EKSTRAK ENZIM CAIRAN RUMEN DOMBA (Ovis aries)
UNTUK BAHAN PAKAN IKAN NILA (Oreochromis niloticus)
INDIRA FITRILIYANI
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar
Doktor pada Program Studi Ilmu Perairan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa disertasi “Peningkatan Kualitas Nutrisi Tepung Daun Lamtoro Dengan Penambahan Ekstrak Enzim Cairan Rumen Domba (Ovis aries) untuk Bahan Pakan Ikan Nila (Oreochromis niloticus) adalah karya saya sendiri dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau kutipan dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka dibagian akhir disertasi ini.
Bogor, Oktober 2010
with Addition of Sheep Rumen Liquor Enzyme for Nila Tilapia (Oreochromis
niloticus) feed. Under Advisory by ENANG HARRIS, ING
MOKOGINTA, NAHROWI.
The aims of this experiment were; (1) to evaluate hydrolyze enzyme activity in exctract rumen liquor of sheep fed forage, (2) to improve the nutrient quality of leucaena leaf meal (LLM) by addition with sheep rumen liquor enzyme; (3) to evaluate the suitability of hydrolized LLM as alternative protein source for Nile tilapia. The first stage of experiment showed that the activity of enzyme hydrolyze in rumen fluid of sheep with forage feeding had the largest cellulase activity than amylase, protease, lipase and phytase activity. The second stage was designed in completely randomized design with 6 treatments and 3 replications with different level enzyme addition (0; 20; 40; 60; 80; and 100 ml/kg LLM) and incubated in 2 and 24 hours.. Results of the first experiment showed that nutrient quality of LLM with addition of sheep rumen liquor enzyme 100 ml/kg LLM, within incubated 24 hours have the best result than that of incubated 2 hours.The enzymes significantly (P<0.05) increased soluble glucose (2127,45%) soluble protein (3538,23%), decreased crude fiber (53,640%) and phytic acid (68,088%); but the enzyme did not affect crude lipid and crude protein content. The third stage experiments was conducted with two experimental diet. The first one contained hydrolyzed LLM at level 10% 15% 20% 25% and 30% ( Group I; Diet A, B, C, D and E respectively) and one diet acting as a control (Diet K, 0% LLM), the second experiments contained non-hydrolized LLM at the same level as the first ezperiment ( Group II; Diet F, G, H, I, J). All diets were isoenergy. A seven weeks feeding trial was carried out on triplicate groups of eight fish (9,38 ± 0,41) in aquarium with a recirculating system for group I and II. Fish were fed twice daily at satiation. Results showed that improvement of the quality of hydrolyzed LLM increased daily growth rate, feed efficiency, protein and fat retention. Similarly, the value of Hepato somatic indeks, glycogen content and enzyme activity of protease, amylase and cellulase in fish feeding hydrolyzed LLM were higher than that of non- hydrolyzed LLM. HIS and liver glycogen storage in fish with hydrolyzed LLM in diet increased with the increasing use of hydrolyzed LLM. Histological liver and intestine of tilapia using hydrolyzed LLM in the diet did not show significant differences with those of fish with feed without LLM. It is was concluded that enzymes from rumen liquer capable of increasing the nutritive quality of LLM. Application of hydrolyzed LLM up to 15% in the diet of tilapia show better performance compared with non-hydrolyzed LLM
K
INDIRA FITRILIYANI. Peningkatan Kualitas Nutrisi Tepung Daun Lamtoro dengan Penambahan Ekstrak Enzim Cairan Rumen Domba ntuk Bahan Pakan Ikan Nila (Oreochromis sp). Dibimbing oleh ENANG
ARRIS, ING MOKOGINTA, NAHROWI. u
H
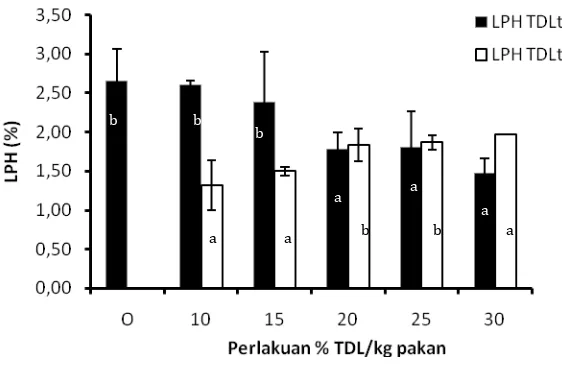
meningkatnya simpanan glikogen dengan meningkatnya kesediaan glukosa/karbohidrat sederhana di dalam pakan. Aktifitas enzim selulase dan amilase menunjukkan kecenderungan terjadinya peningkatan dengan meningkatnya penggunaan TDL terhidrolisis atau TDL tanpa hidrolisis dalam pakan akan tetapi aktifitas enzim selulase ikan perlakuan TDL terhidrolisis lebih rendah dari perlakuan TDL tanpa hidrolisis. Perlakuan kontrol tanpa penggunaan TDL memberikan nilai aktifitas enzim selulase terendah (0,0176 unit/ml/menit) dibandingkan seluruh perlakuan dengan TDL terhidrolisis maupun TDL tanpa hidrolisis. Nilai aktifitas enzim amilase pada saluran pencernaan ikan TDL terhidrolisis berada pada kisaran nilai 0,0758 –0,2148 (unit/ml/menit) sedangkan nilai aktifitas enzim amilase TDL tanpa hidrolisis berkisar 0,06202 – 0,292466 unit/ml/menit. Kisaran nilai aktifitas enzim amilase ini masih lebih rendah dari perlakuan kontrol tanpa penggunaan TDL terhidrolisis dalam pakan. Aktifitas enzim protease perlakuan yang menggunakan TDL terhidrolisis dan tidak terhidrolisis nyata (P<0,05) dipengaruhi oleh perbedaan persentase penggunaan TDL terhidrolisis dalam pakan dimana aktifitas enzim protease perlakuan dengan TDL terhidrolisis lebih tinggi dibandingkan TDL tanpa hidrolisis. Histologi hati dan usus ikan perlakuan dengan TDL terhidrolisis tidak memperlihatkan perbedaan yang signifikan dengan histologi hati dan usus ikan dengan pakan tanpa mengandung TDL. Dapat disimpulkan bahwa enzim selulase, amilase, protease, lipase dan fitase cairan rumen domba dapat meningkatkan kualitas nutrisi TDL. Penggunaan TDL terhidrolisis sampai 15 % dalam pakan ikan nila memperlihatkan kinerja pertumbuhan yang lebih baik dibandingkan penggunaan TDL tanpa hidrolisis pada taraf yang sama.
@ Hak Cipta milik IPB, tahun 2010
Hak Cipta dilindungi Undang-Undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebut sumber
a. Pengutipan hanya untuk kepentingan pendidikan, peneliti, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB 2. Dilarang mengumumkan dan memperbanyak sebagian atau
Dosen Penguji Ujian Tertutup 1. Dr. Ir. Dedi Djusadi, M.Sc
Wakil Dekan Pascasarjana, Institut Pertanian Bogor Staf Pengajar Departemen Budi Daya Perairan Fakultas Perikanan dan Ilmu Kelautan, IPB.
2. Dr. Ir. M. Agus Suprayudi, M.Si
Staf Pengajar Departemen Budi Daya Perairan Fakultas Perikanan dan Ilmu Kelautan, IPB.
Dosen Penguji Ujian Terbuka
1. Dr. Ir. Zafril Imran Azwar, M.Sc
Peneliti di Balai Riset Perikanan Budidaya Air Tawar, Sempur, Bogor.
2. Dr. Nur Bambang Priyo Utomo, M.Si
EKSTRAK ENZIM CAIRAN RUMEN DOMBA (
Ovis aries
)
UNTUK BAHAN PAKAN IKAN NILA (
Oreochromis niloticus
)
INDIRA FITRILIYANI
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar
Doktor pada Program Studi Akuakultur
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Lamtoro dengan Penambahan Ekstrak Enzim Cairan Rumen Domba (Ovis aries) untuk Bahan Pakan Ikan Nila (Oreochromis sp)
Nama : Indira Fitriliyani
NRP : C161060061
Program Studi : Ilmu Akuakultur
Disetujui Komisi Pembimbing
Prof. Dr. Ir. Enang Harris, M.S Ketua
Prof. Dr.Ir. Ing Mokoginta, M.Sc Prof. Dr.Ir. Nahrowi, M.Sc Anggota Anggota
Diketahui
Ketua Departemen Dekan Sekolah Pascasarjana Ilmu Akuakultur
Prof.Dr.Ir.Enang Harris, M.S Prof.Dr.Ir. Khairil Anwar Notodiputro, M.S
kurnia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Nopember 2008 ini adalah dengan judul Peningkatan Kualitas Nutrisi Tepung Daun Lamtoro Dengan Ekstrak Enzim Cairan Rumen Domba (Ovis aries) Untuk Bahan Pakan Ikan Nila (Oreochromis sp).
Ungkapan terimakasih disampaikan kepada
1. Tim komisi pembimbing Bapak Prof. Dr. Ir. Enang Harris, M.S; Ibu Prof. Dr. Ir. Ing Mokoginta, MSc dan Bapak Prof. Dr. Ir. Nahrowi, MSc atas semua bimbingan, saran, koreksi, motivasi dan kebijaksanaan.
2. Departemen Pendidikan Nasional Direktorat Jendral Pendidikan Tinggi (DIKTI) yang telah memberikan Beasiawa Program Pascasarjana, memberikan kesempatan untuk mengikuti Sandwich Programme di Fisheries Faculty Kagoshima University dan bantuan Hibah Program Doktor 2009.
3. Rektor, seluruh staf akademis di Fakultas Perikanan Jurusan Budidaya Perairan Universtitas Lambung Mangkurat atas izin tugas belajar.
4. Kedua orang tua tercinta ayahanda H. Anwar Fauzie (alm) dan ibunda Hj Marliah Chairul dan kepada seluruh keluarga besar, teman-teman di AIR atas bantuan yang diberikan.
5. Suamiku H.Muhammad Mabrur Lc MAg, anak-anak tersayang Muhammad Aliif Azhar; Muhammad Rozien dan Muhammad Nabil,. yang setia mendampingi penulis dalam cinta, kerja dan doa.
6. Prof Shunzuke Koshio dan Associate Prof Manabu Ishikawa, yang telah memberikan banyak bimbingan kepada penulis selama di Kagoshima. Semoga Allah SWT memberikan balasan amal dan kebaikan mereka yang tak terhingga. Disertasi ini penulis persembahkan kepada perkembangan ilmu pengetahuan semoga dapat menjadi ilmu yang bermanfaat.
Bogor, Oktober 2010
Penulis dilahirkan di Banjarmasin 5 Oktober 1975 dari Bapak H.Anwar Fauzie (Alm) dan Ibu Hj Marliah Chairul merupakan anak kelima dari lima bersaudara. Pendidikan sarjana ditempuh di Fakultas Perikanan Jurusan Budidaya Perikanan Universitas Lambung Mangkurat 1993-1997. Kesempatan untuk melanjutkan ke program Magister Sains pada Program Studi Ilmu Perairan pada Program Pascasarjana IPB dengan Beasiswa Pendidikan diperoleh dari beasiswa Departemen Pendidikan Tinggi (DIKTI) tahun 2003-2005.
Tahun 2006 penulis diterima sebagai mahasiswa program doktor di Program Studi Ilmu Perairan Sekolah Pascasarjana Institut Pertanian Bogor. Konsentrasi Ilmu Nutrisi Ikan dengan bantuan beasiswa Departemen Pendidikan Tinggi (DIKTI).
xi
DAFTAR ISI ………. xi
DAFTAR TABEL ……… xii
DAFTAR GAMBAR ……….. xiii
DAFTAR LAMPIRAN ……… xv
I. PENDAHULUAN ……… 1
1.1 Latar Belakang ………... 1
1.2 Perumusan Masalah ………... 3
1.3 Tujuan dan Manfaat Penelitian ………. 4
1.4 Perumusan Hipotesis...………..………... 4
II. TINJAUAN PUSTAKA……… 5
2.1 Kebutuhan Nutrien Ikan nila... 5
2.2 Tepung Daun Lamtoro (Leucaena leucocephala)... 8
2.3 Cairan Rumen sebagai Sumber Enzim ... Enzim Pencernaan dan Peranannya dalam Proses Pencernaan... 13 2.4 17 III. METODELOGI……… 21
3.1 Tahap Pertama: Pengujian Aktifitas Enzim-enzim Hidrolisis Cairan Rumen Domba………. 21 3.1.1 Isolasi dan Produksi Ekstrak Enzim Cairan Rumen Domba………... 21
3.1.2 Uji Aktifitas Ekstrak Enzim Cairan Rumen Domba………... 21
3.2 Tahap Kedua: Pengaruh Penambahan Enzim Cairan Rumen Domba (In Vitro) pada Kualitas Tepung Daun Lamtoro……… 22 3.3 Tahap Ketiga: Pengujian In vivo TDL pada pakan Ikan Nila……… 24
xii
Pembahasan……….. 32
4.2. Tahap Kedua: Pengaruh Penambahan Enzim Cairan Rumen Domba (In Vitro) pada Kualitas Tepung Daun Lamtoro 35 4.2.1. Glukosa Terlarut……….. 36
4.2.2. Protein Terlarut……… 39
4.2.3. Kandungan Nutrient Tepung Daun Lamtoro………... 41
Pembahasan……… 48
4.3. Tahap Ketiga: Pengujian In vivo TDL pada pakan Ikan Nila……… 60
4.3.1. Laju Pertumbuhan Harian………... 60
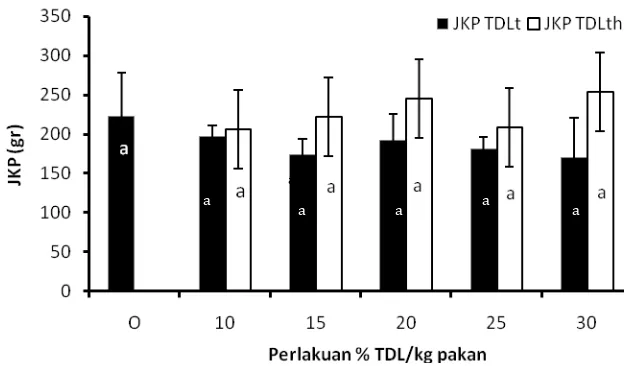
4.3.2. Jumlah Konsumsi Pakan………. 61
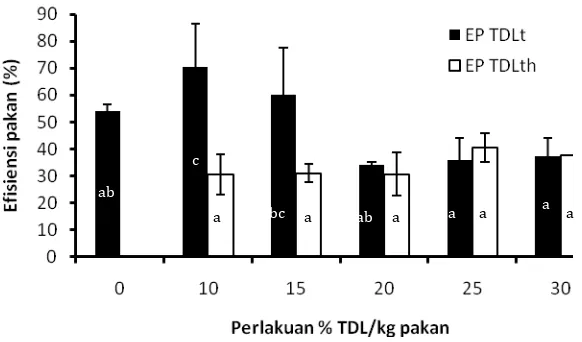
4.3.3 Efisiensi Pakan………. 62
4.3.4. Retensi Protein dan Lemak……….. 63
4.3.5. Hepatosomatik Indeks dan Kadar Glikogen Hati……… 65
4.3.6. Aktifitas Enzim Pencernaan……… 67
4.3.7. Kadar Glukosa Darah Ikan Uji Perlakuan TDL Terhidrolisis………. 70
4.3.8. Histologi Hati dan Usus………... 72
Pembahasan……… 77
V. KESIMPULAN DAN SARAN……… 95
5.1. Kesimpulan………. 96
5.2. Saran……… 96
DAFTAR PUSTAKA... 97
xiii
No Halaman
1 Kebutuhan protein ikan Nila Oreochromis sp dengan bobot tubuh yang berbeda... ...
6
2 Perbandingan komposisi asam amino dan makro, mikro mineral, antara tepung ikan (TI), bungkil kedelai (TBK) dan tepung daun lamtoro
(TDL)………. 9
3 Komposisi enzim cairan rumen sapi……….. 16
4 Komposisi enzim cairan rumen domba………. 16
5 Karakteristik enzim-enzim cairan rumen sapi asal RPH………. 16
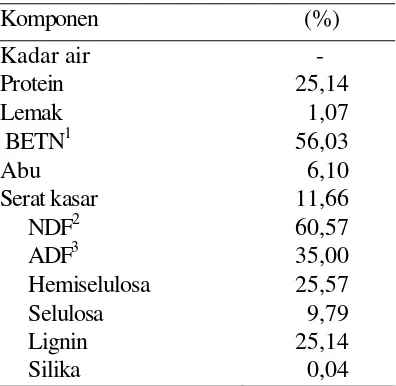
6 Komposisi proksimat dan komponen penyusun serat TDL……….... 23
7 Komposisi pakan perlakuan TDL terhidrolisis ………. Komposisi proksimat pakan perlakuan TDL terhidrolisis……… 25 8 . Komposisi proksimat pakan perlakuan TDL tanpa hidrolisis... 26 9 30 10 Kandungan aktifitas enzim pada setiap perlakuan………. 35
11 Komposisi asam amino tepung daun lamtoro sebelum sesudah inkubasi 24 jam dengan enzim rumen domba sebanyak 100 ml enzim/kg TDL………... 47
12 Komposisi asam amino essensial pakan percobaan ikan nila dengan taraf kandungan TDL terhidrolisis yang berbeda serta kebutuhan asam amino essensil ikan nila (% protein)……….. 77
xiv
No Halaman
1 Struktur molekul asam fitat……….. 11
2 Mekanisme asam fitat mengikat mineral………... 12
3 Mekanisme hidrolisis fosfat oleh enzim fitase (Applegate and Angel,
2004)……… 13
4 Bagian-bagian perut hewan ruminansia……… 13
5 Aktifitas enzim selulase, amilase, protease, lipase dan fitase ekstrak
cairan rumen domba………... 31
6 Nilai rata-rata kadar glukosa terlarut (%) periode inkubasi 2 jam
dan 24 jam………. 37
7 Persentase peningkatan glukosa terlarut perlakuan dibandingkan
kontrol pada inkubasi 2 jam dan 24 jam. ………... 38
8 Kurva respon kadar glukosa terlarut inkubasi 2 jam dengan jumlah
ekstrak enzim rumen berbeda……… 38
9 Kurva respon kadar glukosa terlarut inkubasi 24 jam dengan jumlah
ekstrak enzim rumen berbeda……… 38
10 Nilai rata-rata kadar protein terlarut setiap perlakuan dengan masa
inkubasi 2 jam dan 24 jam………. 39
11 Persentase peningkatan protein terlarut perlakuan dibandingkan
kontrol pada inkubasi 2 jam dan 24 jam……… 40
12 Kurva respon kadar protein terlarut inkubasi 2 jam……….. 41
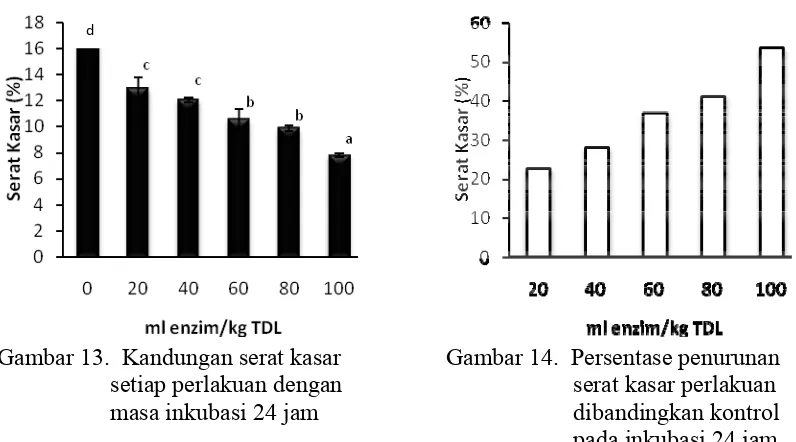
13 Kandungan serat kasar setiap perlakuan dengan masa inkubasi 24
jam………. 42
14 Persentase penurunan serat kasar perlakuan dibandingkan kontrol
pada inkubasi 24 jam………... 42
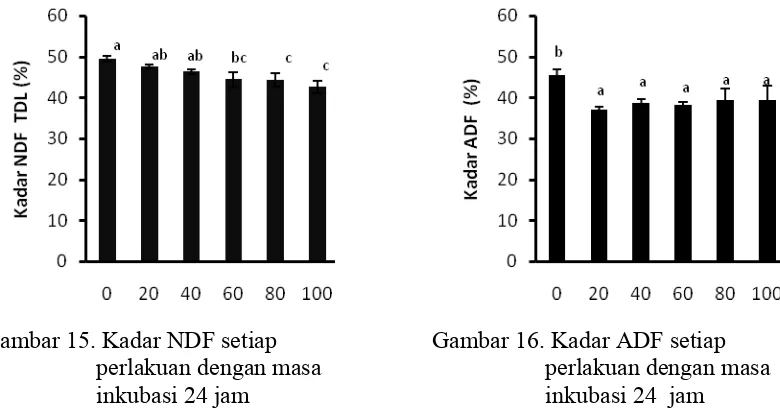
15 Kadar NDF setiap perlakuan dengan masa inkubasi 24 jam…………. 43
16 Kadar ADF setiap perlakuan dengan masa inkubasi 24 jam………… 43
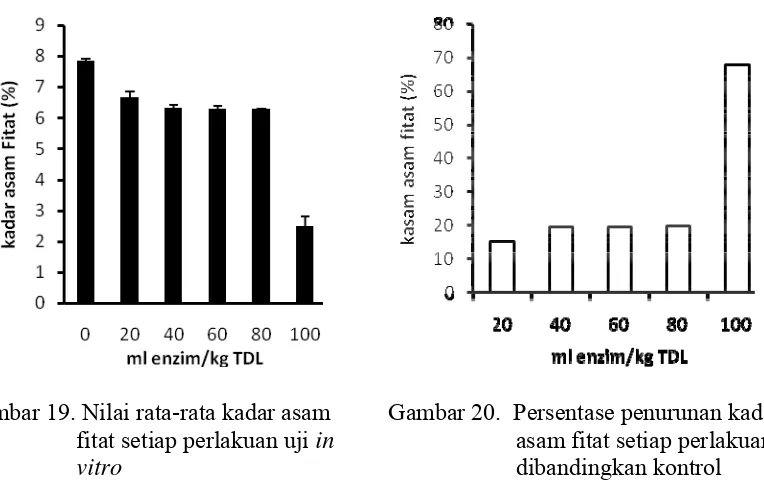
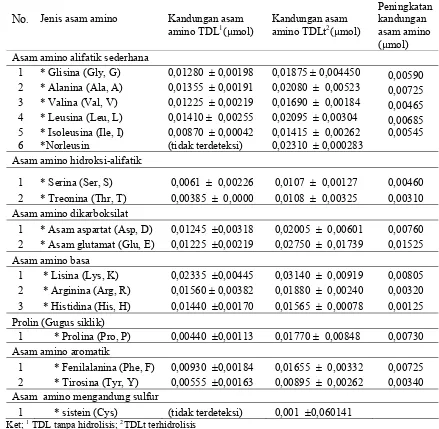
17 Nilai rata- rata kadar protein setiap perlakuan uji in vitro………... 44 18 Nilai rata-rata kadar lemak setiap perlakuan uji in vitro…………... 44 19 Nilai rata-rata kadar asam fitat setiap perlakuan uji in vitro………… 45 20 Persentase penurunan kadar asam fitat setiap perlakuan dibandingkan
kontrol……… 45
21 Kurva respon kadar asam fitat perlakuan uji in vitro……… 46
xv
25 Perbandingan peningkatan kadar glukosa terlarut dan protein terlarut perlakuan dengan penambahan 100 ml enzim/kg TDL dengan perlakuan tanpa enzim pada waktu inkubasi 2 dan 24 jam………
48
26
27
Mekanisme kerja enzim amilase memotong ikatan α-1,4………..
Tiga tipe reaksi yang dikatalisasi oleh enzim selulase………... 50 52
28 Hidrolisis protein oleh enzim eksopeptidase………. 54
29 Nilai Laju pertumbuhan harian perlakuan TDL terhidrolisis dan
TDL tanpa hidrolisis……….. 61
30 Nilai jumlah konsumsi pakan perlakuan TDL terhidrolisis dan TDL
tanpa hidrolisis………... 62
31 Nilai Efisiensi pakan perlakuan TDL terhidrolisis dan TDL tanpa
hidrolisis………. 63
32 Nilai retensi protein perlakuan TDL terhidrolisis dan TDL tanpa
hidrolisis………... 64
33 Nilai retensi lemak perlakuan TDL terhidrolisis dan TDL tanpa
hidrolisis………. 65
34 Nilai hepatosomatik indeks (HIS) perlakuan TDL terhidrolisis dan
TDL tanpa hidrolisis……….. 66
35 Aktifitas enzim selulase (IU/ml.menit) TDL terhidrolisis dan TDL
tanpa hidrolisis………... 68
36 Aktifitas enzim amilase (IU/ml.menit) TDL terhidrolisis dan TDL
tanpa hidrolisis………... 69
37 Aktifitas enzim protease (IU/ml.menit) TDL terhidrolisis dan TDL
tanpa hidrolisis………. 70
38 Pola kadar glukosa darah (mg/100mL) ikan uji setiap perlakuan pada
jam pengamatan 0, 4, 8 dan 24 jam post prandial………. 71
39 Histologi hati ikan nila perlakuan K (0% TDL dalam pakan) dan
perlakuan A (10% TDL dalam pakan)……….. 72
40 Histologi hati ikan nila perlakuan B (15% TDL dalam pakan) dan
perlakuan C (20% TDL dalam pakan)………... 73
41 Histologi hati ikan nila perlakuan D (25% TDL dalam pakan) dan
perlakuan E (30% TDL dalam pakan)………... 73
xvi
44 Histologi usus ikan nila perlakuan B (15% TDL dalam pakan) dan perlakuan C (20% TDL dalam pakan)-penekanan pada
struktur……….. 75
45 Histologi usus ikan nila perlakuan B (15% TDL dalam pakan) dan perlakuan C (20% TDL dalam pakan)-penekanan pada deteksi sel
goblet (GC) dan sel vill (VC)………. 75
46 Histologi usus ikan nila perlakuan D (25% TDL dalam pakan) dan perlakuan E (30% TDL dalam pakan)-penekanan pada struktur……... 76
47 Histologi usus ikan nila perlakuan D (25 % TDL dalam pakan) dan perlakuan E (30% TDL dalam pakan)-penekanan pada deteksi sel
xvii
Uji aktifitas enzim selulase/FP-ase (Metode Ghosse, 1987)……….
Uji aktifitas enzim amilase (Bergmeyer dan Grassi 1983)………
Uji aktifitas enzim protease (Bergmeyer dan Grassi 1983)………..
Uji aktifitas enzim lipase (Tietz dan Friedreck dalamBarlongan, 1990)……..
Uji aktifitas enzim Fitase (Greiner et al. 1997)………
Prosedur analisis kadar glukosa terlarut (Wedemeyer & Yasutake 1977)
Prosedur analisis kadar protein terlarut (Bradford, 1976)...
Prosedur analisis proksimat (kadar air, kadar abu, kadar protein, kadar lemak dan kadar serat kasar) mengikuti metode AOAC (1990)
Penentuan Kadar NDF dan ADF (Van Soest, 1991)………..
Prosedur analisis asam fitat (AOAC, 1990)………
Prosedur analisis HPLC (ICI Intrument Method, 1988)………
Prosedur analisis kadar glikogen (Wedemeyer dan Yasutake, 1977)...
Prosedur memperoleh ekstrak enzim saluran pencernaan untuk analisis aktifitas enzim pencernaan………..
Prosedur persiapan preparat histologis organ hati dan usus………….
Penghitungan aktivitas enzim seluase, amilase, protease, fitase dan
lipase enzim cairan rumen domba………
108
16 Penghitungan kadar glukosa terlarut daun lamtoro gung inkubasi 2 jam………
125
17 Penghitungan kadar glukosa terlarut daun lamtoro gung inkubasi 24 jam………
126
18 Analisis ragam kadar glukosa terlarut (%) tepung daun lamtoro gung pada periode inkubasi 2 jam setelah dihidrolisis dengan berbagai
jumlah ekstrak enzim cairan rumen domba………... 126
19 Uji Duncan kadar glukosa terlarut (%) tepung daun lamtoro gung pada periode inkubasi 2 jam setelah dihidrolisis dengan berbagai
jumlah ekstrak enzim cairan rumen domba……….. 127
20 Analisis ragam kadar glukosa terlarut (%) tepung daun lamtoro gung pada periode inkubasi 24 jam setelah dihidrolisis dengan berbagai
jumlah ekstrak enzim cairan rumen domba………. 127
21 Uji Duncan kadar glukosa terlarut (%) tepung daun lamtoro gung pada periode inkubasi 24 jam setelah dihidrolisis dengan berbagai
xviii
23 Penghitungan kadar protein terlarut tepung daun lamtoro gung inkubasi 24 jam ………
129
24 Analisis ragam kadar protein terlarut (%) tepung daun lamtoro gung pada periode inkubasi 2 jam setelah dihidrolisis dengan berbagai
jumlah ekstrak enzim cairan rumen domba……….. 130
25 Uji Duncan kadar protein terlarut (%) tepung daun lamtoro gung pada periode inkubasi 2 jam setelah dihidrolisis dengan berbagai jumlah ekstrak enzim cairan rumen domba………... 130
26 Analisis ragam kadar protein terlarut (%) tepung daun lamtoro gung pada periode inkubasi 24 jam setelah dihidrolisis dengan berbagai
jumlah ekstrak enzim cairan rumen domba……….. 130
27 Uji Duncan kadar protein terlarut (%) tepung daun lamtoro gung pada periode inkubasi 24 jam setelah dihidrolisis dengan berbagai
jumlah ekstrak enzim cairan rumen domba ………. 131
28 Penghitungan kadar serat tepung daun lamtoro gung inkubasi 24 jam……….
131
29 Analisis ragam kadar serat (%) tepung daun lamtoro gung pada periode inkubasi 24 jam setelah dihidrolisis dengan berbagai
jumlah ekstrak enzim cairan rumen domba……… 132
30 Uji Duncan kadar serat (%) tepung daun lamtoro gung pada periode inkubasi 24 jam setelah dihidrolisis dengan berbagai jumlah ekstrak enzim cairan rumen domba………..
132 31 Penghitungan kadar NDF tepung daun lamtoro gung inkubasi 24
jam……… 132
32 Penghitungan ADF tepung daun lamtoro gung inkubasi 24 jam…….. 133
33 Analisis ragam NDF (%) tepung daun lamtoro gung pada periode inkubasi 24 jam setelah dihidrolisis dengan berbagai jumlah
ekstrak enzim cairan rumen domba………. 133
34 Uji Duncan NDF (%) tepung daun lamtoro gung pada periode inkubasi 24 jam setelah dihidrolisis dengan berbagai jumlah ekstrak
enzim cairan rumen domba………... 134
35 Analisis ragam ADF (%) tepung daun lamtoro gung pada periode inkubasi 24 jam setelah dihidrolisis dengan berbagai jumlah
ekstrak enzim cairan rumen domba……….. 134
36 Uji Duncan ADF(%) tepung daun lamtoro gung pada periode inkubasi 24 jam setelah dihidrolisis dengan berbagai jumlah ekstrak enzim cairan rumen………...
xix
38 Penghitungan kadar lemak (% bobot kering) tepung daun lamtoro gung inkubasi 24 jam………
135
39 Analisis ragam kadar protein (%) tepung daun lamtoro gung pada periode inkubasi 24 jam setelah dihidrolisis dengan berbagai
jumlah ekstrak enzim cairan rumen ……….. 136
40 Analisis ragam kadar lemak (%) tepung daun lamtoro gung pada periode inkubasi 24 jam setelah dihidrolisis dengan berbagai
jumlah ekstrak enzim cairan rumen ……….. 136
41 Penghitungan kadar asam fitat (%) tepung daun lamtoro gung inkubasi 24 jam……….
137
42 Analisis ragam kadar asam fitat (%) tepung daun lamtoro gung pada periode inkubasi 24 jam setelah dihidrolisis dengan berbagai
jumlah ekstrak enzim cairan rumen domba……… 137
43 Uji Duncan kadar asam fitat (%) tepung daun lamtoro gung pada periode inkubasi 24 jam setelah dihidrolisis dengan berbagai jumlah
ekstrak enzim cairan rumen domba……….. 138
44 Perhitungan laju pertumbuhan harian ikan uji pada setiap perlakuan tahap ketiga dengan menggunakan TDL terhidroisis………...
138
45 Perhitungan laju pertumbuhan (%) TDL tanpa hidrolisis…………... 139
46 Analisis ragam laju pertumbuhan harian (%) ikan uji TDL terhidrolisis ……….
139
47 Uji Duncan laju pertumbuhan harian (%)TDL terhidrolisis………… 140
48 Analisis ragam laju pertumbuhan harian (%) TDL tanpa hidrolisis... 140
49 Uji Duncan laju pertumbuhan harian (%)TDL tanpa hidrolisis………
141
50 Nilai jumlah konsumsi pakan (g) dan efisiensi pakan (%) ikan uji TDL terhidrolisis………..
141
51 Nilai jumlah konsumsi pakan (g) dan efisiensi pakan (%) ikan uji TDL tanpa hidrolisis………
142
52 Analisis ragam konsumsi pakan (gr) ikan uji TDL terhidrolisis……. 142
53 Analisis ragam konsumsi pakan (gr) ikan uji TDL tanpa hidrolisis… 142
54 Analisis ragam efisiensi pakan (%) TDL terhidrolisis……… 143
55 Uji Duncan efisiensi pakan (%) TDL terhidrolisis………. 143
xx
59 Nilai retensi protein dan lemak pada ikan TDL tanpa hidrolisis……. 146
60 Analisis ragam retensi protein (%) TDL terhidrolisis ……… 147
61 Uji Duncan retensi protein (%)TDL terhidrolisis ……….. 147
62 Analisis ragam retensi protein (%)TDL tanpa hidrolisis………. 148
63 Uji Duncan retensi protein (%)TDL tanpa hidrolisis……….. 148
64 Analisis ragam retensi lemak (%) TDL terhidrolisis ……… 148
65 Uji Duncan retensi lemak (%)TDL terhidrolisis ……….. 149
66 Analisis ragam retensi lemak (%)TDL tanpa hidrolisis………. 149
67 Uji Duncan retensi lemak (%)TDL tanpa hidrolisis……… 149
68 Hepatosomatik indeks, kecernaan dan kadar glikogen hati ikan uji dengan TDL terhidrolisis………. 150 69 Hepatosomatik indeks, kecernaan dan kadar glikogen hati ikan uji dengan TDL tanpa hidrolisis……… 151 70 Analisis ragam hepatosomatik indeks (%) TDL terhidrolisis………. 151
71 Analisis ragam hepatosomatik indeks (%)TDL tanpa hidrolisis……. 152
72 Uji Duncan hepatosomatik indeks (%)TDL tanpa hidrolisis………… 152
73 Analisis ragam kadar glikogen hati (mg/100mg) TDL terhidrolisis… 152 74 Uji Duncan kadar glikogen hati (mg/100mg)TDL terhidrolisis…….. 153
75 Analisis ragam kadar glikogen hati (mg/100mg) TDL tanpa hidrolisis……….. 153 76 Uji Duncan kadar glikogen hati (mg/100mg)TDL tanpa hidrolisis……….. 153 77 Aktivitas enzim pencernaan pada usus ikan penelitian TDL terhidrolisis……… 154 78 Aktivitas enzim pencernaan pada usus ikan penelitian TDL tanpa hidrolisis……… 155 79 Analisis ragam aktivitas enzim selulase TDL terhidrolisis………….. 155
80 Uji Duncan aktivitas enzim selulase TDL terhidrolisis………... 156
81 Analisis ragam aktivitas enzim selulase TDL tanpa hidrolisis………. 156
82 Analisis ragam aktivitas enzim amilase TDL terhidrolisis…………... 156
83 Analisis ragam aktivitas enzim amilase TDL tanpa hidrolisis……….. 156
xxi
87 Analisis ragam aktivitas enzim protease TDL tanpa hidrolisis……… 158
88 Uji Duncan aktivitas enzim protease TDL tanpa hidrolisis... 158
89 Profil kadar glukosa darah (mg/100ml darah) perlakuan TDL
terhidrolisis………..
159
90 Rekapitulasi data parameter LPH, Konsumsi pakan, efisiensi pakan, retensi protein dan aktifitas enzim usus percobaan in vivo dengan perlakuan perbedaan persentase TDL terhidrolisis dalam pakan……. 160
91 Rekapitulasi data parameter LPH, Konsumsi pakan, efisiensi pakan, retensi protein dan aktifitas enzim usus percobaan in vivo dengan perlakuan perbedaan persentase TDL tanpa hidrolisis dalam pakan… 161
92 Rekapitulasi data parameter retensi lemak, HIS dan glikogen hati percobaan in vivo dengan perlakuan perbedaan persentase TDL
terhidrolisis dalam pakan………. 163
93 Rekapitulasi data parameter retensi lemak, HIS dan glikogen hati percobaan in vivo dengan perlakuan perbedaan persentase TDL
I. PENDAHULUAN
1.1. Latar belakang
Peningkatan permintaan ikan nila sebagai salah satu pilihan sumber protein
hewani dengan harga yang terjangkau untuk masyarakat di negara berkembang seperti
Indonesia, serta di pasar internasional mendorong meningkatnya usaha budidaya ikan
nila. Pada tahun 2004 Amerika Utara mengimpor nila sebesar 112.945 ton pada 2004,
meningkat 25% dibandingkan pada tahun 2003 dan 68% dibandingkan produksi pada
tahun 2002. Setengah dari impor Amerika Utara, dipasok oleh Cina, sedangkan
sisanya oleh Taiwan, Thailand dan Indonesia (FAO 2004). Produksi nila nasional
pada tahun 2004 sebesar 97.116 ton, dan kemudian meningkat sebesar 23,7% dalam
kurun waktu 2004-2008 menjadi sebesar 220.900 ton (DKP 2008). Peningkatan
produksi ini memposisikan Indonesia pada peringkat keempat negara produsen nila
terbesar di dunia setelah Cina, Mesir dan Philipina. Dengan adanya kasus KHV (Koi
Herpes Virus) pada ikan mas, nila menjadi alternatif ikan air tawar yang
dibudidayakan masyarakat dan salah satu andalan dalam program revitalisasi
perikanan. Perkembangan usaha budidaya ikan nila, sangat bergantung dengan pakan
buatan yang dapat mendukung pertumbuhan ikan nila yang optimal, sehingga
peningkatan kualitas pakan buatan menjadi suatu yang sangat penting.
Ikan nila mempunyai kemampuan untuk memanfaatkan pakan buatan yang
mengandung bahan-bahan dari tumbuhan (Fagbenro 1998; Olvera et al. 1997; E1-Sayed 1999; Fontainhas et al. 1999; Main et al. 2002; Ogunji dan Wirth 2000, Chimatiro dan Costa-Pierce 1996), bakteri (Beveridge et al. 1989), fitoplankton hijau (Pantastico et al.
1982). dan Spirulina (Lu dan Takeuchi 2004). Kemampuan dari ikan nila secara morfologi dan fisiologi untuk mencerna bahan pakan dengan kandungan serat tinggi
membuka peluang untuk pengembangan pakan buatan ikan nila dengan menggunakan
bahan pakan dari tepung daun dari berbagai jenis tanaman legumes seperti daun
lamtoro (El Sayed 1999).
Tepung daun lamtoro (selanjutnya disingkat TDL) merupakan sumber
daya hayati lokal yang potensial untuk digunakan sebagai salah satu sumber
protein nabati dalam pakan ikan. Hal ini disebabkan oleh tingginya kandungan
1982), komposisi asam amino yang relatif cukup baik serta kandungan Β
-karoten yang relatif tinggi (Agbede dan Aletor 2004). Menurut NAS (1994)
tanaman ini dapat menghasilkan bahan kering dari unsur-unsur yang dapat
dimakan sebesar 6 - 8 ton per hektar per tahun atau sekitar 20 - 80 ton bahan
segar. Di Indonesia tanaman leguminosa ini mudah ditanam sehingga dapat
membantu penyediaan pakan secara kontinyu sepanjang tahun.
Pemanfaatan bahan baku pakan ikan nila dari daun khususnya daun lamtoro
dibatasi oleh kandungan neutral detergent fiber (NDF) yang tinggi yaitu sebesar 39,5% dan acid detergent fiber (ADF) sebesar 35,10%. (Garcia et al. 1996), defisiensi asam amino esensial (Agr, Thr, He, His, Met) dan kandungan antinutrien
seperti mimosin dan asam fitat (Lim dan Dominy 1991; Wee dan Wang 1987;
Hertrampf dan Pascual 2000). Defisiensi asam amino dapat diatasi dengan
menambahkan asam amino esensial yang menjadi pembatas (Santiago dan Lovell
1988), sedangkan untuk mengatasi mimosin telah dilaporkan beberapa metode yang
efektif untuk mereduksi mimosin seperti perendaman dan pemanasan (Wee dan
Wang, 1987; Widyastuti 2001). Reduksi anti nutrient asam fitat sangatlah penting
sebelum penggunaan TDL dalam pakan, karena asam fitat dapat mengikat protein
dan mineral di dalam digesta dan berpotensi untuk menghambat aktivitas
enzim-enzim pencernaan (Conrad et al. 1996; Ravindran et al. 2000). Beberapa peneliti membuktikan bahwa penambahan enzim fitase mampu menghidrolisis
asam fitat menjadi orthofosfat anorganik dan senyawa inositol yang lebih
rendah. Ikan seperti hewan monogastrik lainnya tidak dapat mencerna asam
fitat karena ketiadaan enzim fitase di saluran pencernaan (Ravindran et al. 2000). Keterbatasan ikan dalam memanfaatkan serat berkaitan dengan ketersediaan
enzim selulotik yang terbatas dalam saluran pencernaan ikan. Kadar serat yang
berlebih pada pakan ikan pada level tertentu dapat menghambat pertumbuhan.
Beberapa penelitian melaporkan bahwa di dalam saluran pencernaan ikan ditemukan
aktifitas selulase dalam jumlah yang kecil (Saha dan Ray 1998; Prejs dan Blaszczyk
2006; Donovan et al. 2004; Li et al 2004; Bairagi et al. 2004; Nibedita et al. 2008). Jalilvand et al. (2008) melaporkan enzim fibrilotik eksogen sangat efektif untuk menurunkan kadar serat bahan baku pakan seperti jerami padi, dan
menghidrolisis tepung daun lamtoro sehingga dapat ditingkatkan kualitas nutriennya.
Penggunaan enzim eksogen ini terkendala dengan harga enzim komersil yang mahal di
pasaran dan biasanya enzim komersil dijual dengan spesifik aktifitas enzim tertentu.
Terkait dengan hal tersebut sangatlah penting dicari sumber enzim yang mengandung multi
enzim, murah dan efektif untuk meningkatkan kualitas nutrien dari tepung daun lamtoro.
Cairan rumen domba merupakan salah satu sumber bahan suplemen alternatif
yang murah dan dapat dimanfaatkan dengan mudah sebagai sumber enzim
hidrolase (Moharrery dan Das, 2002). Cairan rumen domba (Ovies aries) yang didapat dengan memeras isi rumen, merupakan limbah rumah pemotongan hewan
(selanjutnya disingkat RPH) ketersediaan cukup melimpah dan berpotensi sebagai
sumber enzim. Kung et al. (2000) melaporkan bahwa cairan rumen sapi mengandung enzim protease/ deaminase yang menghidrolisis protein atau peptida,
amilase yang menghidrolisis pati, selulase yang menghidrolisis selulosa,
hemiselulase (xylanase) yang menghidrolisis hemiselulosa (xylan), lipase yang
menghidrolisis emak, fitase yang menghidrolisis fitat dan lain-lain.
Potensi multienzim yang terkandung dalam cairan rumen diharapkan dapat
menghidrolisis nutrien dalam TDL sehingga kualitas nutrien TDL dapat
ditingkatkan. Selain mengandung enzim, cairan rumen domba juga mengandung
asam-asam amino, vitamin dan mineral. Peningkatan kulitas nutrient TDL akan
meningkatkan pula kinerja pertumbuhan dan pemanfaatan pakan ikan yang
mengandung TDL. Produk yang diekstraksi dari cairan rumen ini diharapkan dapat
secara langsung digunakan oleh pembudidaya ikan nila sehingga jauh lebih efisien
dibanding bila harus mendirikan sebuah industri enzim yang memerlukan investasi biaya
yang sangat besar.
1.2. Perumusan Masalah
Usaha untuk meningkatkan nilai guna TDL sebagai alternatif sumber
protein nabati pakan ikan nila mengalami kendala karena kandungan serat kasar dan
asam fitat yang tinggi, sedangkan di lain pihak ikan memiliki kemampuan terbatas
untuk mencerna pakan berserat dan dan kandungan asam fitat yang tinggi. Pendekatan
penggunaan enzim untuk menghidrolisis tepung daun lamtoro diharapkan dapat
dari TDL meningkat. Peningkatan kualitan nutrien ini diharapkan dapat meningkatkan
efisiensi pakan sehingga kinerja pertumbuhan ikan nila dapat lebih baik.
Potensi yang dimiliki oleh cairan rumen domba sebagai sumber
enzim-enzim hidrolisis seperti amilase, protease, lipase dan selulase menjadikannya sebagai
salah satu sumber bahan suplemen alternatif yang murah dan dapat dimanfaatkan
dengan mudah untuk meningkatkan kualitas nutrien dari tepung daun lamtoro,
sehingga kecernaan dapat meningkat dan pertumbuhan ikan nila dapat lebih optimal.
1.3. Tujuan Penelitian 1.3.1. Tujuan umum :
Meningkatkan kualitas nutrien TDL sebagai sumber protein nabati alternatif
pakan ikan nila dengan memanfaatkan dan mendayagunakan cairan rumen domba sebagai
sumber enzim hidrolisis (predigestion) 1.3.2. Tujuan khusus :
1. Menguji aktifitas enzim selulase, amilase, protease, lipase dan fitase dalam
ekstrak enzim cairan rumen domba yang dipelihara dengan pakan hijauan.
2. Mengkaji pengaruh hidrolisis (predigestion) TDL dengan ekstrak enzim cairan rumen domba terhadap kualitas nutrien TDL.
3. Menguji efektifitas pemanfaatan TDL terhidrolisis (predigestion) dalam pakan untuk ikan nila serta pengaruhnya pada perubahan aktifitas enzim pencernaan dan
metabolisme nutrient dalam tubuh ikan.
1.4. Manfaat Penelitian
Hasil penelitian ini diharapkan bermanfaat untuk pengembangan ilmu nutrisi
ikan khususnya untuk meningkatkan kualitas nutrien dari TDL dengan menggunakan
ekstrak enzim cairan rumen domba, yang selanjutnya dalam jangka panjang teknologi
ini dapat diaplikasikan dan dikembangkan dalam budidaya ikan nila.
1.5. Perumusan hipotesis
Jika hidrolisis TDL dengan enzim cairan rumen domba dapat meningkatkan kualitas
nutrien TDL maka penggunaan TDL dapat menghasilkan pertumbuhan dan pemanfaatan
pakan ikan nila yang lebih baik dibandingkan dengan pemakaian tepung daun lamtoro tanpa
II. TINJAUAN PUSTAKA
2.1. Kebutuhan Nutrien Ikan Nila
Komponen pakan yang berkontribusi terhadap penyediaan materi dan energi
untuk tumbuh adalah protein, karbohidrat dan lemak. Protein pada ikan
berperan sebagai sumber nutrient dan sebagai sumber energy. Protein
merupakan molekul kompleks yang terdiri dari asam amino esensial dan non
essensial. Protein adalah nutrien yang sangat dibutuhkan untuk perbaikan jaringan
tubuh yang rusak, pemeliharaan protein tubuh, penambahan protein tubuh untuk
pertumbuhan, materi untuk pembentukan enzim dan beberapa jenis hormon, dan
juga sebagai sumber energi (NRC 1993). Kebutuhan ikan akan protein dipengaruhi
oleh berbagai faktor diantaranya ukuran ikan, temperatur air, kadar pemberian
pakan, kandungan energi dalam pakan yang dapat dicerna dan kualitas protein
(Furuichi 1988).
Kebutuhan protein ikan berbeda-beda menurut spesiesnya, namun pada
umumnya ikan membutuhkan protein sekitar 30 - 40 % dalam pakannya (Jobling
1994). Ikan air tawar umumnya dapat tumbuh dengan baik dengan pemberian
pakan yang mengandung kadar protein 25 - 35 % dengan rasio energi protein
adalah sekitar 8 kkal/gram protein. Pada Tabel 1 disampaikan data perbedaan
kebutuhan protein ikan nila dengan bobot tubuh yang berbeda.
Defisiensi asam amino esensial (Agr,Thr, He, His, Met) dan kandungan
mimosin (Lim dan Dominy 1991) merupakan faktor pembatas dalam pemanfaatan
TDL dalam pakan ikan. Defisiensi asam amino dapat diatasi dengan
menambahkan asam amino esensial yang menjadi pembatas sedangkan untuk
mengatasi mimosin telah dilaporkan beberapa metode untuk mereduksi mimosin.
Seperti dilaporkan oleh Hasan et al. (1994), kecernaan TDL pada Labeo rorita
fingerling pada daun yang tidak direndam dalam air adalah 62,7%, sedangkan
kecernaan daun yang direndam dalam air adalah 82,7 %. Pada ikan Lobeo Rohita dan ikan nila, didapatkan bahwa dengan peningkatan kadar TDL dalam pakan baik dengan perendaman maupun tidak, terjadi penurunan pemanfaatan
sinar matahari memberikan pertumbuhan yang lebih baik pada nila
dibandingkan TDL yang ditambahkan sodium hidroksida.
Tabel 1. Kebutuhan protein ikan Nila Oreochromis sp dengan bobot tubuh yang berbeda.
Spesies Bobot tubuh
ikan (g)
Keperluan protein (%)
Pustaka1
O.mossambicus 1,0-2,5 29-38 Cruz dan Laudencia (1977)
Fry 50 Jauncey dan Ross (1982)
0,5-1,0 40 Jauncey dan Ross (1982)
6,0 - 30,0 30-35 Jauncey dan Ross (1982)
1,8 40 Jauncey (1982)
O. niloticus 1,5-7,5 36 Kubaryk (1980)
3,2-3,7 30 Wange et al. (1985)
0,838 40 Siddiqui et al. (1988)
40 30 Siddiqui et al. (1988)
O. aureus 0,3-0,5 36 Davis dan Stickney (1978)
0,16 40 Santiago dan Laron (1991)
di dalam El-Sayed (2006).
Ikan mempunyai kemampuan terbatas untuk mencerna serat, hal ini
berkaitan dengan terbatasnya ketersediaan enzim selulotik dalam saluran
pencernaan. Bahkan pencampuran TDL pada level tertentu dapat menghambat
pertumbuhan ikan. Beberapa penelitian melaporkan bahwa di dalam saluran
pencernaan ikan ditemukan aktifitas selulase dalam jumlah yang kecil (Saha dan Ray
1998; Prejs dan Blaszczyk 2006; Donovan et al. 2009; Li et al. 2004; Bairagi 2004; Nibedita dan Koushik 2008). Pada O. Mossambicus dan tilapia ukuran sejari dilaporkan oleh Jackson et al. (1982), serta Wee dan Wang (1987) bahwa pada penggunaan 25 % TDL dalam pakan berkadar protein 30%
mengakibatkan penurunan kinerja pertumbuhan dan efisiensi pemanfaatan
pakan. Sedangkan pada tilapia ukuran pembesaran (Santiago dan Lovell
(Badawy et al. 1995) pemakaian 15% TDL dalam pakan mengakibatkan pertumbuhan menjadi rendah.
Penyediaan sumber protein pakan baik tepung ikan dan bungkil kedelai
masih bergantung pada impor. Tumbuhan leguminosa dan sereal serta
produknya telah dicoba digunakan sebagai substitusi dari bungkil kedelai di
dalam pakan ikan nila (Meulen et al. 1979). Hal ini sangat memungkinkan digunakan untuk budidaya ikan nila karena ikan nila adalah ikan omnivora
yang cenderung herbivora sehingga lebih mudah beradaptasi dengan jenis
pakan yang dicampur dengan sumber bahan nabati seperti bungkil kedelai,
tepung jagung, tepung biji kapuk, tepung eceng gondok, tepung alfalfa, serta
tepung daun dari berbagai jenis tanaman legumes seperti daun lamtoro (El
Sayed 1999). Popma (1982) menjelaskan kemampuan ikan nila dan ikan air
tawar yang bersifat herbivora dan cenderung omnivora yang dapat mencerna
lebih dari 70% energi kotornya dari bahan non-strach, sedangkan pada ikan
yang bersifat karnivora seperti ikan trout, Wilson dan Poe (1987) melaporkan
hanya mampu mencerna kurang dari 50% .
Karbohidrat merupakan sumber energi yang penting meskipun kandungan
karbohidrat dalam pakan berada dalam jumlah yang relatif rendah. Karbohidrat
dalam pakan dapat berupa serat kasar serta bahan ekstrak tanpa nitrogen (BETN).
BETN mengandung banyak gula dan pati yang bersifat mudah dicerna
sedangkan serat kasar kaya akan lignin dan selulase yang sukar dicerna. Energi
dari karbohidrat sama efektifnya dengan energi dari lemak (NRC 1993).
Pemberian tingkat energi optimum dalam pakan sangat penting karena kelebihan dan kekurangan energi dapat menurunkan pertumbuhan ikan
(Lovell 1988).
Pemanfaatan karbohidrat oleh ikan berbeda-beda bergantung kepada
kompleksitas karbohidrat. Ikan-ikan karnivora tidak mampu memanfaatkan
karbohidrat kompleks/polisakarida sebagai energi utama dalam pakannya pada
kadar yang tinggi. Sedangkan ikan-ikan omnivora dan herbivora dapat
mencerna karbohidrat yang berasal dari tumbuh-tumbuhan. Ikan-ikan karnivora
pakannya sedangkan ikan-ikan omnivora mampu memanfaatkan karbohidrat
optimum sebesar 30-40% dalam pakan (Furuichi 1988).
Bungkil kedelai dan tepung ikan adalah bahan yang umum digunakan
sebagai sumber protein dalam pakan. Tetapi dengan meningkatnya permintaan
manusia akan protein di beberapa negara berkembang, serta diiringi dengan
meningkatnya harga bungkil kedelai dan tepung ikan di tingkat dunia haruslah
dicari bahan dasar lain sebagai sumber protein pakan yang sifat bahan tersebut
adalah lokal sehingga harganya tidak terlalu fluktuatif.
2.2. Tepung Daun Lamtoro (Leucaena leucocephala)
Kale (1987) menyatakan bahwa lamtoro adalah tumbuhan leguminosa
tropis, berasal dari Amerika Tengah. Tumbuhan ini disebarkan oleh orang-orang
Mayan dan Zapotec ke seluruh Amerika Tengah. Klasifikasi Leucaena
leucocephala menurut Brewbaker dan Hylin (1965) adalah, salah satu spesies dari genus Leucaena yang termasuk sub Famili Mimosoideae, Famili Leguminoseae,
sub ordo Rosicae, Ordo Resales, Sub Klas Dycotyledonea, Klas
Angiospermopsidae, sub Divisio Spermatophyta, Divisio Traceophyta dan sub
Kingdom Embryobionta.
Lamtoro (Leucaena) terdiri atas 53 spesies yang digolongkan ke dalam 10 spesies yang telah dikenal. Walaupun seluruh spesies tersebut mungkin sangat
berguna bagi daerah tropis, tetapi hanya Leucaena leucocephala yang telah dimanfaatkan secara luas (NAS 1994). Tanaman lamtoro tumbuh baik di daerah
dengan curah hujan tahunan antara 1000 - 3000 mm. Sementara Garcia et al.
(1996) menyarankan agar tanaman lamtoro ditanam di daerah yang curah
hujannya lebih dari 750 mm per tahun dan ketinggian lebih dari 1500 m dpi.
Selanjutnya dinyatakan pula bahwa tanah yang sesuai dengan tanaman ini adalah
tanah yang netral atau tanah basah (padang penggembalaan ternak), dimana
lamtoro dan rumput makanan ternak ditanam menjadi satu dengan perbandingan
yang tepat. Menurut Bray et al. (1997) tanaman lamtoro dapat ditanam dengan menggunakan berbagai sistem penanaman misalnya sebagai pagar pencegah erosi,
penahan angin atau batas tanah pekarangan. NAS (1994) menyebutkan bahwa
unsur-unsur yang dapat dimakan (daun dan ranting-ranting kecil) sebesar 6-8 ton per
hektar per tahun atau sekitar 20 - 80 ton bahan segar per hektar pertahun.
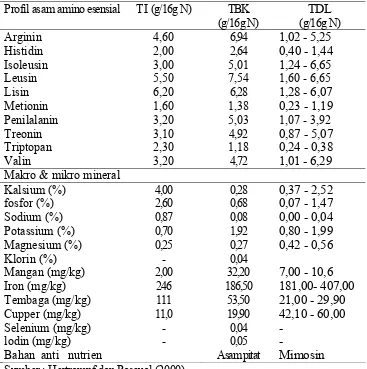
Tabel 2. Perbandingan komposisi asam amino dan makro, mikro serta mineral, antara tepung ikan (TI), bungkil kedelai (TBK) dan tepung daun lamtoro (TDL).
Profil asam amino esensial TI (g/16g N) TBK (g/16g N)
Makro & mikro mineral
Kalsium (%) 4,00 0,28 0,37 - 2,52
Sumber ; Hertrampf dan Pascual (2000)
TDL merupakan sumber daya hayati lokal yang potensial untuk
digunakan sebagai salah satu sumber protein nabati dalam pakan ikan karena
mengandung protein sekitar 34,38 % (Agbede dan Alevator 2004); 25 - 30%
(NAS 1994); 24,2% (Sutardi 1981); 24% (Scott et al. 1982), yang merupakan nilai tertinggi dibandingkan sumber protein nabati lainnya. Komponen asam
amino essensial TDL, bungkil kedelai dan tepung ikan dapat dilihat pada Tabel 2.
amino essensial dibandingkan bungkil kedelai, walaupun masih jauh dari
kandungan asam amino tepung ikan. Tepung daun lamtoro juga merupakan
sumber vitamin A dengan kandungan β-karoten yang relatif tinggi serta
kandungan xantofil yang merupakan sumber pigmentasi pada kulit dan kuning
telur (Hertrampf dan Pascual 2000).
Pemanfaatan bahan baku pakan ikan nila dari daun tumbuhan khususnya
daun lamtoro dibatasi dengan tingginya kandungan neutral detergent fiber (NDF) sebesar 39,5% dan acid detergent fiber (ADF) sebesar 35,10% (Garcia et al. 1996). Serat kasar merupakan komponen karbohidrat yang kaya akan lignin dan selulase
yang sukar dicerna. Selulase merupakan kerangka sel tanaman yang terdiri dari
rantai D-Glukosa dengan derajat polimerasi sebesar lebih kurang 14.000 (Schlegel
1994). Degradasi polisakarida yang terdapat pada dinding sel tanaman yang
merupakan bagian terbesar komponen serat kasar yang kadarnya bervariasi
bergantung kepada jaringan tanaman, jenis tanaman dan umur tanaman (Amin
1997). Pada manusia fungsi utama selulase adalah untuk menyediakan bahan bulky
(tidak dapat dicerna) yang dapat meningkatkan efisiensi kerja saluran pencernaan
yang fungsinya dapat disamakan dengan fungsi serat dalam pakan ternak
(Djojosoebagio dan Pilliang, 1996).
Pemanfaatan TDL di dalam pakan dibatasi pula oleh adanya mimosin
yang merupakan asam amino heterosiklik
(a-amino-(3(N-(3-hidroxy-4-piridon)(asam propionat). Berbagai usaha yang dilakukan untuk menurunkan daya
racun mimosin dalam daun lamtoro adalah dengan pemanasan, penambahan garam
sulfat, penambahan senyawa analog mimosin, pencucian, mendapatkan varietas baru
yang rendah kandungan mimosinnya. Murthy et al. (1994) melaporkan bahwa pengeringan dengan matahari sampai menjadi bahan kering lebih dari 90%.
kemudian dilanjutkan pengovenan pada suhu 100 °C selama 12 jam, dilanjutkan
perendaman dalam air selama 12 jam serta inkubasi dalam larutan 5% NaOH,
menghasilkan penurunan mimosin terbaik dan kehilangan protein yang terkecil. Setelah
dicobakan ke ayam broiler dapat memberikan pertambahan bobot badan dan
dapat memberikan pertumbuhan harian sebesar 2,2%, sedangkan pada kadar
yang sama TDL yang tidak direndam memberikan pertumbuhan harian sebesar
1,1%. Penaflorida et al. (1992) menyatakan bahwa perendaman daun lamtoro selama 30 - 48 jam dapat menurunkan mimosin hingga 90%. Sedangkan penelitian
Widyastuti (2001) menunjukkan bahwa ayam broiler yang ransum dasarnya
dicampur TDL 10% dengan perlakuan pemanasan kering/oven pada suhu 70°C
selama 12 jam memberikan respon pertumbuhan yang terbaik. Dinyatakan pula
bahwa metode ini dinilai paling ekonomis dan mudah diterapkan dibanding
pemanasan lembab atau penambahan komponen zat besi.
Selain mimosin bahan nutrien lain yang terkandung dalam TDL adalah
mio-inositol heksakisfosfat (C6H18O24P6) yang umum disebut asam fitat
mempunyai rumus kimia dan struktur cincin yang mirip dengan glukosa yang
berikatan dengan fosfor untuk membentuk struktur asam fitat. Struktur asam
fitat dapat dilihat pada Gambar 1 Asam fitat mempunyai struktur yang stabil
dan mengandung kira-kira 2/3 fosfor tanaman dalam bentuk fosfor organik
(Ravindran et al. 2000).
Gambar 1. Struktur molekul asam fitat (Ravindran et al. 2000)
Sifat antinutrien asam fitat didasarkan pada kemampuannya untuk
bergabung dengan mineral bervalensi dua, seperti Ca, Zn, Mg dan Fe dan
membentuk fitat mineral yang tidak larut (Wodinski dan Ullah 1996). Sifat
anti nutrien yang lain adalah kemampuan asam fitat untuk berikatan dengan
protein, vitamin dan polisakarida. Kelompok fosfor yang terikat fitat dapat
membentuk ikatan elektrostatik dengan asam-asam amino atau dengan asam
amino bebas dari residu lisin dan arginin yang terdapat pada molekul protein
(Cheryan 1980). Kompleks fitat-mineral-protein dalam bentuk kation
kelompok terminal karboksil pada protein atau kelompok karboksil bebas dari
residu aspartat dan glutamat dalam molekul protein (Cheryan 1980). Fitat
dapat mengikat protein dan mineral di dalam digesta, sangat potensial untuk
menghambat aktifitas enzim-enzim pencernaan seperti protease dan tripsin
(Conrad et al. 1996). Penelitian lain menyatakan bahwa interaksi antara asam fitat dan protein akan menurunkan bioavailability protein pada legum (Davies
1982). Fosfor terikat fitat tidak dapat dimanfaatkan ternak dan terbuang dalam
feses sehingga akan meningkatkan kandungan fosfor dalam tanah dan air.
Mekanisme kerja asam fitat mengikat mineral dapat dilihat pada Gambar 2.
.
Gambar 2. Mekanisme asam fitat mengikat mineral (Cheryan 1980)
Penelitian untuk mereduksi fitat dalam bahan baku pakan dilakukan
dengan menambahkan enzim fitase yang merupakan enzim yang mampu
mengkatalisasi reaksi hidrolisis asam fitat dan menghasilkan orthofosfat
anorganik dan senyawa inositol yang lebih rendah. Enzim fitase ini terdapat
pada jaringan hewan, tanaman dan mikroba (Baruah et al. 2004).
Mekanisme fitase untuk memotong ikatan tersebut dapat dilihat pada Gambar
3. Wodinski dan Ullah (1996) mengemukakan bahwa terdapat 2 jenis enzim
fitase yaitu 3-fitase dari fungi (seperti jenis komersial yang dijual sekarang
yaitu adalah Selenomona ruminantium (Yanke et al. 1998) dan yang terbaru adalah
Mitsuokella jalaludinii (Lan et al. 2002).
Gambar 3. Mekanisme hidrolisis fosfat oleh enzim fitase (Applegate et al. 2004).
2.3. Cairan Rumen sebagai Sumber Enzim
Perut hewan ruminansia terdiri atas rumen, retikulum, omasum dan
abomasums (Gambar 4). Volume rumen pada ternak domba berkisar 10 liter.
Rumen diakui sebagai sumber enzim pendegradasi polisakarida. Polisakarida
dihidrolisis di rumen karena pengaruh sinergis dan interaksi dari komplek
mikro-organisme, terutama sellulase dan xilanase (Trinci et al. 1994). Mikroorganisme terdapat pada cairan rumen (liquid phase) dan menempel pada digesta rumen.
Enzim yang aktif mendegradasi struktural polisakarida hijauan
kebanyakan aktif pada mikro organisme yang menempel pada partikel
pakan. Di dalam retikulo rumen terdapat mikrobia rumen yang terdiri atas
protozoa dan bakteri yang berfungsi melaksanakan fermentasi untuk mensintesis
asam amino, vitamin B-komplek dan vitamin K sebagai sumber zat makanan bagi
hewan induk semang (Hungate 1966). Mikroba rumen dapat dibagi dalam tiga
grup utama yaitu bakteri, protozoa dan fungi (Czerkawski 1986) Beberapa
jenis bakteri yang dilaporkan oleh Hungate (1966) adalah : (a) bakteri
pencerna selulosa (Bakteroidessuccinogenes, Ruminococcus flavafaciens, Ruminococcus albus, Butyrifibriofibrisolvens), (b) bakteri pencerna
hemiselulosa (Butyrivibrio fibrisolvens,Bakteroides ruminocola,
Ruminococcus sp), (c) bakteri pencerna pati (Bakteroides ammylophilus, Streptococcus bovis, Succinnimonas amylolytica, (d) bakteri pencerna gula (Triponema bryantii, Lactobasilus ruminus), (e) bakteri pencerna protein (Clostridium sporogenus, Bacillus licheniformis). Kehadiran fungi dalam rumen diakui sangat bermanfaat bagi pencernaan pakan serat, karena dia
membentuk koloni pada jaringan selulosa pakan. Rizoid fungi tumbuh jauh
menembus dinding sel tanaman sehingga pakan lebih terbuka untuk dicerna
oleh enzim bakteri rumen. Protozoa rumen diklasifikasikan menurut
morfologinya yaitu: holotrichs yang mempunyai silia hampir di seluruh
tubuhnya dan mencerna karbohidrat yang fermentabel, sedangkan oligotrichs
yang mempunyai silia sekitar mulut dan umumnya merombak karbohidrat
yang lebih sulit dicerna (Arora, 1989).
Mikroba-mikroba rumen mensekresikan enzim-enzim pencernaan ke dalam
cairan rumen untuk membantu mendegradasi partikel makanan. Enzim-enzim
tersebut antara lain enzim yang mendegradasi substrat selulase yaitu selulase,
hemiselulase/xylosa adalah hemiselulase/xylanase, pati adalah amilase, pektin
adalah pektinase, lipid/lemak adalah lipase, protein adalah protease dan lain-lain
(Kamra 2005). Aktivitas enzim dalam cairan rumen juga tergantung dari
komposisi atau perlakuan makanan (Moharrery dan Das 2002). Agarwal et al.
(2002) melaporkan, rumen anak domba dengan berat badan 23,5 kg yang diberi
persen rumput sampai umur 24 minggu, pada cairan rumennya didapatkan enzim
carboxymethyl celulase dengan aktivitas enzim 3,60 µmol glukosa per jam per ml,
alpha amilase 0,33 µmol glukosa per menit per ml, xylanase 0,29 µmol xylosa per
menit per ml, beta-glukosidase 0,20 µmol p-nitrophenol per menit per ml,
alpha-glukosidase 0,008 mol p-nitrophenol per menit per ml, urease 0,05 µmol NHs-N
per menit per ml dan protease 452,7 µg hidrolisis protein per jam per ml. Martin
et al. (1999) mendapatkan bahwa enzim-enzim pencerna karbohidrat dalam cairan rumen antara lain adalah amilase, xylanase, avicelase, D-glukosidase,
alpha-L-arabinofuranosidase, beta-D-glukosidase dan beta-D-xylosidase. Martin et al. (1999) juga melaporkan bahwa aktivitas enzim-enzim pencernaan dalam cairan
rumen dipengaruhi oleh posisi rumen, dimana pada (bagian perut (ventral) dan
bagian punggung (dorsal) terdapat protozoa dan bakteri berbeda. Aktivitas
enzim-enzim fibrolitik (xylanase, avicelase, alpha-L-arabinofuranosidase,
beta-D-glukosidase dan beta-D-xylosidase) yang berasal dari mikroba protozoa bagian
punggung (dorsal) yang lebih besar / lebih tinggi sekitar 40 persen dari bagian
perut (ventral), sebaliknya aktivitas enzim-enzim fibrolitik yang berasal dari
bakteri lebih besar di bagian perut (ventral) dari pada bagian punggung
(dorsal). Martin et al. (1999) juga mendapatkan bahwa aktivitas enzim yang berasal dari bakteri lebih tinggi dari pada yang berasal dari protozoa. Di dalam
retikulo rumen Moharrery dan Das (2002) yang mengukur aktivitas enzim protease,
selulase, amilase, lipase dan urease pada cairan rumen domba mendapatkan bahwa
cairan rumen tanpa protozoa tetapi masih mengandung sel-sel bakteri dan cairan
rumen yang berisi enzim-enzim dari sel-sel bakteri, aktivitas enzimnya lebih
tinggi dari cairan rumen tanpa protozoa dan tanpa sel-sel bakteri. Lee et al. (2002) memetakan enzim-enzim dalam cairan rumen sapi. Enzim-enzim yang terdapat
dalam cairan rumen sapi antara lain adalah enzim-enzim selulolitik terdiri atas
beta-D-endoglukanase, beta-D-exoglukanase, beta-D-glukosidase, dan beta-D-fucosida
fucohydrolase, enzim-enzim xylanolitik terdiri atas beta-D-xylanase, beta-D-xylosidase,
acetyl esterase, dan alfa-L-arabinofuranosidase, enzim-enzim pektinolitik terdiri atas
polygalacturonase, pectate lyase dan pectin lyase, dan enzim-enzim lain yang terdiri
atas beta-amilase, endo-arabilase, beta-D-gluanase (laminarinase), beta-D-glucanase
cairan rumen sapi dan aktivitas enzimnya disajikan pada Tabel 3 dan Tabel 4.
Sedangkan pada Tabel 5 disajikan hasil penelitian Budiansyah (2010) tentang
karakteristik enzim rumen sapi lokal yang mendapat makanan hijauan yang kaya serat.
Tabel 3. Komposisi enzim cairan rumen sapi1)
Enzim Lee et al. (2002)1 Agarwal et al
µmol glukosa/ jam /ml
11 83,7 ±20,39
µg hidrolisis protein
125,6 ±3,83
µg hidrolisis protein
452,7 ± 154,3 µg hidrolisis protein /j / l
Keterangan: 1) Sapi dewasa yang diberi makan ransum dasar alfalfa
2)
Anak domba dengan berat badan 23,5 kg yang diberi air susu sampai 8 minggu dan diteruskan dengan 50 persen konsentrat dan 50 persen rumput sampai umur 24 minggu Selulase -Fpase (ug glukosa / ml/ jam)
Tabel 4. Komposisi enzim cairan rumen domba1)
Enzim Cairan Rumen
Tanpa Protozoa
Keterangan1) Sumber : Moharrery dan Das (2002).
Tabel 5. Karakteristik enzim-enzim cairan rumen sapi asal RPH
Parameter Enzim-enzim cairan rumen
Selulase Amilase Fitase Protease
Kisaran pH pH 4 – 9 pH 6 - 9 pH 4 - 9 pH 5 - 9 Sapi impor
Suhu 39 oC 50 oC 39 oC 70 oC
Kisaran Suhu 29-50 oC 50 oC 29-50 oC 39-70 oC
pH optimum pH 4 pH 7 pH 6 pH 7
Kisaran pH pH 4 – 9 pH 6 - 9 pH 4 - 9 pH 4 - 9
Kohn dan Alien (1994) menggunakan enzim protease dari ekstrak cairan
rumen sapi untuk mengukur laju degradasi protein bungkil kedelai dan hay
lucerne. Enzim protease hasil ekstraksi dengan butanol dan aseton hanya tersisa
62 persen aktivitasnya dibanding cairan rumen awal. Tidak ada perbedaan antara
taraf enzim 3, 5 atau 10 ml dalam mendegradasi protein pakan. Protein bungkil
kedelai terdegradasi dengan kecepatan 0,15 mg per gram per jam pada 2 jam
pertama dan turun menjadi 0,01 mg per gram per jam dari 8 sampai 24 jam
berikutnya. Kejadian yang sama juga terjadi pada protein hay lucerne, degradasi
protein dengan kecepatan 0,06 mg per gram per jam pada 2 jam pertama dan turun
menjadi 0,01 mg per gram per jam dari 8 sampai 24 jam berikutnya. Laju
degradasi protein pakan lebih rendah dari yang diukur sebelumnya dengan
menggunakan mikroba hidup. Penggunaan cairan rumen sapi sebagai sumber enzim
kasar telah dicobakan ke dalam ransum unggas berbasis wheat pollard dengan
adanya perbaikan terhadap performa ayam broiler (Pantaya et al. 2005). Budiansyah (2010) melaporkan pula bahwa performa ayam broiler yang lebih baik dengan
penambahan 0,5 % enzim cairan rumen sapi dalam ransum.
2.4. Enzim Pencernaan dan Perannya dalam Proses Pencernaan.
Dalam proses pencernaan, makanan yang dicerna dipecah menjadi
molekul-molekul yang lebih sederhana sehingga mudah diserap melalui dinding usus dan
masuk ke dalam aliran darah. Pencernaan merupakan proses yang berlangsung
terus menerus yang bermula dari pengambilan pakan dan berakhir dengan
pembuangan sisa pakan. Pencernaan pakan meliputi hidrolisis protein menjadi
asam-amino atau polipeptida sederhana, karbohidrat menjadi gula sederhana dan
lipid menjadi gliserol dan asam lemak. Proses pencernaan secara fisika maupun
adanya enzim perncernaan seperti protease, karboksilase dan lipase (Zonneveld
et al. 1991).
Kecernaan didefinisikan sebagai bagian pakan yang diserap oleh hewan
(Lovell 1988). Pengetahuan tentang kecernaan bahan pakan sangat diperlukan
dalam mempelajari kebutuhan energi ikan dan penilaian dari berbagai bahan
pakan yang berbeda. Selama pakan berada dalam usus ikan, nutrient yang
dicerna oleh berbagai enzim menjadi bentuk yang dapat diserap oleh dinding usus
dan masuk ke dalam sistem peredaran darah. Kecernaan ikan terhadap bahan
baku pakan dipengaruhi oleh beberapa faktor yaitu, sifat kimia air, suhu air, jenis
pakan, ukuran, umur ikan, kandungan gizi pakan, frekuensi pemberian pakan,
sifat fisika dan kimia pakan serta jumlah dan macam enzim pencernaan yang
terdapat dalam saluran pencernaan pakan (NRC 1993; Tillman et al. 1991; Hepher 1990).
Enzim adalah katalisator biologis dalam reaksi kimia yang sangat dibutuhkan
dalam kehidupan. Enzim adalah protein, yang disintesis dalam sel dan dikeluarkan dari
sel yang membentuknya melalui proses eksositosis. Enzim yang disekresikan ke luar
sel digunakan untuk pencernaan di luar sel (di dalam rongga pencernaan) atau disebut
"extra cellular digestion", sedangkan enzim yang dipertahankan dalam sel digunakan
untuk pencernaan dalam sel itu sendiri atau disebut "intra celuller digestion" (Affandi
el at. 1992). Enzim pencernaan yang disekresikan dalam rongga pencernaan berasal dari sel-sel mukosa lambung, pilorik kaeka, pankreas dan mukosa usus (Halver dan
Hardy 2002).
Oleh karena itu perkembangan sistem pencernaan erat kaitannya dengan
perkembangan aktivitas enzim dalam rongga saluran pencernaan. Enzim-enzim
tersebut berperan sebagai katalisator dalam hidrolisis protein, lemak dan karbohidrat
menjadi bahan-bahan yang sederhana. Sel-sel mukosa lambung menghasilkan
enzim protease dengan suatu aktivitas proteolitik optimal pada pH rendah. Pilorik
kaeca yang merupakan perpanjangan dari usus yang berfungsi mensekresikan enzim
yang sama seperti yang dihasilkan pada bagian usus yaitu enzim pencernaan
protein, lemak dan karbohidrat yang aktif pada pH netral dan scdikit basa. Cairan
pankreatik kaya akan tripsin, yaitu suatu protease yang aklivitasnya optimal sedikit
dan lipase. Ikan yang tidak memiliki lambung dan pilorik kaeka, aktivitas proteolik
terutama berasal dari cairan pankreatik. (Watford dan Lam 1993 ).
Kecernaan (digestibility) dipengaruhi oleh tiga faktor yaitu (1) jenis pakan yarg dimakan dan kadar kepekaan pakan terhadap pengaruh enzim pencernaan, (2)
aktivitas enzim-enzim pencernaan dan (3) lama waktu pakan yang dimakan
terkena aksi enzim pencernaan. Masing-masing faktor di atas dipengaruhi oleh
berbagai faktor sekunder yang berkaitan dengan ikan itu sendiri (spesies, umur,
ukuran) dan kondisi fisiologis, yang berkaitan dengan lingkungan (temperatur),
dan yang berkaitan dengan pakannya (komposisi pakan, ukuran partikel dan
jumlah pakan yang dimakan). Kecernaan berbeda antar spesies ikan, hal ini terjadi
akibat perbedaan sistem dan enzim-enzim pencernaan (De Silva dan Anderson 1995).
Kemampuan ikan dalam mencerna makanan sangat bergantung pada
kelengkapan organ pencernaan dan ketersediaan enzim pencernaan.
Perkembangan saluran pencenaan tersebut berlangsung secara bertahap dan
setelah mencapai ukuran/umur tertentu saluran pencernaan mencapai
kesempurnaan. Perkembangan struktur alat pencernaan ini diikuti oleh
perkembangan enzim pencernaan dan perubahan kebiasaan makan (food habit). Kandungan nutrien pakan nampaknya berpengaruh pada aktivitas enzim pencernaan.
Kuzmina (1996) mengungkapkan bahwa tersedianya substrat merupakan faktor
yang nyata dalam pengaturan aktivitas enzim pada ikan dan mamalia. Kandungan
protein pakan yang tinggi dikaitkan dengan kandungan selulase yang rendah
dapat meningkatkan aktivitas protease pada ikan rainbow trout . Peningkatan
proporsi pati kentang dalam pakan dari 10 menjadi 90% yang diikuti penurunan
proporsi tepung ikan akan meningkatkan aktivitas enzim maltase dan amilase pada
ikan mas (Hepher 1990).
Stickney dan Shumway (1974) menyatakan bahwa enzim selulase
diproduksi oleh mikroflora usus, yang dihubungkan dengan aktivitas selulase
dalam usus dengan jumlah selulase/bakteri selulitik. Bairagi et al. (2004) tentang aktifitas dua mikroba dari strain Bacillus yaitu Bacillus subtilus dan Bacillus circulans dari saluran pencernaan ikan mas dan ikan nilayang memproduksi enzim amilase, selulase, protease dan lipase. Das dan Tripathi (1991)
diberi pakan dari makanan yang mengandung tetrasiklin. Pemanfaatan daun
lamtoro sangat dibatasi oleh kecernaan ikan yang terbatas terhadap jenis
dedaunan ini. Hal ini berkaitan dengan ketersediaan enzim selulotik yang
terbatas dalam saluran pencernaan ikan. Beberapa penelitian telah melaporkan
bahwa ikan tidak memiliki enzim selulase dan kemungkinan adanya populasi
mikroba selulotik di saluran pencernaan ikan juga masih menjadi kontroversi di
kalangan peneliti (Stickney dan Shumway 1974; Prejs dan Blaszczyk 2006;
Linsday dan Harris 1980; Lessel dan Lesel 1986; Luczkovich dan Stellway 1993;
Saha dan Ray 1998). Kontrofersi tersebut terbantahkan dengan penelitian Prejs
dan Blaszczyk 2006; Donovan dan Rosalie 2009; Li et al. 2004; Nibedita dan Koushik 2008 yang mendapatkan aktifitas enzim selulase pada saluran
pencernaan ikan.
Enzim protease menguraikan rantai-rantai peptida dari protein. Peptidase
diklasifikasikan menjadi endopeptidase dan eksopeptidase yang bergantung pada
letak ikatan peptida pada tengah atau akhir molekul. Endopeptidase
menghidrolisis protein dan peptida-peptida rantai panjang menjadi peptida-peptida
pendek. Endopeptidase penting antara lain pepsin yang dihasilkan dari zimogen
pepsinogen, tripsin dari tripsinogen, dan kimotripsin dari kimotripsinogen.
Eksopeptidase menghidrolisis peptida menjadi asam-asam amino.
Karboksipeptidase, aminopeptidase, dan dipeptidase termasuk dalam kelompok
eksopeptidase. Alfa-amilase adalah enzim yang bertanggung jawab menghidrolisis
pati menjadi glukosa. Enzim ini memutuskan ikatan 1,4-α -glukosidik dan
mengubah pati menjadi glukosa dan maltosa. Lipase adalah enzim penting dalam
pencemaan lemak. Lipase memecah lemak menjadi gliserol dan asam lemak
(Steffens 1989; Hepher 1990).
Enzim berperan dalam mengubah laju reaksi, sehingga kecepatan reaksi
yang diperlihatkan dapat dijadikan ukuran keaktifan enzim. Satu unit enzim
adalah jumlah enzim yang mengkatalisis transformasi 1 mikromol substrat dalam
waktu 1 menit pada suhu 25°C dan pada keadaan pH optimal. Aktivitas enzim
bergantung pada konsentrasi enzim dan substrat, suhu, pH dan inhibitor.
Dinyatakan pula bahwa enzim pencernaan yang dihasilkan oleh lambung ikan aktif
III. METODOLOGI
Penelitian dilakukan dalam tiga tahap.
Tahap Pertama : Pengujian aktifitas enzim-enzim hidrolisis pada ekstrak enzim cairan rumen domba
Tahap kedua : Pengaruh penambahan ekstrak enzim cairan rumen domba (in vitro) terhadap kualitas tepung daun lamtoro (TDL).
Tahap ketiga : Efektifitas pemanfaatan TDL terhidrolisis (predigestion) dalam pakan buatan untuk ikan nila serta pengaruhnya pada perubahan aktifitas enzim pencernaan dan metabolisme nutrien.
3.1. Tahap Pertama
Pengujiaan Aktifitas Enzim-enzim Hidrolisis Cairan Rumen Domba
Pada percobaan ini sasaran yang ingin dicapai adalah mendapatkan eksrak enzim cairan rumen domba yang dipelihara dengan pakan hijauan, dan menguji diuji aktifitas enzim selulase, fitase, amilase dan protease. Data aktifitas enzim yang didapatkan akan menjadi dasar perhitungan jumlah enzim yang ditambahkan pada substrat TDL.
3.1.1. Isolasi dan Produksi Ekstrak Enzim Cairan Rumen Domba.
3.1.2. Uji Aktifitas Ekstrak Enzim Cairan Rumen Domba
Ekstrak cairan rumen domba yang telah dilarutkan dalam cairan buffer phosfat dengan perbandingan 1 : 1, selanjutkan akan diuji kandungan aktifitas enzim selulase, amilase, protease dan lipase. Uji aktifitas enzim selulase/FP-ase dilakukan menurut metode Ghosse, (1987) dengan prosedur kerja pada Lampiran 1, aktifitas enzim amilase dan enzim protease dilakukan menurut metode Bergmeyer dan Grassi (1983) dengan prosedur kerja pada Lampiran 2 dan 3. aktifitas enzim lipase dilakukan menurut metode Tietz dan Friedreck dalam Barlongan, (1990) dengan prosedur kerja pada Lampiran 4 dan aktifitas enzim fitase dilakukan menurut metode Greiner et al. (1997) dengan prosedur kerja pada Lampiran 5.
3.2. Tahap Kedua
Pada percobaan tahap kedua, sasaran yang ingin dicapai adalah untuk mendapatkan informasi kualitas TDL yang mendapat perlakuan dengan penambahan ekstrak enzim cairan rumen domba dengan taraf dosis dan waktu inkubasi yang berbeda. TDL dengan kualitas terbaik selanjutnya akan dievaluasi pada pengujian in vitro untuk bahan campuran pakan ikan nila.
3.2.1. Pengaruh penambahan enzim cairan rumen domba (in vitro) terhadap kualitas tepung daun lamtoro
Penelitian dilakukan di Laboratorium Nutrisi Ternak Perah Fakultas Peternakan Institut Pertanian Bogor, untuk menguji pengaruh penambahan enzim cairan rumen domba dengan taraf dosis dan waktu inkubasi yang berbeda terhadap kualitaas TDL.