Impact of the rhizosphere on soil microarthropods in
agroecosystems on the Georgia piedmont
Carol J. Garrett
a,1, D.A. Crossley Jr.
a,∗, David C. Coleman
a,
Paul F. Hendrix
a,b, Keith W. Kisselle
a,2, Robert L. Potter
aaInstitute of Ecology, University of Georgia, Athens, GA 30602, USA bDepartment of Crop and Soil Sciences, University of Georgia, Athens, GA 30602, USA
Accepted 5 July 2000
Abstract
We pulse-labeled corn (Zea mays) and weed plants with photosynthetically fixed Carbon-14 to investigate the importance of the rhizosphere as a food source for soil microarthropod food webs. In field samples, we followed the movement of14C from plant shoots to roots to microarthropods. In conventionally tilled (CT) agroecosystems, the soil microarthropods accumulated radioactive tracer and reached concentrations as high as those in roots. In no-tillage (NT) agroecosystems, radioisotope concentrations in microarthropods were not as high. The CT systems lack surface organic litter, an alternate food base for microarthropods. Results suggest that feeding in the rhizosphere is more important in CT systems. Weeds transferred higher concentrations of tracer into the rhizosphere than did crop plants, suggesting that weeds in CT systems may be important in fueling food webs. © 2001 Elsevier Science B.V. All rights reserved.
Keywords: Carbon-14; Mite; Collembola; Weed; Maize
1. Introduction
Most current models of soil food webs depict plant residues as the main resource base, with living roots contributing relatively little. Failure to include roots explicitly in models is probably due to a dearth of information about their influences on food webs (Lussenhop, 1992). Curry and Ganley (1977) found that higher microarthropod numbers were associ-ated with grass roots than in bulk soil in pastures. Lussenhop and Fogel (1991) reported that
inverte-∗Corresponding author. Tel.:+1-706-542-2968; fax:+1-706-543-0646.
1Current address: Rt 1 Box 33-E, Huttonsville, WV 26273, USA. 2Current address: US Environmental Protection Agency, Athens,
GA 30605, USA.
brate densities were concentrated on roots of Bigtooth Aspen and Bracken in measurements made from a rhizotron. Preliminary research at the Horseshoe Bend Agroecosystem Facility, University of Georgia has suggested to us that the influence of roots on the soil community may be especially significant in agri-cultural systems (Crossley et al., 1992; Holland et al., 1996), where microarthropods may become numerous in the rhizosphere (Crossley, unpublished data).
We investigated the influence of the rhizosphere on soil microarthropods by use of a radioactive tracer. We used a pulse-labeling technique (Kisselle et al., 1999) to introduce radioactive Carbon-14 into corn plants (Zea mays L.), and then followed the movement of the tracer from above-ground plant stem to roots and then into soil microarthropods. In this way we were able to identify the root sources of input into the soil food web.
The rhizosphere contains labile carbon substances exuded by plant roots and is a zone of intense mi-crobial activity. Recent studies have shown that plant growth and development are controlled to a large ex-tent by the soil environment within the region of the root, an environment which the plant itself helps to create and where microbial activity has a major influ-ence (Curl and Truelove, 1986).
The usual definition of the term rhizosphere (the distance within 2 mm of the root surface) needs to be extended for studies of soil food chains that involve arthropods. If the term is intended to convey the con-cept “zone of influence of roots”, then the 2 mm dis-tance is inadequate. Our sampling methods extracted soil microarthropods from 5 cm dia. soil cores contain-ing crop or weed roots. We consider the microarthro-pods contained in these cores to be capable of entering the rhizosphere and feeding there, if not permanent residents. Microarthropods in laboratory cultures can easily move 5 cm or more in a day.
The amount of net primary production (NPP) reach-ing the roots of plants has been estimated to be as much as 60–80% (Coleman et al., 1988). While much of the photosynthate is converted to structural material, some is exuded as organic compounds. The amount of exudate reported varies widely (0.1–25% of NPP) (McCully and Canny, 1985). Corn root exudates have been reported to consist of mostly sugars, organic acids and a small amount of amino acids (Kraffcyzk and Beringer, 1985). Although mucilaginous material, root cap cells and root debris are supplied to the soil, the water soluble and volatile compounds are the most readily decomposable contributions of roots. Exudates appear to be responsible for the increase in microbial numbers in the rhizosphere as compared to nearby sites (Katznelson, 1965). Bacteria and fungi in soil systems are severely limited by the availability of en-ergy sources (Coleman et al., 1993). The rhizosphere is a zone of intense microbial activity and a source of energy for soil food chains.
Many of the soil fauna are microphagous or my-cophagous and occur in greater numbers in the vicin-ity of the rhizosphere. At the Horseshoe Bend Facilvicin-ity significantly more arthropods have been found in sam-ples centered over the stems of both crop and weed plants as compared with samples taken between crop rows (Crossley, unpublished data). Cropping systems maintained under a no-tillage regime (NT systems)
contain a large amount of litter residue on the soil sur-face, which provides soil food chains with an abundant food base. Conventionally tilled (CT) systems, in con-trast, lack such a surface cover of plant debris; plant residues are incorporated into the top 15 cm of soil. Our use of a radioactive tracer enabled us to compare root influences on food chains in these two different kinds of cropping systems.
2. Materials and methods
The research was conducted at Horseshoe Bend Agroecosystem Facility near the campus of The Uni-versity of Georgia, Athens, GA. The soil type is a Hiwassee sandy clay loam soil (clayey, kaolinitic, thermic, Typic Kanhapludult) as described by Groff-man et al. (1987). The research plots used in this ex-periment have been termed the “side plots” (Jerkins, 1994). They consists of six plots, 3 CT and 3 NT, col-lectively occupying approximately 0.1 ha. The tillage regimes have been maintained for 15 years.
Prior to this study, the plots were planted in crim-son clover (Trifolium incarnatum) as a winter cover crop. The summer crop, field corn (Zea mays), was planted in June 1996. In the CT plots the clover crop was mowed and plowed under to a depth of about 15 in. with a moldboard plow. The plots were then disked and tilled and corn seed was planted by direct drilling. In the NT plots the winter cover crop was mowed and the corn seed was directly drilled using a no-till planter. Both treatments were fertilized at a rate of 150 kg 10–10–10 ha−1.
Within each of the plots a mini-plot, 1 m×2 m, was established for tagging with14C. Each mini-plot was surrounded by a 20 cm wide plexiglass strip that extended 15 cm into the ground. Corn was re-planted in the mini-plots on June 9, 1996. They were watered regularly and some weeds removed. Each mini-plot initially contained 28 corn plants and about 28 weeds of various species.
2.1. Labeling procedures
mini-plot. A closed loop system of tubing with an in-line pump allowed injection of14CO2and unlabeled
CO2, continuously subsampled with an infrared gas
analyzer.
The labeling was begun on July 11, 1996 when the corn plants were about a meter tall. The labeling started each day at approximately 12:00 EDT and con-tinued for about 2 h. We used two chambers so that two plots could be labeled simultaneously, one CT and one NT. Thus, six plots were labeled on three consec-utive days as follows: July 11, Plots CT 9 and NT 10; July 12, CT 11 and NT 12; July 13, CT 13 and NT 14).
2.2. Sampling procedures
Sampling was begun three days after labeling, with each group of plots sampled in sequence, i.e., Plots CT 9 and NT 10 on July 13, CT 11 and NT 12 on July 14, and CT 14 and NT 14 on July 15. The next sequence of samples was taken 3 days after tagging (July 16–18), with further sample sequences at 13, 20, 28 and 42 days after tagging, for a total of six sampling dates. Two corn plants and two weeds were sampled from each mini-plot on each sample date. The weeds sampled were four species: Johnson grass (Sorghum
halepense (L.) Persoon), sicklepod (Cassia obtusifolia
L.), Bermuda grass (Cynodon dactylon (L.) Persoon) and morning glory (Ipomea purpurea (L.) Roth). The various weed species were combined for analysis. The weeds were cut at the soil surface level and a 5 cm×
5 cm soil core was taken directly over the cut stem. The shoot itself was bagged separately. The corn plant was sampled similarly, except that three 5 cm×5 cm soil cores were taken immediately adjacent to the plant stalk rather than directly over it, because of the large size of the corn stalk.
2.3. Sample processing
Soil cores were immediately placed in high-effi-ciency, Tullgren-type extractors (Crossley and Blair, 1991) for 4–6 days. Roots were then removed from the cores by hand sorting.
Microarthropods from the three soil cores taken around each corn plant were combined into a single sample. They were enumerated and sorted into the following taxa: Collembola, Prostigmata, Mesostig-mata, immature Oribatida, and adult Oribatida. (Other
microarthropod taxa were too scarce to yield sufficient numbers for estimating radiocarbon content.) Vials containing each faunal group in 70% alcohol were placed in a hood at room temperature to evaporate the alcohol. The arthropods were then digested with 1 ml of Scintigest and 1 ml of deionized water in a 55–60◦C oven overnight. The samples were then neutralized with 1 ml of 0.6 M acetic acid and sonicated for 3 min to aid the digestion process. Approximately, 20 ml of Scintiverse was added to each vial and 14C activity
was measured with a Beckman scintillation counter. The corn plants were oven-dried at 55◦C. They were then weighed, sorted into stem and leaf, ground, and subsamples oxidized in a Harvey biological ox-idizer. 14C activity was measured on a Beckman scintillation counter. The roots were processed with similar procedures.
Biomass values for functional groups of micro-arthropods were calculated using the average individ-ual dry weights suggested by Petersen and Luxton (1982) as follows: Collembola, 2.7mg; Oribatida, 5.3mg; Mesostigmata, 7.7mg; Prostigmata, 1.0mg.
3. Results
3.1. Microarthropod numbers
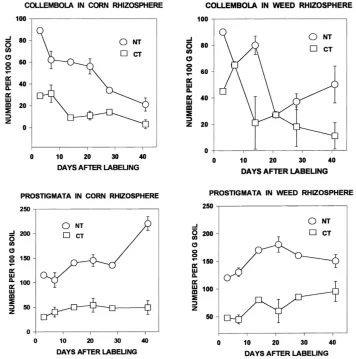
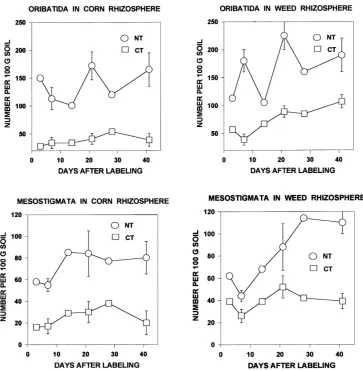
Microarthropod populations were consistently larger in rhizosphere samples from no-tillage systems compared with conventional tillage (Figs. 1 and 2). Samples from NT plots usually contained more than 500 microarthropods per 100 g soil, but samples from CT plots rarely exceeded 250 per 100 g soil. Further-more, each of the microarthropod groups was more abundant in NT versus CT rhizospheres (Figs. 1 and 2). The differences in abundance between NT and CT were statistically significant (P <0.01, t-test of paired comparisons) except for the Collembola.
Fig. 1. Abundance of Collembola and Prostigmata in rhizospheres of corn and weed plants grown under no-tillage and conventional tillage regimes. Standard errors shown for representative points.
Populations of collembolans (Fig. 1) declined dur-ing the sampldur-ing period (July–August 1996), but mite populations generally increased (Figs. 1 and 2). These trends are clear in the NT plots but less apparent in the samples from CT plots, where population sizes were smaller. The lower populations in the CT plots were also less variable between sampling dates. Oribatida in the NT plots showed the largest date-to-date varia-tion in numbers.
3.2. 14C radioactivity
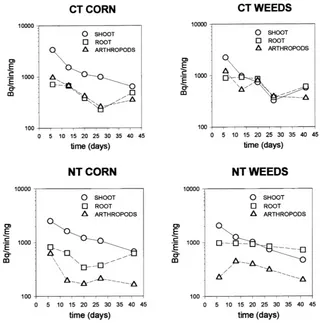
The pulse of radioactive14C generated by fixation of 14CO2 was detected in its movement from plant
shoots to roots to arthropods feeding in the rhizosphere (Fig. 3). The tracer appeared in the rhizosphere food web within 5 days of labeling. After labeling, corn and weed shoots showed similar levels of radioactivity (2500–3500 Bq/min/mg; Fig. 3) despite considerable variation from plant to plant (Kisselle et al., 1999). Radioactivity of the corn and weed shoots declined rapidly and at similar rates of approximately 0.03 per day. Radioactivity of the roots declined only slightly during the 42-day field study. In general,14C concen-trations in microarthropods tracked closely the con-centrations in the roots.
Fig. 2. Abundance of Oribatida and Mesostigmata in rhizospheres of corn and weed plants grown under no-tillage and conventional tillage regimes. Standard errors shown for representative points.
Under CT, microarthropods rapidly attained14C con-centrations very similar to those in the plant roots (Fig. 3). Under NT conditions,14C concentrations in microarthropods were significantly lower than those in roots. This difference persisted in both corn and weed rhizospheres. There was considerable variation in radioactivity among components in the CT weed systems but much less in the NT weed systems.
4. Discussion
The higher number of microarthropods found in NT than in CT has been previously reported for
Fig. 3. Concentrations of14C (Bq/min/mg) in shoots, roots and microarthropods from rhizospheres of corn and weed plants grown under
no-tillage and conventional tillage regimes.
higher C, N and water contents in the surface soil under no-till and consequently higher microbial pop-ulations (Coleman, 1986). NT systems have been shown to be primarily fungal-based, which support a higher population of mycophagous microarthropods (Hendrix et al., 1986; Beare et al., 1992).
Instances have been reported in which CT does not result in lower numbers of microarthropods (Parmelee, 1985), possibly due to drought conditions (Perdue and Crossley, 1989). Precipitation data for July and August 1996 reveal that there was no significant departure from normal rainfall during this time.
Our finding of declines in collembolan populations and increases in mite numbers, especially oribatid and prostigmatic mites, is a usual mid-summer sequence (Crossley et al., 1992; Beare et al., 1995; Parmelee,
1985; Stinner et al., 1986). Some families of Prostig-mata respond rapidly to disturbances, such as plowing or cultivation (Crossley et al., 1992). A small contri-bution of oribatid mites in proportion to the total mite population is most often associated with high numbers of Prostigmata (Petersen and Luxton, 1982).
Radioactivity of microarthropods appeared to reach a transient equilibrium with radioactivity of the roots, since root and arthropod14C concentrations varied in synchrony over most of the sampling periods. This rapidly attained equilibrium implies that14C acquired by microarthropods is turned over rapidly, so that in-gestion soon equals loss of tracer. The primary means of loss of tracer is probably via respiration.
significant feeding in the rhizosphere. Labeled exu-dates (organic compounds) entering the rhizosphere to be accumulated by microbes, or lysates released from senescent root cells, or direct feeding on the roots themselves are possible mechanisms by which the microarthropods could accumulate radioactivity.
Oribatida and Collembola are saprophagous or my-cophagous and may feed upon the many groups of fungi, saprobic, pathogenic, and mycorrhizal, which may colonize active roots. Klironomos and Kendrick (1995) suggest that the arthropod community may not be directly attracted to the root biomass, but may respond positively to roots that are colonized by a high diversity of darkly pigmented microfungi. In the litter zone, where few roots are found the animals still respond positively to a high diversity of darkly pigmented microfungi colonizing decaying leaf litter (Klironomos and Kendrick, 1995). The mesostigmatid mites in the Horseshoe Bend soils are mostly preda-tors of nematodes but some (Uropodini) are saprovores (Mueller et al., 1990).
Our tracer study revealed that rhizosphere inputs were more important in CT plots than in NT plots. Faunal concentrations of14C in NT plots were signifi-cantly lower than those of either the fauna or the roots in the CT systems. The lower tracer concentrations in the fauna in NT are probably due to feeding upon un-labeled sources, such as fungi in the litter layer. Previ-ous work at Horseshoe Bend found that, during July, 80% of surface residues remained undecomposed but only 50% of buried residues remained (Beare et al., 1992). The surface mulch in NT thus provided an al-ternate food source that was rare or absent in the CT plots.
Weed rhizospheres contained roots with 14C con-centrations nearly as high as concon-centrations in shoots. Soil fauna had equally high concentrations of tracer in CT plots. In weed rhizosphere samples from the NT plots, faunal14C concentrations were lower than sam-ples from the CT plots. However, these weed plot val-ues were higher than the14C concentrations in fauna from corn root samples. These findings suggest that weed rhizospheres may be more important than crop rhizospheres in supporting soil food webs. This is log-ical, since crop plants are selected to maximize their above-ground NPP, unlike weeds. Weeds may be a sig-nificant factor for the protection of soil biodiversity, especially in CT agroecosystems.
Acknowledgements
This research was supported by grants from the National Science Foundation. We thank Betty Weise, Jennifer Rice and Glen Stevens for their assistance in field and laboratory.
References
Beare, M.H., Parmelee, R.W., Hendrix, P.F., Cheng, W., Coleman, D.C., Crossley Jr., D.A., 1992. Microbial and faunal interac-tions and effects on litter nitrogen and decomposition in agroecosystems. Ecol. Monographs 62, 569–591.
Beare, M.H., Coleman, D.C., Crossley Jr., D.A., Hendrix, P.F., Odum, E.P., 1995. A hierarchical approach to evaluating the significance of soil biodiversity to biogeochemical cycling. Plant Soil 170, 5–22.
Coleman, D.C., 1986. The role of microfloral and faunal interactions in affecting soil processes. In: Mitchell, M.J., Nakas, J.P. (Eds.), Microfloral and Faunal Interactions in Natural and Agroecosystems. Martinus/Junk, Dordrecht. Coleman, D.C., Crossley Jr., D.A., Beare, M.H., Hendrix, P.F.,
1988. Interactions of organisms at root/soil and litter/soil inter-faces in terrestrial ecosystems. Agric. Ecosyst. Environ. 24, 117–134.
Coleman, D.C., Hendrix, P.F., Beare, M.H., Cheng, W.X., Crossley Jr., D.A., 1993. Microbial and faunal interactions as they affect soil organic matter in subtropical agroecosystems. In: Paoletti, M.G., Foissner, W., Coleman, D.C. (Eds.), Soil Biota, Nutrient Cycling and Farming Systems. Lewis Publishers, Boca Raton, FL, pp. 1–14.
Crossley Jr., D.A., Blair, J.M., 1991. A high-efficiency, “low-technology” Tullgren-type extractor for soil microarthropods. Agric. Ecosyst. Environ. 34, 187–192.
Crossley Jr., D.A., Mueller, B.R., Perdue, J.C., 1992. Biodiversity of microarthropods in agricultural soils: relations to processes. Agric. Ecosyst. Environ. 40, 37–46.
Curl, E.A., Truelove, B., 1986. The Rhizosphere. Springer, New York.
Curry, J.P., Ganley, J., 1977. The arthropods associated with the roots of some common weed and grass species. Ecol. Bull. 25, 330–339.
Groffman, P.M., Hendrix, P.F., Crossley Jr., D.A., 1987. Nitrogen dynamics in conventional and no-tillage agroecosystems with inorganic fertilizers or legume nitrogen inputs. Plant Soil 97, 315–332.
Hendrix, P.F., Parmelee, R.W., Crossley Jr., D.A., Coleman, D.C., Odum, E.P., Groffman, P.M., 1986. Detritus food webs in conventional and no-tillage agroecosystems. Bioscience 36, 374–380.
Holland, J.N., Cheng, W., Crossley Jr., D.A., 1996. Herbivore-induced changes in plant carbon allocation: assessment of below-ground C fluxes using carbon-14. Oecologia 107, 87– 94.
Jerkins, D.J., 1994. Agroecosystem response of soil and microbial components to different tillage and nutrient applications. Ph.D. dissertation. University of Georgia, Athens, GA.
Katznelson, H., 1965. Nature and importance of the rhizosphere. In: Baker, K.F., Snyder, W.G. (Eds.), Ecology of Soil-Borne Plant Pathogens. Murray, London, pp. 187–209.
Kisselle, K.W., Garrett, C.J., Hendrix, P.F., Crossley Jr., D.A., Coleman, D.C., 1999. Method for14C-labeling maize field plots and assessment of label uniformity within plots. Commun. Soil Sci. Plant Anal. 30, 1759–1771.
Klironomos, J.R., Kendrick, B., 1995. Relationships among microarthropods, fungi, and their environment. Plant Soil 170, 183–197.
Kraffcyzk, G.T., Beringer, H., 1985. Soluble root exudates of maize: influence of potassium supply and rhizosphere microorganisms. Soil Biol. Biochem. 16 (4), 315–322. Lussenhop, J., 1992. Mechanisms of microarthropod–microbial
interactions in soil. Adv. Ecol. Res. 23, 1–33.
Lussenhop, J., Fogel, R., 1991. Soil invertebrates are concentrated on plant roots. In: Keister, D.L., Cregan, P.B. (Eds.), The
Rhizosphere and Plant Growth. Kluwer Academic Publishers, Dordrecht, p. 111.
McCully, M.E., Canny, M.J., 1985. Localisation of translocated C-14 in roots and root exudates of field grown maize. Physiol. Plant 65, 380–392.
Mueller, B.R., Beare, M.H., Crossley Jr., D.A., 1990. Soil mites in detrital food webs of conventional and no-tillage agroecosystems. Pedobiologia 34, 389–401.
Parmelee, R.W., 1985. Decomposition and microarthropod response in conventional tillage and no-tillage agroecosystems. MS Thesis. University of Georgia, Athens, GA, 48 pp. Perdue, J.C., Crossley Jr., D.A., 1989. Seasonal abundance of
soil mites (Acari) in experimental agroecosystems. Effects of drought in no-tillage and conventional tillage. Soil Till. Res. 15, 117–124.
Petersen, H., Luxton, M., 1982. A comparative analysis of soil fauna populations and their role in decomposition processes. Oikos 39, 288–388.
Stinner, B.R., Krueger, G.R., McCartney, D.A., 1986. Insecticide and tillage effects on pest and non-pest arthropods in corn agroecosystems. Agric. Ecosyst. Environ. 15, 11–21. Winter, J.P., Voroney, R.P., Ainsworth, D.A., 1990. Soil