L
Journal of Experimental Marine Biology and Ecology 251 (2000) 227–238
www.elsevier.nl / locate / jembe
Ammonium and phosphate excretion in three common
echinoderms from Philippine coral reefs
*
Danilo T. Dy , Helen T. Yap
The Marine Science Institute, University of the Philippines, Diliman, 1101 Quezon City, Philippines
Received 15 November 1999; received in revised form 20 February 2000; accepted 20 April 2000
Abstract
The ammonium and phosphate excretion and oxygen consumption of three species of echinoderms (Tripneustes gratilla, Protoreaster nodosus and Ophiorachna incrassata) commonly encountered in Philippine coral reefs were investigated in relation to time of day (i.e. daytime between 10:00 and 12:00 h vs. nighttime between 22:00 and 24:00 h) and their recent feeding history (i.e. recently-collected vs. short-term starvation for 361 days). The experiment used whole organism incubations and followed a nested hierarchical design. Ammonium excretion rates were
21 21
14476310 nmol g DW h (mean6S.E., n524) for T. gratilla, 361633 for O. incrassata and 492638 for P. nodosus. Ammonium excretion differed significantly among species, time of incubation and recent feeding history. Interaction between species and recent feeding history was also significant. The organisms excreted more ammonium during daytime except for starved specimens of O. incrassata. In addition, animals that were starved in the laboratory for a few days had a tendency to excrete more ammonium than recently-collected specimens. Phosphate excretion
21 21
rates were 25613 nmol g DW h for T. gratilla, 1062 for O. incrassata and 461 for P.
nodosus. There were no significant differences in phosphate excretion among the three species of
echinoderms, their recent feeding history and time of day. Oxygen consumption rates were 21 21
286624mg O g2 DW h for T. gratilla, 6463 for O. incrassata and 5463 for P. nodosus. Oxygen consumption differed significantly among species and recent feeding history but differed only slightly with time of incubation. There was a significant correlation between oxygen consumption and ammonium excretion (r50.48, P50.018), and between oxygen consumption and phosphate excretion (r50.41, P50.047) for T. gratilla. The nutrient excretion by tropical echinoderms is another pathway by which inorganic nutrients are regenerated in coral reef communities. However, the quantity of nutrients excreted is dependent on the species of echinoderms, their nutritional status and time of day. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Ophiorachna incrassata; Nutrient excretion; Protoreaster nodosus; Tripneustes gratilla
*Corresponding author. Present address: Biology Department, University of San Carlos, 6000 Cebu City, Philippines. Tel.: 163-32-346-1128.
E-mail address: [email protected] (D.T. Dy).
1. Introduction
Coral reefs, which are productive ecosystems that grow in low nutrient waters, are composed of communities of corals, other animals and plants, and produce moderately sustainable fisheries with the input of few extra nutrients (Wilkinson, 1994). In most tropical reefs remote from big land masses, the growth of reef macrophytes is limited either by the availability of inorganic nutrients or the extent of grazing by fishes and sea urchins. Marine organisms excrete soluble inorganic nutrients to their surrounding waters in the form of ammonia (Wright, 1995) and phosphate (Pomeroy and Bush, 1959). These soluble excretory products are available to nutrient-limited primary producers and possibly contribute to the nutrient pool in coral reef waters. The role of macroinvertebrates in nutrient recycling has been studied by several authors. For example, Qian et al. (1996) showed that the alga, Kappaphycus alvarezii, treated with wastes of the pearl oyster, Pinctada martensi, grew much faster than when it was not exposed to oyster wastes. Taylor and Rees (1998) observed that 79% of the nitrogen required by a subtidal bed of fucalean algae came from ammonium excreted by mobile epifauna. Ambler et al. (1988) showed that productivity in Gracilaria was higher when ammonium excreted by the scallop Argopecten purpuratus was introduced. Ammonium excreted by sea urchins is another mechanism that maintains the high rates of primary productivity of algal turf on coral reefs (Williams and Carpenter, 1988). Mukai et al. (1989) pointed out that nitrogen released from megabenthic animals in a tropical seagrass meadow did not satisfy the nitrogen required for seagrass production but was enough to sustain planktonic and epiphytic algal production.
There are several factors affecting the regeneration of nutrients by macroinvertebrates in algal-dominated coral reefs. Aside from purely species differences and occupancy of different trophic guilds, other factors may come into play (Diehl and Lawrence, 1979; Stickle and Bayne, 1982; Davoult et al., 1991; Mingoa-Licuanan, 1993). We studied nutrient excretion and oxygen consumption rates of three common species of ech-inoderms found in tropical coral reefs, namely: Protoreaster nodosus (starfish),
Tripneustes gratilla (sea urchin), and Ophiorachna incrassata (brittle star). We
hypoth-esize that aside from taxonomic affiliation, the recent feeding history of the organism as well as time of day are factors that affect the above.
2. Materials and methods
2.1. Experimental design
D.T. Dy, H.T. Yap / J. Exp. Mar. Biol. Ecol. 251 (2000) 227 –238 229 Table 1
Nested hierarchical design outlining the various independent and dependent variables where D5daytime, N5nightime, F5recently collected (presumed fed), S5starved for 361 days
Independent variables Echinoderm species
T. gratilla O. incrassata P. nodosus
(sea urchin) (brittle star) (starfish)
Time of day D N D N D N
Recent feeding history F S F S F S F S F S F S
Dependent variables ←—————————— Oxygen consumption ——————————→
←—————————— Ammonium excretion ——————————→
←—————————— Phosphate excretion ——————————→
same species. The experimental run for each factor combination was conducted twice to give a total of six replicates.
2.2. Collection of the test organisms
Echinoderms were collected from a coral reef flat in the eastern side of Mactan Island, Central Philippines (Fig. 1). Collection involved skin diving and careful handpicking of the organisms, and placing them in plastic containers. They were immediately brought to the laboratory and acclimated for at least 6 h before subjecting them to the incubation experiments. The test animals were not fed during acclimation to minimize the release of fecal matter during the incubation proper which would possibly affect the measurements.
2.3. Experimental procedures
Before each experiment, seawater was collected at the same site as the organisms, passed through a 10-mm filter bag and poured into 2.7-l reaction flasks. The salinity ranged between 32.3 and 34.0‰. After initial water samples for dissolved oxygen, and ammonium and phosphate measurements were collected from each flask, the acclimated organisms, previously washed with filtered seawater, were placed in the reaction flasks. The flasks were filled with filtered seawater and made to overflow for 1 min and then covered, after which they were placed in a water bath (temperature between 23.2 and 25.08C) for 1–2 h. At the end of the incubation period, each flask was placed on top of a magnetic stirrer with gentle stirring for 3 min to homogenize the medium, after which final samples for dissolved oxygen, ammonium and phosphate concentration were collected.
After incubation, the volume of the organism was measured by seawater displace-ment. The organism was dried in an oven at 908C for at least 48 h or until constant weight.
2.4. Seawater analysis
Fig. 1. Map of the collecting site (Maribago, Mactan Is., Central Philippines).
collected in duplicate and analyzed using the classic Winkler method adapted by 21
D.T. Dy, H.T. Yap / J. Exp. Mar. Biol. Ecol. 251 (2000) 227 –238 231
2.5. Statistical analysis
Tests for significant differences due to the recent feeding history of the organisms which was nested under time of incubation which in turn was nested under species were carried out using nested (hierarchical) analysis of covariance (ANCOVA) at P50.05 with temperature and salinity acting as covariates. Dependent variables (oxygen consumption, ammonium and phosphate excretion) were initially tested for normality and homoscedasticity. Data were transformed (i.e. oxygen consumption using log ,10
ammonium excretion using square root and phosphate excretion using the sine function) because the dependent variables were heteroscedastic. In cases where significant differences were detected among the dependent variables after ANCOVA, Tukey’s H.S.D. test was used for post-hoc comparisons of the main effects. Planned comparisons were carried out to compare effects of recent feeding history and time of incubation for each echinoderm species. To check whether there was a correlation between oxygen consumption and nutrient excretion in each species, we used Pearson product-moment correlation (Zar, 1984).
3. Results
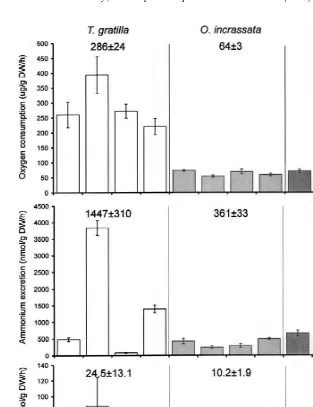
Oxygen consumption differed significantly among species and their recent feeding history, but less so with time of day (Fig. 2; Table 2). T. gratilla had significantly higher rates of oxygen consumption than the other two echinoderm species. Significantly higher oxygen consumption rates were also measured for starved T. gratilla during the day compared to night. Both O. incrassata and P. nodosus consumed more oxygen when recently collected than when the organisms were subjected to short-term starvation. Interaction between species and recent feeding history was significant during the day.
Ammonium excretion differed significantly among species, time of day and recent feeding history (Fig. 2; Table 2). Interaction between species and recent feeding history was also significant. T. gratilla had the highest rates of ammonium excretion followed by P. nodosus and then by O. incrassata. Post-hoc analysis indicated significant differences among the three species of echinoderms. Ammonium excretion by T. gratilla varied significantly in terms of time of day and recent feeding history. Starved organisms of this species excreted significantly more ammonium than recently collected in-dividuals. Ammonium excretion was significantly higher during the day compared to night.
Recently collected specimens of the brittle star, O. incrassata excreted significantly more ammonium during the day, while starved specimens excreted significantly more ammonium at night. Similarly, higher values were measured during the day compared to night for the starfish, P. nodosus but only recently collected specimens had significantly higher ammonium excretion during the day. Rates of ammonium excretion by P.
nodosus did not vary significantly with recent feeding history. There was a significant
interaction between time of day and recent feeding history for ammonium excretion by
Fig. 2. Rates (mean6S.E.) of oxygen consumption, and ammonium and phosphate excretion. (D5day, N5night, F5recently collected, S5starved). Top values are mean6S.E. of four treatments.
There were no significant differences in phosphate excretion among the three species of echinoderms, their recent feeding history and time of day (Fig. 2; Table 2).
D
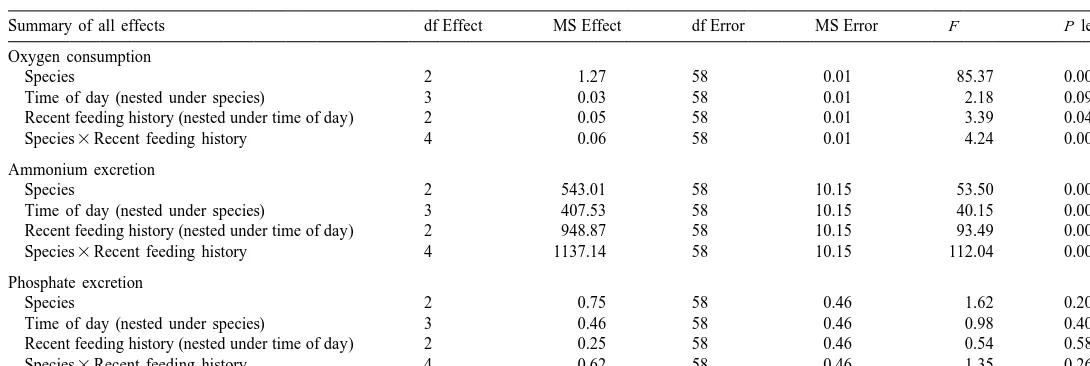
Summary of nested ANOVA results testing for the effect of species, time of day and recent feeding history on oxygen consumption, and ammonium and phosphate excretion
Summary of all effects df Effect MS Effect df Error MS Error F P level
Oxygen consumption
Species 2 1.27 58 0.01 85.37 0.0000*
Time of day (nested under species) 3 0.03 58 0.01 2.18 0.0999
Recent feeding history (nested under time of day) 2 0.05 58 0.01 3.39 0.0405*
Species3Recent feeding history 4 0.06 58 0.01 4.24 0.0044*
Ammonium excretion
Species 2 543.01 58 10.15 53.50 0.0000*
Time of day (nested under species) 3 407.53 58 10.15 40.15 0.0000*
Recent feeding history (nested under time of day) 2 948.87 58 10.15 93.49 0.0000*
Species3Recent feeding history 4 1137.14 58 10.15 112.04 0.0000*
Phosphate excretion
Species 2 0.75 58 0.46 1.62 0.2061
Time of day (nested under species) 3 0.46 58 0.46 0.98 0.4065
Recent feeding history (nested under time of day) 2 0.25 58 0.46 0.54 0.5854
Species3Recent feeding history 4 0.62 58 0.46 1.35 0.2631
true for P. nodosus (oxygen consumption vs. ammonium excretion r50.24, P50.250; oxygen consumption vs. phosphate excretion r5 20.27, P50.198).
3.1. O:N ratio
An estimate of the atomic ratio of oxygen-consumed to ammonia-nitrogen excreted (O:N ratio) was computed to provide an indication of the nutritional requirement of the three species of echinoderms. T. gratilla had an O:N ratio of 68.7619.5 while O.
incrassata had 13.261.3 and P. nodosus had 7.660.7.
3.2. Dry wt /volume ratio
Of the three echinoderm species used in this study, the brittlestar, O. incrassata had the highest dry wt / volume ratio (0.6060.04) followed by the starfish, P. nodosus (0.3760.01) and then by the sea urchin, T. gratilla (0.1560.00). The P. nodosus individuals used in this study were heavier than the other two echinoderms.
4. Discussion
Because echinoderms lack morphologically differentiated excretory organs and are ammonotelic (Lewis, 1967; Jangoux, 1982), the higher rates of ammonium excretion relative to phosphate excretion were expected. Phosphate excretion by the three echinoderms was, in most cases, minor or negligible and, in some instances, values were negative indicating extreme conservation of phosphate. Our phosphate excretion data for
21 21
T. gratilla (24.5613.1 nmol g DW h ) are higher than the value of 5.364.1 21 21
nmol g DW h reported by Vink and Atkinson (1985). The ophiuroid, Ophiocoma 21 21
scolopendrina, had a phosphate excretion rate of 7264 nmol g DW h (Ryabushko 21
and Propp, 1985) which is higher than our data for O. incrassata (10.261.9 nmol g 21
DW h ). On the other hand, the seastar, Echinaster luzonicus, had a phosphate 21 21
excretion rate of 562 nmol g DW h (Ryabushko and Propp, 1985) which is within 21 21
the same order of magnitude as our data for P. nodosus (4.161.2 nmol g DW h ). The low phosphate excretion rates by echinoderms are presumably due to the adaptation of the animals to the low amount of phosphorus in their natural food and is evidence of the effectiveness of phosphorus utilization by the organism (Ryabushko and Propp, 1985).
4.1. Nutrient excretion and oxygen consumption
D.T. Dy, H.T. Yap / J. Exp. Mar. Biol. Ecol. 251 (2000) 227 –238 235
T. gratilla is an herbivorous grazer. It feeds on seagrass and macroalgae which are known to have low caloric content. When T. gratilla is subjected to short-term starvation, catabolism ensues which not only leads to higher ammonium excretion but also to an active search for food (greater tube feet activity which entails more oxygen consumption) to meet its metabolic requirements. During daytime, changes in light intensity lead to increased tube feet and spine activities as observed by Millott (1956) in the covering reaction of sea urchins. Hence, the covering reaction should lead to more energy utilization (i.e. more oxygen uptake and more physiological output such as nutrient excretion) by T. gratilla. This explain the positive correlation between oxygen consumption and ammonium excretion in T. gratilla. A very low metabolic rate but high nitrogen diet could be a reason why this correlation was not significant for the other two species.
4.2. O:N ratio
The atomic oxygen:nitrogen ratio of 68.7619.5 for T. gratilla indicates that carbohydrate and / or lipid is the primary catabolic substrate of this species. T. gratilla is an herbivorous grazer widely found in seagrass meadows (Klumpp et al., 1993), coral–algal reefs and Philippine seaweed (mostly Eucheuma) farms (Gomez et al., 1983). On the other hand, the atomic O:N ratios of 13.261.3 for O. incrassata and 7.660.7 for P. nodosus indicate that these two species oxidize protein to some extent. The O:N ratios of the two species determined in this study are within the same order of magnitude reported by Ryabushko and Propp (1985) for the ophiuroid, Ophiocoma
scolopendrina (11.0) and for the starfish, Echinaster luzonicus (11.3). Brittle stars such
as O. incrassata are detritivores feeding on organic debris or small animals. Starfishes are locally regarded as nuisance predators in mussel farms. In coral–algal reefs, P.
nodosus probably feeds on both bivalves and gastropods. The differences in trophic
status exhibited by the three echinoderms in this study could account for the differences in their excretion rates and thus their different roles in nutrient recyling.
4.3. Excretion rates as a function of time
Daytime excretion rates were higher than nighttime rates for T. gratilla. This trend was similar for the sea urchin Centrostephanus coronatus in a temperate reef community off southern California (Bray et al., 1988). Ammonium excretion of recently collected
O. incrassata was also significantly higher during the day. Daytime ammonium excretion rates were higher for P. nodosus as well although this was not significant for starved specimens.
In the field, higher daytime excretion rates could be associated with higher daytime temperatures. The three echinoderm species inhabit coral reef flats where water temperatures are 1–28C higher during the day. Other organisms such as the Manila clam,
Tapes japonica (Mann and Glomb, 1978), are known to excrete more ammonium when
4.4. Excretion rates as a function of recent feeding history
Starved specimens of T. gratilla had significantly higher ammonium excretion. Starved specimens of both O. incrassata and P. nodosus had significantly higher ammonium excretion compared to recently collected individuals at least during the night. Phosphate excretion was also higher in starved specimens. This could be due to catabolism of tissue (Diehl and Lawrence, 1979). The relative increase in ammonium excretion by starved specimens was also noted by Davoult et al. (1991) for Ophiothrix
fragilis. In the reef, the varying degrees of starvation experienced by echinoderms during
a diurnal period should affect their ammonium excretion capacity. Davoult et al. (1991) suggested an effect of feeding activity and also tide on the excretion rates of echinoderms. He noted that excretion by Ophiothrix fragilis was always higher after slack water than before slack water.
4.5. Other considerations
The higher excretion rates of ammonium relative to phosphate indicate the potential contribution of ammonium to the immediate surroundings. In some reef areas where nutrients are limited, the ammonium excreted by macroinvertebrates could supplement the pool available to macrophytes. Excretion by macrofauna in temperate reef com-munities accounts for the general increase in ammonium near the reef substratum (Bray et al., 1988). Except for the brittle star, O. incrassata, which usually lives underneath rocky substrates and coral rubble, the two other echinoderm species can be seen wandering among algal beds. Since macroalgae in coral–algal reefs are already nutrient-limited, ammonium excreted by the wandering echinoderms can be taken up immedi-ately.
The macrophyte, Ulva curvata, for example, can utilize transient increases in ammonium concentrations resulting in higher growth rates (Ramus and Venable, 1987). The growth rate of a cultured species, Kappaphycus alvarezii, was higher when intermittently applied with ammonium fertilizer compared to the control (Li et al., 1990). In tropical coral reefs with low nutrient concentrations or in seaweed farms located in coral reef platforms, reef macrophytes such as K. alvarezii have the potential to absorb more inorganic nutrients than are available. Excretion by macroinvertebrates could possibly supplement nutrients in the water column in this nutrient-limited condition and during slack tide when water turnover rates are low.
The higher ammonium excretion during daytime could have positive effects on macrophytes which are actively photosynthesizing at this time. If this can be established and confirmed under field conditions, a possible coupling between macrophyte pro-duction and echinoderm excretion can be defined for coral reef systems as a form of beneficial relationship.
D.T. Dy, H.T. Yap / J. Exp. Mar. Biol. Ecol. 251 (2000) 227 –238 237
the physiology of the organisms (i.e. reproductive status) could also be important and will be the subject of future research.
Acknowledgements
˜
We thank C.R. Wilkinson, P. Alino and R. Dizon for reviewing the manuscript. We also thank E. Barrios and T. Punzalan for discussions on statistical analysis. Members of the International Association of Marine Science Libraries and Information Centers (IAMSLIC) were helpful in providing references. This study was financially supported by the Department of Science and Technology — Philippine Council for Aquatic and Marine Research and Development (DOST-PCAMRD), the Department of Agriculture — Bureau of Agricultural Research (DA-BAR) and the University of San Carlos. This is contribution no. 307 of the Marine Science Institute, University of the Philippines. [SS]
References
Ambler, R., Edding, M., Rodriguez, A., Escobar, S., 1988. Excretion of ammonium by Argopecten purpuratus (Mollusca, Pectinidae) and their utilization by Gracilaria sp. (Rhodophyta, Gracilariaceae): a possibility of polyculture. Gayana Bot. 45, 309–315.
Bray, R.N., Miller, A.C., Johnson, S., Krause, P.R., Robertson, D.L., Westcott, A.M., 1988. Ammonium excretion by macroinvertebrates and fishes on a subtidal rocky reef in southern California. Mar. Biol. 100, 21–30.
Davoult, D., Gounin, F., Janquin, M.A., 1991. Ammonium excretion by the ophiurid Ophiothrix fragilis as a function of season and tide. Mar. Biol. 111, 425–429.
Diehl, W.J., Lawrence, J.M., 1979. Effect of nutrition on the excretion rate of soluble nitrogenous products of Luidia clathrata (Say) (Echinodermata; Asteroidea). Comp. Biochem. Physiol. 62A, 801–806.
Gomez, E.D., Guieb, R.A., Aro, E., 1983. Studies on the predators of commercially important seaweeds. Fish. Res. J. Philipp. 8, 1–17.
Grasshoff, K., Erhardt, M., Kremling, K. (Eds.), 1983. Methods of Seawater Analysis, 2nd Edition. Verlag Chemie, Berlin, Germany, p. 419.
Jangoux, M., 1982. Excretion. In: Jangoux, M., Lawrence, J.M. (Eds.), Echinoderm Nutrition. A.A. Balkemia, Rotterdam, pp. 437–445.
Klumpp, D.W., Salita-Espinosa, J.T., Fortes, M.D., 1993. Feeding ecology and trophic role of sea urchins in a tropical seagrass community. Aquat. Bot. 45, 205–229.
Lewis, J.B., 1967. Nitrogenous excretion in the tropical sea urchin Diadema antillarum Philippi. Biol. Bull. 132 (1), 34–37.
Li, R., Li, J., Wu, C.Y., 1990. Effect of ammonium on growth and carrageenan content in Kappaphycus alvarezii (Gigartinales, Rhodophyta). Hydrobiologia 204–205, 499–503.
Mann, R., Glomb, S.J., 1978. The effect of temperature on growth and ammonia excretion of the Manila clam Tapes japonica. Estuar. Coast. Mar. Sci. 6, 335–339.
Millott, N., 1956. The covering reaction of sea urchins. I. a preliminary account of covering in the tropical echinoid Lytechinus variegatus (Lamarck), and its relation to light. J. Exp. Biol. 33, 508–523.
Mingoa-Licuanan, S.S., 1993. Oxygen consumption and ammonia excretion in juvenile Tridacna gigas (Linne, 1758): effects of emersion. J. Exp. Mar. Biol. Ecol. 171, 119–137.
Pomeroy, L.R., Bush, F.M., 1959. Regeneration of phosphate by marine animals. Int. Oceanogr. Cong. 1, 893–894.
Qian, P.Y., Wu, C.Y., Wu, M., Xie, Y.K., 1996. Integrated cultivation of the red alga Kappaphycus alvarezii and the pearl oyster Pinctada martensi. Aquaculture 147, 21–35.
Ramus, J., Venable, M., 1987. Temporal ammonium patchiness and growth rate. In: Codium and Ulva (Ulvophyceae). J. Phycol. 23, 518–523.
Ryabushko, V.I., Propp, L.N., 1985. Rates of respiration and nitrogen and phosphorus metabolism in echinoderms in the South China Sea. Biologiya Morya 11 (6), 331–335.
Stickle, W.B., Bayne, B.L., 1982. Effects of temperature and salinity on oxygen consumption and nitrogen excretion in Thais (Nucella) lapillus (L.). J. Exp. Mar. Biol. Ecol. 58, 1–17.
Strickland, J.D.H., Parsons, T.R. (Eds.), 1972. A Practical Handbook of Seawater Analysis. Fisheries Board of Canada, Bull. No. 167.
Taylor, R.B., Rees, T.A.V., 1998. Excretory products of mobile epifauna as a nitrogen source for seaweeds. Limnol. Oceanogr. 43, 600–606.
Vink, S., Atkinson, M.J., 1985. High dissolved C:P excretion ratios for large benthic marine invertebrates. Mar. Ecol. Prog. Ser. 21, 191–195.
Wilkinson, C.R., 1994. Living coastal resources of Southeast Asia: status and management. In: Report of the consultative forum third ASEAN–Australia symposium on living coastal resources. Chulalongkorn University, Bangkok, Thailand, p. 131.
Williams, S.L., Carpenter, R.C., 1988. Nitrogen-limited primary productivity of coral reef algal turfs: potential contribution of ammonium excreted by Diadema antillarum. Mar. Ecol. Prog. Ser. 47, 145–152. Wright, P.A., 1995. Nitrogen excretion: three end products, many physiological roles. J. Exp. Biol. 198,
273–281.