KULTUR AKAR TRANSGENIK
Trichosanthes cucumerha
L.
SERTA PENGARUH DENSITAS EKSPLAN AWAL, UMUR
PANEN DAN KASEIN HIDROLISAT TERHADAP PRODUKSI
BIOMASSA DAN PROTEIN TOTAL AKAR TRANSGENIK
AbstrakPenelitian dilakukan di Laboratorium Kultur Jaringan Tanaman, Jurusan Budidaya Pertanian, Fakultas Pertanian IPB. Tujuan penelitian adalah untuk mendapatkan kultur akar transgenik (akar berambut) yang tumbuh stabil dalam media tanpa penarnbahan zat pengatur tumbuh dari kecambah Trichosanthes cucumerina L. dengan bantuan Agrobacterium rhizogenes 9457, mempelajari pengaruh densitas eksplan awal, umur panen serta kasein hldrolisat terhadap produksi biomassa dan hasil protein total dari akar transgenik.
Penelitian tediri dari tiga tahap yaitu inisiasi kultur akar berambut, uji pengaruh densitas eksplan dan umur panen terhadap produksi biomassa dan hasil protein total dari akar berambut, dan uji pengaruh penarnbahan kasein hidrolisat terhadap produksi biomassa dan hasil protein total dari akar berambut.
Hasil infeksi kecambah in vitro dengan A. rhizogenes menunjukkan sebanyak 42 kecambah dari selutar 108 kecambah yang diinfeksi menghasilkan akar pada lokasi infeksi. Namun setelah melalui tahap penapisan dalarn media Murashige-Skoog tanpa zat pengatur tumbuh (MS-0), hanya dihasilkan satu galur akar berarnbut yang tumbuh stabil dalam media tersebut. Galur akar berambut tersebut tumbuh stabil dalam jurnlah sub kultur yang ti& terbatas dalam media MS-0 dan menunjukkan indikasi yang kuat sebagai akar transgenik. Kesulitan mengeliminasi sisa-sisa Agrobacterium dari media kultur menyebabkan galur akar berambut yang dihasilkan sangat terbatas.
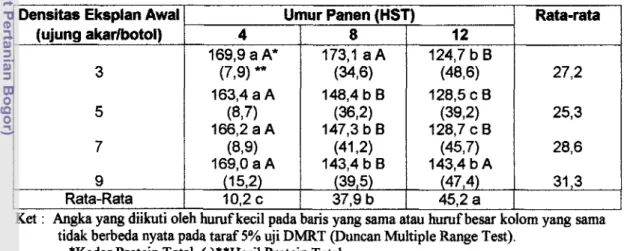
Galur akar berambut yang tumbuh stabil digunakan dalam uji pengaruh densitas eksplan awal, umur panen dan kasein hidrolisat. Densitas eksplan awal3, 5, 7 dan 9 akar per botol dikombinasikan dengan umur panen 4, 8 dan 12 HST. Biomassa yang dihasilkan pada semua perlakuan densitas eksplan awal pada 12 HST, berbeda nyata antara eksplan awal3 dengan yang lainnya. Sebaliknya hasil protein total cenderung menurun dengan meningkatnya umur panen pada setiap densitas eksplan awal. Perbedaan densitas eksplan mempengaruhi pola perturnbuhan karena pada densitas eksplan 7 dan 9 akar berarnbut cenderung lebih cepat memasuki fase stasioner. Kadar protein total menurun dengan meningkatnya umur kultur sedangkan hasil protein total tergantung pada waktu panen.
Kasein hidrolisat tidak berpengaruh nyata terhadap produksi bomassa, namun cenderung menekan produksi biomassa dibanding kontrol. Kadar protein total dapat ditingkatkan pada konsentrasi kasein hldrolisat 50 mgll
.
Peningkatan kasein hidrolisat menjadi 100 mg/l menglaasilkan kadar protein yang sama dengan kasein hidrolisat 50 mgll. Peningkatan menjadi 150 mgA menyebabkan penurunan kadar protein total. Hasil protein total tertinggi dihasilkan pada media tanpa kasein hidrolisat.Pendahuluan
Kultur akar merupakan suatu bentuk kultur jaringan akar yang hidup dan berdiferensiasi secara terorganisir membentuk biomassa akar tanpa kehadran tipe organ lain dari tanaman seperti batang, daun ataupun bunga. Terdapat dua tipe kultur akar yaitu kultur akar non transgenik dan kultur akar transgenik. Kultur akar non transgenik diperoleh dengan memotong ujung akar tanaman di lapangan lalu disterilkan maupun akar kecambah tanarnan in vitro lalu ditanam dalam media kultur jaringan (Payne et al. 1992).
Kultur akar transgenik merupakan kultur akar yang dihasilkan dengan menginfeksi tanaman dengan Agrobacterium rhizogenes. Infeksi tanaman dengan
Agrobacterium rhizogenes akan mengakibatkan ditransfernya T-DNA dari Ri (Root inducing) plasmid ke genom tanaman yang menyebabkan induksi proliferasi akar transgenik yang juga disebut akar berambut (Payne et al., 1992).
Akar berambut mempunyai ciri percabangan lateral yang ekstensif dan tidak dipenganh geotropisme. Percabangan yang ekstensif tersebut menghasilkan banyak meristem sehingga memungkinkan pertumbuhan biamassa yang cepat dibandingkan akar tanaman normal. Sifat penting lainnya dari akar berambut adalah kemampuannya untuk tumbuh stabil dalam media kultur in vitro walaupun tanpa penambahan zat pengatur tumbuh.
Pada kultur akar bukan transgenik, pemantapan dan pemeliharaannya memerlukan tambahan auksin yang dapat menekan produktivitasnya dalam menghasilkan metabolit sekunder. Kultur akar berambut cenderung lebih stabil dibandingkan kultur kalus dan suspensi sel dan merupakan alternatif yang lebih
baik dibandingkan ekstraksi protein langsung dari tanaman in vivo maupun Mtur kalus dan suspensi sel (Toppi et al. 1996).
Induksi akar berambut dengan bantuan Agrobacterium rhizogenes telah berhasil dilakukan pada beberapa spesies Cucurbitaceae. Diantaranya adalah
Lufla cylrndrica L. ( Toppi, 1996; Turnilisar 2001), Cucurbita pep0 L. (Toppi, 1997), dan Trichosanthes spp.(Savary dan Flores, 1994). Akar berambut pada beberapa spesies Cucurbitaceae tersebut digunakan mtuk mempelajari identitas protein aktif yang ada pada akar tanaman.
Protein aktif dari berbagai tanaman dilaporkan bermanfaat bagi tamman penghasilnya sebagai suatu mekanisme ketahanan terhadap serangan patogen maupun cekarnan abiotik. Protein aktif tersebut dapat berupa enzim, defensin,
antimicrobial peptide, maupun Ribosome Inactivating Proteins (RIP). Kultur akar
berambut dari Trichosanthes sp. dilaporkan menghasilkan trichosantin dan
chtinase yang menunjukkan aktivitas anticendawan dalam uji in vitro.
Vivanco et al. (1997) menyatakan bahwa protein aktif berupa Ribosome
In-activating Protein (RIPS) beptensi besar untuk dimanfmtkan dalam bidang pertanian. Pendapat ini didukung oleh adanya peningkatan ketahanan terhadap cendawan pada tanaman tembakau transgenik yang mengekspresikan RIP dari biji barley (Logeman et al. 1992).
Efisiensi penggunaan kultur akar berambut untuk produksi metabolit sekunder sangat tergantung pada kemampuan produksi biomassa dan kadar senyawa target dari akar berambut. Dua ha1 tersebut kadang-kadang tidak terjadi secara bersamaan. Produksi biomassa yang tinggi belwn tentu diiringi oleh kadar senyawa target yang tinggi. Usaha mengoptimalkan produksi biomassa maupun
kadar senyawa target kadang-kadang perlu dilakukan. Perbaikan produktivitas biomassa dapat dilakukan dengan penapisan dan seleksi galur akar berambut, optimasi komposisi media dan induksi produksi senyawa yang diinginkan dengan perlakuan elisitasi (Fu, 1999).
Densitas eksplan awal merupakan faktor penting yang berpengaruh terhadap kapasitas produksi biomassa akar berambut per satuan waktu tertentu. Laju proliferasi jaringan dalam kultur juga dipengaruhi oleh rasio eksplan dengan medium, rasio eksplan dengan udara, volume botol kultur, dan tipe penutup botol kultur, disamping faktor-faktor utama seperti nutrisi media (McClelland
dan
Smith, 1990)Optimasi komposisi media dapat dilakukan antara lain dengan penambahan senyawa organik maupun anorganik. Penarnbahan berbagai bentuk senyawa yang mengandung nitrogen fteroksidasi atau tereduksi, organik ataupun anorganik) diketahui dapat berpengaruh nyata terhadap pertumbuhan dan diferensiasi eksplan yang ditanam secara m vitro (Narayanaswarny, 1994).
Kasein hidrolisat merupakan senyawa organik kompleks yang mengandung berbagai asam amino. Asam amino berfimgsi sebagai sumber N dan berpengaruh terhadap aktivitas nitrat reduktase, pertumbuhan kultur, serta biosintesis produk sekunder. Berbagai senyawa organik sumber N juga mempunyai pengaruh yang kompleks dan kemunglunan berkaitan dengan peranannya sebagai sumber karbon, prekursor produk clan molekul regulator (Cresswell et al. 1989).
Percobaan yang dilakukan bertujuan untuk untuk mengnduksi akar berarnbut pada hipokotil kecambah paria belut (Trichosanthes cucumerina L.)
dengan bantuan Agrobacterium rhizogenes strain 9457, menyeleksi galur akar berambut yang hunbuh stabil dalarn media tanpa zat pengatur tumbuh dan mengevaluasi pengaruh densitas eksplan dan waktu panen serta penambahan kasein hidrolisat terhadap produksi biomassa, k& protein dan hasil total protein dari akar berambut.
Bahan dan Metode Percobaan
Percobaan dilakukan dalam tiga tahap, yaitu inisiasi kultur akar berambut, uji pengaruh densitas eksplan dan waktu panen, dan uji pengaruh penambahan kasein hidrolisat terhadap produksi biomassa, kadar protein
dan
hasil protein total dari akar berambut Trichosanthes cucumerina L.Eksplan dan Media Tanam
Eksplan awal berupa benih tanaman Trichosanthes cucumerina L. yang diperoleh dari daerah Weleri, Jawa Tengah, dicuci bersih dan dibuang kulit bijinya. Kemudian benih direndarn dalam bakterisida Agrep dan k g i s i d a Dithane selama 24 jam untuk mengurangi kontaminasi cendawan. Selanjutnya benih disterilkan dengan larutan sodium hipoklorit (Baycline 20%, 15% dan 10%) berturut-turut selama 20, 15 dan 10 menit, dibilas dengan air steril lalu Qtanarn dalarn media dasar Murashige-Skoog (MS) yang dtambah gula (30 g/l) dan agar (7 gll). Setelah benih berkecambah, pada umur 2 minggu setelah perkecambahan bagian hipokotil diinfeksi dengan Agrobacterium rhizogenes.
Inisiasi Kultur dan Penapisan Galur Akar Berambut
Agrobacterium rhizogenes strain 9457 disegarkan dalam media Yeast Manitol Broth
(YMB)
padat dengan komposisi yeast extract ( 0.4 gll), mannitol(10 gh), NaCl (0.1 gh), MgS04. 7 H20 (0.2 g/l) dan KH2P04 (0.5 gll), dan agar (7 gA). pH media diatur sehingga mencapai 7.0 sebelum diautoklaf pada suhu 12 1 OC, tekanan 1.5 psi selama 20 menit. Koloni berumur 3 hari digunakan untuk menginokulasi hipokotil kecambah in vitro yang berumur 2 minggu. Infeksi dilakukan dengan melukai bagian hipokotil kecambah dengan jarum preparat yang telah dicelupkan ke koloni Agrobacterium rhizogenes 9457.
Pertumbuhan dan perkembangan kecambah yang diinokulasi dengan
Agrobacterium diamati selama 3 minggu setelah inokulasi. Akar berambut yang muncul dipindahkan ke media MS tanpa ZPT (MS-0) padat dengan penambahan antibiotika cefotaxime (250 mg). Akar disubkultur ke media yang masih segar sampai 4-5 kali dan dievaluasi pertumbuhannya. Akar yang dapat turnbuh baik &lam MS-0 padat selanjutnya ditanam dalam media MS-0 cair dan diinkubasikan dalam ruangan dengan suhu 2527°C dengan cahaya terang dan dikocok dengan kecepatan 100 rpm pada rotary shaker.
Pemeliharaan Kuitur Akar berambut
Pemeliharaan kultur akar berambut dilakukan untuk menyediakan sumber eksplan bagi beberapa percobaan yang akan dilakukan. Media tanam untuk pemeliharaan kultwr akar berambut adalah media MS-0, terdiri dari garam-gararn yang mengandung hara makro, hara mikro, dan ditarnbah vitamin serta bahan organik dengan komposisi seperti yang dikembangkan oleh Murashige clan Skoog (1962). Karbohidrat diberikan dalam bentuk gula sebanyak 30 g/l. Media dalarn bentuk cair dm pH media diatur sehingga mencapai 5.8 dengan menggunakan KOH (1 N) atau HCl (IN) sebelum diautoklaf pa& suhu 12 1°C, tekanan 1.5 psi selama 20 menit.
Kultur akar berambut dipelihara dalam media MS-0 seperti tersebut diatas dengan mensubkultur ujung-ujung akar berambut setiap 12-14 hari sekali. Kultur akar berambut diinkubasi dalam ruangan dengan suhu 25
- 27OC dengan cahaya
terang dan dikocok dengan kecepatan 100 rpm pada rotary shaker.Uji Pengaruh Densitas Eksplan Awal dan Umur Panen.
Akar berambut yang tumbuh stabil selama 15-20 kali sub kultur dalam media pemeliharaan MS-0 digunakan dalarn percobaan ini. Eksplan ujung akar sepanjang 1-1.5 cm ditanam dalam 25 ml media MS-0 dan dievaluasi perkembangannya. Dalam percobaan ini dievaluasi pengaruh berbagai densitas eksplan (3, 5, 7 dan 9 eksplan awal) dan umur panen 4, 8 dan 12 hari sesudah tanam (HST) terhadap produksi biomassa dan protein total yang dipanen. Setiap kombinasi perlakuan dalam percobaan ini diulang 4 kali dan tiap ulangan terdiri dan satu botol. Kultur diinkubasikan pada gyratory shaker inkubator dengan cahaya rendah dan dikocok dengan kecepatan 100 rprn.
Pengamatan dilakukan terhadap bobot segar, bobot kering akur berambut
setelah dikeringdinginkan dengan fieeze dryer dan kadar protein total akar berambut yang dilakukan pada masing-masing umur panen. Hasil protein total dihitung berdasarkan perkallan bobot kering akar berambut dengan kadar protein total.
Uji Pengaruh Kasein Hidrolisat
Dalam percobaan ini, diuji pertumbuhan dan perkembangan akar berambut dalam media MS-0 dengan penambahan kasein hidrolisat. Eksplan ujung akar (1- 1.5 cm, 3 eksplanhtol) ditanam dalarn 25 ml media MS-0 dengan penambahan
berbagai konsentrasi casein hydrolisate (0,50, 100 an 150 mg/l). Setiap perlakuan diulang 12 kali dengan satu botol sebagai satu ulangan. Kultur diinkubasikan pada
gyratory shaker inkubator dengan cahaya rendah clan dikocok dengan kecepatan 100 rpm.
Ekstraksi Protein dan Pengukuran Kadar Protein Total
Protein diekstmk dari akar berambut dengan menggerus contoh yang dianalisis (0.2 g bobot kering) dalam 10 ml buffer fosfat (NaH2P04 10 mM, Na2HP04 15 mM, KC1 100 mM, Na2EDTA 2 mM dan PVP 0.75%, pH 7.5) dingin dan disentrifugasi dengan kecepatan 8000 rpm selama 15 menit. Supernatan di ambil dan ke dalam endapan ditambahkan kernbali 10 rnl buffer fosfat dan disentrifugasi kembali pada suhu 4°C dengan kecepatan 8000 rpm selama 15 menit.. Supernatan yang didapat digabungkan dan ditera hingga 25 ml dengan penambahan buffer fosfat. Total protein terlarut dalam gabungan supernatan yang didapat ditentukan dengan metode Bradford (1976). Kadar protein total dan hasil protein total yang didapat dilutung berdasarkan bobot kering total biomassa dari contoh yang dianalisis.
Pegamatan dan Analisis Data
Pengamatan dilakukan terhadap bobot segar, bobot kering hairy root setelah dikeringdinman dan kadar protein total hairy root yang dipanen pada 12 HST. Hasil protein total dihitung berdasarkan perkalian bobot kering akar berambut dengan kadar protein total. Data hasil percobaan dianalisis sidik ragam dengan selang kepercayaan 95%. Jika perlakuan berpengaruh nyata dilakukan analisis lanjut dengan Duncan Multiple Range Test (DMRT).
Hasil Inisiasi Kultur Akar Berambut
Kecambah yang diinokulasi dengan Agrobacterium rhizogenes LBA 9457 menunjukkan gejala pembengkakan seperti membentuk kalus pada bagian yang diinfeksi pada 4-7 hari setelah infeksi (HSI). Respon pembentukan tumor pada jaringan kecambah berkisar antara 47% pada 7 HSI sarnpai dengan 56% pada 21 HSI (Lampiran Tabel 1).
Akar berambut biasanya terbentuk pada jaringan kalus yang berkembang pada bagian kecambah yang diinokulasi dengan Agrobacterium, meshpun ada yang berkembang pada jaringan tanpa pembentukan kalus. Morfologg akar rambut yang terbentuk bervariasi dar~ antara tebal, kaku dan pendek atau langsing dan memanjang (Gambar 2). Akar rambut juga berkembang dari bagian kecambah yang tidak diinokulasi dengan Agrobacterium, yang biasanya mempunyai morfologi akar rambut yang lebih kecil dan halus.
Contoh akar rambut yang diisolasi darr m a n yang tidak diinfeksi biasanya tidak berhasil tumbuh dalam media MS-0 padat dengan cefotaxime (250 mg/l) yang mengindikasikan akar tersebut bukan akar transgenik yang dicari. Sebaliknya akar yang muncul dari jaringan kalus biasanya mampu tumbuh dalam media MS-0 dengan cefotaxime (250 mg/l). Kesulitan dalam mematikan
Agrobacterrum dalam kultur akar menyebabkan keberhasilan mendapatkan kultur akar rambut transgenik menjadi rendah. Pada akhir tahapan percobaan ini hanya didapat satu klon galur akar berambut yang mampu tumbuh cepat secara terus menerus &lam media MS-0.
Gambar 2. Akar adventif yang muncul di lokasi infeksi dengan Agrobacterium rhizogenes. A. Akar adventif dengan morfologi yang lebih tebal dan gemuk, B. Akar adventif dengan morfologi yang lebih halus dan langsing. C. Akar berambut yang telah tumbuh stabil dalam media MS 0
+
3% sukrosa tanpa ZPT dengan kondisi cahaya terang terus menerus. D. Akar berambut yang telah tumbuh stabil dalam media MS 0 + 3%Kondisi lingkungan terutama cahaya cenderung berpengaruh terhadap morfologi akar berambut dalam satu galur. Hal tersebut ditemukan secara tidak sengaja ketika akar berambut diinkubasi &lam kondisi cahaya yang relatif berbeda. Ketika akar berarnbut diinkubasi &lam ruangan dengan cahaya terang terus-menerus, morfologi akar berambut menunjukkan percabangan yang intensif, pendek-pendek, tebal dan kaku, dan berwarna kehijauan. Sebaliknya ketika diinkubasi dalam shaker incubator dengan cahaya rendah, akar berambut tersebut mempunyai percabangan yang intensif yang cenderung tumbuh memanjang dan berwarna putih (Gambar 2).
Pengaruh Densitas Eksplan Awal dan Umur Panen
Dalam percobaan ini, akar berambut mulai berkembang 4 hari setelah tanam yang ditandai dengan terjadinya pemanjangan akar sekunder. Pada umur 8 hari akar sekunder makin memanjang
dan
membentuk percabangan tertier dan akar hampir memenuhi seluruh ruangan media. Pada 12 hari setelah tanam akar berambut telah memenuhi seluruh ruangan media.Hasil analisis data menunjukkan densitas eksplan awal atau umur panen berpengaruh nyata terhadap berbagai peubah yang diamati. Sedangkan persentase bahan kering hanya dipengardu oleh umur kultur.
Rata-rata bobot segar akar berambut nyata meningkat dengan meningkatnya densitas eksplan awal ataupun umur panen. Penggunaan 3 eksplan awal nyata memberikan bobot segar lebih rendah dibanding 7 atau 9 eksplan awal namun tidak berbeda nyata dengan bobot segar pada 5 eksplan awd. Bobot segar
secara nyata meningkat antara waktu panen 4 , 8 dan 12 HST seperti terlihat pada Tabel 1.
Persentase bahan kering tidak dipengaruhi oleh densitas eksplan awal, tetapi hanya dipengaruhi oleh umur panen. Umur panen 12 hari nyata mempunyai persentase bahan kering lebih tinggi dibandingkan umur panen 4 dan 8 HST (Tabel 1).
Tabel 1. Bobot segar akar rambut (g) dan persentase bahan kering (%) pada beberapa densitas eksplan awal dan umur panen
Pengaruh umur panen terhadap hasil bobot kering biomassa akar berambut Rata-rata Densitas Eksplan Awal
fuiuna akarlbotol) 3 5 7 9 Rata-Rata
tergantung densitas eksplan awal yang digunakan. Pada penggunaan 3 eksplan awal per botol, bobot kering biomassa nyata meningkat pada umur panen 4
Umur Panen (HST)
Ket : Angka yang diikuti oleh huruf yang sama pada baris atau kolom yang sama tidak berbeda nyata pada taraf uji 5% (DMRT).
*
Bobot segar **Persentase Bahan Kering0,61* (7.0)" 0,72 (6.5) 0,88 (6.6) 1,33 (6.1) 0,88 c (6.6B)
sampai dengan 12 HST. Sebaliknya pada penggunaan 5, 7 atau 9 eksplan awal 4
per botol, umur panen 8 dan 12 HST nyata menghasilkan bobot kering biomassa
8
I
12 2,46 (6.0) 3 , s (7.1) 4,64 (7.0) 4,11 (7.2) 3,77 b (6.88)lebih tinggi (Tabel 2).
Tabel 2. Bobot kering akar rambut (g) pada beberapa densitas eksplan awal dan umur panen
4,20 (8.3) 4,39(8.1) 5,11 (7.7) 5,07(7.1) 4,69 a (7.8A) 2,42 B (7.1) 2,99AB(7.2) 3,54A (7.1) 3,51A (6.8)
Densitas Eksplan Awal (ujung a k a r l W )
3 5 7 9
Ket : Angka yang diikuti oleh huruf kecil pada baris yang sama atau huruf besar kolom yang sama tidak berbeda nyata pada taraf uji 5% (DMRT).
Umur Panen (HST) 4 0,04cA 0,05 b A 0,06 b A 0,08 b A 8 0,14bB 0,27 a A 0,32 a A 0,30 a A 12 0,34aA 0,35 a A 0,38 a A 0,35 a A
Sebaliknya, pengaruh jumlah eksplan awal terhadap bobot kering biornassa akar berambut tergantung pada umur panen yang dilakukan. Pada umur panen 4 HST, jumlah eksplan awal yang digunakan tidak berpengaruh nyata dan biornassa kering masih relatif rendah. Dernikian pula pada umur panen 12 HST, jumlah eksplan awd tidak berpengaruh nyata, tetapi biomassa yang didapat relatif tinggi. Pada umur panen 8 HST, penggunaan 3 eksplan per botol nyata menghasilkan total biomassa kering yang lebih rendah dibandingkan jumlah eksplan yang lainnya (Tabel 2).
Tabel 3. Kadar protein total (mg/g BK) dan hasil protein total (mg) dari akar berambut pada beberapa densitas eksplan awal clan umur Panen
Kn : Angka yang diikuti o l k huruf kecil pads baris ya*g sama ahu humf besar kilom yang sama ' tidak berbeda nyata pada taraf 5% uii DMRT (Duncan Multivle Range Test).
Densitas Eksplan Awal (ujung akarlbotol) 3 5 7 9 Rata-Rata
Kadar protein total yang diisolasi dan akar rarnbut dipengaruhi oleh interaksi antara densitas eksplan awal yang digunakan dan umur panen. Pada penggunaan 3 eksplan awal per botol, kadar protein total nyata menurun pada umur panen 12 HST. Pada penggunaan eksplan awal 5,7 dan 9 per botol, kadar proteinnya telah nyata menurun pada urnur panen 8 HST (Tabel 3).
Pada umur panen 4 HST, jumlah eksplan awal tidak berpengaruh nyata terhadap kadar protein total. Pada umur panen 8 HST, penggunaan 3 eksplan awal Rata-rata 27,2 25,3 28,6 31,3 Umur Panen (HST) 4 169,9 a A* (7,9) " 163,4 a A (8,7) 166,2 a A (8,9) 169,O a A (1 5,2) 10.2 c 8 173,l a A (3416) 148,4 b B (362) 147,3 b B (41 $2) 143,4 b B (39,5) 37.9 b 12 124,7 b B (48,6) 128,5 c B (392) 128,7 c B (45,7) 143,4 b A (47,4) 45.2 a
memberikan kadar protein total tertinggi. Sebaliknya pada umur panen 12 HST, kadar protein total tertinggi didapat ketika penggunaan 9 eksplan awal per botol.
Hasil protein total hanya dipengaruhi oleh umur panen. Meningkatnya
umur panen nyata meningkatkan hasil protein total yang didapat dan rata-rata hasil protein total tertinggi diperoleh pada umur panen 12 HST (Tabel 3).
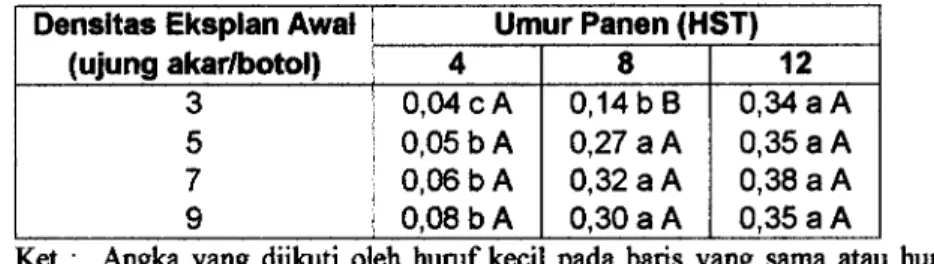
Morfologi akar berambut pada berbagai densitas eksplan awal dan urnur panen seperti terlihat pada Gambar 3, 4 dan 5. Setiap eksplan mempunyai kecepatan pertumbuhan dan perkembangan yang relatif sama seperti terlihat pada Gambar 3. Tetapi pada penggunaan 7 dan 9 eksplan awal per botol, biomassa akar telah memenuhi r u n g media pada 8 HST, sebaliknya pada penggunaan 3 dan 5 eksplan awal per botol, biomassa akar baru memenuhi ruang media pada 12 HST. Pengaruh Kasein Hidrolisat
Penarnbahan berbagai konsentrasi kasein hidrolisat tidak berpengaruh nyata terhadap rata-rata bobot segar, bobot kering, dan persen bahan kering akar rambut yang dihasilkan. Rata-rata bobot segar, bobot kering dan persentase bahan kering yang dihasilkan disajikan pada Tabel 4. Morfologi akar berambut pada berbagai konsentrasi kasein hidrolisat terlihat pada Gambar 6.
Tabel 4. Bobot segar, bobot kering dan persentase bobot kering akar berarnbut pada berbagai konsentrasi kasein hidrolisat
Kasein hidrolisat (mgll) dak berbeda 0 50 100 150 Bobot Segar (9)
Ket : Angka yang diikuti oleh huruf yang sama pada kolom yang sama t nyata pada taraf uji 5% (DMRT)
4,40 3,58 3,50 3,05 Bobot Kering (9) Persen Bahan Kering ( % ) 0,32 0,27 0,26 0,25 7,5 7,6 7,7 8.4
Tabel 5. Kadar protein total dan hasil protein total dari berambut pada beberapa konsentrasi kasein hidrolisat
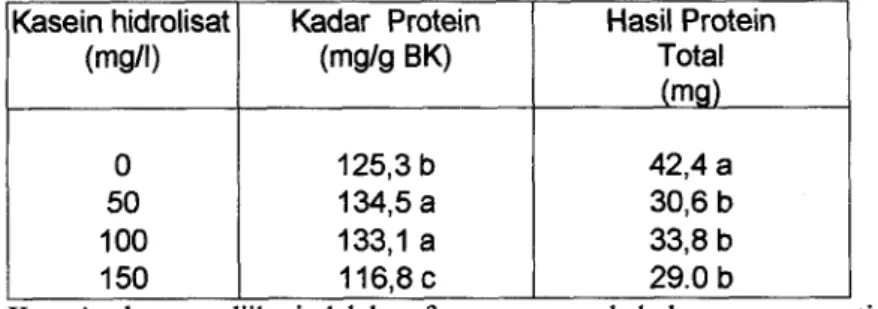
Kadar protein total nyata dipengaruhi oleh penambahan berbagai Hasil Protein Total (mg) Kasein hidrolisat (mg/l) 0 50 100 150
konsentrasi kasein hdrolisat Penambahan kasein hidrolisat sampai dengan 100 Kadar Protein
(mg/g BK)
mg/l nyata meningkatkan kadar protein total yang diisolasi dari akar berambut. Ket : Angka yang diikuti oleh huruf yang sama pada kolom yang sama tidak berbeda
nyata pada taraf uji 5% (DMRT) 125,3 b 134,5 a 133,l a 116,8 c
(Tabel 5). Pada penambahan kasein hdrolisat 150 mgll, kadar protein justru nyata 42,4 a
30,6 b 33,8 b 29.0 b
lebih rendah dibanding kontrol.
Hasil protein total nyata dipengaruhi penambahan kasein hidrolisat. Hasil protein total cenderung menurun dengan penambahan kasein hidrolisat. Hasil protein total tertinggi dihasilkan pada kontrol dan berbeda nyata dengan yang dihasilkan pada penarnbahan kasein hidrolisat 50, 100, maupun 150 mg/l (Tabel
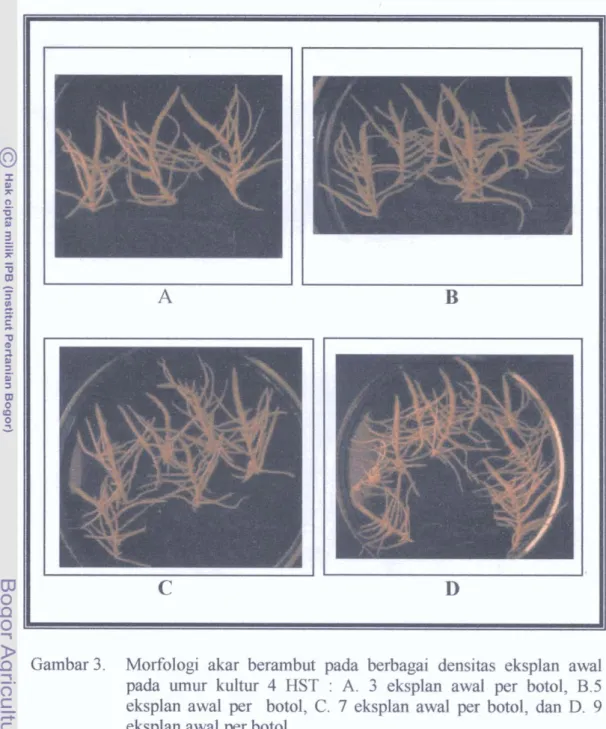
Gambar 3. Morfologi akar berambut pada berbagai densitas eksplan awal pada urnur kultur 4 HST : A. 3 eksplan awal per botol, B.5 eksplan awal per botol, C. 7 eksplan awal per botol, dan D. 9 eksplan awal per botol.
Gambar 4. Morfologi akar berambut pada berbagai densitas eksplan awal pada umur kultur 8 HST: A.3 eksplan awal per botol, B. 5 eksplan awal per botol), C. 7 eksplan awal per botol dan D. 9 eksplan awal per botol.
Gambar 5. Morfologi akar berambut pada berbagai densitas eksplan awal pada umur kultur 12 HST: A.3 eksplan awal per botol), B. 5 eksplan awal per botol), C. 7 eksplan awal per botol, dan D. 9 eksplan awal per botol.
Gambar6. Morfologi akar berambut hasil perlakuan kasein hidrolisat: A. kasein hidrolisat 0 mfl, B. kasein hidrolisat 50 mfl, C. kasein hidrolisat 100 mfl), D. kasein hidrolisat 150 mfl, Umur Kultur
Pem bahasan
Akar transgenik (akar berambut) dapat terbentuk sebagai ahbat integrasi T-DNA dari Ri-plasmid pada Agrobacterium rhizogenes ke dalam genom sel tanaman. T-DNA tersebut mengandung gen yang fungsinya mernungkinkan terjadinya proliferasi akar adventif dari sel transgenik.
Dalam percobaan ini gejala pembengkakan dalam bentuk turnorlgall telah terlihat pada 4-7 hari setelah infeksi dengan Agrobacterium. Pada beberapa spesies tanaman, akar akan muncul secara langsung pada tempat inokulasi bakteri, tetapi pada spesies tertentu mula-mula akan muncul kalus pada tempat inokulasi, selanjutnya dari kalus tersebut akan muncul akar. Dalam keadaan normal 1-4 minggu setelah inokulasi akan muncul akar (Ernawati, 1992)
Efisiensi pembentukan akar berambut berdasarkan jumlah tanaman yang diinfeksi yang membentuk akar berambut pada lokasi infeksi yaitu sekitar 43%. Hasil tersebut terlihat masih rendah dibandingkan efisiensi transformasi pada LuHa cyllindrica L. Roem yang mencapai 80% (Toppi et al. 1996) dan pada Cucurbita pep0 L. yang mencapai 94% dengan bantuan A. rhizogenes 1855 (Toppi et al., 1997).
Menurut Gelvin (2000), sejumlah tahapan dalam proses transformasi dapat membatasi transformasi pada tanaman tertentu. Tahapan tersebut meliputi sintesis fenolik oleh tanaman yang berperan dalam induksi gen vir, kolonisasi sel tanaman oleh Agrobacterium, transfer T-DNA ke dalam sitoplasma tanaman, translokasi T- DNA ke inti sel tanaman dan integrasi T-DNA.
Terbatasnya jumlah galur akar berambut yang diperoleh yang turnbuh stabil dalam media MS-0, yaitu hanya satu galur dari sekitar 43 galur awal, lebih
banyak disebabkan sulitnya menghilangkan kontaminasi oleh Agrobacterium. Shackelford dan Chlan (1996) menyatakan bahwa kontaminasi karena
Agrobacterium merupakan suatu masalah setelah proses transformasi. Agrobacterium dapat mengalami pertumbuhan yang berlebihan sehingga
menghambat pertumbuhan jaringan atau akar hasil transformasi.
Akar berambut yang tumbuh stabil dalam media tanpa penambahan zat pengatur tumbuh dalam jumlah sub kultur yang tidak terbatas merupakan indikasi yang kuat bahwa telah terjad transformasi pada sel tanaman. Akar berambut bersifat autotrof terhadap hormon karena ekspresi dari T-DNA yang telah terintegrasi dalarn sel tanaman. Hormon yang tinggi dalam sel tanaman yang memungkinkan sel berproliferasi dan berdiferensiasi secara cepat diduga karena adanya aksi dari berbagai roE gene pada T-DNA.
Perbedaan morfologi akar berambut pada dua kondisi lingkungan dengan intensitas cahaya yang berbeda sebagaimana digambarkan pada bagian hasil, menunjukkan bahwa pada akar berambut yang ditempatkan dibawah cahaya terang terus-menerus, aktivitas auksin yang berperan dalam pemanjangan sel kemungkinan lebih rendah. Akibatnya pemanjangan akar relatif terhambat. Sedangkan akar berambut yang ditempatkan pada shaker dengan cahaya rendah, kemungkinan aktivitas auksin lebih tinggi dan akar berambut mengalami etiolasi sehingga pemanjangan tidak terhambat.
Cahaya dapat menginduksi sel-sel untuk membentuk klorofil yang mengakibatkan warna kehjauan pada akar berambut. Hal ini juga dilaporkan oleh Savary dan Flores (1994) pada sebagian besar klon akar berambut dari
kelihatan berlangsung sampai 12 HST. Hal tersebut menunjukkan bahwa pada densitas eksplan awal yang lebih tinggi, fase stasioner lebih cepat hperoleh. Perbedaan densitas inokulum merupakan suatu faktor yang mempengaruhi produksi biomassa dan juga waktu yang dibutuhkan untuk mencapai fase stasioner (Sikuli dan Demeyer, 1 997).
Berdasarkan bobot segar awal dari eksplan yaitu 3 eksplan awal(0.08 g), 5 eksplan awal(0.12 g), 7 eksplan awal(0.13 g) dan 9 eksplan awal(0.19 g), maka waktu penggandaan biomassa atau doubllng time dari akar berambut kurang dari 4 hari. Peningkatan biomassa segar akar berambut mencapai 6-7 kali bobot awal pada 4 HST, 21-36 kali bobot awal pada 8 HST clan 26 - 52 kali bobot awal pada 12 HST. Peningkatan biomassa segar tertinggi dihasilkan pada densitas eksplan awal yang paling rendah (3 eksplan awal). Hal tersebut kemungkinan disebabkan pada densitas eksplan yang lebih rendah, kompetisi sel atau jaringan dalam mendapatkan hara dan faktor pertumbuhan lainnya lebih rendah sehingga pertumbuhan &pat berlangsung cepat dalam jangka waktu yang lebih panjang.
Persentase bahan kering meningkat dengan bertambahnya urnur kultur. Hal ini kemunglunan pa& umur kultur yang lebih lanjut, akar berambut telah mensintesis berbagai senyawa organik disamping bertambahnya biomassa sel , sehingga akumulasi bahan kering juga meningkat.
Kadar protein total dari akar berambut cenderung menurun dengan meningkatnya umur kultur. Hal ini sejalan dengan tahap perkembangan sel dan jaringan, dimana pada awal pertumbuhan kultur, terjadi pembelahan sel secara aktif. Pembelahan sel yang cepat tersebut sejalan dengan peningkatan sintesis
protein primer yang memang diperlukan untuk untuk pertumbuhan dan perkembangan sel.
Protein yang tinggi pada fase awal pertumbuhan tersebut kemungkinan bukan merupakan protein yang berhubungan dengan mekanisme ketahanan sel terhadap patogen, tetapi merupakan protein struktur untuk pembangun sel. Savary dan Flores (1994), menyatakan bahwa protein yang berhubungan dengan ketahanan pada Trichosanthes kirilowii var japonicum yaitu trichosanthin lebih banyak dihasilkan pada fase pertumbuhan sekunder dari akar.
Pertumbuhan sekunder dari akar tanaman in vivo dicirikan oleh bentuk akar yang menyerupai umbi yang disebut storage root. Sedangkan pada akar berambut, pertumbuhan sekunder dari akar ditandai oleh morfologi akar berambut yang memendek, melebar dan bertambah gemuk weshy), kaku, dan densitas rarnbut akar yang rendah (Savary dan Flores, 1994).
Hasil protein total meningkat dengan bertambahnya umur panen disebabkan adanya peningkatan biomassa. Walaupun kadar protein total dari akar berambut menurun dengan bertarnbahnya urnur panen tetapi karena biomassanya yang tingg, maka hasil protein total tetap lebih tinggi.
Perbedaan densitas eksplan awal akan mempengaruhi pola pertumbuhan akar berambut. Pada densitas eksplan awal yang lebih tinggi, akar berambut akan lebih cepat mencapai fase stasioner. Senyawa metabolit sekunder maupun protein aktif pada tanaman diduga lebih banyak dihasilkan pada fase pertumbuhan stasioner. Karena itu dengan mempercepat tercapainya fase stasioner dan memelihara kultur akar berambut lebih lama pada fase tersebut kemungkinan dapat meningkatkan kadar protein aktifnya. Hal tersebut dapat dilakukan dengan
menggunakan densitas eksplan awal yang lebih tinggi. Meskipun demikian diperlukan identifikasi lebih lanjut tentang profil dan keberadaan protein aktif pada berbagai fase pertumbuhan.
Hasil uji pengaruh kasein hidrolisat menunjukkan bahwa kasein hidrolisat tidak berpengaruh nyata terhadap produksi biomassa akar berambut dari T. cucumerina L. Namun terdapat kecenderungan penurunan biomassa segar maupun biomassa kering dengan penambahan kasein hidrolisat hingga 150 mg/l. P e n m a n biomassa tersebut disebabkan percabangan lateral dan pemanjangan akar berambut yang cenderung tertekan pada kasein hidrolisat 150 mgll seperti terlihat pada Gambar 6.
Kasein hidrolisat merupakan senyawa organik kompleks sumber asam amino. Pengaruh negatif dari asam amino terhadap produksi biornassa pernah dilaporkan Wielanek dan Urbanek (1999) pada akar berambut dari Tropaleum
majus. Asam amino berupa L-cystein 0.6
m M
menekan produksi biomassa hingga 35% dari kontrol, sedangkan phenilulunine menekan produksi biomassa akar berambut hingga 40% di bawah kontrol. Sebaliknya kedua asam amino tersebut meningkatkan glucotropaeolin yang dihasilkan sekitar 40%-70% di atas kontrol.Penambahan kasein hidrolisat dalam kultur akar berambut dari
Trichosanthes cucumerina L. dalam penelitian ini hanya efektif sekitar 50 mg/l.
Pada konsentrasi 100 mg/l kadar protein tidak berbeda dengan yang dihasilkan pada 50 mg/l, sedangkan pada konsentrasi 150 mgll kadar protein menurun bahkan lebih rendah dan kontrol.
Hasil protein total sangat tergantung pada produksi bahan kering. Kasein hidrolisat cenderung menurunkan hasil protein total, karena pengaruhnya yang
negatif terhadap produksi biomassa. Dengan demikian, meskipun kasein hidrolisat 50 mgll meningkatkan kadar protein akar berarnbut, tetapi biomassa yang dihasilkan rendah, sehingga hasil akhir protein total lebih rendah dari kontrol.
Kesimpulan
Akar berambut dapat diinduksi dan tanaman Trichosanthes cucumerina L.
Efisiensi induksi akar berambut mencapai 43% dan terbatasnya galur akar berarnbut yang diperoleh disebabkan kesulitan menghilangkan kontaminasi
Agrobacterium rhizogenes.
Densitas eksplan awal yang optimum untuk menghasilkan biomassa yang adalah sebanyak 5 ujung akar per botol. Pada densitas eksplan awal 5 ujung akarhotol, produksi biomassa lebih tingg dibandingkan 3 ujung akarlbotol, namun tidak berbeda nyata dengan yang dihasilkan pa& 7 maupun 9 ujung akar per botol.
Hasil protein total tidak dipengaruhi oleh densitas eksplan, namun protein total tertinggi dihasilkan pada densitas eksplan 9 ujung akar per botol. Peningkatan umur panen hingga 12 HST masih dapat meningkatkan hasil protein total, karena pada waktu masih terjadi peningkatan produksi biornassa walaupun akar berarnbut sudah cenderung memasuki fase stasioner.
Penambahan kasein hidrolisat 50 dan 100 mg/l dapat meningkatkan kadar protein total dari akar berarnbut. Tetapi karena kasein hidrolisat cenderung menekan produksi biomassa, maka untuk mendapatkan biomassa dan hasil protein total tertinggi cukup dengan menggunakan media MS-0 tanpa penambahan kasein hidrolisat.
Narayanaswamy S. 1994. Plant Cell and Tissue Culture. T a t . McGraw-Hill Publishing Company Limited. New Delhi. 652p.
Ng TB, Liu-Wk, Tsao Sw, Yeung HW. 1994. Effect of trichosanthin and momorcharin on isolated rat hepatocytes. J Ethnopharm 43;2:8 1-87.
Payne J, Hamill JD, Robins
RJ,
Rhodes JC. 1987. Production of hyoscyamin by hairy root cultures of Datura stramonium. Planta medica 53:474-478. Payne GF, Bringi V, Prince CL, Shuler MI,. 1992. Plant Cell and Tissue Culturein Liquid Systems. New York: John Wiley and Sons.Bab 8, Root Cultures; hlm 227-277.
Savary BJ, Flores HE. 1994. Biosynthesis of defense-related protein in transformed root cultures of Trichosanthes kirilowii Maxim. Var.
japonicum (Kitam). Plant Physiol 106: 1 195- 1204
Shackelford NJ, Chlan CA. 1996. Identification of antibiotics that are effective in eliminating Agrobacterium tumefaciens. Plant Mol Biol Rep 14; 1 50-57. Sikuli
NN,
Demeyer K. 1997. Influence of ion composition of medium onalkaloid production by "hairy roots" of Dutura stramonium. Plant Cell Tissue Organ Cult 4 1 :26 1-267.
Terras FRG et ul. 1995. Small cysteine-rich antifungal proteins from radish: their role in host defense. The Plant CeIl7: 573-588.
Toppi LS di, Gorini P, Properzi G, Barbieri L, Spano L. 1996. Production of ribosome in-activating protein from hairy-root cultures of Lufla cyllindrica
(L.) Roem. Plant Cell Reports 15 : 9 10-91 3.
Toppi LS di, Pecchioni N, Durantee M. 1997. Cucurbita pepo L. can be transformed by Agrobacterium rhizogenes. Plant Cell. Tissue Organ Cult 5 1 : 89-93.
Tumilisar C et al.. 2001. Protein Bioaktif Asal Kultur Akar Transgenik Blustru
(Lufla cycyldrica (L.). Roem dan Mvitasnya dalam Menghambat
Proliferasi Sel Tumor Secara In Vitro. Disertasi. Program Pascasarjana Institut Pertanian Bogor.
Vivanco JM, Weitzel D, Flores HE. 1997. Characterization of a major storage root protein isolated from the andean root crop Mirabilis apansa. Di dalarn: Flores HE, Lynch JP, Eissenstat D. Radical biology: Advances and Perspectives on the Function of Plant Roots. Proceedings llth Annual Penn State Symposium in Plant Physiology. Amarican Society of Plant Physiologists, Rockville, Maryland U.S.A.
Wielanek M. Urbanek H. 1999. Glucotropaelin and myrosinase production in hairy root cultures of Tropaeolum majus. Plant Cell Tissue Organ Cult 57 : 39-45.