DURYADI SOLIHIN, DONDIN SAJUTHIand R. R. DYAH PERWITASARI.
Tarsius is an endemic species of Indonesia that is threatened. Conservation of this species would yield better results if its genetic make up and diversity determined. The objective of this research was to study the specific genetic marker on Cyt b gene and D-loop region of Tarsius sp. Sequencing of PCR product using primer H15149 on Cyt b gene yielded base sequence of 276 nts (coding 92 amino acids), while sequencing using primer DLTARPROF on D-loop resulted in base sequence of 270 nts. Results of Cyt b and D-loop fragments sequencing were put on multiple alignment with other primates from Genbank with the aid of software Genetyc-Win Version 3.0 and Clustal W, and were analyzed using MEGA program version 3.1.Fourteen different amino acid sites were found. Tarsius dianae had 12 amino acid sites (amino acid no. 2, 6, 9, 22, 23, 29, 39, 41, 42, 45, 55, and 85), T. spectrum had 7 amino acid sites (amino acid no. 2, 6, 9, 41, 45, 55 and 85) and T. bancanus had 2 amino acid sites (amino acid no. 23 and 45) which can be used as genetic marker. Five unique amino acids were found on T. dianae, such as amino acid site: 6 (valine), 22 (alanin), 29 (alanin), 39 (serin) and site 42 (valine). The genetic distance based on nucleotide Cyt b calculated using Kimura 2-parameter model indicated that in the smallest value of 0.7%, biggest 22.3% and average 13.1%, while for D-loop, the smallest genetic distance is 0% and biggest 11.8% and average 2.3%. The phylogenetic tree using neighbor Joining Method based on the sequence of nucleotide and amino acid Cyt b reveded differentiation among
ABSTRAK
RINI WIDAYANTI. Kajian Penanda Genetik Pada Gen Cytochrome B dan Daerah D-loopTarsius sp.. Dibimbing oleh DEDY DURYADI SOLIHIN, DONDIN SAJUTHI, R.R. DYAH PERWITASARI
Tarsius merupakan salah satu satwa endemik Indonesia yang keberadaannya mulai memprihatinkan. Konservasi sebagai salah satu cara untuk pelestarian satwa ini akan lebih terarah dan berhasil guna apabila karakteristik dan keragaman sumber genetiknya diketahui dengan pasti. Tujuan dari penelitian ini adalah mengkaji penanda genetik spesifik pada gen Cyt b dan daerah D-loopTarsius sp.
Penentuan runutan hasil PCR menggunakan primer H15149 pada gen
Cyt b didapatkan runutan basa sebesar 276 nt (menyandi 92 asam amino), sedangkan hasil pengurutan menggunakan primer DLTARPROF pada daerah D-loop didapatkan runutan basa sebesar 270 nt. Fragmen Cyt b dan D-loop hasil pengurutan disejajarkan berganda dengan primata lain dari data Genbank
dengan bantuan perangkat lunak Genetyx-Win versi 3.0 dan Clustal W, kemudian dianalisis dengan menggunakan program MEGA versi 3.1. Dari hasil analisis diperoleh 14 situs asam amino yang berbeda. Tarsius dianae memiliki 12 situs asam amino (asam amino ke 2, 6, 9, 22, 23, 29, 39, 41, 42, 45, 55, 85),
T. spectrum memiliki 7 situs asam amino (asam amino ke 2, 6, 9, 41, 45, 55 dan 85) dan T. bancanus memiliki 2 situs asam amino (ke 23 dan 45) yang dapat digunakan sebagai penanda genetik. Lima asam amino unik ditemukan pada T. dianae, yaitu pada situs asam amino ke 6 (valina), ke 22 (alanina), ke 29 (alanina), ke 39 (serina) dan ke 42 (valina).
Jarak genetik berdasar nukleotida Cyt b yang dihitung menggunakan model 2 parameter Kimura ditemukan nilai paling kecil sebesar 0,7%, nilai paling besar 22,3% dan rata-rata 13,1%, sedangkan pada D-loop jarak genetik paling kecil adalah 0% dan nilai paling besar 11,8% dengan rata-rata 2,3%. Filogram menggunakan metode neighbor joining berdasar hasil penentuan runutan nukleotida dan asam amino Cyt b tersebut dapat dijadikan pembeda masing-masing spesies Tarsius, tetapi filogram berdasar runutan nukleotida pada daerah
D-loop tidak dapat digunakan untuk membedakan spesies Tarsius tersebut.
KAJIAN PENANDA GENETIK GEN CYTOCHROME B
DAN DAERAH D-LOOP PADA Tarsius sp.
OLEH :
RINI WIDAYANTI
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Primatologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Disertasi : Kajian Penanda Genetik Gen Cytochrome B dan Daerah D-loop pada Tarsius sp.
Nama : Rini Widayanti NRP : P30600001 Program Studi : Primatologi
Menyetujui,
1. Komisi Pembimbing
Dr. Ir. Dedy Duryadi Solihin, DEA
Prof. drh. Dondin Sajuthi, MST, Ph.D Dr. Ir. R.R. Dyah Perwitasari, MSc
Mengetahui,
2. Ketua Program Studi Primatologi 3. Dekan Sekolah Pascasarjana
Dr. Ir. Sri Supraptini Mansjoer Prof. Dr.Ir. Syafrida Manuwoto, MSc
SURAT PERNYATAAN
Saya menyatakan dengan sebenar-benarnya bahwa segala pernyataan dalam disertasi saya yang berjudul:
“KAJIAN PENANDA GENETIK GEN CYTOCHROME B DAN DAERAH
D-LOOP PADA Tarsius sp .”
Merupakan gagasan atau hasil penelitian disertasi saya sendiri dengan bimbingan Komisi Pembimbing, kecuali yang dengan jelas ditunjukkan rujukannya. Disertasi ini belum pernah diajukan untuk memperoleh gelar pada program sejenis di perguruan tinggi lain. Semua data dan informasi yang digunakan telah dinyatakan secara jelas dan dapat diperiksa kebenarannya.
Bogor, 26 Januari 2006
RIWAYAT HIDUP
Penulis dilahirkan di Magelang pada tanggal 25 Januari 1963 dari ayah Djuwari (alm) dan ibu Hidajati. Penulis merupakan anak ketiga dari lima bersaudara.
Pendidikan Sekolah Dasar diselesaikan pada tahun 1974 di SD Cacaban III Magelang, Sekolah Menengah Pertama lulus tahun 1977 di SMP Negeri I Magelang dan Sekolah Menengah Atas di SMA Negeri I Magelang lulus pada tahun 1981.
Sarjana Kedokteran Hewan Universitas Gadjah Mada Yogyakarta diselesaikan pada tahun 1986, selanjutnya profesi Dokter Hewan diperoleh pada tahun 1987. Pada tahun 1999 penulis menyelesaikan S2 untuk program studi Sain Veteriner di Sekolah Pascasarjana Universitas Gadjah Mada Yogyakarta. Penulis pada tahun 2000 mendapat kesempatan untuk mengikuti program S3 pada program studi Primatologi di Sekolah Pascasarjana Institut Pertanian Bogor. Beasiswa pendidikan Pascasarjana diperoleh dari Direktorat Jenderal Pendidikan Tinggi Republik Indonesia.
PRAKATA
Puji syukur ke hadirat Allah SWT penulis panjatkan atas segala karunia-Nya sehingga berhasil menyelesaikan disertasi yang berjudul “ Kajian Penanda Genetik Gen Cytochrome B dan Daerah D-loop Pada Tarsius sp.”. Disertasi
ini diajukan sebagai salah satu syarat untuk memperoleh gelar doktor pada Sekolah Pascasarjana di Institut Pertanian Bogor.
Terima kasih penulis ucapkan kepada Dr. Ir Dedy Duryadi Solihin , DEA, selaku ketua komisi pembimbing serta Prof. drh. Dondin Sajuthi, MST, Ph.D dan Dr. Ir. R.R. Dyah Perwitasari. M.Sc. selaku anggota komisi pembimbing atas waktu, bimbingan, perhatian dan nasehat yang telah diberikan kepada penulis sejak awal rencana penelitian hingga selesainya penulisan disertasi ini.
Terima kasih penulis sampaikan juga kepada Dr. Ir. Achmad Farajallah M.Si selaku penguji luar komisi pada sidang tertutup dan kepada Prof. Dr. Ir. H. Abdullah Syam, M.Sc dan Ir. Muladno, M.Sc., Ph.D selaku penguji luar komisi pada sidang terbuka atas masukan dan koreksinya, sehingga lebih sempurnanya disertasi ini.
Terima kasih juga penulis sampaikan kepada Dr. Ir. Sri Supraptini Mansjoer selaku Ketua Program Studi Primatologi dan staf pengajar Program Studi Primatologi, Dr. drh. Joko Pamungkas, M.Sc selaku Kepala Pusat Studi Satwa Primata, LPPM-IPB dan Dr. Ir Komang G Wiryawan selaku Kepala Pusat Studi Ilmu Hayati, PAU-IPB yang telah memberi ijin penulis melakukan penelitian beserta fasilitasnya di laboratorium Biologi Molekuler, PSIH-PAU, IPB. Dalam kesempatan ini pula, penulis mengucapkan terima kasih kepada bapak Drs. Yulius Duma, MSi, Dr. Saroyo dan Dr. Hengki Johannis Kiroh yang telah membantu dalam memperoleh sampel Tarsius. Penulis berterima kasih pula kepada rekan-rekan PRM, teman seperjuangan di lab (mbak Niken, dik Evi, Ari, dan Chule, bu Amin, bu Donata, pak Is, pak Asep, pak Jack, pak Sinaga, pak Heri) atas dukungan dan kerjasamanya.
Untuk mas Prapto, Galih, Wida dan Tika, ibu dan saudaraku terima kasih atas dukungan dan doanya.
DAFTAR ISI
DAFTAR GAMBAR ………..
DAFTAR TABEL ………...………..
DAFTAR LAMPIRAN ………...……..
PENDAHULUAN ………..………..
Latar Belakang ……… Ruang Lingkup Penelitian ………. Tujuan Penelitian ……… Manfaat Penelitian ………. Hipotesis ……….. TINJAUAN PUSTAKA ………..…….…
Tarsius sp. ………. Klasifikasi ………... Morfologi ……… Karakteristik dan Anatomi T. spectrum dan T. dianae ……... Ekologi ……… Penyebaran ……… Usaha Konservasi ………...……….. DNA Mitokondria (mt-DNA) ……….. Cytochrome b ……….………. D-loop ………..……….. BAHAN DAN METODE ………..
Tempat dan Waktu Penelitian ……….. Rancangan Penelitian ……… Koleksi Contoh Darah dan Otot ………... Isolasi DNA Total ……… Amplifikasi DNA dengan PCR ……….. Penentuan Runutan Nukleotida ……….……….. Analisis Data ………... HASIL DAN PEMBAHASAN ………....
DNA Total ……….…….………….……. Amplifikasi Gen Cyt b dan D-loop ……….……….. Amplifikasi Gen Cyt b ………....…..………….…….. Amplifikasi daerah D-loop ………..……….
Penentuan Runutan Nukleotida Gen Cyt b Parsial …..………... Keragaman Runutan Asam Amino ……….... Keragaman Runutan Nukleotida ……… Hubungan Kekerabatan Tarsius sp. Berdasar Runutan
Nukleotida dan Asam Amino Cyt b Parsial ……….…… Penentuan Runutan Nukleotida D-loop Parsial ……… Keragaman Runutan Nukleotida ………... Hubungan Kekerabatan Tarsius sp. Berdasar Runutan Nukleotida D-loop Parsial ……….……… Keragaman Runutan Nukleotida Gen Cyt b dan D-loop ….….… Hipotesis Terjadinya Keragaman Tarsius sp. di Sulawesi ………. Konsistensi Hubungan Kekerabatan Tarsius sp. ...……… Strategi Konservasi untuk Tarsius sp. ……… SIMPULAN DAN SARAN ………..…...
Simpulan ……….. Saran ……… DAFTAR PUSTAKA ………...………
29 29 35
40 42 42
DAFTAR GAMBAR
Gambar 1. Gambar 2. Gambar 3. Gambar 4. Gambar 5. Gambar 6. Gambar 7. Gambar 8. Gambar 9. Gambar 10. Gambar 11. Gambar 12. Gambar 13. Gambar 14. Gambar 15. Gambar 16.Tarsius sp. ………..
Bentuk jumbai pada ekor T. bancanus, T. spectrum
dan T. dianae ………
Bentuk telinga pada T. spectrum dan T. dianae …...
Tiga hipotesis biogeografi Sulawesi dan sekitarnya ..
T. spectrum sebagai maskot di Sulawesi Utara ……
Susunan gen dari organisasi genom mitokondria
T bancanus ………
Skema organisasi Gen Cyt b pada T. bancanus …..
Posisi asam amino (asam amino ke 47 sampai dengan 126) hasil amplifikasi menggunakan primer L14841 dan H15149 ………
Skema organisasi daerah kontrol mt-DNA pada mamalia ………..………..
Peta pengambilan contoh T. spectrum, T. dianae
dan T. bancanus ………
Profil DNA Total Tarsius sp. menggunakan gel agarose 1,2% ………
Profil DNA Tarsius sp. hasil amplifilkasi pasangan primer L14841dan H15149 ………
Skema letak penempelan primer L14841 dan H15149 untuk mengamplifikasi daerah Gen Cyt b
pada Tarsius sp. ………..
Profil DNA Tarsius sp. hasil amplifikasi pasangan primer DLTARPROF-DLTARPHER dan
DLTARPROF-DLTARBFR ……….
Skema letak penempelan primer DLTARPROF, DLTARBFR, DLTARPHER untuk mengamplifikasi fragmen D-loopTarsius sp. ………
Skema Gen Cyt b dan sebagian daerah hasil perunutan DNA (berukuran 276 pb) yang dipakai untuk analisis keragaman genetik pada Tarsius sp.
Gambar 17.
Gambar 18.
Gambar 19.
Gambar 20.
Gambar 21.
Gambar 22.
Gambar 23.
Gambar 24.
Posisi asam amino hasil amplifikasi menggunakan primer L14841& H15149 ………...
Filogram menggunakan metode Neighbor joining dari nukleotida daerah Gen Cyt b parsial (berukuran 276 nt) Tarsius sp. hasil penelitian dengan spesies primata lain sebagai pembanding ………
Filogram menggunakan metode Neighbor joining dari asam amino sebagian daerah gen Cyt b
(berukuran 92 aa) Tarsius sp. hasil penelitian
dengan spesies primata lain ……….
Skema daerah D-loop dan sebagian daerah hasil perunutan DNA (berukuran 270 nt) yang dipakai untuk analisis keragaman genetik pada Tarsius sp.
Filogram menggunakan metode Neighbor joining
dari nukleotida daerah D-loop parsial (berukuran 270 nt).Tarsius sp hasil penelitian dengan spesies primata lain) ………..
Hipotesis terjadinya keragaman Tarsius sp. di Sulawesi ………
Deforestasi sejak masa prapertanian sampai tahun 1997 ………...
Perubahan tutupan hutan di Sulawesi ………..
33
40
41
42
45
48
54
DAFTAR TABEL
Tabel 1. Tabel 2. Tabel 3. Tabel 4. Tabel 5. Tabel 6. Tabel 7. Tabel 8. Tabel 9. Tabel 10. Tabel 11. Tabel 12. Tabel 13. Tabel 14.Data yang mendukung 16 taksa di Sulawesi ………...
Pengukuran telinga T. dianae dan T. spectrum ………
Urutan basa dan suhu penempelan primer untuk
mengamplifikasi Gen Cyt b dan D-loopTarsius sp. ……
Asam amino beserta kodon penyandi yang mengalami mutasi pada Cyt b parsial dari T. bancanus,
T. spectrum dan T. dianae hasil penelitian dan spesies primata lain ……….
Komposisi dan situs asam amino beragam pada Tarsius
hasil penelitian dan T. bancanus asal Malaysia
(Genbank) ………
Perubahan asam amino Cyt b parsial hasil penjajaran
Tarsius hasil penelitian dengan T. bancanus asal
Malaysia berdasar sifat kimianya ……….
Rata-rata komposisi asam amino hasil translasi gen Cyt b parsial T. bancanus, T. spectrum, T. dianae dan primata lain ………...……….
Rata-rata komposisi nukleotida pada gen Cyt b parsial
Tarsius sp. hasil penelitian dan T. bancanus dari
Genbank ………..………..
Jumlah dan posisi basa dari triplet kodon beragam (79 situs) gen Cyt b parsial Tarsius sp. hasil penelitian dan
T. bancanus dari Genbank ……..……..………...
Rasio transisi terhadap transversi (di bawah diagonal) dan rata-rata perbedaan asam amino Tarsius sp. dan primata lain ……….………
Rata-rata transisi basa ke 1, 2 dan 3 (di bawah diagonal) dan transversi basa ke 1, 2, 3 (di atas diagonal) setiap kodon pada Gen Cyt b parsial Tarsius dan primata lain .
Rasio antara asam amino total, asam amino kekal, asam amino sinonimus dan asam amino non sinonimus gen
Cyt b parsial Tarsius sp. ………..….
Persentase rataan komposisi nukleotida daerah D-loop
parsial (berukuran 270 nt) T. bancanus dan T.spectrum
Jarak genetik T. bancanus, T. spectrum dan T.bancanus
Tabel 15.
* berdasar runutan nukleotida D-loop parsial (berukuran 270 nt) ……….………..
Hutan alam, hutan yang sudah terdegradasi, dan kawasan yang hutannya sudah gundul pada
pertengahan 1990-an di Sulawesi ………
44
DAFTAR LAMPIRAN
Lampiran 1. Lampiran 2. Lampiran 3. Lampiran 4. Lampiran 5. Lampiran 6. Lampiran 7. Lampiran 8. Lampiran 9. Lampiran 10. Lampiran 11. Lampiran 12.Letak penempelan primer L14841 dan H15149 pada urutan basa nukleotida Gen Cyt bT. bancanus …….
Letak penempelan primer DLTARPROF, DLTARBFR pada runutan basa nukleotida daerah D-loop
T. bancanus ………..…
Penjajaran berganda nukleotida dari Cyt b parsial (276 nt) T. bancanus, T. spectrum, T. dianae hasil penelitian terhadap T. bancanus dan spesies
primata lain ……….…..
Penjajaran berganda asam amino dari Cyt b parsial (92 aa) T. bancanus, T. spectrum,T.dianae hasil penelitian terhadap T. bancanus dan beberapa spesies primata lain ……….
Jumlah penggunaan kodon dan nilai indeks
penggunaan kodon dalam mt-DNA Tarsius ………..
Keluarga asam amino berdasar rantai samping ….
Komposisi asam amino hasil translasi nukleotida gen
Cyt b parsial (92 asam amino) T. bancanus,
T. spectrum, T. dianae hasil penelitian, T. bancanus
dan N. coucang ……….………….
Matriks perbedaan nukleotida dari gen Cyt b parsial antara T. bancanus, T. spectrum, T. dianae terhadap beberapa spesies primata lain ………..
Matriks perbedaan asam amino dari Cyt b parsial (92 asam amino) antara T. bancanus, T. spectrum, T. dianae terhadap T. bancanus dan beberapa spesies primata lain ……….
Komposisi nukleotida yang menyusun DNA daerah Gen Cyt b parsial (berukuran 276 nt) T. bancanus,
T. spectrum, T. dianae hasil penelitian dan
T. bancanus * ………..………..
Jarak genetik (di bawah diagonal) menggunakan metode 2 parameter Kimura dan rasio transisi terhadap transversi (di atas diagonal) berdasarkan urutan nukleotida pada Gen Cyt b parsial (276 pb)
Penjajaran berganda dari D-loop parsial antara T. bancanus, T. spectrum terhadap primata lain ……...
Lampiran 13. Lampiran 14. Lampiran 15. Lampiran 16. Lampiran 17. Lampiran 18. Lampiran 19. Lampiran 20. Lampiran 21. Lampiran 22. Lampiran 23. Lampiran 24. Lampiran 25.
Matriks perbedaan nukleotida dari D-loop parsial (berukuran 270 nt) antara T. bancanus, T. spectrum hasil penelitian dan beberapa spesies primata …….
Persentase komposisi nukleotda yang menyusun DNA daerah D-loop parsial (berukuran 270 nt)
T. bancanus, T. spectrum hasil penelitian dan
T. bancanus* ………..……
Jarak genetik (di bawah diagonal) menggunakan metode 2 Parameter Kimura dan rasio transisi terhadap transversi (di atas diagonal) berdasar runutan nukleotida pada D-loop parsial (270 nt) ……
Runutan basa nukleotida Gen tRNA Glu, Cyt b, tRNAThr, tRNAPro, daerah D-loop dan tRNAPhe dari Tarsius bancanus ………
Runutan basa nukleotida Gen tRNA Glu, Cyt b, tRNAThr, tRNAPro, daerah D-loop dan tRNAPhe dari Nycticebus coucang ……….
Runutan basa nukleotida Gen tRNA Glu, Cyt b, tRNAThr, tRNAPro, daerah D-loop dan tRNAPhe dari Cebus albifrons ……….
Runutan basa nukleotida Gen tRNA Glu, Cyt b, tRNAThr, tRNAPro, daerah D-loop dan tRNAPhe dari Macaca sylvanus ………..
Runutan basa nukleotida Gen tRNA Glu, Cyt b, tRNAThr, tRNAPro, daerah D-loop dan tRNAPhe dari Gorilla gorilla ……….
Runutan basa nukleotida Gen tRNA Glu, Cyt b, tRNAThr, tRNAPro, daerah D-loop dan tRNAPhe dari Pan troglodytes ……….
Runutan basa nukleotida Gen tRNA Glu, Cyt b, tRNAThr, tRNAPro, daerah D-loop dan tRNAPhe dari Pan paniscus ……….
Runutan basa nukleotida Gen tRNA Glu, Cyt b, tRNAThr, tRNAPro, daerah D-loop dan tRNAPhe dari Hylobates lar ………...
Runutan basa nukleotida Gen tRNA Glu, Cyt b, tRNAThr, tRNAPro, daerah D-loop dan tRNAPhe dari Homo sapiens ………
Runutan basa nukleotida Gen tRNA Glu, Cyt b, tRNAThr, tRNAPro, daerah D-loop dan tRNAPhe dari Lemur catta ………..
Lampiran 26.
Lampiran 27.
Runutan basa nukleotida daerah D-loopLagothrix lagotricha ………..
Data spesies primata yang digunakan dalam analisis kekerabatan Tarsius ………..
103
PENDAHULUAN
Latar BelakangIndonesia merupakan salah satu negara yang memiliki ragam jenis primata terkaya di dunia. Dari sekitar 195 jenis primata yang ada, 40 jenis ditemukan di Indonesia dan 24 jenis di antaranya merupakan satwa yang hanya ditemukan di Indonesia (endemik). Salah satu jenis satwa endemik yang hidup di Indonesia adalah Tarsius sp. dari genus Tarsius, famili Tarsiidae (Supriyatna & Wahyono, 2000).
Genus Tarsius berdasar data morfologi yang didukung dengan data vokalisasi dikelompokkan ke dalam lima spesies, yaitu Tarsius pumilus, T. dianae, T. spectrum, T. bancanus dan T. syrichta (Musser dan Dagosto 1987).
Empat spesies diantaranya terdapat di Indonesia dengan daerah penyebaran Sumatera dan Kalimantan (T. bancanus), Sulawesi (T. spectrum, T. pumilus dan
T. dianae) dan satu spesies ada di Filipina (T. syrichta). Menurut Shekelle dan Leksono (2004) sampai saat ini telah ditemukan 16 populasi Tarsius di Sulawesi yang kemungkinan dapat menjadi spesies sendiri dan baru lima spesies di antaranya sudah mempunyai nama (T. spectrum, T. dianae, T. pumilus, T. sangiriensis dan T. pelengensis). Satwa ini memiliki daya tarik tersendiri karena memiliki bola mata yang besar, tubuh yang sangat kecil dan warna yang menarik. Dalam usaha mempertahankan keberadaan Tarsius sp. sebagai sumber keragaman hayati Indonesia dan ditunjang dengan statusnya sebagai satwa langka, maka pemerintah telah lama mengkategorikan satwa ini sebagai satwa yang dilindungi sejak tahun 1931 yaitu berdasar Peraturan Perlindungan Binatang Liar tahun 1931 dan Peraturan Pemerintah No. 7 Tahun 1999. Meskipun dalam daftar buku yang dikeluarkan International Union for Conservation Nature and Natural Resources (IUCN) karena data yang kurang, maka status konservasi satwa ini adalah “Data Deficient”.
Dalam upaya mengetahui sejarah perkembangan yang pasti dari
Penelitian tentang eksplorasi molekuler untuk analisis genom sitoplasma yaitu mitokondria berupa runutan utuh dari DNAnya, telah dilakukan pada
T. bancanus oleh Schmitz et al. (2002). Dengan diketahuinya runutan utuh DNA mitokondria pada T. bancanus maka terbuka kesempatan untuk mengkaji penanda genetik untuk setiap gen penyandi protein maupun bagian yang tidak menyandi protein pada spesies lain dari Tarsius ini. Fragmen penyandi protein di dalam genom mitokondria yaitu Cytochrome b (Cyt b), Cytochrome oxsidase-c dan gen lainnya banyak digunakan untuk penelitian mengenai hubungan spesies dari genus atau famili yang sama, seperti yang dilakukan oleh Ozawa et al.
(1997) yaitu pada gajah Asia dan gajah Afrika; Randi dan Lucchini (1998) pada genus Alectoris. Namun demikian penelitian keragaman daerah penyandi protein untuk melihat hubungan antar spesies pada Tarsius sp. belum dilakukan.
Fragmen bukan penyandi protein di dalam genom mitokondria yang sering dipakai dalam penelaahan keragaman genetik dan hubungan kekerabatan di antara spesies hewan adalah daerah D-loop. Daerah D-loop ini menarik untuk ditelaah karena dua dari ketiga domainnya memiliki mutasi yang tinggi sehingga perubahan runutan basa-basa nukleotidanya terjadi tidak saja pada tingkatan inter-spesies tetapi juga pada tingkatan intra-spesies. Dengan demikian penelaahan daerah D-loop banyak digunakan untuk penelitian biologi populasi dan evolusi hewan. Misalnya, seperti yang dilakukan oleh Randi et al. (1997) yaitu pada populasi Capreolus pygargus dan C. capreolus; Fumagalli et al.
(1996) pada tupai; Savolaenin et al. (2000) pada anjing domestik dan srigala; Wilkinson dan Chapman(1991) pada Nycticebus humeralis; Casane et al. (1997) pada lagomorph dan Tamura (2000) untuk mempelajari evolusi pada manusia. Namun demikian penelaahan keragaman genetik daerah bukan penyandi ( D-loop) pada Tarsius sp. belum dilakukan.
Pemanfaatan penanda genetik pada Tarsius sp. diharapkan dapat membantu dalam melengkapi data yang kurang sehingga dapat menunjang usaha konservasi.
Ruang Lingkup Penelitian
Penelitian yang dilakukan adalah mengkaji penanda genetik pada genom mitokondria dari T. bancanus, T. spectrum dan T. dianae dengan menganalisis runutan DNA mitokondria pada gen penyandi protein (Cyt b) dan pada daerah
Untuk menganalisis runutan gen penyandi Cyt b dan daerah D-loop yang terdapat pada DNA mitokondria digunakan teknik Polymerase Chain Reaction
(PCR) yang dilanjutkan dengan penentuan runutan DNA memakai alat “automatic sequencing” (ABI Prism versi 341 dan ALF Express II). Contoh
Tarsius diperoleh dari beberapa lokasi yang secara alami dihuni oleh spesies
Tarsius seperti: T. spectrum (Tangkoko dan Air Madidi, Sulawesi Utara),
T. dianae (Kamarora, Sulawesi Tengah) dan T. bancanus diperoleh dari Lampung, Sumatera Selatan.
Tujuan Penelitian
Penelitian ini bertujuan untuk mengkaji penanda genetik DNA mitokondria (gen penyandi Cyt b dan D-loop) dari T. spectrum; T. dianae dan T. bancanus.
Manfaat Penelitian
Penelitian ini diharapkan dapat membantu memperjelas pengelompokan spesies Tarsius yang ada di Indonesia. Informasi tersebut dapat dipakai dalam usaha pengelolaan program penangkaran atau konservasi.
Hipotesis
Penanda genetik spesifik pada gen Cyt b dan D-loop T. spectrum, T dianae dan T. bancanus memiliki fragmen DNAunik dan mencirikan
TINJAUAN PUSTAKA
Tarsius sp.
[image:31.612.133.265.157.318.2]Klasifikasi
Gambar 1. Tarsius sp. (Abdullah 2000)
Tarsius syrichta terdapat di Filipina, T. spectrum ditemukan di hutan primer maupun hutan sekunder di Tangkoko-Batuangus Sulawesi Utara yang terletak pada ujung sebelah utara dari semenanjung utara Sulawesi pada ketinggian 500 m di atas permukaan air laut. Tarsius spectrum juga ditemukan di Sulawesi Selatan, pulau Togian dan pulau kecil lainnya (Musser dan Dagosto 1987). Tarsius pumilus ditemukan endemik di Sulawesi Tengah yaitu di gunung Rano-rano (ketinggian 1800 m), di gunung Rorekatimbu Sulawesi Tengah (ketinggian 2200 m), dan di hutan gunung Latimojong Sulawesi Selatan pada ketinggian sekitar 2200 m (Maryanto dan Yani 2000, Jones et al. 2004). Tarsius dianae ditemukan di hutan primer tidak jauh dari batas sebelah utara Taman Nasional Lorelindu pada 001o10’S, 120 09’E, 2 km di sebelah Tenggara
Kamarora pada ketinggian 700 m di atas permukaan laut (Niemitz et al. 1991), dan T. bancanus ditemukan di Sumatera Selatan, Borneo dan sekitar Bangka dan Belitung (Musser dan Dagosto 1987). Menurut Shekelle (2003) sampai saat ini telah ditemukan 16 populasi Tarsius di Sulawesi yang kemungkinan dapat menjadi spesies sendiri dan baru lima spesies di antaranya yang sudah mempunyai nama yaitu T. spectrum, T. sangiriensis, T. pumilus, T. pelengensis
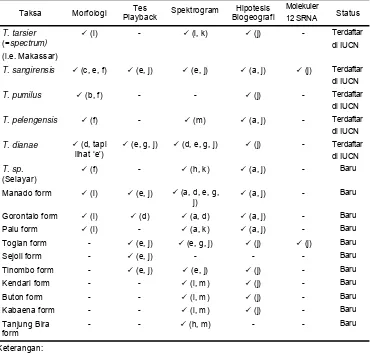
dan T. dianae. Sebelas spesies lainnya masih perlu pemberian nama untuk keperluan konservasi, seperti terlihat pada Tabel 1.
Tarsius sp. atau monyet hantu diklasifikasikan dalam Kelas Mammalia, Ordo Primata, Subordo Prosimian,
Superfamili Tarsoidea, Famili Tarsiidae,
Tabel 1. Data yang mendukung 16 Taksa di Sulawesi (Shekelle 2003)
Taksa Morfologi Tes
Playback Spektrogram
Hipotesis Biogeografi
Molekuler
12 SRNA Status
T. tarsier
(=spectrum)
(i.e. Makassar)
9 (l) - 9 (l, k) 9 (j) - Terdaftar di IUCN
T. sangirensis 9 (c, e, f) 9 (e, j) 9 (e, j) 9 (a, j) 9 (j) Terdaftar di IUCN
T. pumilus 9 (b, f) - - 9 (j) - Terdaftar di IUCN
T. pelengensis 9 (f) - 9 (m) 9 (a, j) - Terdaftar di IUCN
T. dianae 9 (d, tapi lihat ‘e’)
9 (e, g, j) 9 (d, e, g, j) 9 (j) - Terdaftar di IUCN
T. sp.
(Selayar)
9 (f) - 9 (h, k) 9 (a, j) - Baru
Manado form 9 (l) 9 (e, j) 9 (a, d, e, g, j)
9 (a, j) - Baru
Gorontalo form 9 (l) 9 (d) 9 (a, d) 9 (a, j) - Baru Palu form 9 (l) - 9 (a, k) 9 (a, j) - Baru Togian form - 9 (e, j) 9 (e, g, j) 9 (j) 9 (j) Baru Sejoli form - 9 (e, j) - - - Baru Tinombo form - 9 (e, j) 9 (e, j) 9 (j) - Baru Kendari form - - 9 (i, m) 9 (j) - Baru Buton form - - 9 (i, m) 9 (j) - Baru Kabaena form - - 9 (i, m) 9 (j) - Baru Tanjung Bira
form
- - 9 (h, m) - - Baru
Keterangan: - : belum diteliti 9 : sudah diteliti oleh:
(a) MacKinnon dan MacKinnon (1980)
(f) Groves (1998) (k) Shekelle (in review)
(b) Musser danDagosto (1987) (g) Nietsch dan Kopp (1998) (l) Groves (in review) (c) Feiler (1990) (h) Nietsch dan Babo (2001) (m) Nietsch (pers. comm.) (d) Niemitz et al. (1991) (i) Nietsch dan Burton
(2002)
(e) Shekelle et al. (1997) (j) Shekelle (2003)
Di Indonesia Tarsius memiliki bermacam-macam nama lokal. Tarsius spectrum disebut juga Tangkasi (Minahasa), Ngasi (Sulteng), Tanda-bona Passo (Wana), Podi (Tolaki), Wengu (Mornene), Tenggahe (Sangir) dan Tanda-bana
(Sulut). Tarsius bancanus disebut juga Kera buku, Singapuar (Bengkulu), Krabuku (Lampung), Palele (Belitung), Mentiling ingkir, Ingkit, Linseng (Ngaju),
Page (Tidung), Makikebuku (Karimata), Singaholeh (Kutai), Tempiling (Kalbar),
Morfologi
Tarsius sp. merupakan primata yang mempunyai ukuran tubuh paling kecil dan dapat hidup sampai berumur sekitar 13,5 tahun. Diberi nama Tarsius
karena mempunyai tulang tarsal panjang membentuk pergelangan kaki yang
mampu meloncat hingga 3 meter dari pohon ke pohon. Tarsius dapat memutar kepalanya hingga 180 derajat. Tibia dan fibula pada bagian bawah bergabung menjadi satu, berfungsi sebagai alat penahan goncangan pada saat hewan
meloncat dari pohon ke pohon. Jari kaki dan jari tangan sangat panjang dan
ramping, sehingga Tarsius dapat berpegangan erat pada pohon dan ranting.
Tarsius memiliki tungkai belakang yang pendek dan tangan yang kecil. Jari kaki dan tangan memiliki bantalan, tetapi pada T. pumilus jari tangan mengalami reduksi dalam ukurannya sehingga T. pumilus memiliki genggaman yang lebih bagus daripada spesies Tarsius lainnya. Tarsius pumilus juga memiliki ujung kuku yang perluasannya melebihi tepi bantalan jari. Pada jari kaki kedua dan
ketiga memiliki cakar yang dipergunakan untuk grooming, sedangkan jari lainnya memiliki kuku. Bola mata sangat besar dan setiap mata memiliki lapisan
postorbital di belakangnya, yang akan melindungi bola mata dari musculus temporalis yang sangat kuat ke arah samping. Telinga sangat lebar dan aktif (mobile), memiliki gigi taring yang besar dan tajam (Musser dan Dagosto 1987).
Kunci identifikasi Tarsius (Storr 1780) menurut Niemitz dan Verlag (1984) adalah sebagai berikut: Tarsius spectrum (Pallas 1778) mempunyai ciri-ciri antara lain muka seperti Galago senegalensis, ekor berambut dengan panjang jumbai kurang-lebih 110 mm, panjang rambut 5-12 mm dan terdapat juga
kelompok rambut pendek memiliki sisik menyerupai struktur kulit ekor. Tarsius pumilus (Miller dan Hollister 1921) mempunyai ciri-ciri antara lain hewan dewasa memiliki panjang kepala-badan 95-105 mm, panjang kranial maksimum 31mm,
lebar kranial maksimum 26,3 mm, kulit bagian tarsal berambut dengan baik, dan
terdapat titik warna kekuning-kuningan dibelakang telinga. Tarsius bancanus
(Horsfield 1821) mempunyai ciri-ciri antara lain panjang kaki belakang antara
59-74 mm, panjang ekor antara 180-245 mm, jumbai pada ekor berkembang
baik, rambut jumbai memiliki panjang sekitar 7 mm dan kulit bagian tarsal
ditumbuhi rambut dengan baik. Tarsius syrichta (Linnaeus 1758) memiliki ciri- ciri antara lain kaki belakang panjangnya antara 56-69 mm, panjang ekor antara
200-240 mm, jumbai pada ekor tidak berkembang baik, rambut pada jumbai
pendek dan sangat sedikit. Tarsius bancanus, T. spectrum dan T. dianae dapat dilihat pada Gambar 2. Menurut Supriatna dan Wahyono (2000), ukuran tubuh
[image:34.612.203.433.135.389.2]T. bancanus T. spectrum T. dianae
Gambar 2. Bentuk jumbai pada ekor T. bancanus, T. spectrum dan T. dianae (Shekelle 2003)
T. dianae sama dengan T. spectrum, namun sedikit lebih besar dibanding dengan T. pumilus. Panjang badan antara 110-120 mm, panjang ekor antara 215-225 mm, berat badan antara 95-110 gram, warna kulit keabu-abuan dengan
bintik-bintik hitam pada kedua sisi, hidung sedikit menonjol dibanding
T. spectrum dan bagian atas bibir terdapat rambut putih yang tumbuh. Tarsius
sangiriensis memiliki warna rambut abu-abu kekuningan, menyerupai
T. spectrum; panjang tubuh antara 115-125 mm, panjang ekor 225-240 mm, jumbai ditumbuhi rambut yang tumbuhnya jarang, berat tubuh 110-120 gram,
telinga besar bila dibandingkan dengan kepala.
Secara morfologi, antara T. bancanus dan Tarsius sp. yang berasal dari Sulawesi dapat dibedakan dengan melihat panjang jumbai pada ekor dan dari
lebar telinga, namun diantara spesies Tarsius yang ada di Sulawesi sangat sulit
untuk dibedakan. Gambar 3, memperlihatkan perbedaan telinga antara
T. bancanus T. spectrum T.dianae
Gambar 3. Bentuk telinga pada T. bancanus , T. spectrum dan T. dianae (Stevan Merker)
Karakteristik dan Anatomi T. spectrum dan T. dianae.
Rambut abu-abu kekuningan dengan spot hitam T. dianae ditemukan di kedua sisi hidung dan tampak sangat jelas apabila dibanding pada T. spectrum.
Rambut pendek keputihan ditemukan di kedua sisi bibir atas dan di bagian
tengah bibir bawah. Ada bagian yang tak berambut pada bagian bawah telinga
(pada T. spectrum tidak dijumpai). Terdapat celah di bagian tengah yang membagi hidung menjadi dua belahan. Pada saat pelupuk mata tidak terbuka
secara penuh bentuk mata tampak lebih tidak simetris dibanding dengan
T. spectrum. Kuku jari berwarna gelap dan tajam (tirus). Mata lebih tertarik ke arah samping (kurang pada T. spectrum). Warna rambut pada tubuh hampir sama, tetapi T. dianae memiliki pigmen yang lebih gelap pada ekor, jari kaki, jari tangan dan kuku daripada T. spectrum. Kulit bersisik di bawah ekor pada
T. dianae berwarna coklat, sedangkan pada T. spectrum tidak berwarna. Kuku tangan T. dianae lebih sempit dan lebih melengkung daripada T. spectrum
(Niemitz et al. 1991).
Telinga T. dianae lebih pendek dan lebih sempit dibanding T. spectrum.
Tabel 2: Pengukuran telinga T. dianae dan T. spectrum (Niemitz et al. 1991)
Deskripsi, Pengukuran (mm) T. dianae T. spectrum
1. Panjang telinga dari ujung pinna ke dalam incisura intertagrica
2. Panjang telinga dari ujung pinna sampai titik paling basal dari telinga luar
3. Lebar telinga yang terbesar
4. Panjang maksimum daerah tak berambut dari ujung tragus ke titik paling basal dari telinga luar
5. Area yang paling lebar
6. Kedalaman incisura intertagrica
7. Lebar incisura intertagrica
28,8
31,7
19,3
7,9
6,0
4,1
2,0
32,2
33,5
23,4
5,0
3,6
3,6
1,0
Index: Lebar x 100____ Panjang maksimum
60,8 70,0
Ekologi
Tarsius sp. hidup di hutan tropis, hutan primer, hutan sekunder, hutan bambu, semak-semak, perkebunan dan kadang hidup di perkampungan. Tarsius
hidup berkelompok, dengan jumlah anggota antara 2-6 individu. Komposisi
kelompok bervariasi dari hanya 2 individu (1 ekor jantan dan 1 ekor betina), 1
ekor jantan dewasa, 2 ekor betina dewasa dan anak-anaknya. Struktur sosial
sebagian besar monogami dan hanya sedikit yang poligami (Gursky 1995).
Penyebaran
Data sebaran biogeografi primata dan kodok (Macaca dan Bufo) di Sulawesi, menurut Evans et al. (2003) dapat digunakan untuk memperkirakan 7 daerah endemisitas (oleh Shekelle dan Leksono dianggap 8 daerah
endemisitas). Menurut Mac Kinnon dan Mac Kinnon (1980) daerah endemisitas
tersebut masih ditambah daerah yang tidak dihuni Macaca tetapi dihuni Tarsius
yaitu kepulauan Sangihe, Kepulauan Togian, Kepulauan Banggai, pulau Selayar
dan pulau Kabaena. Menurut Shekelle dan Leksono (2004) diperkirakan ada 13
Menurut Hall (2001), di zaman Miosen sampai dengan Pleistosen
Sulawesi merupakan kepulauan yang berasal dari beberapa daratan Asia,
Australia dan daratan yang timbul dari dasar lautan. Melalui proses pergeseran
lempeng (playtectonic) daratan tersebut (disebut “microplates”) membentuk pulau Sulawesi sekarang (Gambar 4b). Pulau Sulawesi sekarang, diduga selesai
terbentuk pada zaman Pleistosen yaitu kira-kira 1-2 juta tahun yang lalu.
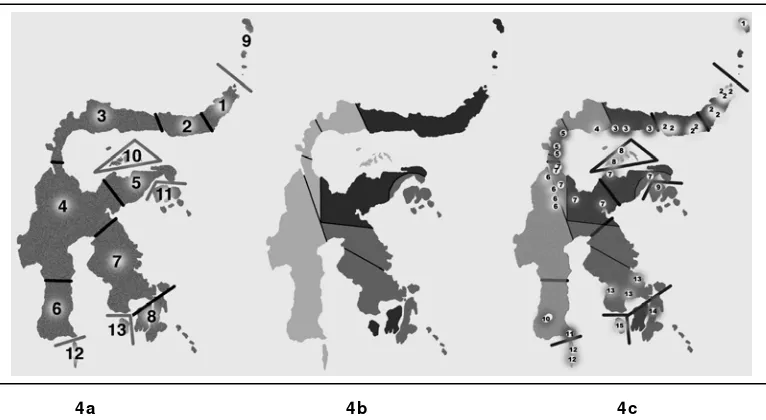
Menurut Shekelle dan Leksono (2004), daerah sebaran Tarsius
mempunyai banyak kesamaan dengan sebaran hipotesis biogeografi berdasar
data biologi (Gb 4a) dan mempunyai banyak kesamaan dengan “microplates”
Sulawesi seperti terlihat pada gambar 4b. Daerah sebaran Tarsius di Sulawesi
[image:37.612.129.512.267.477.2]4a 4b 4c
Gambar 4. Tiga Hipotesis Biogeografi Sulawesi dan Sekitarnya (Shekelle dan Leksono 2004)
4a : Berdasarkan Data Biologi, 4b : Berdasarkan Data Geologi,
4c : Hipotesis hibrid (Biologi dan Geologi) dengan data daerah sebaran Tarsius
Keterangan: (kiri) Daerah 1-8 berdasarkan data genetik Macaca dan Bufo (Evans et al. 2003). Daerah 9-13 berdasarkan hipotesis MacKinnon dan MacKinnon (1980) merupakan daerah yang dihuni Tarsius tetapi tidak dihuni Macaca yang secara alami mempunyai daerah endemisitas sendiri. (tengah) Rekonstruksi geologi peristiwa tektonik dari zaman Cenozoik bahwa Sulawesi terbentuk dari kepulauan dengan beberapa asal (abu-abu muda = Asia, abu-abu sedang = Australia, abu-abu tua =daratan yang timbul dari dasar lautan). (kanan) Hipotesis 4a di padukan dengan 4b dan dibandingkan dengan data sebaran kelompok akustik Tarsius (Shekelle dan Leksono 2004).
Keterangan Gambar 4c.(1) T. sangiriensis, (2) Manado form, (3) Gorontalo form, (4) Sejoli form, (5) Tinombo form, (6) Palu form, (7) T. pumilus, (8) Togian form, (9)
T. pelengensis, (10) T. spectrum, (11) Tanjung Bira form, (12) Tarsius sp. (Selayar), (13) Kendari form, (14) Buton form, (15) Kabaena form.
dan sekitarnya sangat sesuai dengan kombinasi hipotesis biogeografi berdasar
yang berarti hipotesis biogeografi Sulawesi secara menyeluruh. Hipotesis
berdasar data biologi berasumsi bahwa spesies-spesies yang lain pindah ke
Sulawesi hampir bersama dengan Macaca dan Bufo, sedangkan hipotesis berdasar data geologi berasumsi bahwa spesies-spesies lain pindah ke Sulawesi
pada saat proses pergeseran lempeng dimulai.
Menurut Shekelle (2003), berdasar pada data molecular clock
kedatangan bajing diperkirakan 11,5 juta tahun yang lalu dan evolusi semua
spesies Macaca dari Afrika Utara sampai Sumbawa terjadi kurang dari 7 juta tahun yang lalu. Berdasar data jarak genetik, kedatangan Tarsius ke Sulawesi diperkirakan pada zaman pertengahan Miosen (sekitar 15 juta tahun yang lalu)
yang hampir bersama dengan kedatangan bajing. Oleh karena itu menurut
Shekelle dan Leksono (2004), hipotesis hibrid biogeografi lebih sesuai untuk
sebaran kelompok Tarsius dan merupakan hipotesis yang paling masuk akal dibanding dengan hipotesis biografi lainnya.
Usaha Konservasi
Sulawesi memiliki luas 187,882 km2 dan merupakan pulau terbesar dan
terpenting di daerah biogeografi Wallacea. Ditinjau dari sejarah geologi,
pulau Sulawesi diduga tidak pernah bersatu dengan daratan manapun (Hall
2001). Keadaan terisolasi dalam waktu yang lama menimbulkan terjadinya
evolusi pada berbagai spesies, sehingga satwa di pulau Sulawesi mempunyai
tingkat endemisitas yang tinggi. Tingkat endemisitas paling tinggi terjadi pada
taksa vertebrata. Pada mamalia, 61% dari 127 jenis yang ada di Sulawesi
bersifat endemik. Sedangkan pulau Kalimantan yang mempunyai endemisitas
paling tinggi di daratan Asia hanya 18% mammalia yang bersifat endemik
(Whitten et al. 1987).
Daerah Wallacea mempunyai 529 spesies vertebrata endemik (1,9% dari
jumlah di dunia) dan spesies-spesies ini mengalami ancaman yang serius karena
hanya 15% habitat alami yang masih tersisa. Sekitar 39,2% dari habitat alami
yang tersisa terdapat dalam kawasan konservasi. Habitat alami yang masih
tersisa tersebut hanya akan efektif untuk melindungi biodiversitas di Sulawesi jika
tersebar sesuai dengan distribusi, sehingga upaya konservasi di Sulawesi harus
dirancang secara komprehensif (Shekelle dan Leksono 2004).
Menurut Shekelle dan Leksono (2004), data biogeografi dapat membantu
daerah endemisitas di Sulawesi. Sebaran daerah endemisitas ini sangat penting
untuk merancang kawasan konservasi yang sesuai dengan sebaran daerahnya.
Selain itu, upaya konservasi juga dapat dibantu dengan menggunakan hewan
maskot. Hewan maskot biasanya berupa hewan yang mempunyai karisma yang
mempunyai nilai tertentu seperti bentuk yang lucu, langka, unik, mudah diingat,
berukuran besar dan endemis. Tarsius tidak berukuran besar tetapi dijadikan maskot di Sulawesi (terutama di Sulawesi Utara, dapat dilihat pada Gambar 5),
[image:39.612.248.356.231.390.2]karena (1) mempunyai sebaran yang luas, sampai ke pulau-pulau sekitar
Gambar 5. T. spectrum sebagai maskot di Sulawesi Utara
Sulawesi, (2) mempunyai banyak taksa endemik yang tersebar di hampir seluruh
daerah endemisitas, (3) berada di lebih banyak tipe habitat, (4) bukan
merupakan hama sehingga tidak mengancam produk pertanian, (5) tidak
mempunyai nilai ekonomi, misalnya daging atau bagian tubuh lainnya (kecuali
sebagai hewan peliharaan), (6) merupakan hewan yang karismatik. Menurut
Shekelle dan Leksono (2004), program yang akan dilakukan untuk membantu
usaha konservasi adalah melakukan pemberian nama pada 11 spesies Tarsius
baru, yaitu dengan menangkap Tarsius hidup-hidup dan memeliharanya dalam kandang sehingga memungkinkan semua untuk melakukan penelitian, pelatihan,
pendidikan, penangkaran dan sebagai sumber analisis genetik, sebelum hewan
tersebut mati untuk dijadikan contoh spesimen.
DNA Mitokondria (mt-DNA)
Mitokondria merupakan organela di luar inti sel yang mengandung sekitar
16,5 x 103/3,3 x 109 pbdari DNA yang ditemukan di dalam sel manusia (Wertz
dari sampel biologis dalam jumlah kecil atau dari sampel biologis yang sudah
terdegradasi adalah lebih besar daripada DNA inti karena molekul mt-DNA
terdapat dalam ratusan sampai ribuan kopi dibanding dengan DNA inti yang
hanya dua kopi pada setiap selnya. Oleh karena itu, otot, tulang, rambut, kulit,
darah dan cairan tubuh lainnya dapat digunakan sebagai sumber materi untuk
penentuan lokus mt-DNA apabila terjadi degradasi oleh karena peralatan atau
karena waktu.
Runutan mt-DNA dipilih sebagai penanda genetik karena berukuran relatif
kecil dan jumlah kopi yang banyak sehingga mudah didapat dari sel, diturunkan
dari induk betina (maternal) dan beberapa gen dalam mitokondria mutasinya
lebih cepat daripada gen inti karena rendahnya aktivitas proof reading dari γ DNA
polymerase selama replikasi di dalam mitokondria (Wertz 2000). Menurut Reyes
et al. (1998), mutasi pada genom mitokondria juga karena kurangnya sistem reparasi DNA dan adanya kerusakan oksidatif yang disebabkan oleh radikal
bebas, hasil selama transpor elektron ke oksigen pada rantai respirasi kompleks
di membran dalam mitokondria. Menurut Majerus (1996) genom mitokondria
mempunyai kecepatan evolusi 5-10 kali lebih cepat daripada genom inti, dan
satu bagian dari genom mitokondria yang disebut D-loop mengalami evolusi sepuluh kali lebih cepat lagi.
Genom mt-DNA mamalia berbentuk sirkuler, beruntai ganda, memiliki
panjang sekitar 16,5 kb yang mengandung basa guanine (G) dan cytosine (C)
berkisar antara 32-45,6%. Kedua basa G dan C terdistribusi secara tidak merata
diantara kedua untai DNA. Berdasar kandungan basa guaninnya, mt-DNA dibagi
menjadi 2 untai yaitu, untai yang kaya G disebut untai berat (heavy strand) dan yang mengandung sedikit G disebut untai ringan (light strand) (Reyes et al.
1998).
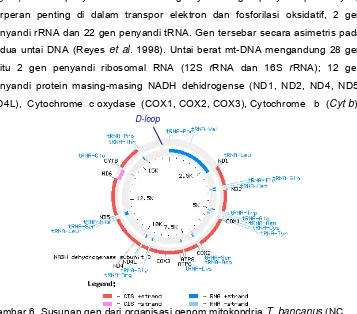
Berdasarkan data dari GenBank, organisasi genom mitokondria pada primata yang telah diketahui urutan nukleotidanya masing-masing memiliki
jumlah dan susunan gen yang sama dengan mamalia lain, sedangkan
perbedaan yang ada hanya jumlah nukleotida penyusun mt-DNA. Jumlah
nukleotida penyusun genom mt-DNA T. bancanus adalah 16927 pb (GenBank
NC 002811), Gorilla gorilla 16364 pb (GenBank NC 001645), Pan troglodytes
Cebus albifrons 16554 pb (GenBank NC 002763) dan Homo sapiens 16571 pb (GenBank DQ 246833). Susunan gen dari organisasi genom mitokondria
T. bancanus dapat dilihat pada Gambar 6.
Berdasar jenis gennya, genom mitokondria dibagi menjadi 2 bagian, yaitu
daerah penyandi (coding region) dan daerah bukan penyandi (non coding region). Daerah penyandi terdiri dari 37 gen yaitu 13 gen penyandi protein yang berperan penting di dalam transpor elektron dan fosforilasi oksidatif, 2 gen
penyandi rRNA dan 22 gen penyandi tRNA. Gen tersebar secara asimetris pada
kedua untai DNA (Reyes et al. 1998). Untai berat mt-DNA mengandung 28 gen yaitu 2 gen penyandi ribosomal RNA (12S rRNA dan 16S rRNA); 12 gen penyandi protein masing-masing NADH dehidrogense (ND1, ND2, ND4, ND5,
[image:41.612.144.501.179.493.2]ND4L), Cytochrome c oxydase (COX1, COX2, COX3), Cytochrome b (Cyt b),
Gambar 6. Susunan gen dari organisasi genom mitokondria T. bancanus (NC 002811).
ATPase (ATP6, ATP8) dan 14 gen penyandi tRNA masing-masing fenilalanina (tRNAPhe), valina (tRNAVal), leusina (tRNALeu), isoleusina (tRNAIle), metionina (tRNAMet), triptofana (tRNATrp), asam aspartat (tRNAAsp), lisina (tRNALys), glisina (tRNAGly), arginina (tRNAArg), histidina (tRNAHis), serina (tRNASer), leusina (tRNALeu) dan treonina (tRNAThr). Sedangkan untai ringan mt-DNA mengandung sisanya (9 gen) yaitu, 1 gen penyandi protein yaitu NADH dehidrogense6 (ND6)
dan 8 gen penyandi tRNA yaitu asam glutamat (tRNAGlu), prolina (tRNAPro), serina (tRNASer), tirosina (tRNATyr), sistina (tRNACys), asparagina (tRNAAsn), alanina (tRNAAla) dan glutamina (tRNAGln). Daerah bukan penyandi genom
mitokondria hanya terdiri dari daerah kontrol (control region) (Reyes et al. 1998; Schmitz et al. 2002).
Cyt b
Gen penyandi Cyt b mempunyai ukuran 1140 pb, terletak diantara gen penyandi tRNAGlu (di sebelah kiri atau depan) dan gen penyandi tRNAThr (di sebelah kanan atau belakang) pada mt-DNA (Gambar 7) (Schmitz et al. 2002). Adanya variasi urutan pada Cyt b menyebabkan gen ini banyak digunakan untuk membandingkan spesies dalam genus yang sama atau famili yang sama (Randi
1996).
Protein Cyt b dibagi dalam 3 domain fungsional, yaitu intermembran, matriks dan transmembran. Domain intermembran terbentang antara membran
dalam dan luar mitokondria, mengalami evolusi yang lebih lambat daripada ke 2
domain fungsional lainnya. Keadaan ini diduga karena fungsinya sebagai pusat
redox Qo (Irwin et al. 1991 dan Griffiths 1997). Domain intermembran pada mamalia mengandung 105 residu asam amino dengan komposisi terbesar dari
ND6 tRNA Pro
Cyt b (1140 pb)
5’ 3’
tRNAGlu tRNAThr
Gambar 7. Skema organisasi gen Cyt b pada T. bancanus (Howell 1989 dan Schimtz et al. 2002)
asam aminonya adalah yang berhubungan dengan fungsi pusat redox Qo
(Zhang et al. 1998 diacu dalam McClellan and McCracken 2001) dan sekitar 29% bersifat kekal pada metazoa (Degli Esposti et al. 1993 diacu dalam McClellan
Residu asam amino ke 20 – 40
Residu asam amino ke 130 - 150
and McCracken 2001). Domain matriks berlokasi pada permukaan dalam dari
membran dalam mitokondria, terdiri dari 65 residu asam amino dan proporsi
terbesar adalah residu asam amino polar dan asam amino dasar (Griffiths 1997
diacu dalam McClellan and McCracken 2001). Pada bagian matriks relatif sedikit
residu yang bersifat kekal kemungkinan karena sebagian besar bagian dari
domain ini tidak diketahui fungsinya. Domain transmembran terdiri dari protein
Cyt b yang ditransfer ke membran dalam mitokondria, pada mammalia mengandung 209 residu asam amino yang sebagian besar termasuk asam
amino hidrofobik. Penggantian asam amino pada daerah transmembran terjadi
antara asam amino hidrofobik leusina, isoleusina dan valina (Irwin et al., 1991; Kornegay et al. 1993 diacu dalam McClellan dan McCracken 2001), dan 19% dari residu asam amino ini bersifat kekal pada sebagian besar metazoa (Degli
Esposti et al. 1993 diacu dalam McClellan and McCracken, 2001). Banyak dari residu tersebut dikaitkan dalam fungsinya untuk ligasi heme, aktivitas redoks
atau stabilitas struktural.
Menurut Howell (1989), hasil analisis urutan asam amino pada golongan
prokariotik dan eukariotik disimpulkan terdapat 5 region protein yang terdiri atas
sekitar 20 residu asam amino yang bersifat kekal selama evolusi. Dua region
protein yang sangat kekal diperkirakan terdapat pada residu protein ke 130 - 150
dan residu protein ke 270 – 290. Kedua region protein ini berlokasi pada loop
ekstramembran dan diduga merupakan bagian pusat reaksi Qo. Region residu
protein ke 20 – 40 yang merupakan bagian dari pusat reaksi Qi juga bersifat
kekal di dalam tingkat spesies.
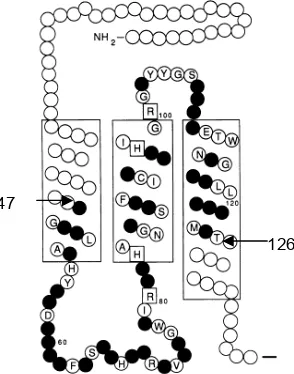
Runutan DNA yang menyandi 80 asam amino (posisi ke 47 sampai
dengan ke 126) dari Cyt b rodensia, burung, ikan dan manusia (hasil amplifikasi menggunakan primer L14841 dan H15149) setelah disejajarkan berganda
didapatkan 129 posisi nukleotida beragam dari 240 nukleotida. Komposisi
nukleotida dari ke 4 contoh individu tersebut kandungan Gnya adalah yang
paling sedikit, seperti yang dilaporkan pada mt-DNA vertebrata. Perubahan
nukleotida yang terjadi pada interspesies sebagian besar adalah terjadi secara
substitusi transisi, sedangkan untuk antar genera dalam famili atau ordo yang
sama perubahan terjadi sebagian besar adalah transversi. Menurut hipotesis
struktural Cyt b, runutan DNA hasil amplifikasi primer L14841 dan H15149 terletak pada intermembran mitokondria, yaitu dengan ditandai tidak adanya
posisi ke 83 yaitu histidina (H), posisi ke 97 yaitu H, dan posisi ke 100 adalah R,
Tidak adanya penggantian keempat asam amino ini disebabkan fungsi asam
amino histidina dalam ligasi heme di domain intermembran (Gambar 8) (Kocher
et al. 1989). Berdasar penjajaran urutan basa gen Cyt b, daerah tersebut dapat memberikan informasi filogenetik pada tingkat intraspesies sampai tingkat antar
genera.
Menurut Schmitz et al. (2002), hasil analisis asam amino pada primata dan nonprimata menunjukkan bahwa primata besar memiliki afinitas yang lebih
tinggi basa G+C sedangkan Tarsius dan Slow loris memiliki lebih banyak basa A+T pada gen yang menyandi asam amino seperti yang tampak pada non
[image:44.612.233.380.279.466.2]primata.
Gambar 8. Posisi 80 asam amino (asam amino ke 47 sampai dengan 126) hasil amplifikai menggunakan primer L14841 dan H15149.
Keterangan: asam amino variatif ; asam amino tidak dirunut;
(ada huruf di dalamnya) asam amino tidak variatif diantara vertebrata; asam amino kekal yang berhubungan dengan fungsi
Cyt b (Kocher et al. 1989)
D-loop
Daerah bukan penyandi (non coding region) merupakan genom yang tidak membawa urutan informasi untuk pembentukan protein maupun RNA.
Tingginya angka dari polimorfisme nukleotida atau adanya perbedaan runutan
pada kedua bagian hipervariabel dari non coding region digunakan untuk membedakan di antara individu dan atau sampel biologis (Melton 1999).
Daerah bukan penyandi (non coding region) atau disebut juga daerah kontrol (control region) atau D-loop terletak di antara gen penyandi tRNAPro (di
sebelah kiri atau depan) dan gen penyandi tRNAPhe (di sebelah kanan atau belakang). Jumlah dan runutan nukleotida penyusun D-loop dari beberapa spesies primata dapat dilihat pada Lampiran 16 –24 dan 26.
D-loop merupakan bagian dari mt-DNA yang sangat variatif dalam substitusi nukleotida, insersi atau delesi pendek (indels) dan memiliki variable number tandem repeat (VNTRs) yang dinamis yang terletak pada bagian yang hipervariatif dan domain yang khusus (Fumagalli et al. 1996). Daerah D-loop
dibagi menjadi 3 domain, yaitu domain I yang berbatasan dengan tRNAPro, terdiri
dari runutan yang diasosiasikan dengan termination of H-strand replication (TAS)
yang sering mengandung VNTRs (R1 repeats), domain II yang terdapat di bagian
sentral dan bersifat kekal, terdapat conserved sequence block (CSB B, C, D,E dan F) dan domain III yang berbatasan dengan tRNAPhe, terdiri dari runutan yang
variabilitasnya tinggi karena substitusi nukleotida, insersi dan delesi (indels) serta
VNTRs (R2 repeat) dan runutan nukleotida bersifat kekal yang merupakan
promotor untuk transkripsi untai berat (heavy strand) dan untai ringan (light strand) berturut-turut HSP dan LSP, titik awal replikasi strand H (OH) dan daerah blok runutan pendek yang kekal (short conserved sequence block) (CSB1, CSB2 dan CSB3) (Sbisa et al. 1997 diacu dalam Randi dan Lucchini 1998). Menurut Southern et al. (1988), blok runutan kekal (Conserved sequence block, CSB) merupakan daerah yang homologinya tinggi yang ditemukan pada mt-DNA
lumba-lumba, sapi, manusia dan tikus. Variasi ukuran panjang dari D-loop mt-DNA disebabkan oleh variasi jumlah runutan kopi berulang (sequence tandem repeat) (Fumagalli et al. 1996). Menurut hasil penelitian yang dilakukan oleh Wilkinson dan Chapman (1991), bahwa adanya runutan nukleotida yang berbeda
panjangnya antara anak dan induk evening bat disebabkan oleh duplikasi atau delesi runutan sebesar 81pb yang berulang dengan kopi sebanyak 5-8 kali di
daerah D-loop. Demikian juga menurut Greenberg et al. (1983), runutan nukleotida di sekitar daerah awal replikasi untai H pada manusia lebih dari 96%
perubahan basanya adalah transisi dan variasi panjang nukleotida disebabkan
oleh penambahan atau delesi mono atau dinukleotida dalam repeat yang
berurutan.
Pada vertebrata, susunan dari repeat tersebut ditemukan dalam 5 posisi,
membentuk untai tiga D-loop. RS3, RS4 dan RS5 berada di ujung 3’ dari CR,
upstream dari titik awal replikasi untai H (Hoelzel et al. 1994). Susunan pada RS1, RS2, RS4 dan RS5 memiliki kopi antara 40-160 pb, sedangkan pada RS3
memiliki kopi yang lebih pendek yaitu antara 6-22 pb dengan jumlah
pengulangan yang lebih tinggi (lebih dari 40) dan tingkat heteroplasmiknya juga
lebih tinggi. Susunan kopi berulang pada RS3 ditemukan pada beberapa
mamalia yaitu, anjing dan srigala (Savolainen et al. 2000), pada 14 spesies tupai (Fumagalli et al. 1996), babi (Ghivizzani et al. 1993), kelinci (Mignotte et al.
1990), Lagomorph (Casane et al. 1997), Tupaia belangeri (Schmitz et al. 2000),
Nycticebus coucang (GenBank NC 002765) dan T. bancanus (Schmitz et al.
2002).
Gambar 9. Skema orga