67
PENGARUH PENAMBAHAN EKSTRAK ENZIM CAIRAN RUMEN DOMBA PADA
KOMPONEN SERAT KASAR, KANDUNGAN ASAM FITAT
TEPUNG DAUN LAMTORO GUNG (Leucaena leucocephala)
The Effect of Addition Sheep Rumen Liquor Enzyme Extract
On Fiber Component and Fitate Acid Content Leucaena Leaf Meal
Indira Fitriliyani
11
Jurusan Budidaya Perairan Fakultas Perikanan Ilmu Kelautan Universitas
Lambung Mangkurat Banjarmasin,
Coresspondent : Email : [email protected]
ABSTRACT
The aim of this experiment was to evaluate the nutrient quality of leucaena leaf meal
(LLM) with addition of sheep rumen liquor enzyme for nile tilapia feed which incubated
24 hours (in vitro); This experiment was designed in completely randomized design with
6 treatments and 3 replications each with different level of enzyme addition (0; 20; 40;
60; 80; and 100 ml/kg LLM). Results of the experiment showed that nutrient quality of
LLM with addition of sheep rumen liquor enzyme which incubated for 24 hours, where
significantly affected (P<0.05) on decrease of crude fiber (53,640%), NDF, ADF
component and phytic acid (68,088%). Over all conclusion is a great potential for using
sheep rumen liquor enzyme for improving nutrition quality of leucaena leaf meal
Keywords:
leucaena leaf meal (LLM), sheep rumen liquor enzyme, nutrient quality
.
PENDAHULUAN
Tepung
daun
lamtoro
gung
(selanjutnya disingkat TDL) merupakan
sumber daya hayati lokal yang potensial
untuk digunakan sebagai salah satu
sumber protein nabati dalam pakan ikan.
Hal ini disebabkan tingginya kandungan
protein yaitu sekitar 34,38 %, komposisi
asam amino yang hampir seimbang
dengan bungkil kedelai serta
merupakan sumber vitamin A dengan
-karoten yang relatif tinggi
serta kandungan xantofil yang merupakan
sumber pigmentasi pada kulit dan kuning
telur (Agbede dan Aletor, 2004) ; 25
-30% (NAS 1984); 24,2% (Sutardi 1981);
24% (Scott
et al
1982). Tanaman ini
dapat menghasilkan bahan kering dari
unsur-unsur yang dapat dimakan sebesar
6-8 ton per hektar per tahun atau sekitar
20 - 80 ton bahan segar (NAS, 1994). Di
Indonesia tanaman leguminosa ini mudah
ditanam
sehingga
dapat
membantu
penyediaan
pakan
secara
kontinyu
sepanjang tahun.
Pemanfaatan bahan baku pakan
ikan nila dari daun tumbuhan khususnya
daun lamtoro gung salah satunya dibatasi
oleh
kandungan
yang
tinggi
dari
komponen
neutral detergent fiber
(NDF)
68
39,5% dan
acid detergent fiber
(ADF)
35,10%. (Gracia
et al
. 1996). Sedangkan
ikan
mempunyai
kemampuan
yang
terbatas dalam memanfaatkan serat. Hal
ini berkaitan dengan ketersediaan enzim
selulotik yang terbatas dalam saluran
pencernaan ikan, bahkan pada level
tertentu dapat menghambat pertumbuhan
ikan. Beberapa peneliti telah melaporkan
bahwa ikan tidak memiliki enzim selulosa
dan
kemungkinan
adanya
populasi
mikroba selulotik di saluran pencernaan
ikan juga masih menjadi kontrofersi di
kalangan peneliti (Stickney & Shumway
1974; Prejs & Blaszczyk 1977; Linsday
dan Harris 1980; Lessel dan Lesel 1986;
Luczkovich & Stellway 1993; Saha & Ray
1998).
Jalilvand
et al.
(2008) melaporkan
enzim fibrilotik eksogen sangat efektif
untuk menurunkan kadar serat bahan
baku pakan seperti jerami padi, dan silase
jagung. Penggunaan enzim eksogen
diharapkan dapat menghidrolisis tepung
daun lamtoro gung sehingga dapat
ditingkatkan
kualitas
nutrisi
dan
kecernaannya.
Penggunaan
enzim
eksogen ini terkendala dengan harga
enzim komersil yang mahal di pasaran,
sehingga sangatlah penting dicari sumber
enzim yang murah dan efektif untuk
meningkatkan kualitas nutrisi dari tepung
daun lamtoro gung.
Cairan rumen domba merupakan
salah satu sumber bahan suplemen
alternatif
yang
murah
dan
dapat
dimanfaatkan dengan mudah sebagai
sumber
enzim-enzim
hidrolase
(Moharrery dan Das, 2001) diantaranya
enzim yang bersifat selulitik (Kung, 2006).
Hal ini berkaitan dengan kemampuan
domba untuk mencerna hijauan dengan
kandungan serat yang tinggi. Isi rumen
domba sebagai sumber ekstrak enzim
kasar
akan
dimanfaatkan
untuk
menghidrolisis (predigestiori) TDL yang
akan digunakan sebagai bahan campuran
pakan ikan nila. Produk yang diekstraksi
dari cairan rumen ini diharapkan dapat
secara langsung digunakan sehingga
jauh lebih efisien dibanding bila harus
menggunakan enzim dengan mendirikan
sebuah industri enzim, asam amino,
vitamin dan mineral, serta dapat menjadi
salah satu sumber bahan suplemen
alternatif
yang
murah
dan
dapat
dimanfaatkan
dengan
mudah
untuk
meningkatkan kualitas nutrisi dari tepung
daun lamtoro gung, sehingga kecernaan
meningkat dan pertumbuhan ikan nila
dapat lebih optimal.
MATERI DAN METODE
Eksperimen
Penelitian dilakukan selama ± 6
bulan dari Juni - Desember 2008, di
69
Laboratorium
Nutrisi
Ternak
Perah
Fakultas Peternakan IPB. Bahan yang
digunakan adalah tepung daun lamtoro
gung yang diperoleh dari daerah Bogor,
cairan rumen domba yang diperoleh dari
peternak domba tradisional di wilayah
Ciampea, Bogor (dari ternak yang sudah
dipotong yang isi rumennya dikeluarkan
dan
kemudian
diperas
untuk
mendapatkan cairan rumennya).
Persiapan Enzim Cairan Rumen
Cairan rumen sapi yang diambil
dari RPH disentrifus dengan kecepatan
12.000 rpm selama 10 menit pada suhu
4°C.
Kemudian
supernatan
yang
terbentuk direaksikan dengan ammonium
sulfat dengan menggunakan magnetic
stirer dan didiamkan selama semalam
pada suhu 4
oC. Cairan rumen kemudian
disentrifus kembali dengan kecepatan
12.000 rpm selama 15 menit pada suhu
4°C. Selanjutnya endapan diambil
sebagai enzim kasar (Pantaya, 2003).
Enzim kasar tersebut langsung digunakan
untuk hidrolisis TDL.
Rancangan Penelitian
Percobaan
ini
didesain
menggunakan rancangan acak lengkap
(RAL) dengan 6 perlakuan dan 3 ulangan
dan
setiap
ulangan
dilakukan
pengulangan dua kali (duplo). Dilakukan
inkubasi 24 jam pada 6 level jumlah
ekstrak enzim kasar yang ditambahkan
yaitu, 0 (K), 20 (A), 40 (B); 60 (C); 80 (D);
100 (E) mL/kg TDL. Parameter yang
diamati adalah komponen serat kasar, NDF
dan ADF, serta kadar asam fitat dari TDL
sebelum dan sesudah dihidrolisis dengan
ekstrak enzim rumen.
Data perubahan
kualitas nutrien dianalisis menggunakan
ANOVA dengan software SAS versi 6.12
(1997). Uji lanjut Duncan dilakukan pada
data yang menunjukkan perbedaan nyata.
Analisa kadar serat kasar dan kadar asam
fitat dilakukan dengan metode AOAC
(1990), sedangkan analisa komponen serat
kasar NDF dan ADF dilakukan dengan
metode Van Soest
et al
.(1991).
HASIL DAN PEMBAHASAN
Hasil
Serat Kasar
Kandungan serat kasar dalam
TDL setelah dihidrolisis oleh enzim kasar
cairan rumen domba yang mengandung
enzim selulase. Hasil pengukuran kadar
serat TDL yang dihasilkan pada akhir
periode inkubasi 24 jam disajikan pada
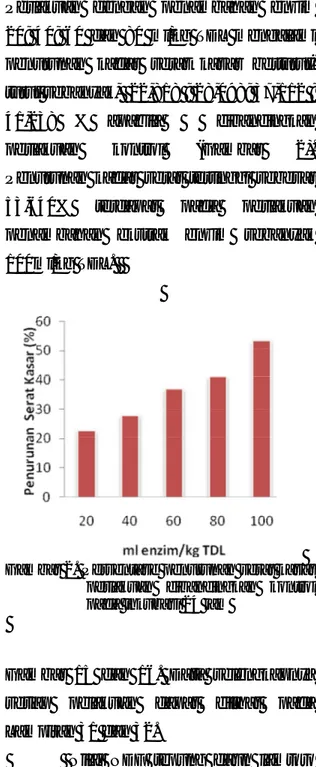
Gambar 1.
Kandungan serat kasar nyata
dipengaruhi oleh penambahan enzim
rumen, dimana kandungan serat kasar
mengalami
penurunan
dengan
peningkatan penambahan jumlah enzim(P
< 0,05). Nilai kadar serat kasar tertinggi
yaitu 16,769% terdapat pada perlakuan
70
tanpa penambahan enzim yang berbeda
nyata dengan semua perlakuan dengan
penambahan enzim. Nilai kadar serat
kasar yang terendah yaitu 7,774% dicapai
oleh
perlakuan
pemberian
enzim
100ml/kg. Nilai 7,774% ini berbeda nyata
dengan
perlakuan
lainnya
yaitu
pemberian enzim 20, 40, 60, dan 80 ml/kg
TDL yang menghasilkan nilai kadar serat
kasar berturut-turut adalah 12,942%;
12,057%; 10,546% dan 9,854% .
Perlakuan dengan penambahan enzim
20; 40; 60 dan 80 ml/kg TDL mengalami
penurunan kadar serat kasar
berturut-turut sebanyak, 22,818 ; 28,098; 37,112 ;
41,238 % apabila dibandingkan
perlakuan
kontrol
(Gambar
2).
Penurunan kadar serat tertinggi sebesar
53,640%
terdapat
pada
perlakuan
penambahan ekstrak enzim sebanyak
100ml/kg TDL.
Gambar 1. Kandungan serat kasar setiap
perlakuan
dengan
masa
inkubasi 24 jam
Gambar 2. Persentase penurunan serat kasar
perlakuan dibandingkan kontrol
pada inkubasi 24 jam
Ket : huruf yang sama pada diagram batang
Menunjukkan nilai tidak berbeda nyata
(P >0,05)
NDF dan ADF
Hasil pengukuran kadar NDF dan
ADF tepung daun lamtoro gung yang
telah mendapat penambahan eksrak
enzim kasar cairan rumen disajikan pada
Gambar 3 dan 4.
Hasil pengukuran kadar NDF dan
ADF tepung daun lamtoro gung yang
telah mendapat penambahan eksrak
enzim kasar cairan rumen disajikan
Gambar 15 dan 16. Data selengkapnya
setiap pelakuan dapat dilihat pada
Lampiran 31 dan 32.
Nilai NDF tepung daun lamtoro
gung nyata (P < 0,05) dipengaruhi oleh
penambahan
ekstrak
enzim
rumen
domba. Terdapat perbedaan yang nyata
antara
nilai
NDF
perlakuan
tanpa
penambahan enzim dengan nilai NDF
perlakuan yang mendapat penambahan
enzim 40ml/kg TDL. Nilai NDF tertinggi
d71
yaitu 46,32% dicapai pada perlakuan
tanpa penambahan enzim yang tidak
berbeda
nyata
dengan
nilai
NDF
perlakuan yang mendapat penambahan
enzim 20 dan 40ml/kg TDL. Sedangkan
nilai NDF terendah yaitu 42,54 % dicapai
pada perlakuan 100 ml/kg TDL yang tidak
berbeda nyata dengan nilai NDF dengan
penambahan
enzim 80
ml/kg
TDL
(44,31%) dan 60 ml/kg TDL (44,42%).
Semakin
banyak
enzim
yang
ditambahkan,
nilai
NDF
mengalami
penurunan.
Perlakuan
dengan
penambahan enzim sebanyak 20; 40; 60;
80 dan 100 ml/kg dibandingkan dengan
kontrol, menghasilkan penurunan nilai
NDF berturut-turut sebanyak 3,66; 6,29;
10,14; 10,36 dan 13,92%.
Penambahan
ekstrak
enzim
rumen
domba
nyata
(P
<
0,05)
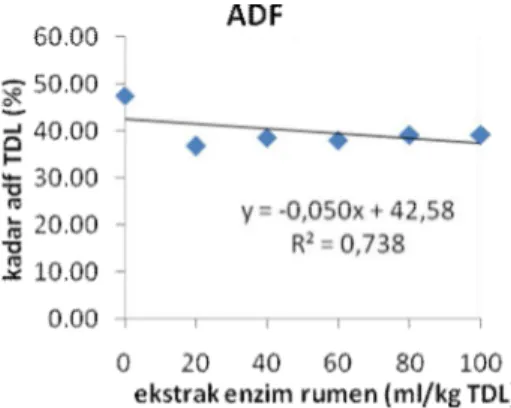
mempengaruhi nilai ADF tepung daun
lamtoro. Nilai ADF perlakuan tanpa
penambahan enzim lebih tinggi dari nilai
ADF seluruh perlakuan yang mendapat
penambahan
enzim.
Perlakuan
penambahan enzim pada taraf taraf 20,
40, 60, 80 dan 100 ml/kg tidak
menghasilkan nilai ADF yang berbeda
tetapi lebih rendah dan nyata berbeda
dengan perlakuan tanpa penambahan
enzim. Peningkatan jumlah enzim yang
ditambahkan
cenderung
akan
menurunkan nilai ADF. Secara
berturut-turut penambahan enzim sebanyak 20;
40; 60; 80 dan 100 ml/kg akan
menurunkan nilai ADF sebanyak 18,70 ;
14,63; 15,83 ; 13,43 dan 13,46%.
Gambar 3. Kadar NDF setiap perlakuan
dengan masa inkubasi 24 jam
Gambar 4. Kadar ADF setiap perlakuan
dengan masa inkubasi 24 jam
Ket : huruf yang sama pada diagram batang
menunjukkan nilai tidak berbeda nyata
(P >0,05)
Kadar Asam Fitat
Hasil pengukuran kadar asam fitat
tepung daun lamtoro gung yang telah
72
mendapat penambahan eksrak enzim
kasar cairan rumen disajikan pada
Gambar 5.
Kandungan
asam
fitat
nyata
(P<0,05) dipengaruhi oleh penambahan
ekstrak enzim rumen domba. Kadar
asam fitat pada perlakuan penambahan
ekstrak enzim rumen sebanyak 20, 40, 60
dan 80 ml/kg berbeda nyata dengan
perlakuan kontrol dan perlakuan dengan
penambahan
eksrak
enzim
rumen
sebanyak 100ml/kg TDL. Nilai asam fitat
tertinggi
yaitu 7,84%
dicapai
pada
perlakuan tanpa penambahan enzim dan
perlakuan terendah yaitu 2,50% dicapai
pada pelakuan yang menggunakan enzim
terbanyak (100ml/kg TDL).
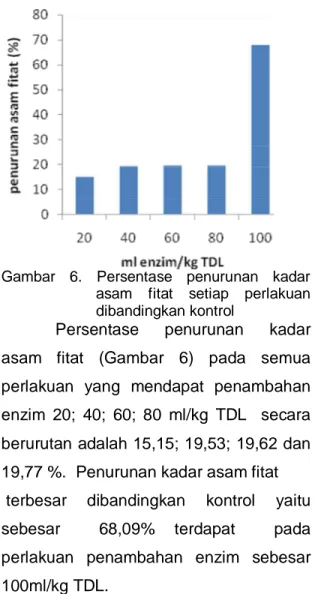
Gambar 5. Nilai rata-rata kadar asam fitat
setiap perlakuan uji in vitro
Gambar 6. Persentase penurunan kadar
asam fitat setiap perlakuan
dibandingkan kontrol
Persentase
penurunan
kadar
asam fitat (Gambar 6) pada semua
perlakuan yang mendapat penambahan
enzim 20; 40; 60; 80 ml/kg TDL secara
berurutan adalah 15,15; 19,53; 19,62 dan
19,77 %. Penurunan kadar asam fitat
terbesar
dibandingkan
kontrol
yaitu
sebesar 68,09% terdapat pada
perlakuan penambahan enzim sebesar
100ml/kg TDL.
Uji Respon Parameter Kualitas Nutrien
Respon kadar serat dan kadar
asam fitat pada periode inkubasi 24 jam
membentuk pola persamaan garis linier.
Pola respon kadar serat (Gambar 7) dan
asam fitat (Gambar 8) membentuk pola
respon
linier
negatif
yang
menggambarkan semakin banyak ekstrak
enzim rumen yang ditambahkan maka
kadar serat dan kadar asam fitat akan
menurun.
73
Gambar 7. Kurva respon kadar serat
perlakuan uji in vitro
Gambar 8. Kurva respon kadar asam
fitat perlakuan uji in vitro
Respon kadar NDF dan kadar
ADF pada periode inkubasi 24 jam
membentuk pola persamaan garis linier
negatif yang mengambarkan semakin
banyak ekstrak enzim rumen yang
ditambahkan maka kadar serat akan
menurun.
Gambar 9. Kurva respon kadar NDF perlakuan uji in vitro
Gambar 10. Kurva respon kadar ADF perlakuan uji in vitro
Pembahasan
Hasil percobaan secara
in vitro
menunjukkan bahwa eksrak enzim
cairan
rumen
domba
efektif
menghidrolisis tepung daun lamtoro
gung untuk bahan formulasi pakan ikan
nila dan sangat dipengaruhi oleh jumlah
enzim yang ditambahkan
Pada penelitian ini, penambahan
ekstrak enzim cairan rumen akan
menurunkan
kadar
serat
TDL.
Penambahan 100 ml enzim/kg TDL
dapat
menurunkan
kadar
serat
mencapai 53,40%. Penurunan kadar
serat ini merupakan hasil dari kerja
enzim amilase dan selulase yang
disekresikan
oleh
mikroba
yang
terkandung pada cairan rumen. Dimana
enzim amilase akan menghidrolisis
-1,4 menjadi pati cair dan
maltose sedangkan enzim selulase
akan
menghidrolisis
selulosa
yang
memiliki rantai yang lebih pendek.
Enzim amilase akan menghidrolisis
74
ikatan -1,4 menjadi D-glukosa, maltosa
dan sejumlah kecil destrin. Proses
penghidrolisisan ini merupakan kerja
kelompok
endo
amilase
dan
eksoamilase. Endo amilase yaitu enzim
amilase
yang
bekerja
dengan
memecah ikatan pada bagian tengah
substrat dengan pH optimum 5-7 dan
suhu optimum 60 – 70
oC. Endo amilase
banyak ditemukan pada tanaman dan
mikroorganisme,
terutama
Bacillus
stearothermophilus,
B-subtilus,
Apergilus
niger
dan
A.oryzae.
Sedangkan kelompok ekso amilase
adalah menghidrolasis
unit-unit dari
ujung non reduksi substrat menjadi
maltose dan maltotriosa dengan pH 4,5
– 5,5 dan suhu 40- 60
oC. Ekso amilase
banyak ditemukan pada tanaman dan
mikroorganisme,
terutama
Bacillus
stearothermophilus,
B-subtilus,
Apergilus niger dan A.oryzae. Jenis
mikroorganisme
ini
sangat
banyak
didapatkan di rumen, sehingga ketika
cairan
rumen
dieksraksi
untuk
mendapatkan enzim kasar, jenis enzim
amilase
selulase
akan
terdeteksi
aktifitasnya.
Enzim
selulase
merupakan
kompleks enzim (multi komponen) yang
terdiri dari beberapa enzim yang bekerja
bertahap
atau
bersama-sama
menguraikan
selulosa
menjadi
D-glukosa (Kim et al. 1994). Ada empat
kelompok enzim utama yang menyusun
selulosa berdasarkan substrat
masing-masing enzim, yaitu: Pertama; Endo
-
-4
D-glukanohidrolase, EC 3.2.1.4),
Cx-selulase, menghidrolisis ikatan glikolisik
-4) secara acak. Enzim ini tidak
menyerang
selobiosa
tapi
menghidrolisis selodekstrin. Enzim ini
juga aktif menyerang selulosa yang
telah
disubstitusi
misalnya
-4)
D-glukan selobiohidrolase (EC 3.2.1.91),
Cl
yang
menyerang
ujung
rantai
selulosa
non
pereduksi
dan
menghasilkan selobiosa. Enzim ini
dapat menyerang selodekstrin tapi tidak
menyerang
selulosa
yang
telah
disubstitusi
serta
tidak
dapat
-4)
D-glukan glukohidrolase (EC 3.2.1.74),
menyerang ujung rantai selulosa non
pereduksi dan menghasilkan glukosa.
Enzim ini menyerang selooligosakarida
dan CMC. Sedangkan yang ke empat
-
-4)
D-glukosida
glukohidrolase
(EC
3.2.1.21), menghidrolisis selobiosa dan
rantai pendek selooligosakarida dan
glukosa.
Cone
(1990)
melakukan
observasi dengan scanning elektron
mikroskop yang memperlihatkan hasil
bahwa telah terjadi degradasi granula
bahan pati dengan penambahan cairan
rumen yang telah bebas dari sel-sel
mikroba rumen.
Analisa kualitas nutrien TDL
dengan
periode
inkubasi
24
jam
memperlihatkan
penurunan
nilai
kandungan serat kasar dan serta
75
perubahan komponen serat yaitu ADF
dan NDF. Nilai
Acid Detergent Fiber
(ADF) dan
Neutral Detergent Fiber
(NDF) adalah nilai yang dihasilkan untuk
mengambarkan komponen dari serat
kasar dimana kedua metode ini hanya
dapat menentukan kadar total serat
yang tak larut dalam larutan deterjen.
ADF hanya dapat untuk menentukan
kadar
total
selulosa
dan
lignin,
sedangkan
dengan
NDF
dapat
menententukan
kadar
total
dari
selulosa,
hemiselulosa
dan
lignin.
Selisih jumlah serat dari analisis NDF
dan ADF dianggap jumlah kandungan
hemiselulosa,
meski
sebenarnya
terdapat
juga
komponen-komponen
lainnya selain selulosa, hemiselulosa
dan lignin. Pada penelitian ini nilai
NDF dan ADF perlakuan kontrol tanpa
penambahan enzim memperlihatkan
nilai kandungan NDF yang lebih tinggi
dibandingkan dengan perlakuan 20, 40,
60
dan
100
ml
enzim/kg
TDL,
penurunan nilai NDF dan ADF ini dapat
menggambarkan meningkatnya bagian
bahan pakan yang dapat dicerna.
Menurunnya kadar serat serta
komponen
NDF,
ADF
dengan
bertambahnya jumlah ekstrak enzim
cairan rumen yang ditambahkan, terjadi
karena peluang substrat untuk bertemu
dengan
katalisator
biologis
dalam
proses hidrolisis protein, karbohidrat
dan lemak semakin besar. Abu
et al
(2005) melaporkan bahwa peningkatan
konsentrasi enzim secara umum akan
memberikan pengaruh yang lebih besar
pada waktu hidrolisis dibandingkan
dengan peningkatan temperatur. Lama
proses
hidrolisis
berlangsung
menyebabkan
substrat
yang
terdegradasi
semakin
banyak
dan
produk yang dihasilkan akan semakin
meningkat. Vijaya
et al. (2002)
melaporkan
adanya
indikasi
peningkatan derajat hidrolisis dengan
peningkatan waktu inkubasi.
Dilaporkan oleh Pantaya (2005)
bahwa perlakuan tanpa enzim rumen
mengandung polisakarida lebih tinggi
dibandingkan
perlakuan
dengan
penambahan enzim rumen, dimana
perlakuan dengan penambahan enzim
rumen 620 dan 1240 U/kg pada
wheat
pollard
menurunkan kadar polisakarida
sebesar 4 dan 3,9%. Hidrolisis enzim
1240
U/kg
terhadap
komponen
polisakarida
wheat pollard juga akan
meningkatkan kandungan oligosakarida
dan
monosakarida
sebesar
5,5%
dibandingkan pada perlakuan tanpa
penambahan enzim (Pantaya, 2005).
Kemampuan
bakteri
rumen
untuk
meningkatkan kualitas bahan baku
pakan telah
dibuktikan pula oleh
Purnomohadi
(2006),
dimana
fermentasi jerami padi selama 7 hari
dengan
bakteri
selulitik
rumen
menghasilkan penurunan bahan kering
10,6%, kadar serat 15,98% .
Penambahan
enzim
cairan
rumen ini akan merombak komponen
bahan yang sulit dicerna menjadi mudah
76
dicerna,
dimana
selulosa
dipecah
menjadi komponen glukosa yang dapat
dimanfaatkan sebagai sumber energi
bagi hewan (Twoney
et al.
2003).
Didukung pula oleh Alemawor (2009)
yang mendapatkan peningkatan kualitas
nutrien
yang
lebih
baik
pada
penggunaan multi enzim pada bahan
baku pakan dengan nilai total gula
meningkat, serat kasar, NDF, ADF,
selulase dan lignin yang menurun.
Pada penelitian ini TDL tanpa
perlakuan
penambahan
enzim
mengandung
asam
fitat
sebanyak
7,839%. Sedangkan terjadi penurunan
kadar fitat sebanyak 68,088 % TDL
setelah
diinkubasi
dengan
ekstrak
cairan rumen domba sebanyak 100
ml/kg TDL. Satu mili ekstrak ekstrak
cairan rumen domba mengandung
enzim
fitase
2,7388
Unit/menit.ml
(Fitriliyani,
unpublisded). Satu unit
fitase didefenisikan sebagai kuantitas
enzim itu membebaskan 1 mikromol dari
fosfor per menit dari 0,0015mol/L
sodium phytate pada pH 5,5 dan 37
oC
(Simon
et al. 1990). Dalam enzim
rumen didapatkan aktifitas enzim fitase.
Fitase dapat menghidrolisis asam fitat
secara
bertahap
menjadi
senyawa
turunannya, yang dapat larut dan
terserap dalam sistem pencernaan.
Fitase
(mio-inositol
heksakisfosfat
fosfohidrolase, E.C. 3.1.3.8) merupakan
suatu fosfomonoesterase yang mampu
menghidrolisis
asam
fitat
menjadi
ortofosfat anorganik dan ester-ester
fosfat dari mio-inositol yang lebih
rendah. Cole (2001) mengemukakan
bahwa terdapat 2 jenis enzim fitase
yaitu; 3-fitase yang diperoleh dari fungi
dan
6-fitase
yang
diperoleh
dari
tumbuhan. Perbedaan khas dari kedua
jenis
ini
adalah
tempat
hidrolisis
pertama molekul fitat, 3-fitase pertama
memotong asam fitat pada posisi 3 dan
6-fitase pertama memotong asam fitat
pada posisi 6.
Kadar asam fitat memperlihatkan
pola respon linier yang kadarnya
semakin
menurun
dengan
meningkatnya jumlah ekstrak enzim
cairan
rumen
yang
ditambahkan.
Penurunan kadar asam fitat merupakan
hasil kerja dari enzim fitase yang
terkandung dalam ekstrak enzim cairan
rumen domba. Asam fitat merupakan
zat anti nutrisi yang secara alamiah
terdapat pada tanaman leguminosa dan
kacang-kacangan.
Mio-inositol
heksakisfosfat
(C
6H
18O
24P
6)
adalah
rumus kimia dari asam fitat dengan
struktur cincin yang mirip dengan
glukosa, yang berikatan dengan fosfor
unruk membentuk struktur asam fitat.
Selain fosfor unsur-unsur lain juga
ditemukan terikat dalam asam fitat
(Ravindra,
2000)
seperti
mineral
bervalensi dua (Ca, Zn, Fe dan Mg)
yang akan membentuk fitat mineral
yang tidak larut (Cole, 2001). Asam fitat
tidak larut dalam pH netral dan
menurunkan aktifitas enzim protease
dengan protein yang mengikat asam
77
fitat, sehingga akan menurunkan pula
bioavailability dari protein di dalam
pakan (Ravindra, 2000).
Kandungan enzim fitase yang
menghidrolisis
asam
fitat
akan
melepaskan bahan-bahan mineral dari
gugusnya seperti P, Ca, Zn, Mg dan Fe
. Hal ini sejalan pula dengan analisa
asam fitat yang terkandung pada
perlakuan inkubasi TDL dengan 80ml/kg
TDL yang mengalami penurunan 19,776
dibandingkan
perlakuan
kontrol.
Armani dan Refilda (2005) melaporkan
bahwa penambahan enzim fitase
pada gandum, bekatul dan kedelai
dapat meningkatkan ketersediaan
mineral Ca, Mg, Fe dan Zn. Mineral Ca,
Mg, Fe dan Zn yang dibebaskan dari
gandum, berturut-tuurut 70 %, 7,1 %,
17,5 % dan 89,6 %; pada bekatul
berturut-turut mencapai 60 %, 17,5 %,
7,7 % dan 86,8 % dan pada Kedelai
berturut-turut mencapai 77 %, 7,7 %,
12,1 % dan 88,9 %.
Ikan mempunyai keterbatasan
dalam menyerap fosfor dari air karena
konsentrasi fosfor dalam air sangat
kecil, sehingga kebutuhan fosfor ikan
sebagian besar dipenuhi dari pakan
(NRC, 1993). Mineral fosfor penting
sebagai komponen dari fosfolipid,
asam-asam nukleat, senyawa berenergi tinggi
(ATP). Fosfor berperanan penting
dalam metabolism karbohidrat, lemak
dan asam amino, sedangkan dalam otot
dan jaringan syaraf berperan dalam
menjaga tekanan osmotic cairan tubuh
(Lall. 2002). Keseimbangan fosfor
dalam tubuh dijaga dengan jalan
pertukaran antara senyawa fosfor dalam
tulang dan fosfor yang ada dalam
makanan (Djodjosubagio dan Piliang,
1990).
KESIMPULAN DAN SARAN
Kesimpulan
Peningkatan
penambahan
ekstrak
enzim cairan rumen domba untuk
menghidrolisis TDL efektif menurunkan
kandungan serat dan kandungan asam
fitat.
Saran
Perlunya kajian peluang pemanfaatan
eksrak enzim rumen dan isi rumen
untuk feed additive pakan ikan.
DAFTAR PUSTAKA
Abu EA, Ado SA and James DB. 2005.
Raw starch degrading amylase
production by mixed culture of
Aspergillus
niger
and
Saccharomyces cerevisae
grown
on
Sorghum pomace,
Afr. J.
Biotechnol. 4(8):785-790.
Agbede JO and Aletor VA. 2004.
Chemical
characterization
and
protein quality evaluation of leaf
protein
concentrates
from
Gliricidia sepium
and
Leucaena
leucocephala.
International
Journal of Food Science and
Technology, 39: 253-261.
Alemawor
F,
Victoria,
Dzogbefia,
Emmanuel
OK,
Oddoye
and
James HO. 2009. Enzyme
cocktail for enhancing poultry
utilisation of cocoa pod husk.
Scientific Research and Essays,
4(6):555-559.
78
AOAC. 1990. Official Methods of
Analysis. Association of Official
Analitycal Chemist. AOAC.
Washington DC. USA.
Armaini dan Refilda. 2005. Pengaruh
fitase
terhadap
peningkatan
ketersediaan mineral dalam bahan
pangan yang berasal dari
biji-bijian. Working Paper. Fakultas
Matematika
dan
Ilmu
Pengetahuan Alam. (Unpublished)
URL:
http://repository.unand.ac.id/id/epri
nt/1618
Cole SJ. 2001. Phytase. www.phytase
.net. 6 Maret 2010.
Djojosoebagio S dan Piliang GW.
1996. Fisiologi nutrisi. Jakarta UI
Press.
Gracia GW, Ferguson TU, Neckles FA,
and Archibald KAE. 1996. The
nutritive
value
and
forage
productivity
of
Leucaena
leucocephala.
Anim
Feed
Sci
Technol. 60:29-41.
Jalilvand G, Odongo NE, López S,
Naserian A, Valizadeh R, Eftekhar
Shahrodi, F, Kebreab E and
France J. 2008. Effects of
different levels of an enzyme
mixture on in vitro gas production
parameters of contrasting forages.
Anim. Feed Sci. Tech.
146:289-301.
Kung LJr, Treacher RJ, Nauman GA,
Smagala AM, Endres KM and
Cohen MA, 2000. The effect of
treating forages with fibrolytic
enzymes on its nutritive value and
lactation performance of dairy
cows. J. Dairy Sci. 83:115-122.
Lall SP. 2002. Mineral nutrition, p
260-308. In JE Halver and RW Hardy
(eds), Fish nutrition, 3
rded,
Academic Press, San Diego, USA.
Lessel R, Frogeot C, Lesel M. 1986.
Cellulose digestibility in grass
carp
Ctenopharyngodon idella
and goldfish Carassius auratus.
Aquaculture,
54;11-17.
Lindsay GJH and Harris JE. 1980.
Carboxymethylcellulase activity in
the digestive tracts of fish.
Journal
of Fish Biology. l6:219-233.
Luczkovich JJ and Stellwag EJ. 1993.
Isolation of cellulolytic microbes
from the intestinal tract of Lagodon
rhomboides: size-related changes
in
diet and microbial abundance.
Marine Biology. 16:381 -388.
Moharrery A and Das Tirta K. 2002.
Correlation
between
microbial
enzyme activities in the rumen fluid
of sheep under different treatments.
Reprod. Nutr. Dev,41:513-529.
NAS. 1994. Leucaena: Promising forage
and tree crop for the tropics. Second
Edition.
National
Academy
of
Sciences. Washington.
Pantaya Dadik, Nahrowi, Lily Amalia
Sofyan. 2005. Penambahan
enzim cairan rumen pada pakan
berbasis
wheat pollard
dengan
proses
pengolahan
steam
pelleting pada performans
Prejs A and Blaszczyk M. 2006.
Relationships between food and
cellulase activity in freshwater
fishes.
Journal of Fish Biology.
Vol 11;5; 447–452.
Purnomohadi M. 2006. Peranan bakteri
selulotik
cairan
rumen
pada
fermentasi jerami padi terhadap
mutu pakan.
Jurnal Protein, Vol
13, No 2.
Ravindran V, Cabahung S, Ravindran
G, Sell PH and Bryden WL. 2000.
Respose of broiler chickens to
microbial phytase supplementation
as influenced by dietary phytic
acid
and
non-phytate
phosphorous level. II. Effects on
apparent metaboliazable energy,
nutrient digestibility and nutrient
retention.
Br. Poult. Sci, 41:
193-200.
Saha A and Ray AK. 1998. Cellulase
activity
in
rohu
fingerlings.
Aquaculture
Internationale,
79
Scott JR, Newton SH and Katayama
RW. 1982. Evaluation of sunflower
meal
as
a
soybean
meal
replacement in rainbow trout diets.
Proceeding of Thirty-Sixth Annual
Conference.
South-Eastern
Association of Fish and Wildlife
Agencies, Jacksonville. Florida.
Stickney RR and Shumway SE. (1974)
Occurrence of cellulase activity in
the stomachs of fish. Journal of Fish
Biology, 6:779-790.
Sutardi T. 1981. Sapi perah dan
pemberian
makanannya.
Dep.
Ilmu
Makanan
Ternak.
Fak.
Petcrnakan. Inst. Pcrtanian Bogor.
Bogor
Twoney LN, Muske JR, Kowe JB, Choct
M, Brown W, Mc Connell MF and
Pethick DW. 2003. The effect of
increasing level of soluble non
starch polysaccharide on inclusion
of feed enzyme in dog diet on
fecal quality and digestibility.
Animal
Feed
Science
and
Technology ,108: 71-82.
Van Soest PJ, Robertson JB, Lewis BA.
1991. Methods for dietary fiber,
neutral detergent fiber, and
non-starch polysaccharides in
relation to animal nutrition.
J.
Dairy Sci. 74: 3583-3597.
B.
Vijaya GV, Gireesh T and Gajanan
SB. 2002. Effect of enzymatic
hydrolysis of proteins on growth
and milk production.
Journal of
the
Science
of
Food
and