EKSPRESI GEN CmBG1 MELON (Cucumis melo L. ‘TACAPA’) TERHADAP PENGARUH LAHAN KRITIS KARST
Ganies Riza Aristya1)*, Yuanita Rachmawati2), dan Budi Setiadi Daryono1)
1)Laboratorium Genetika, Fakultas Biologi Universitas Gadjah Mada, 2)
Mahasiswa Pascasarjana Program Studi Biologi, Fakultas Biologi Universitas Gadjah Mada
*e-mail : [email protected]
ABSTRAK
CmBG1, pengkode enzim β-Glucosidase peregulasi hormon asam absisat (ABA) pada Cucumis melo L. ABA berperan dalam respons adaptif tumbuhan terhadap cekaman abiotik. Lahan kritis karst memberikan cekaman pada tumbuhan berupa dehidrasi dan hambatan penyerapan nutrisi. Gen CmBG1 dapat digunakan sebagai indikator molekular suatu tanaman berada dalam kondisi cekaman. Melon unggul TACAPA Lab. Genetika Fakultas Biologi UGM potensial untuk dibudidayakan di lahan kritis karst. Penelitian ini dilakukan untuk membandingkan ekspresi gen CmBG1 secara kualitatif dan kuantitatif melon TACAPA yang ditanam pada medium tanah karst dari wilayah Agroekosistem II dan III Yogyakarta. cDNA library diperoleh dari reverse transcription isolat RNA. cDNA diamplifikasi dengan primer spesifik, kemudian dispektrofotometri pada λ260 nm untuk mengetahui konsentrasi gen CmBG1. Ekspresi gen dianalisis dengan real time PCR dengan gen referensi Cm-actin. Uji kualitatif dilakukan dengan elektroforesis gel agarosa 1,5%. Band CmBG1 TACAPA terdeteksi pada semua perlakuan dengan ukuran ±1258 bp dan sebagai kontrol Cm-actin menunjukkan band DNA sebesar ±445 bp. Konsentrasi dan ekspresi gen CmBG1 melon TACAPA yang ditanam pada medium tanah karst jauh lebih besar dibandingkan tanah kontrol.
Kata kunci: Ekspresi gen, CmBG1, melon TACAPA, real time PCR
PENDAHULUAN
Balitbang Pertanian tahun 1997 menunjukkan bahwa terdapat kurang lebih 158.600 ha lahan kritis yang tersebar pada tiga Zona Agroekosistem di Daerah Istimewa Yogyakarta (DIY), yaitu Agroekosistem II, III, dan Va. Dua diantara tiga agroekosistem ini berjenis perbukitan karst, yaitu Agroekosistem II meliputi Perbukitan Gunungsewu, Gunungkidul dan Agroekosistem III meliputi Perbukitan Dlingo Bantul serta Perbukitan Sentolo Kabupaten Kulon Progo. Apabila lahan kritis wilayah agroekosistem tersebut tidak segera diberdayakan, akan dapat memberikan dampak semakin buruk terhadap kualitas lahan.
Tanah-tanah yang dibentuk di atas lahan induk seperti batuan kapur, kapur dan endapan glasial, mengandung kalsium karbonat (CaCO3) dengan kadar tinggi dan
tanah ini cenderung memiliki pH sekitar 7
atau lebih [1] sehingga dapat memberikan cekaman pada tanaman. Karst merupakan kawasan yang memiliki karakteristik relief dan drainase spesifik dan unik, karena sistem drainase kawasan karst didominasi oleh air bawah permukaan yang sebagian besar masuk ke jaringan bawah tanah sehingga pada musim penghujan, air tidak dapat tertahan pada permukaan dan langsung masuk ke dalam tanah [2]. Kondisi ini juga memberikan cekaman dehidrasi pada tanaman karena air tidak dapat terserap secara optimum oleh tanaman disebabkan langsung masuk ke dalam jaringan bawah tanah.
dan perkecambahan seperti didiskusikan pada [3, 4, 5]. ABA juga berperan dalam mediasi adaptif tanaman terhadap pengaruh cekaman biotik maupun abiotik [6, 7]. Ekspresi BG1 pada Citrulus lanatus (ClBG1) yang diberi perlakuan cekaman kekeringan menunjukkan peningkatan yang signifikan dibandingkan tanaman kontrol pada saat fase akhir perkembangan yaitu 25 Day After Full Bloom (DAFB) [3].
Beberapa jenis tanaman termasuk melon, dapat tumbuh dan bertahan pada kondisi yang tidak optimum bagi pertumbuhannya. Hal ini disebabkan terdapat beberapa mekanisme atau strategi, dapat dilakukan oleh suatu organisme dalam lingkungan tercekam, antara lain: Strategi pelarian (Escaping), penghindaran (Avoidance), dan toleransi (tolerance) [8, 9, 10, 11].
Melon TACAPA merupakan melon klimakterik hasil pemuliaan tanaman.
Dihasilkan dari ♀A4 yang merupakan melon
komersial dengan cita rasa manis dan ♂PI
371795 dengan karakter buah tidak manis, namun memiliki ketahanan terhadap jamur tepung (powdery midew). Melon kultivar TACAPA memiliki sifat unggul hasil penggabungan sifat dari induknya, yaitu bentuk buah bulat lonjong dengan net halus, daging buah berwarna hijau kekuningan, rasa manis, dan tahan terhadap jamur tepung powdery mildew [12, 13, 14, 15]. Melon ini sedang dikembangkan untuk tahan diberbagai lahan kritis diantaranya lahan kritis karst dan lahan labil berabu-vulkanik. Berdasarkan sifat unggul yang dimiliki oleh Melon TACAPA, sangat dimungkinkan melon ini untuk dibudidayakan lebih luas untuk memenuhi permintaan pasar melon yang setiap tahun semakin meningkat.
Pada penelitian ini dilakukan penanaman Melon TACAPA pada greenhouse KP4 menggunakan perlakuan media tanah karst wilayah lahan kritis Gunungkidul, Dlingo, dan Sentolo. Kemudian dilakukan karakterisasi molekular. Karakter molekular ditentukan dengan analisis kualitatif dan kuantitatif ekspresi gen CmBG1 yang dilakukan untuk mengetahui pengaruh lahan kritis karst terhadap ekspresi gen tersebut. Deteksi gen diperlukan sebagai penanda molekular pengkode Asam Absisat (ABA) yang berperan dalam ketahanan tanaman terhadap faktor stress abiotik.
Metode Penelitian Waktu dan Tempat
Penelitian dilaksanakan pada 2014 di greenhouse KP4 UGM di Berbah Sleman Yogyakarta. Penelitian skala laboratorium dilakukan di Lab. Genetika Fakultas Biologi, Lab. FALITMA Fakultas Biologi, Lab. Biologi dan Molekular Fakultas Kedokteran Umum, dan LPPT di Universitas Gadjah Mada Yogyakarta.
Skala Greenhouse
Percobaan dilakukan dengan 4 macam perlakuan media tanam Kontrol (K) tanah dari KP4, Gunungsewu (G) tanah dari Gunungkidul pada koordinat BT: 110o32’15.7” LS: 08o03’36.7” dengan elevasi 265 m dpl, Dlingo (D) BT: 110o27’56.2” LS:
07o55’58.7” Elevasi: 230 m dpl, dan
Sentolo(S) BT: 110o11’11.3” LS:
07o49’56.5” beserta ketinggian pada Elevasi:
116 m dpl, dengan 4 kali ulangan. Penyiraman dilakukan 3 kali seminggu, pemberian pupuk NPK 2 kali dalam sebulan.
Skala Molekular
Cm-actin 5’-TGGCTGGAATAGAACTT CTGGGC-3’ (AB640865) [16]. Dilakukan dalam 40 siklus dengan optimasi temperatur sebagai berikut: 50OC - 2 menit, 95OC - 10 menit, 95OC - 1 menit, 60OC - 1 menit.
Analisis Data
Analisis kualitatif dengan melihat pita atau band. Gen CmBG1 akan menunjukkan band DNA dengan ukuran ±1258 bp. Analisis kuantitatif dengan melihat absorbansi DNA dan RNA pada panjang gelombang (λ) 260 nm dengan rasio kemurnian dengan perbandingan nilai absorbansi A260/A280 nm pada kisaran nilai
1,8 - 2,0 [17]. Ekspresi gen CmBG1 dianalisis menggunakan software CFX Manager 3.1 Bio Rad Real Time PCR (q-PCR). Ekspresi gen dianalisis dengan
menggunakan relative quantitation dengan metode compare Ct method/Ct).
Hasil dan Pembahasan Spektrofotometri
Kemurnian DNA dan RNA dianggap tinggi pada rasio pengukuran 1,8 - 2,0 [17]. Nilai rasio DNA kurang dari 1,8 menunjukkan adanya kontaminasi dari protein dan fenol sedangkan nilainya bila lebih dari 2,0 menunjukkan adanya kontaminasi RNA [18]. Pada penelitian, rasio kemurnian dan konsentrasi isolat RNA, cDNA library yang dihasilkan dari reverse transcription, serta hasil amplifikasi gen
CmBG1 dengan template cDNA library
tersaji pada Tabel 1.
Tabel 1. Hasil spektrofotometri rasio dan konsentrasi isolat RNA, cDNA, dan PCR product pada melon kultivar TACAPA
Perlakuan
Hasil Isolat RNA cDNA Hasil PCR
Rasio Konsentra
si (µg/ml) Rasio
Konsentrasi
(µg/ml) Ratio
Konsentra si (µg/ml)
Kontrol (K) 1.856 186.7 1.417 1031.5 1.602 578.5
Gunungsewu (G) 2.512 93.3 1.500 1670.2 1.602 598.4
Dlingo (D) 1.876 131.3 1.542 2524.9 1.611 743.4
Sentolo (S) 2.175 87.5 1.593 3757.4 1.683 749.8
Hasil isolat RNA perlu diukur konsentrasinya untuk menentukan apakah hasil tersebut memungkinkan untuk dibentuk cDNAnya. Hasil uji kuantitatif spektrofotometri yang didapat pada rasio kemurnian isolat RNA berkisar antara 1,2 hingga 2,5. Nilai kemurnian RNA yang seharusnya didapat berdasarkan [17] adalah 1,8 - 2.0. Pada uji kuantitatif hasil spekrofotometri, hasil yang didapatkan masih banyak kultivar yang rasio isolat RNA di bawah nilai 1,8. Hal ini besar kemungkinannya bahwa pada isolat RNA banyak mengandung kontaminan DNA dan protein serta fenol. Sementara Hasil uji kuantitatif spektrofotometri yang didapat tentang rasio kemurnian cDNA berkisar antara 1,4 hingga 1,5 dan rasio kemurnian hasil PCR 1,6. Hasil yang didapatkan ini dapat dikatakan seragam, karena tidak jauh berbeda satu sama lain. Berbeda bila kita bandingkan dengan hasil isolat RNA yang
memiliki rentang/range perbedaan yang lebih besar.
Hasil konsentrasi cDNA yang diperoleh berkisar antara 1031.5 hingga 3757.4 µg/mL. Hasil konsentrasi cDNA yang didapatkan jauh lebih besar dibandingkan dengan konsentrasi RNA yang didapatkan. Hal ini disebabkan pada proses reverse transcription menggunakan kit Fermentas, ditambahkan sequence DNA berupa primer Oligo dT sejumlah 1µL, kemudian dilakukan proses amplifikasi dengan PCR.
penelitian pada Citrulus lanatus oleh Li et al. [3] bahwa terjadi peningkatan ekspresi gen ClBG1 selama akhir fase pematangan dalam kondisi cekaman kekeringan. Peningkatan ekspresi gen ini cerminan dari proses mediasi respon adaptif terhadap stress abiotik, walaupun belum banyak diteliti tentang sejauh apa gen tersebut mempengaruhi respon adaptif terhadap stress abiotik Li et al. [3]. Tanah Dlingo dan Sentolo memberikan stress yang lebih besar bila dibandingkan
dengan Gunungsewu. Hal ini ditunjukkan dengan relatif lebih tingginya konsentrasi gen CmBG1 pada kultivar melon yang ditanam pada lahan tersebut.
Deteksi CmBG1
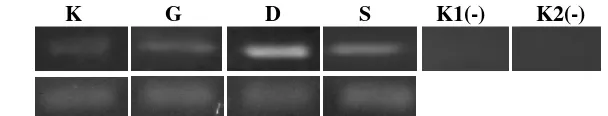
Hasil PCR gen CmBG1 kemudian dirunning menggunakan elektroforator untuk mendeteksi band yang terbentuk. Hasil deteksi gen CmBG1 secara kualitatif tersaji pada Gambar 1.
K G D S K1(-) K2(-) CmBG1 (±1258 bp)
CmActin (±445 bp)
Gambar 1. Hasil deteksi gen CmBG1 Melon TACAPA dengan kontrol CmActin pada gel agarosa 1,5%. Keterangan gambar K: Kontrol, G: Gunungsewu, D: Dlingo, S: Sentolo, K1(-): Kontrol negatif, Mentimun Cucumis sativus, K2(-): Kontrol negatif, Melon dari Isolasi DNA
Hasil yang didapatkan menunjukkan terbentuk band dengan ukuran ±1258 bp pada semua perlakuan baik kontrol maupun perlakuan lahan kritis. Hal ini disebabkan karena semua tanaman mengekspresikan hormon ABA, namun ekspresi dari peran ABA pada kultivar melon yang ditanam pada kondisi normal (kontrol) tentu akan berbeda dengan kultivar melon yang ditanam dengan cekaman lahan kritis (Gunungsewu, Dlingo, dan Sentolo). Secara hipotesis, ekspresi gen CmBG1 akan meningkat pada tanaman yang diberi cekaman atau stress. Hasil melalui uji kualitatif tidak dapat menunjukkan seberapa besar gen tersebut diekspresikan, oleh karena
itu selanjutnya dilakukan uji kuantitatif dengan Real Time PCR.
Pada uji kualitatif gen CmBG1 digunakan kontrol negatif untuk mengetahui spesifisitas primer. Kontrol Negatif 1 adalah sampel dari genus yang sama yaitu mentimun (Cucumis sativus). Mentimun diisolasi RNA kemudian di RT-PCR dan dilakukan amplifikasi gen CmBG1. Kontrol Negatif 2 yaitu template dari isolat DNA melon. Tidak terbentuk band untuk hasil PCR dari kedua kontrol negatif ini. Hal ini menunjukkan bahwa primer yang digunakan memang spesifik untuk isolat RNA tanaman melon (Cucumis melo L.).
Real time PCR
Tabel 2. Hasil nilai Cq, relative quantities (RQ) metode Pfaffl dan Livak 2-ΔΔCq, serta efisiensi reaksi melon kultivar TACAPA pada nilai Treshold 106,433
Target Sampel RQ (Metode
Pfaffl) Cq
Efisiensi Reaksi
RQ (Metode Livak 2-ΔΔCq)
Actin Actin (referensi) 1 22.86
CmBG1 Kontrol (Kalibrator) 1 28.42 1
CmBG1 D.T 56139.92 12.64 1.9997 56266.94
CmBG1 S.T 17925.90 14.29 2.0000 17928.91
CmBG1 G.T 1827.35 17.58 1.9994 1833.01
CmBG1 K.T 16.40 24.38 1.9984 16.45
Keterangan:
Kuantitas mRNA dalam sel merupakan parameter jumlah gen yang terekspresi. Untuk menganalisa tingkat ekspresi gen, cDNA yang telah disintesis dari mRNA diuji secara kuantitatif menggunakan real-time PCR. Tabel 2 di bawah ini memberikan gambaran nilai Cq, relative quantities metode Pfaffl dan Livak, serta efisiensi reaksi. Nilai Cq didapat dari jumlah siklus pada proses
PCR yang berpotongan dengan garis threshold. Angka relative quantities menunjukkan jumlah gen relatif yang terekspresi dalam seluruh proses PCR. Efisiensi amplifikasi (E) menjelaskan berapa banyaknya target yang diproduksi dalam setiap siklus PCR. Jika proses PCR efisien 100%, maka setiap siklus akan memproduksi dua kali lipat hasil dari template semula.
Gambar 2. Cq pada siklus amplifikasi melon kultivar TACAPA pada perlakuan media tanam tanah kontrol, Gunungsewu, Dlingo, dan Sentolo. Garis merah tanpa point menunjukkan amplifikasi gen referensi Actin. Gambar 2 menunjukkan pada kultivar
TACAPA, media tanam yang menginduksi ekspresi gen CmBG1 paling cepat ditunjukkan perlakuan Dlingo ditambah dengan nilai RQ yang didapat adalah yang paling besar. Sementara TACAPA pada media tanam kontrol memiliki ekspresi gen paling lambat dengan jumlah ekspresi gen CmBG1 paling sedikit seluruh perlakuan.
Hasil data ekspresi gen CmBG1 menggunakan analisis kuantitatif real time PCR secara umum menunjukkan bahwa konsentrasi gen CmBG1 pada kultivar TACAPA yang ditanam pada media kontrol memiliki nilai yang lebih rendah bila dibandingkan media tanam dengan perlakuan lahan kritis baik Gunungsewu, Dlingo, maupun Sentolo. Hasil ini senada dengan data kuantitatif menggunakan spektrofotometri. Li et al. [3] menyatakan bahwa terjadi peningkatan ekspresi gen ClBG1 pada semangka selama akhir fase pematangan dalam kondisi cekaman kekeringan. CmBG1 adalah gen peregulasi hormon ABA pada Cucumis melo L. Sun et al., [4] menyatakan bahwa ekspresi gen CmBG1 konsisten dengan akumulasi jumlah
ABA. ABA juga berperan serta toleransi adaptif terhadap stress abiotik [3, 19]. ABA berperan dalam meregulasi terbuka dan tertutupnya stomata dengan regulasi kanal ion [19]. Mekanisme tersebut dapat mencegah transpirasi air berlebih, pada saat kondisi cekaman berlangsung.
Peningkatan ekspresi gen CmBG1 menunjukkan akumulasi peningkatan ABA pada tanaman. Hal tersebut akan memicu tanaman untuk menjaga kadar air agar tetap optimum dalam tubuh tanaman dengan melakukan regulasi stomatal aperture. Pada penelitian ini, kondisi stress air pada tanaman terbukti meningkatkan ekspresi gen CmBG1.
Melalui hasil real time PCR yang menjelaskan ekspresi gen CmBG1 dapat diketahui bahwa perlakuan Dlingo dan Sentolo memberikan stress yang lebih besar bila dibandingkan dengan Gunungsewu. Hal ini ditunjukkan dengan hasil relative quantitative ekspresi gen CmBG1 yang lebih besar bila dibandingkan dengan perlakuan tersebut. Dengan demikian, tanaman yang ditanam pada lahan kritis Dlingo dan Sentolo memerlukan perlakuan yang lebih dari lahan kritis Gunungsewu atau lahan bukan kritis.
Simpulan
Gen CmBG1 dengan ukuran ±1258 bp terdeteksi pada semua kultivar. Konsentrasi gen CmBG1 pada kultivar TACAPA, induk TACAPA dan hasil persilangan TACAPA pada media kontrol lebih rendah bila dibandingkan media tanam dengan perlakuan tanah karst baik Gunungsewu, Dlingo, maupun Sentolo. Hasil ini sama dengan uji ekspresi gen CmBG1 menggunakan analisis kuantitatif real time PCR. Perlakuan Dlingo dan Sentolo memberikan stress yang lebih besar bila dibandingkan dengan Gunungsewu, sehingga tanaman yang ditanam pada lahan kritis Dlingo dan Sentolo memerlukan perlakuan yang lebih dari lahan kritis Gunungsewu atau lahan bukan kritis. Langkah-langkah konservasi lahan kritis dengan tepat akan dapat mendayagunakan dan memaksimalkan potensi yang ada baik potensi lahan maupun potensi tanaman, khususnya melon.
Ucapan Terima Kasih
Ucapan terima kasih tim peneliti berikan kepada Kementrian Riset dan Teknologi Republik Indonesia (Kemenristek) atas bantuan dana penelitian yang diberikan melalui program INSINAS (RT-2014-123).
Daftar Pustaka
[1] Sutara, P.K., (1996), Struktur Anatomi Eichornia crassipes (Mart.) Solm. dan Pistia stratiotes L. Akibat Pemberian Naphtol dan Kalsium Karbonat, Tesis Fakultas Matematika dan Ilmu Pengetahuan Alam Universitas Gadjah Mada Yogyakarta.
[2] Nahdi, M.S., (2012), Konservasi Ekosistem Lahan Kritis Berbasis Kearifan Masyarakat di Kawasan Imogiri Yogyakarta, Disertasi Fakultas Biologi Universitas Gadjah Mada Yogyakarta.
[3] Li, Q., Ping, L., Liang S., Yanping, W., Kai, J., Yufei, S., Shengjie, D., Pei C., Chaorui, D., & Ping, L., (2012), Expression Analysis of β -galactosidase that Regulate Abscisic Acid Homeostatis during Watermelon (Citrulus lanatus) Development and Under Stress Condition, Journal of Development and Ripening, Journal Plant Growth Regulation, DOI 10.1007/s00344-012-9293-5.
[5] Ren, J., Sun, L., Wu, J.F., Zhao, S.L.,Wang, C.L., Wang, Y.P., (2010), Cloning and Expression Analysis of
cDNAs for ABA8’-Hydroxylase
during Sweet Cherry Fruit Maturation and Under Stress Conditions, Journal of Plant Physiology 167:1486–1493. [6] Chernys, J.T., Zeevaart, J.A.D., (2000),
Characterization of The 9-Cis-Epoxycarotenoid Dioxygenase Gene Family and The Regulation of Abscisic Acid Biosynthesis in Avocado, Journal of Plant Physiology 124:343-353. [7] Zhu, J.K., (2002), Salt and drought
stress signal transduction in plants, Annu. Rev. Plant Biol. 53:247-273. [8] Morgan, J.M., (1984), Osmoregulation
and Water Stress in Higher Plants, Annual Review Plant Physiology, Journal of Plant Molecular Biology 35:299-319.
[9] Mooney, H.A., Pearcy, R.W., & Ehleringer, J., (1987), Plant Physiology Ecology Today, Journal of Bioscience 7:18-20.
[10] Ingram, J. & Bartels, D., (1996), The Molecular Basis of Dehydration Tolerance in Plants, Annual Review Plant Physiology, Journal of Plant Molecular Biology 47:377-403.
[11] Chaves, M.M., Maroco, J.P., & Pereira, J.S., (2003), Understanding Plant Responses to Drought from Genes to The Whole Plant Function, Journal of Plant Biology 30:239-264. [12] Aristya, G.R., (2006), Skrining dan
Pewarisan Sifat Ketahanan Tanaman Melon (Cucumis melo L.) terhadap Powdery Mildew (Jamur Tepung), Skripsi Fakultas Biologi Universitas Gadjah Mada Yogyakarta.
et Shiskoff) pada Tanaman Melon (Cucumis melo L.), Tesis Fakultas Biologi Universitas Gadjah Mada Yogyakarta.
[14] Daryono, B.S. & Qurrohman, M.T., (2009), Pewarisan Sifat Ketahanan Melon (Cucumis melo L.) Terhadap Powdery Mildew (Podosphaera xantii (Castag.) Braun et Shishkoff), Jurnal Perlindungan Tanaman Indonesia 15(1):1-6.
[15] Qurrohman, M.T., (2011), Analisis Keterpautan Gen Ketahanan terhadap Powdery Mildew pada Tanaman Melon (Cucumis melo L.) Hasil Test Cross dengan Penanda Sequence Characterized Amplified Region (SCAR), Tesis Fakultas Biologi Universitas Gadjah Mada Yogyakarta.
[16] Okuda, S., Okuda, M., Sugiyama, M., Sakata, T., Takeshita, M., & Iwai, H., (2011), Screening for Resistance Melon Germplasms to Cucurbit Chlorotic Yellows Virus Transmitted
by Bemisia tabaci.
(http://www.ncbi.nlm.nih.gov
/nuccore/AB640865). Diakses pada 26 Juni 2013.
[17] Sambrook, J. & Russel, D.W., (2001), Molecular Cloning: A Laboratory Manual 3rd Ed. Volume 3, New York: Cold Spring Harbour.
[18] Clark, D.P., (2010), Molecular Biology: Academic Cell Update, USA: Elsevier Inc.
[19] Hubbard, K. E., Nishimura, N. & Hitomi, K. 2010. Early Abscisic Acid Signal Transduction Mechanisms: Newly Discovered Components and Newly Emerging Questions. Genes Dev. 24:1695-1708.