___________________________
Corresponding author:Babić Vojka, Maize Research Institute, Zemun Polje, Slobodana Bajića 1, 11185 Belgrade, Serbia; Tel: +38111 37 56 704; Fax: +38111 37 56 707; e-mail:
vbabic@mrizp.rs
UDC 575:633.15 DOI: 10.2298/GENSR1302589B
Original scientific paper
THE IDENTIFICATION OF BANDS RELATED TO YIELDS AND STABILITY IN MAIZE HYBRIDS AND THEIR PARENTAL COMPONENTS
Vojka BABIĆ1, Slaven PRODANOVIĆ2, Milosav BABIĆ1, Nenad DELIĆ1, Violeta ANĐELKOVIĆ1
1
Maize Research Institute, Zemun Polje, Belgrade, Serbia
2
Faculty of Agriculture, University of Belgrade, Belgrade, Serbia
Babić V., S. Prodanović, M. Babić, N. Delić and V. Anđelković (2013): The identification of bands related to yields and stability in maize hybrids and their parental components. Genetika, Vol 45, No. 2, 589-599.
Geneticists have been trying to explain adaptability and stability of a genotype in terms of a desirable combination of alleles expressed via epistasis. Stability determined in one set of data is very often stability of a genotype to a prevalent stress factor in a moment of carrying out the experiment. However, grain stability of a certain genotype can be a result of different factors, such as tolerance to drought, or to some important diseases and pests. Yield and yield stability of 15 maize hybrids were observed in 24 environments. The level of the interaction effect of studied maize hybrids was identified by the AMMI analysis, while a number of bands positively related to yield and stability of studied maize hybrids were identified by a genetic characterisation by RAPD markers. Bands positively related to yields were present to a greater extent in parents originating from the BSSS population, while bands positively related to stability were more present in parents originating from the Lancaster population.
Key words: maize hybrids, RAPD markers, yield stability INTRODUCTION
Yields stability was recognised as an important objective of plant breeding. It is partially influenced by a genetic structure, and partially by heterogeneity and heterozygosity of a variety. The majority of scientists define variety stability as genetic ability of a variety to accomplish stable and high yield under various environmental conditions. Stability, in terms of agronomy, is ability of genotypes to always result in the uniform yield regardless of effects of environmental conditions. Geneticists have been trying to explain adaptability in terms of a desirable combination of alleles expressed via epistasis, while physiologists emphasised the response of a
genotype to stress and genes that were activated by this response (DIMITRIJEVIĆ and PETROVIĆ, 2000). CATTIVELLI et al. (2002) concluded that little was known about mechanisms that control stress as each response to stress is influenced by a great number of genes. Although great attention has been paid to this issue, it is still little known about genetic components that determine stability and about effects of the selection process on stability (LEE et al., 2003). Early concepts of stability based on linear regression (LR) much affected former breeding programmes. The interaction could be qualitatively analysed by these concepts only if certain data were well fit to the regression model (ZOBEL et al., 1988). In recent times, many researches leaved the concept that tried to summarise a great number of vectors included into the interaction into one universal parameter of stability. So called hybrid models such as Additive Main effects and Multiplicative Interactions (АММI) model, then Sites Regression Analysis (SREG), Genotype main effects and Genotype x Environment interaction model (GGE) (GUNJAČA, 2001; STOJAKOVIĆ et al., 2012) have been applied. By comparing АММI analysis, GGE analysis and PCA analysis, GAUCH (2006) stated in his study that these related models basically had similar results. The best results could be achieved when agronomic understanding of yield trials (genotypes and environments) was combined with knowledge of statistical and experimental design.
Following the literature impression, LAMKEY and LEE, (1993) stated, that the issue of the GxE interaction was accessed more from the statistical than form the biological aspect. They considered that such an approach do not play a great role in the plant breeding improvement. The genetic analysis of complex traits such as yield and yield stability are a great challenge. A polygenic character of complex traits in the combination with the inter locus interaction makes this task hard and complicated. Furthermore, besides some general objects related to biology and the structure that are known, there are series of unknown magnitudes such as: the number of loci that control the trait; the number of segregating alleles per locus; allele frequency; effects of allelic substitution; linkage of loci; epistatic interactions between loci; gene expression and regulation. Therefore, quantitative traits are still a great challenge and difficult task in studies. Application of quantitative trait loci (QTL) methodology opens a discussion on a genetic basis of stability. VIA et al., 1995, described two types of genetic control. The “allelic sensitivity model” suggests that the constitutive gene itself regulates a direct response to environmental conditions, while the “gene regulation model” suggests that there are one or more regulatory genes under the direct influence of the environment and that the constitutive gene is activated or deactivated by these regulatory genes.
The main objective of the present study was to reveal the link between certain bands from genetic characterisation performed by the RAPD markers of observed maize hybrids and its parent components and quantitative traits, grain yield and yield stability. Yield stability was defined by the value of the interaction vector obtained in the АММI analysis.
MATERIALS AND METHODS
The study encompassed 15 commercial maize hybrids ranging from medium late maturity (FAO 400) to late maturity (FAO 700) (Table 1). The origin of parental lines and the affiliation with heterotic groups are presented in Tables 2.
Two-year four-replicate trials were set up according to the randomised block design in six locations and two sowing densities (D1=54,000 plants ha-1 and D2=64,900 plants ha-1) (24 environments). The grain yield (kg ha-1) was observed. According to data obtained on the yield,
the AMMI analysis was done and by this analysis the level of the interaction effect of observed maize hybrids was identified.
Table 1. Formulae and average yields of observed maize hybrids
Hybrid Hybrid formula Average yield (t ha-1)
ZP-677 (ZPL13 x ZPL14) x ZPL3 10.170 ZP-580 ZPL9 x ZPL10 10.002 ZP-735 ZPL11 x ZPL17 9.994 ZP-704 (ZPL13 x ZPL14) x ZPL2 9.879 ZP-753 ZPL15 x ZPL13 9.827 ZP-732 (ZPL13 x ZPL14) x ZPL16 9.692 ZP-701 ZPL13 x ZPL5 9.559 ST-500 NKL2 x LN2 9.423 ZP-633 ZPL11 x ZPL12 9.357 ZP-570 (ZPL4 x ZPL7) x ZPL8 9.314 ST-600 NKL1 x LN1 9.256 ZP-599 ZPL6 x ZPL2 9.250 ZP-480 ZPKL1 x ZPL4 8.802 ZP-533 (ZPL3 x ZPL5) x ZPL4 8.712 ZP-42а ZPL1 x ZPL2 8.663 Overall mean 9.460
Genetic characterisation of F1 hybrids and their parental components was done by the application of the RAPD markers. The DNA was isolated from grain following the modified protocol of SAGHAI – MAROOF et al., (1984). Depending on the DNA concentration after dilution of samples, random amplified polymorphic DNA polymerase chain reaction (PCR) was applied. The reaction was done following the protocol of WILLIAMS et al., (1990). Twenty-eight commercial arbitrary RAPD primers were used. The presence (1) and the absence (0) of bands were visually determined for observed maize genotypes.
According to information on yield, the observed maize hybrids were classified into two groups: with the yield above ("+ yield hybrids") and with the yield below overall mean ("- yield hybrids"). Based on the length of the interaction vector in the AMMI analysis hybrids were classified into three groups: within one ("stable hybrids"), two and three ("unstable hybrids") standard deviation. The percentage presence of all obtained bands from the RAPD analysis was calculated in all hybrids, and then bands that were present by more than 50% in "+yield" and "+stable" hybrids were separated. The presence of separated bands was also presented in parental components.
Inbred Origin Heterotic group
ZPL1 Version of the inbred B14 *BSSS
ZPL2 Мо17 Lancaster
ZPKL1 Sister combination derived by a cross of inbred line Lancaster and the inbred derived by pedigree selection from the cross of the Lancaster inbred and an inbred originating from Pećki dent
Lancaster
NKL2 Sister combination derived by a cross of inbreds (B84xCM109)xB84. CM109 is one of the В14 versions.
BSSS
LN2 Version of the inbred Мо17 Lancaster
ZPL3 Developed by pedigree selection from the cross of the inbred Мо17 and the inbred originating from Pećki dent
It combines with BSSS inbreds but it gives heterosis and it combines with Lancaster ZPL4 Developed by pedigree selection from the cross of the two
inbreds originating from Iowa
Independent source
ZPL5 Virginia inbred Lancaster
ZPL6 Developed by the cross of the inbred А632 (BSSS ) to the inbred А619 originating from Ohio
Results in hybrids with
Lancaster inbreds ZPL7 Developed by pedigree selection from the cross of the two
BSSS inbreds
BSSS
ZPL8 Version of the inbred L75 Lancaster
ZPL9 Originates from Istrian large-seeded population It combines with Lancaster
inbreds but results in hybrids also with inbreds from the independent source
ZPL10 Derived by hybrid self-pollination Independent source
ZPL11 Public inbred FR21-28 derived from self-pollination of a Pioneer hybrid
It combines best with Lancaster but results in good hybrids also with BSSS inbreds
ZPL12 Developed by pedigree selection from the Argentinean population
It combines with BSSS inbreds but also expresses heterosis with Lancaster
ZPL13 В73 BSSS
ZPL14 Version of the inbred В84 BSSS
ZPL15 Developed by pedigree selection from the cross (hybrid Arizona хС103) хМо17
Lancaster
ZPL16 Version of С103 Lancaster
ZPL17 Developed by backcrosses of a local inbred and an early version of Мо 17
Lancaster
NKL1 Developed by the cross of В73 to В84 BSSS
LN1 Version of the inbred Мо17 Lancaster
*
(BSSS-Iowa Stiff Stalk Synthetic)
RESULTS AND DISCUSSION
The AMMI model is a hybrid model that in the first step applies the analysis of variance, by which total variability is divided into main additive effects of genotypes and environments and additive interaction. Then the principal component analysis (PCA) is applied to the non-additive remain, GxE interaction. In such a way, several interaction PCA axes (IPCA) or components are also obtained. The essential question in all models that include the Singular
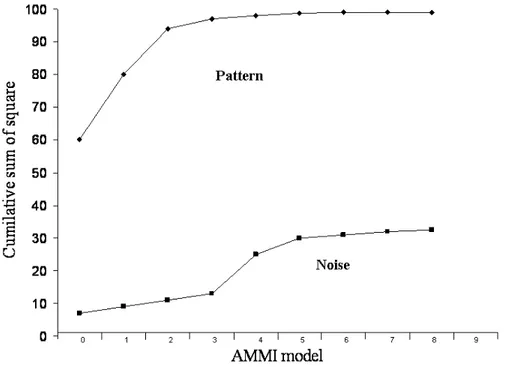
Value Decomposition (SVD) method is how many axes will be included into the model. Since the greatest amount of noise is placed in the interaction, the cumulative curves of the sum of squares of pattern and noise can facilitate in making the decision which number of axes is the best to be kept in the analysis in a concrete case. Figure 1 shows that the participation of pattern sharply increase when the first two interaction axes are included into the model. On the other hand, the participation of the noise gradually increased when the third interaction axis is included into the model and later it sharply increase (Figure 1). This led to a conclusion that for the given set of data the best result would be achieved when the first two IPCA axes are considered (the more detailed presentation of the results on the AMMI analysis can be seen in the manuscript BABIĆ et al., 2010).
Figure 1. Pattern and noise recovered by different AMMI models for grain yield of observed maize hybrids
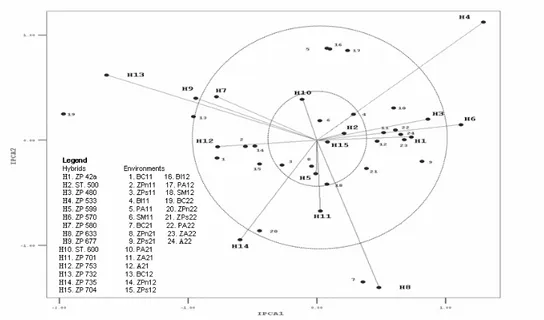
The AMMI2 biplot shows hybrids and environments according to values of the interaction effect (IРСА1-IРСА2). Concentric circles represent regions of one, two, or over two standard deviations of interaction vector length of observed maize hybrids. The lowest interactions were recorded in the hybrids ZP-599 (Н5), ZP-704 (Н15), ST-500 (Н2) and ST-600 (Н10), which were placed within a region of one standard deviation of interaction vectors. The following hybrids were placed within the region of two standard deviations: ZP-42а, 480, 580, ZP-677, ZP-701 and ZP-753. The highest values of interaction vectors (outside of two standard
deviations) were detected in the hybrids: ZP-533, ZP-570, ZP-633, ZP-735 and ZP-732, which points out the lowest stability of this group (Figure 2).
Figure 2. AMMI2 biplot for grain yield of the observed maize hybrids
The application of molecular markers in the improvement of complex quantitative traits, such as yield or stability, is very limited in the breeding programmes in which phenomenon of heterosis is used. The problem of the GxE interaction in commercial breeding programmes is solved by studying commercial hybrids in a great number of locations and years. Due to an unknown mechanism of genetic control and an unknown mechanism of gene regulation by environmental factors, the GxE interaction is not a simple problem within a field of molecular marker studies. On the other hand, we assumed that molecular markers could provide a different insight into the GxE interaction, the insight which was not possible by applied statistical analyses. Therefore, genetic characterisation by RAPD markers was done for both, hybrids and their parental components, with the aim to determine the link between results on the level and stability of yields obtained in field trials and results of genetic characterisation. The analysis was performed with 28 RAPD primers out of which primes ОРВ05, ОРВ09 and ОРВ02 did not provide bands; primers GEN4-70-7 and GEN1-70-9 did not provide data for a greater number of genotypes, while the primer ОРВ19 had one band for all observed genotypes. Therefore, all these primers were rejected. Twenty one primers were used for the further analysis. The total number of bands with 21 selected markers amounted to 141 that is 6.7 bands per primer.
According to field trials and presented statistical analyses it was tried in a simple way to find out the connection between the level and stability of yield and results obtained by the application of molecular markers. Observed maize hybrids were grouped according to obtained
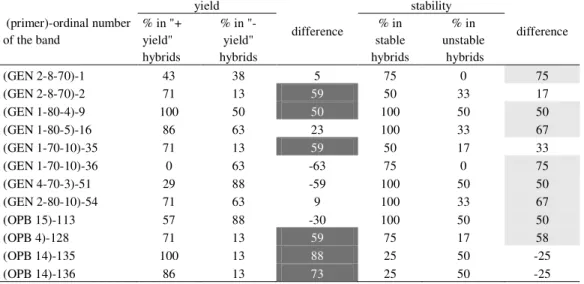
yield into high yielding "+ yield" (whose yield was above the overall mean) and low yielding "- yield" hybrids (whose yield was below the overall mean), and according to stability into "stable" (whose interaction was within one standard deviations of the interaction vectors) and "unstable" (whose interaction was outside values of two standard deviations of the interaction vectors). Then, the percentage of presence of each of 141 bands obtained by RAPD markers was estimated. Six bands (2, 9, 35, 128, 135 and 136) were separated as they were present by more than 50% in hybrids achieving yield above the average. Also, eight bands (1, 9, 16, 36, 51, 54, 113 and 128) were separated as they were present by more than 50% in hybrids that expressed small values of the interaction (Table 3). It was also noticeable that except bands 9 and 128 all others were different. This can lead to a conclusion that the yield and stability are the frequently defined by a different set of genes, i.e. by DNA fragments of different lengths and locations on a genome. This is in agreement with studies carried out by some authors who stated that the yield and stability were the most probably defined by different genes and, despite negative correlation, stable genotypes with high yielding potential can be developed. Moreover, loci in the interaction with the environment, so called stability loci, were identified in some recent studies dealing with QTL mapping of complex quantitative traits. Some of such loci were in the vicinity of the locus regulating the main trait, while others were positioned in sites with no QTLs regulating the main trait (UNGERER et al., 2003; SARANGA et al., 2001; HITTALMANI et al., 2003).
Table 3. Presence of "+ yield" (deep grey cells) and "+ stability" (light grey cells) bands in hybrids (%)
(primer)-ordinal number of the band yield difference stability difference % in "+ yield" hybrids % in "- yield" hybrids % in stable hybrids % in unstable hybrids (GEN 2-8-70)-1 43 38 5 75 0 75 (GEN 2-8-70)-2 71 13 59 50 33 17 (GEN 1-80-4)-9 100 50 50 100 50 50 (GEN 1-80-5)-16 86 63 23 100 33 67 (GEN 1-70-10)-35 71 13 59 50 17 33 (GEN 1-70-10)-36 0 63 -63 75 0 75 (GEN 4-70-3)-51 29 88 -59 100 50 50 (GEN 2-80-10)-54 71 63 9 100 33 67 (OPB 15)-113 57 88 -30 100 50 50 (OPB 4)-128 71 13 59 75 17 58 (OPB 14)-135 100 13 88 25 50 -25 (OPB 14)-136 86 13 73 25 50 -25
The presence of so called "+ yield" bands (deep grey) and "+stability" bands (light grey) is also presented for parental components (Table 4). It is noticeable that the "+ yield" bands were more present in inbreds of the ВЅЅЅ heterotic groups, while "+stability" bands were more often present in the inbreds of the Lancaster background. The most stable hybrids have most often a high presence of "+stability" bands in both parents. A great success of hybrids developed by the crosses of these two heterotic groups might be explained by this result. If we consider the known heterotic pair В73 and Мо17 (ZPL2 and ZPL13) we shall notice that Мо17 has all separated
"+stability" bands and one "+ yield" band, while В73 has all "+ yield" bands and few "+stability" band. According to practice the combination of these two inbred lines results in the stable high yielding hybrid whose modifications are still cultivated in large regions under the moderate climate conditions.
Table 4. Presence of "+ yield" (deep grey cells) and "+ stability" (light grey cells) bands in parental components marker/ genotype 1 2 9 16 35 36 51 54 113 128 135 136 NKL2 1 0 1 0 0 0 0 0 0 0 1 1 LN2 1 0 0 0 0 1 1 1 1 0 1 0 ZPL6 1 0 / 0 / / 0 1 1 0 0 1 ZPL2 1 0 0 1 0 1 1 1 1 0 1 0 ZPL13xZPL14 1 1 1 1 1 0 0 1 0 1 1 1 ZPL13 1 1 1 1 1 0 1 1 0 1 1 1 ZPL14 1 0 1 1 0 1 0 1 0 1 0 1 NKL1 1 1 1 0 1 0 0 1 0 1 1 1 LN1 1 0 0 1 0 1 1 1 1 1 0 0 ZPL4xZPL7 1 0 1 0 0 0 1 1 0 0 1 1 ZPL4 1 0 0 1 0 0 1 1 0 0 1 1 ZPL7 1 0 1 0 0 0 1 1 0 0 0 1 ZPL8 1 0 0 0 0 1 1 1 1 0 0 0 ZPL3xZPL5 1 0 0 0 0 1 1 1 1 0 0 0 ZPL3 1 0 0 0 0 1 1 1 1 0 0 0 ZPL5 1 0 0 0 0 1 0 1 1 0 / / ZPL11 1 0 0 0 0 1 1 1 0 0 0 0 ZPL12 1 0 0 0 0 1 1 1 1 0 0 0 ZPL16 0 0 0 1 / / 1 1 1 0 0 0 ZPL17 1 0 0 0 0 0 0 1 0 0 1 1 ZPL1 1 0 1 0 0 1 0 1 0 0 1 1 KL1 1 0 0 1 0 1 1 1 1 0 0 0 ZPL15 0 0 0 1 0 1 1 1 1 0 0 0 ZPL9 1 0 0 0 0 0 0 1 1 1 1 0 ZPL10 1 0 1 1 1 0 1 1 0 1 0 1
Two "+ yield" bands were present in the male component (ZPL4) of the hybrid ZP-533, while there were no "+ yield" bands present in the female component (ZPL3хZPL5) that had only "+stability" bands. This hybrid ranked last based on the yield, and its stability vector was in the region outside two standard deviations. The female component of the hybrid ZP-704 (ZPL13xZPL14) had all bands for the yield, as well as, five bands for stability, while the male component had six bands for stability and one for the yield. This hybrid had the lowest value of the interaction vector, while its yield was above the overall mean (it ranked fourth). For the
observed maize hybrids can be stated that parents of the ВЅЅЅ background contributed more to the yield, while the parents belonging to the Lancaster heterotic group contributed more to stability. However, such presence of bands can be simply a consequence of a great divergence of parental components. Due to this, as wall as, due to a small number of observed genotypes, general conclusions can not be drawn, but obtained results are to a great extent in accordance with results on field trials, as well as, with empirical data in practice (BABIĆ et al., 2011b).
When genetic potential of the yield is studied and genotypes are compared by the yield it is very rare to simultaneously observe parameters that can be a cause of stability and any biotic or abiotic stress. Therefore, when we consider yield stability of a certain genotype, we have to bear in mind that stability can be a consequence of various factors, such as tolerance to drought or resistance to the most important diseases or pests (BABIĆ et al., 2011a). Hence, genotype stability, determined in one set of experiments, cannot be generalised. It primarily presents certain stability of a genotype to a prevalent stress factor in the moment of carrying out the experiment.
The comprehension of the GхE interaction of quantitative traits from the genetic aspect requires knowledge on its genetic structure. Therefore, the GхE interaction will not be a simple problem from a molecular markers aspect, except in cases when the regulation of the gene activity by environments is simple. However, regardless of all difficulties that accompany the application of molecular markers in breeding for complex quantitative traits, such as yield and stability, their application in breeding programmes can provide a different insight into the nature of these traits, the insight that statistical analyses by themselves do not provide.
ACKNOWLEDGEMENT
Part of this work was supported by the Ministry of Education, Science and Technological Development of the Republic of Serbia through the Project TR 31068. Authors would like to thank the Ministry for the support.
Received March 20th , 2013 Accepted July 10th
, 2013
REFERENCES
ALLARD, R.W. (1997): Genetic basis of the evolution of adaptedness in plants. In: P.M.A. Tigerstedt, Adaptation in Plant Breeding,. Selected papers from the XIV EUCARPIA Congress on Adaptation in Plant Breeding, Jyvaskyla, Sweden, pp.1-12.
BABIĆ, V., M. BABIĆ, M. IVANOVIĆ, M. KRALJEVIĆ-BALALIĆ andM. DIMITRIJEVIĆ(2010): Understanding and utilization of genotype-by-environment interaction in maize breeding. Genetika, 42 (1): 303-312.
BABIĆ, M., V. BABIĆ, N. DELIĆ, S. PRODANOVIĆ and V. ANĐELKOVIĆ (2011a): The comparison of stability parameters according to the Finlay-Wilkinson, Еberhart-Russell and AMMI model. Selekcija i semenarstvo, Vol XVII, 2: 35-40.
BABIĆ, V., S. PRODANOVIĆ, M. BABIĆ, N. DELIĆand V. ANDJELKOVIĆ (2011b): The identification of bands related to yields and stability in maize hybrids. XXII EUCARPIA Maize and Sorghum Conference. Resources in Maize and Sorghum Breeding. Opatija, Croatia, June 19-22. Conference Book, pp. 94.
BECKER, H.C. (1981): Corelation among some statistical measures of phenotypic stability. Euphytica, 30: 835-840. CATTIVELLI, L., P. BALDI, C. CROSSATI, M. GROSSI and G. VALE (2002): Genetic bases of barely physiological response to
stressful conditions. In: Slafer G.A., Molina–Cano J.L, Savin R., Araus J.L., Romagosa I., Barely Sience, The Haworth Press, Binghamton, NY. USA. p. 307-360.
DIMITRIJEVIĆ, M. and S. PETROVIĆ (2000): Adaptability and stability of genotypes. Selekcija i semenarstvo VII (1-2): 21-28.
GAUCH, H.G.JR. (2006): Statistical Analysis of Yield Trials by AMMI and GGE. Crop. Sci. 46: 1488-1500.
GUNJAČA, J. (2001): Interakcija genotip x okolina u nebalansiranim serijama pokusa. Doktorska teza , Sveučilište u Zagrebu.
HITTALMANI, S., N. HUANG, B. COURTOIS and H.E. SHASHIDHAR (2003): Identification of QTL for growth-and grain yield-related traits in rice across nine locations of Asia. Theor. Appl. Genet. 107: 679-690.
LAMKEY, K.R. and M. LEE (1993): Quantitative genetics, molecular markers and plant improvement. „Proc. 10th Australian plant breeding conf.“, pp. 104-115 Gold coast, 18-23 April..
LEE, E.A., T.K. DOERKSEN and L.W. KANNENBERG (2003): Genetic components of yield stability in maize breeding populations. Crop Sci. 43: 2018-2027
SAGHAI-MAROOF, M., A. SOLIMAN, R.A. JORGENSEN and R.W. ALLARD (1984): Rihosomal DNA spacer-length polymorphisms in barely: mendelian inheritance, chromosomal location and population dynamics. Proc. Natl. Acad. Sci., USA, 81: 8014-8018.
SARANGA, Y., M. MENZ, C.X. JIANG, R.J. WRIGHT and D. YAKIR (2001): Genomic dissection of genotype x environment interactions conferring adaptation of cotton to arid conditions. Genome Res., 11: 1988-1995.
STOJAKOVIĆ, M., M. IVANOVIĆ, G. BEKAVAC, A. NASTASIĆ, B. PURAR, B. MITROVIĆ and D. STANISAVLJEVIĆ (2012): Evaluation of new NS maize hybrids using biplot analysis. Genetika, Vol. 44, No. 1: 1-12.
UNGERER, M.C., S.S. HALLDORSDOTTIR, M.D. PURUGGANAN and T.F.C. MACKAY (2003): Genotype-environment interactions at quantitative trait loci affecting inflorescence development in Arabidopsis thaliana. Genetics 165: 353-365.
VIA, S., R. GOMULKIEWICZ, G. DE JONG, S.M. SCHEINER and C.D. SCHLICHTING (1995): Adaptive phenotypic plasticity; consensus and controversy. Trends Ecol. Evol. 10: 212-217.
WILLIAMS, W.A., M.B. JONES and M.W. DEMMENT (1990): A concise table for path analysis statistics. Agron. J. 82: 1022-1024.
IDENTIFIKOVANJE TRAKA VEZANIH ZA PRINOS I STABILNOST HIBRIDA KUKURUZA I NJIHOVIH RODITELJSKIH KOMPONENTI
Vojka BABIĆ1, Slaven PRODANOVIĆ2, Milosav BABIĆ1, Nenad DELIĆ1, Violeta ANĐELKOVIĆ1
1
Institut za kukuruz, Zemun Polje, Beograd, Srbija
2
Poljoprivredni fakultet, Univerzitet Beograd, Beograd, Srbija Izvod
Gentičari pokušavaju da objasne adaptabilnost i stabilnost genotipa u smislu poželjne kombinacije alela ispoljenih kroz epistazu. Međutim, stabinost definisana u jednom setu podataka je vrlo često zapravo stabilnost genotipa na dominantni stresni faktor u momentu izvođenja ogleda. Stoga, stabilnost rodnosti određenog genotipa može biti rezultat različitih faktora kao što su tolerantnost na sušu ili neku značajnu bolest ili štetočinu. U ovom radu ispitivan je nivo i stabilnost prinosa 15 hibrida kukuruza u 24 poljne sredine. Nivo interakcijskog efekta definisan je AMMI analizom. Genetička karakterizacija je urađena RAPD markerima. Trake pozitivno vezane za visok nivo prinosa su najčešće bile prisutne u roditeljima koji vode poreklo iz BSSS populacije dok su trake pozitivno vezane za stabilnost bile u značajnom procentu prisutne kod roditelja Lancaster populacije.
Primljeno 20. III 2013.