STUDI PENDAHULUAN: POTENSI PADANG LAMUN SEBAGAI KARBON
ROSOT DAN PENYERAP KARBON DI PULAU PARI, TELUK JAKARTA
oleh
WAWAN KISWARA
Pusat Penelitian OseanografiReceived 9 March 2010, Accepted 16 November 2010
ABSTRAK
Analisis siklus karbon pada ekosistem pesisir menunjukkan pentingnya keberadaan vegetasi pesisir seperti alga makro, lamun, dan mangrove, tetapi diabaikan dari penghitungan siklus karbon di lautan. Pengabaian ini dimungkinkan akibat terbatasnya luasan vegetasi pesisir, yang hanya sekitar < 2 % dari permukaan samudera. Penelitian ini bertujuan mengetahui peranan jenis-jenis lamun (Enhalus acoroides, Cymodocea rotundata dan Thalassia hemprichii) sebagai karbon rosot (carbon stock) dan penyerapan karbon (carbon sink) yang dilakukan pada bulan April dan Juli 2009 di Pulau Pari, Teluk Jakarta. Analisis kandungan karbon dalam bagian tanaman lamun dilakukan dengan metode Walkley & Black. Persentase karbon pada bagian tanaman lamun tertinggi didapat pada rimpang yang berkisar antara 41,53 – 50,63 % dan terendah pada akar antara 26,79 – 37,77 %. Karbon rosot oleh lamun Enhalus acoroides, Cymodocea rotundata dan Thalassia hemprichii di padang lamun Pulau Pari berkisar antara 30,62 – 545,33 gr.C.m-2 dan penyerapannya berkisar antara 0,20 – 1,83 gr.C.m-2.h-1. Untuk mendapatkan informasi yang lebih lengkap tentang karbon rosot dan penyerapannya di padang lamun Pulau Pari, masih diperlukan penelitian lebih lanjut dengan mengukur kandungan karbon pada lamun C. serrulata, H. ovalis, H. uninervis, dan S. isoetifolium serta pemetaan untuk mengukur luasnya.
Kata kunci: Lamun, karbon rosot, penyerapan karbon, Pulau Pari, Teluk Jakarta.
ABSTRACT
PRELIMINARY STUDY: ABILITY OF SEAGRASS BEDS AS CARBON
STOCK AND CARBON SINK AT PARI ISLAND, JAKARTA BAY. Analysis of carbon
cycle in the coastal ecosystems shows the important of marine vegetation including macro algae, \seagrass and mangrove, although they are neglected from accounting of carbon cycle in the ocean. The neglect is possibly a consequence of the limited extent of marine vegetation, which cover only < 2 % of the ocean surface. The study aims to understand the role of seagrasses (Enhalus acoroides, Cymodocea rotundata and Thalassia hemprichii) as carbon stock and carbon sink, which carried out in April and July 2009 at Pari Island, Jakarta Bay. Percentage of carbon in the plant parts were analyzed by method of Walkley & Black. The highest percentage of carbon in the plant part of seagrasses was found in the rhizome 41,53 – 50,63 % and the lowest in the roots 26,79 – 37,77 %.. Carbon stock of Enhalus acoroides, Cymodocea rotundata and Thalassia hemprichii at seagrass beds of Pari Islands varied from 30,62 to 545,33 g.C.m-2 and their carbon sink varied from 0,20 to 1,83 g.C.m-2.d-1. To have more information on carbon stock and carbon sink in the seagrass beds of Pari Island needs measurement of carbon content in the seagrass of C. serulata, H. ovalis, H. uninervis and S. isoetifolium and mapping to measure their area.
PENDAHULUAN
Ekosistem lautan mencakup komunitas yang didominasi oleh vegetasi pantai,
termasuk alga makro yang dominan pada dasar berbatu, lingkungan estuaria dan terumbu karang serta tanaman tingkat tinggi yang mendominasi area pasang surut (hutan bakau di daerah tropis, rawa payau di daerah temperate) dan padang lamun yang tumbuh pada dasar pasir dan lumpur dan kadang-kadang pada dasar berbatu yang dijumpai sepanjang daerah pesisir dunia (DUARTE et al. 2005).Walaupun vegetasi tersebut diketahui merupakan suatu yang sangat penting dalam memelihara biodiversitas laut (ALONGI 2002; DUARTE et al. 2005), vegetasi pantai (alga makro, lamun, rawa payau dan mangrove) diabaikan dari penghitungan siklus karbon di lautan. Pengabaian ini dimungkinkan akibat terbatasnya luasan vegetasi pesisir, yang mana hanya sekitar < 2 % dari permukaan samudera (DUARTE & CEBRIAN 1996).
Analisis yang ada dalam siklus karbon ekosistem yang didominasi oleh vegetasi pesisir menunjukkan pentingnya keberadaan alga makro, padang lamun, dan mangrove, seperti respirasi global yang mencapai 7-8 PgCth-1 (MIDDELBURG et al. 2004). Jumlah yang besar ini menyebabkan karbon di pesisir dapat dianggap sebagai bagian dari timbunan karbon di lautan. Lebih lanjut dari analisis timbunan karbon dari komunitas vegetasi dasar laut menunjukkan ekspor dalam jumlah yang besar ke ekosistem sekelilingnya dan juga menyimpan sejumlah karbon organik di sedimennya. Waktu pulih vegetasi pesisir sangat lambat, seperti yang sering didominasi oleh organisme yang berumur panjang seperti bakau yang dapat mencapai umur puluhan tahun (TOMLINSON 1994; DUARTE et al. 2005) dan lamun yang dapat mencapai umur sampai beberapa dekade (DUARTE & HEMMINGA 2000).
Kecepatan penguburan karbon organik di area vegetasi pesisir dari DUARTE et al. 2005 dapat dilihat di Tabel 1. NIENHUIS et al. (1989) berdasarkan riset di Laut Flores melaporkan kandungan karbon beberapa jenis lamun (Enhalus acoroides Cymodocea rotundata dan Thalassia hemprichii), berkisar antara 31,0 – 32,5 % C dan 26,2 – 32,9 % C masing-masing untuk bagian bawah (akar dan rimpang) dan bagian atas (pelepah dan helaian daun).
Tabel 1 : Taksiran penguburan karbon organik di area vegetasi pantai (DUARTE et al. 2005).
Table 1: Estimate of organic carbon burial in vegetated areas (DUARTE et al. 2005). Component Area (Tm2) g Cm-2 y-1 Tg y-1 Mangrove Salt marsh Seagrass 0.2 0.4 0.3 139.0 151.0 83.0 23.6 60.4 27.4 Keterangan : T = 1012
Informasi mengenai peran padang lamun sebagai penyimpan dan laju penyerapan karbon dari perairan Indonesia belum pernah terungkap. Penelitian ini merupakan penelitian pendahuluan yang bertujuan untuk mengetahui peranan beberapa jenis lamun sebagai penyimpan karbon melalui kajian luas tutupan, kerapatan dan biomasanya, laju penyerapan karbon melalui kecepatan pertumbuhan dan produksinya, serta sumber karbon melalui gugur serasahnya.
BAHAN DAN METODE
Penelitian lapangan pengukuran cadangan karbon pada padang lamun di rataan
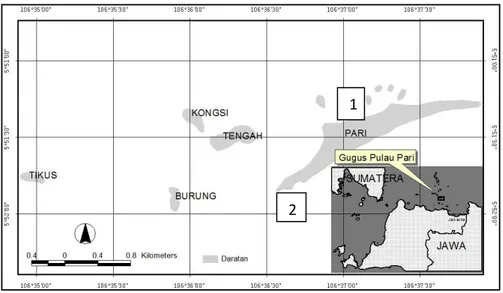
terumbu Pulau Pari dilaksanakan pada bulan April dan Juli 2009. Observasi pendahuluan dilakukan untuk mengetahui komposisi jenis lamun, sebaran, tipe vegatasi, tipe substrat dankedalaman perairan. Stasiun penelitian untuk mengukur luas tutupan, kerapatan, biomas, pertumbuhan dan produksi serta gugur serasah lamun ditentukan berdasarkan hasil observasi pendahuluan, yakni Stasiun 1 di Utara untuk tegakan murni Enhalus acoroides dan Stasiun 2 di Barat Pulau Pari untuk tegakan campuran E. acoroides, Thalassia hemprichii dan Cymodocea rotundata serta tegakan campuran T. hemprichii dan C. rotundata (Gambar 1). Pengukuran luas tutupan, kerapatan dan biomas lamun dilakukan dengan menggunakan bingkai besi ukuran 50 x 50 cm2 untuk E. acoroides dan ukuran 20 x 20 cm2 untuk T. hemprichii dan C. rotundata. Substrat tempat pengambilan sampel dicuplik untuk diukur butirannya.
Pengukuran pertumbuhan dan produksi daun lamun dilakukan dengan metode penandaan pertumbuhan daun dari ZIEMAN (1974) yang dimodifikasi BROUNS (1985) dan ERFTEMEIJER (1993). Pengamatan pertumbuhan lamun dilakukan selama lima hari. Metode ini dapat mengukur pertumbuhan daun lamun (cm/hari) dan produksi neto daun lamun (g.bobot kering.m-2.hari-1) yang dapat dikonversi dengan %C/bobot kering menjadi g.C.m-2.h-1 (BARRÓN et al. 2004).
Pengukuran gugur serasah lamun dilakukan dengan menggunakan kurungan dari jaring kelambu 1x1 m2 (NOJIMA & MUKAI 1996). Pengambilan serasah dilakukan pada contoh yang melayang dan terhampar di dasar kurungan.
Pengambilan contoh lamun untuk mengukur luas tutupan, kerapatan dan biomas dilakukan dengan tiga ulangan, sedangkan contoh untuk pengukuran pertumbuhan dan produksi lamun dilakukan sebanyak 10 tunas.
Analisis karbon dalam sampel berat kering bagian-bagian tanaman (akar, rimpang, pelepah daun dan helaian daun) dan serasah lamun (% C/berat kering) dikerjakan di laboratarium Departemen Ilmu Tanah dan Sumberdaya Lahan Fakultas Pertanian IPB dengan metode Walkley & Black (SCHUMACHER, 2002).
Metode Walkley & Black dilakukan sebagai berikut, satu gram sampel kering dimasukkan ke dalam labu takar 50 ml, ditambahkan 10 ml 0,167 M K2Cr2O7 dan 10 ml H2SO4 pekat lalu dikocok. Warna merah jingga pada larutan harus tetap terjaga. Jika terjadi perubahan warna menjadi hijau atau biru maka ditambahkan K2Cr2O7 dan H2SO4, jumlah penambahan ini harus dicatat. Penambahan untuk blangko juga harus sama banyak. Larutan kemudian didiamkan sekitar 30 menit hingga dingin. Setelah dingin kemudian ditambahkan 5 ml H3PO3 85% dan 1 ml indikator difenilamin, dan larutan diencerkan dengan akuades hingga volume larutan mencapai 50 ml. Lima ml larutan dipipet ke dalam erlenmeyer 50 ml dan ditambahkan 15 ml akuades, kemudian ditritasi dengan larutan FeSO4 1N atau 0,5 N hingga warna menjadi kehijauan. Prosedur tersebut dilakukan terhadap sampel dan blangko. Rumus menghitung karbon adalah sebagai berikut:
(B-A) x M FeSO4 x 12 x 100 % C = --- gr sampel x 4000 Keterangan : B = ml titrasi blanko A = ml titrasi sampel
12/4000 = miliequivalent berat dari C dalam gram
. Untuk menghitung biomas dalam satuan gram karbon per meter persegi (g.C.m-2) mengikuti BARRÓN et al. (2004), yang menghitung bobot karbon biomas per m-2 (g.C.m-2) bagian tanaman lamun dari g.bobot kering (g.BK.m-2) yang dikonversi ke mol C dengan persentase kandungan karbon (% C) bobot kering biomasnya, sehingga biomas dan produksi neto lamun dapat ditentukan dalam satuan g.C.m-2.
Gambar 1: Lokasi penelitian penyimpanan dan penyerapan karbon di rataan terumbu Pulau Pari, Teluk Jakarta, 1 = Utara; 2 = Barat Figure 1: Study site of carbon stock and carbon sink at reef flat of Pari Island, Jakarta Bay, 1 = North; 2 = West
Pengukuran ukuran butiran pasir dilakukan dari substrat yang diambil dengan plexiglas ukuran 5 cm sedalam 20 cm. Contoh disaring secara basah dengan saringan bertingkat, hasil saringan dikeringkan dengan oven pada suhu 80o C, kemudian ditimbang bobot keringnya. Hasil penimbangan bobot kering digunakan untuk menghitung persentase ukuran butiran pasir.
Pengukuran salinitas perairan lokasi penelitian dilakukan dari contoh air laut yang diambil dengan menggunakan botol sampel salinitas. Nilai salinitas contoh air laut diukur dengan salinometer merk Beckman RS7C. Suhu air laut diukur dengan memakai thermometer air raksa.
HASIL DAN PEMBAHASAN
Data butiran pasir, salinitas dan suhu serta kerapatan, luas tutupan, persentase kandungan karbon dalam bagian tanaman lamun, biomasa lamun dan serasah serta pertumbuhan dan produksi lamun disajikan dalam Tabel 1 s.d. 8. Tipe subtrat stasiun Pari Barat lebih banyak dicirikan oleh subtrat pasir kasar (45,87 % dan 46,47 %), sementara Stasiun Pari Utara lebih banyak dicirikan oleh substrat pasir halus dan lumpur dengan komposisi masing-masing 42,10 % dan 33,83 % (Tabel 2). Karakteristik pasir kasar dan pasir halus di lokasi penelitian merupakan ciri khas substrat karbonat dari pulau-pulau karang seperti yang dilaporkan oleh ERFTEMEIJER (1993) yang mendapat 28,7 – 46,1 % pasir di Kepulauan Spermonde.
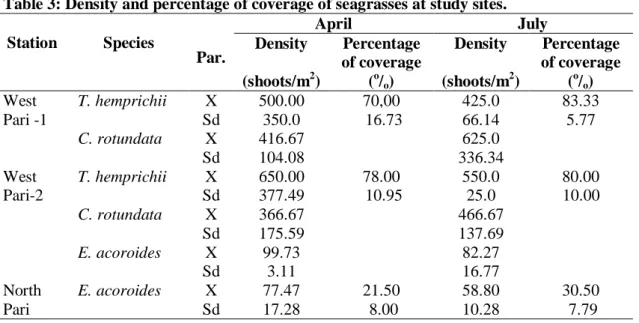
Luas tutupan rata-rata lamun di stasiun penelitian Pari Utara berkisar antara 21,50 % -
30,50 % dan luas tutupan di stasiun Pari Barat berkisar antara 70 % - 83,33 %. Luas tutupan rata-rata lamun di stasiun penelitian Pulau Pari Barat lebih tinggi dari pada di Pulau Pari Utara (Tabel 3). Komposisi jenis lamun di Pari Utara adalah monospesifik E. acoroides sementara komposisi jenis lamun di Pari Barat adalah campuran dua jenis lamun T. hemprichii dan C. rotundata serta campuran tiga jenis lamun, yaitu T. hemprichii, C.1
rotundata dan E. acoroides (Tabel 3). Luas tutupan lamun dipengaruhi oleh genangan air, lamun yang selalu tergenang air saat surut mempunyai luas tutupan yang lebih tinggi dibandingkan lamun yang terbuka saat surut. Lamun yang terbuka saat surut daun-daunnya terbakar panas matahari sehingga menurunkan luas tutupannya (DE IONGH 1996).
Tabel 2: Ukuran butiran pasir substrat di stasiun penelitian. Table 2: Grain size of substrates at stations study sites.
Station Par. Peeble (4-8 mm) Coarse sand (0.5 – 2.0 mm) Fine sand (0.125 – 0.25 mm) Mud (Residuals – 0.063 mm) West Pari-1 X Sd 1.01 0.47 46.47 2.82 29.88 1.40 22.63 4.50 West Pari-2 X Sd 0.78 1.10 45.87 15.97 25.51 1.92 27.85 13.68 North Pari X Sd 1.56 0.73 22.50 0.31 42.10 4.50 33.83 4.17
Kerapatan rata-rata lamun di stasiun penelitian berkisar 58,80 tunas.m-2 (E. acoroides di Pari Utara) sampai dengan 650,00 tunas.m-2 (T. hemprichii di Pari Barat). Kerapatan rata-rata lamun pada bulan April lebih tinggi daripada bulan Juli 2009 (Tabel 3). NIENHUIS et al. (1989) dalam penelitiannya di Laut Flores menemukan bahwa kerapatan tunas lamun per luasan area tergantung pada jenisnya. Jenis lamun yang mempunyai morfologi besar (E. acoroides) mempunyai kerapatan yang rendah (140 tunas.m-2) dibandingkan dengan jenis lamun yang mempunyai morfologi kecil (Halodule uninervis) dengan kerapatan yang tinggi (14.800 tunas.m-2).
Tabel 3: Kerapatan dan luas tutupan lamun di stasiun penelitian. Table 3: Density and percentage of coverage of seagrasses at study sites.
Station Species Par. April July Density (shoots/m2) Percentage of coverage (o/o) Density (shoots/m2) Percentage of coverage (o/o) West Pari -1 T. hemprichii C. rotundata X Sd X Sd 500.00 350.0 416.67 104.08 70,00 16.73 425.0 66.14 625.0 336.34 83.33 5.77 West Pari-2 T. hemprichii C. rotundata E. acoroides X Sd X Sd X Sd 650.00 377.49 366.67 175.59 99.73 3.11 78.00 10.95 550.0 25.0 466.67 137.69 82.27 16.77 80.00 10.00 North Pari E. acoroides X Sd 77.47 17.28 21.50 8.00 58.80 10.28 30.50 7.79
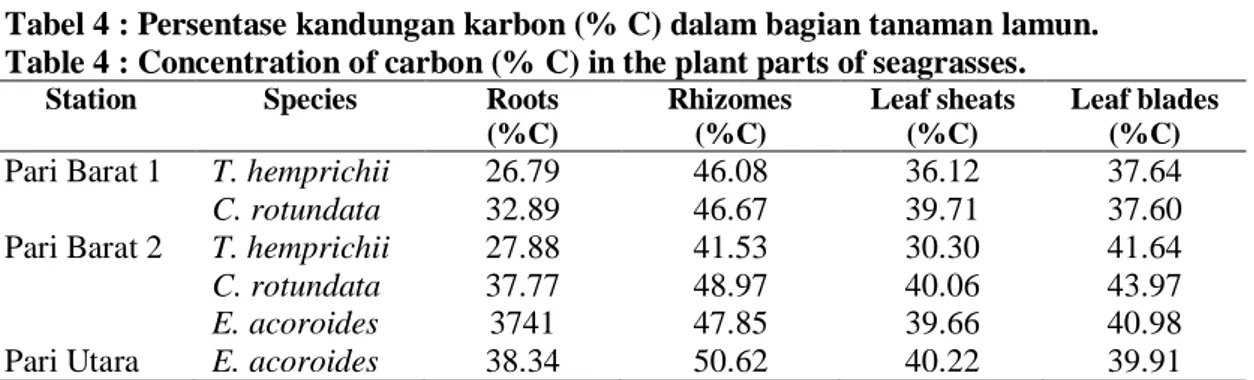
Persentase karbon (% C) bagian-bagian tanaman lamun (E. acoroides, C. rotundata dan T. hemprichii) dari sampel berat keringnya yang dianalisis di Laboratorium Departemen Ilmu Tanah dan Sumberdaya Lahan Fakultas Pertanian IPB disajikan dalam Tabel 4. Persentase kandungan karbon dalam bagian-bagian tanaman lamun yang tertinggi terdapat dalam rimpang yang berkisar antara 41,53 % (T. hemprichii) sampai 50,62 % (E. acoroides). Nilai % karbon dalam bagian tanaman lamun yang lebih rendah ditemukan oleh NIENHUIS et al. (1989) dimana akar dan rimpangnya tidak dibedakan dalam lamun yang ditelitinya di Laut Flores.
Tabel 4 : Persentase kandungan karbon (% C) dalam bagian tanaman lamun. Table 4 : Concentration of carbon (% C) in the plant parts of seagrasses.
Station Species Roots (%C) Rhizomes (%C) Leaf sheats (%C) Leaf blades (%C) Pari Barat 1 T. hemprichii
C. rotundata 26.79 32.89 46.08 46.67 36.12 39.71 37.64 37.60 Pari Barat 2 T. hemprichii
C. rotundata E. acoroides 27.88 37.77 3741 41.53 48.97 47.85 30.30 40.06 39.66 41.64 43.97 40.98
Pari Utara E. acoroides 38.34 50.62 40.22 39.91
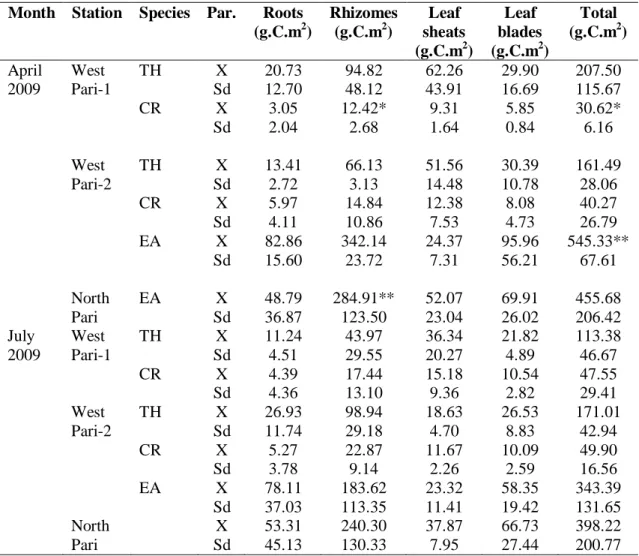
Biomasa bagian tanaman lamun dalam berat kering sampel (g.berat kering.m
-2 ) yang dikonversi dengan percentase nilai karbonnya (% C) (Tabel 4) menjadi gram C.m-2 tertera dalam Tabel 5. Biomasa lamun rata-rata dalam rimpang mempunyai nilai tertinggi dibandingkan dengan nilai dalam bagian tanaman lainnya, yaitu berkisar antara 12,42 gr.C.m -2(C. rotundata) sampai dengan 284,91 gr.C.m-2 (E. acoroides). Sementara itu biomasa total rata-ratanya berkisar antara 30,62 gr.C.m-2 pada C. rotundata di stasiun Pari Barat 1 dan 545,33 gr.C.m-2 pada E. acoroides di stasiun Pari Barat 2 (Tabel 5).
NIENHUIS et al. (1989) melaporkan bahwa biomasa lamun bagian bawah (akar dan rimpang) adalah 6 – 10 kali lebih besar dari pada biomasa bagian atasnya (seludang daun dan helaian daun). Mereka menemukan korelasi positip antara luas tutupan lamun dengan biomasanya. Sementara itu DE BOER (2000) menemukan perbedaan biomasa bagian bawah lamun Zostera capensis yang mencapai 91 %. Porsi biomasa lamun didapat pada rimpang yang berkisar antara 32,41 % (T. hemprichii) sampai 63,01 % (E. acoroides).
DE BOER (2000) melaporkan adanya variasi musiman pada morfologi, kerapatan dan biomas lamun dari hasil penelitiannya di Inhaca Island, Mozambique, dia membedakannya menurut musim panas dan musim dingin. Penelitian yang sama dilakukan di Pulau Pari, yaitu mengamati variasi musiman pada kerapatan dan biomas lamun berdasarkan perbedaan antara musim hujan dan musim kemarau serta surut terendah yang terjadi siang hari (terbakarnya daun).
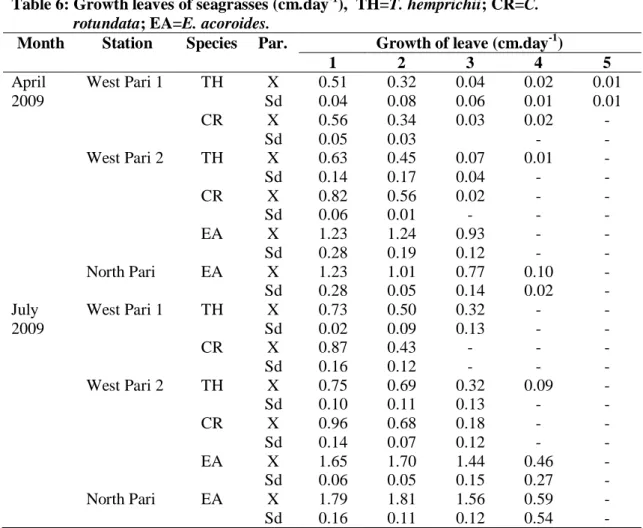
Laju pertumbuhan daun rata-rata berkisar antara 0,01 cm/hari (daun ke-5 T.
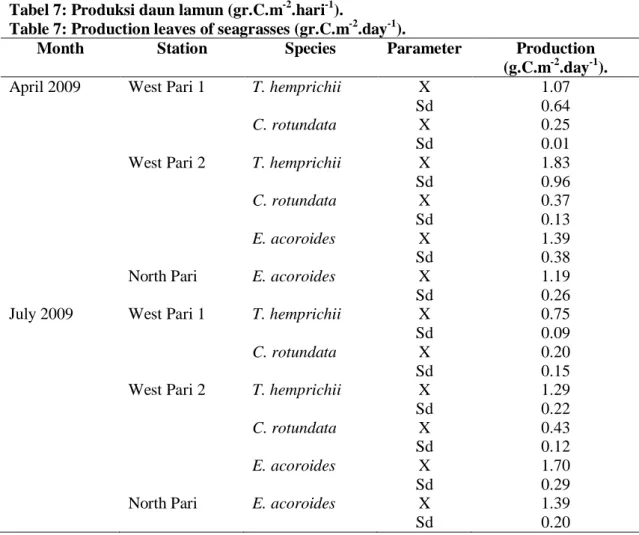
hemprichii) sampai 1,79 cm/hari (daun ke-1 E. acoroides) (Tabel 6). Laju pertumbuhan daun lamun yang tinggi pada daun muda banyak dilaporkan oleh peneliti lain seperti BROUNS (1985) yang melaporkan pertumbuhan T. hemprichii di Papua New Guinea, ESTACION & FORTES (1988) pada pertumbuhan E. acoroides di Lag-it, North Bais Bay-Philipina dan ERTEMEIJER (1993) pada C. rotundata, T. hemprichii dan E. acoroides di Kepulauan Spermonde, Sulawesi Selatan.Produksi daun lamun rata-rata berkisar antara 0,20 g.C.m
-2.h
-1(C. rotundata)
sampai 1,83 g.C.m
-2.h
-1(T. hemprichii) (Tabel 7). Produksi ini merupakan produksi
neto dari daun lamun sehingga dapat dikatagorikan sebagai laju kecepatan penyerapan
karbon oleh lamun, seperti yang dilakukan oleh ZIEMAN (1974) pada Thalassia
testudinum yang pertumbuhannya diukur dengan tehnik penandaan daun. Apabila
produksi neto daun lamun di Pulau Pari dihitung untuk satu tahun besarnya berkisar
antara 73 – 668 g.C.m
-2.th
-1.
Tanaman dapat menyerap CO2 dari atmosfer. Tanaman mampu menyimpan bagian karbon dari molekul CO2 untuk membangun strukturnya. Berat atom karbon (C) adalah 12, oksigen (O) adalah 16 sehingga bobot atom CO2 menjadi 44. Pengikatan satu ton karbon oleh tanaman dapat mengikat 44/12, atau 3,67 ton CO2 dari atmosfer dan melepaskan sebanyak 2,67 ton O2 ke atmosfer (www.olivotto.com/carbon/index.html). Produksi lamun di Pulau Pari yang berkisar antara 73 – 668 g.C.m-2.th-1 berarti sama dengan menyerap CO2 dari atmosfer sebesar 267,67 – 2.449,33 g.CO2. m
-2
.th-1. Hasil penelitian ini menunjukkan bahwa penyerapan karbon oleh lamun di Pulau Pari ternyata lebih tinggi daripada yang dilaporkan oleh KALDY et al. (2002) yang mendapatkan nilai produksi jenis-jenis lamun Halodule
wrightii (47 – 246 g C th-1), Syringodium filiforme (15 – 101 g C th-1) dan Thalassia testidinum (60 – 342 g C th-1) di Lower Laguna Madre, Amerika.
Tabel 5: Biomas lamun (g.C.m2).
Table 5: Biomass of seagrasses (g.C.m2). Month Station Species Par. Roots
(g.C.m2) Rhizomes (g.C.m2) Leaf sheats (g.C.m2) Leaf blades (g.C.m2) Total (g.C.m2) April 2009 West Pari-1 TH CR X Sd X Sd 20.73 12.70 3.05 2.04 94.82 48.12 12.42* 2.68 62.26 43.91 9.31 1.64 29.90 16.69 5.85 0.84 207.50 115.67 30.62* 6.16 West Pari-2 TH CR EA X Sd X Sd X Sd 13.41 2.72 5.97 4.11 82.86 15.60 66.13 3.13 14.84 10.86 342.14 23.72 51.56 14.48 12.38 7.53 24.37 7.31 30.39 10.78 8.08 4.73 95.96 56.21 161.49 28.06 40.27 26.79 545.33** 67.61 North Pari EA X Sd 48.79 36.87 284.91** 123.50 52.07 23.04 69.91 26.02 455.68 206.42 July 2009 West Pari-1 TH CR X Sd X Sd 11.24 4.51 4.39 4.36 43.97 29.55 17.44 13.10 36.34 20.27 15.18 9.36 21.82 4.89 10.54 2.82 113.38 46.67 47.55 29.41 West Pari-2 TH CR EA X Sd X Sd X Sd 26.93 11.74 5.27 3.78 78.11 37.03 98.94 29.18 22.87 9.14 183.62 113.35 18.63 4.70 11.67 2.26 23.32 11.41 26.53 8.83 10.09 2.59 58.35 19.42 171.01 42.94 49.90 16.56 343.39 131.65 North Pari X Sd 53.31 45.13 240.30 130.33 37.87 7.95 66.73 27.44 398.22 200.77
Catatan : (n=3, TH = T. hemprichii; CR= C. rotundata; EA= E. acoroides).
* = Nilai terendah, ** = Nilai tertinggi
Note : (n=3, TH = = T. hemprichii; CR= C. rotundata; EA= E. Acoroides. * = Lowest value, ** = Highest value
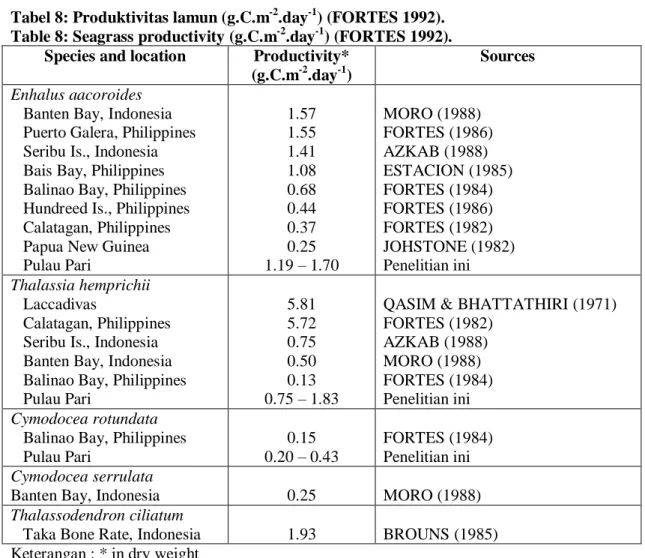
Produksi lamun di Pulau Pari mempunyai nilai yang berada dalam kisaran yang dilaporkan FORTES (1992) untuk E. acoroides dan T. hemprichii, tetapi untuk C. rotundata mempunyai nilai yang lebih tinggi. FORTES (1992) menemukan bahwa bobot produksi lamun dalam karbon (gr.C.m-2.day-1) dengan mengkonversi nilai produktivitas lamun dalam bobot kering (gr.DW.m-2.day-1) dengan faktor 0,88 dan kemudian dengan faktor 0,47 menurut WESTLAKE (1963) (Tabel 8).
Persentase karbon (% C) dalam serasah yang
masih segar dan melayang, serasah yang terdapat di atas substrat yang sudah mulai
lapuk dan dalam sedimen hasil analisis tertera dalam Tabel 9.
Tabel 6: Pertumbuhan daun lamun (cm/hari), TH=T. hemprichii; CR=C. rotundata; EA=E. acoroides.
Table 6: Growth leaves of seagrasses (cm.day-1), TH=T. hemprichii; CR=C. rotundata; EA=E. acoroides.
Month Station Species Par. Growth of leave (cm.day-1)
1 2 3 4 5 April 2009 West Pari 1 TH CR X Sd X Sd 0.51 0.04 0.56 0.05 0.32 0.08 0.34 0.03 0.04 0.06 0.03 0.02 0.01 0.02 - 0.01 0.01 - - West Pari 2 TH CR EA X Sd X Sd X Sd 0.63 0.14 0.82 0.06 1.23 0.28 0.45 0.17 0.56 0.01 1.24 0.19 0.07 0.04 0.02 - 0.93 0.12 0.01 - - - - - - - - - - - North Pari EA X Sd 1.23 0.28 1.01 0.05 0.77 0.14 0.10 0.02 - - July 2009 West Pari 1 TH CR X Sd X Sd 0.73 0.02 0.87 0.16 0.50 0.09 0.43 0.12 0.32 0.13 - - - - - - - - - - West Pari 2 TH CR EA X Sd X Sd X Sd 0.75 0.10 0.96 0.14 1.65 0.06 0.69 0.11 0.68 0.07 1.70 0.05 0.32 0.13 0.18 0.12 1.44 0.15 0.09 - - - 0.46 0.27 - - - - - - North Pari EA X Sd 1.79 0.16 1.81 0.11 1.56 0.12 0.59 0.54 - -
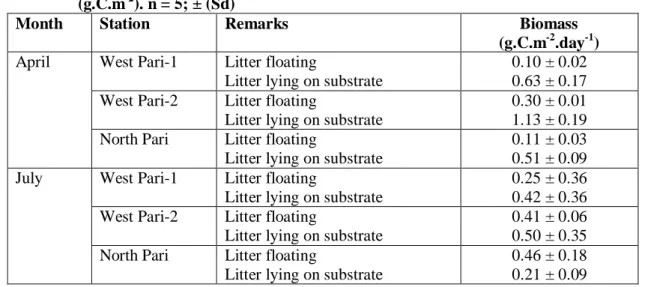
Biomasa serasah (g.bobot kering.m-2) yang dikonversi dengan percentase nilai karbonnya (% C) (Tabel 9) menjadi gram C.m-2 disajikan pada Tabel 10. Biomasa serasah dalam g.C.m-2 hasil konversi dari berat keringnya dengan % C hasil analisa bagian atas (melayang) yang berkisar antara 32,10 – 45,15 % C yang merupakan serasah yang masih segar yang lepas langsung dari lamun. Biomasa serasah di dasar yang berkisar antara 27,81 – 36,56 % C yang merupakan serasah yang sudah lebih lapuk dan yang terdapat pada substrat yang berkisar antara 0,37 – 0,41 % C menunjukkan kecenderungan bahwa karbon hasil produksi daun lamun lebih banyak diekspor ke sistem lain. Hal yang sama dilaporkan oleh PERGENT et al. (1994) yang menunjukkan bahwa sebanyak 29 % materi hasil produksi dari lamun Posidonia oceanica di Laut Mediterania diekspor ke sistem lain.
Penelitian ini baru menghitung produksi harian dari tiga jenis lamun yang ada
di Pulau Pari. Untuk mendapatkan informasi yang lengkap tentang karbon rosot dan
penyerapannya di Pulau Pari, masih diperlukan penelitian lebih lanjut untuk
mengukur produksi harian jenis-jenis lamun lainnya seperti C. serrulata, H. ovalis, H.
uninervis dan S. isoetifolium serta pemetaan sebarannya untuk mengukur luasnya.
Data tersebut digabung dengan hasil pemetaan dapat dipakai untuk menghitung
karbon rosot dan penyerapannya oleh padang lamun di Pulau Pari. Hasil penelitian ini
dapat dipakai sebagai dasar penghitungan karbon rosot dan penyerapannya di padang
lamun berbagai lokasi di perairan Indonesia.
Tabel 7: Produksi daun lamun (gr.C.m-2.hari-1).
Table 7: Production leaves of seagrasses (gr.C.m-2.day-1).
Month Station Species Parameter Production
(g.C.m-2.day-1). April 2009 West Pari 1 T. hemprichii
C. rotundata X Sd X Sd 1.07 0.64 0.25 0.01 West Pari 2 T. hemprichii
C. rotundata E. acoroides X Sd X Sd X Sd 1.83 0.96 0.37 0.13 1.39 0.38
North Pari E. acoroides X
Sd
1.19 0.26 July 2009 West Pari 1 T. hemprichii
C. rotundata X Sd X Sd 0.75 0.09 0.20 0.15 West Pari 2 T. hemprichii
C. rotundata E. acoroides X Sd X Sd X Sd 1.29 0.22 0.43 0.12 1.70 0.29
North Pari E. acoroides X
Sd
1.39 0.20
Tabel 8: Produktivitas lamun (g.C.m-2.day-1) (FORTES 1992). Table 8: Seagrass productivity (g.C.m-2.day-1) (FORTES 1992).
Species and location Productivity* (g.C.m-2.day-1)
Sources
Enhalus aacoroides Banten Bay, Indonesia Puerto Galera, Philippines Seribu Is., Indonesia Bais Bay, Philippines Balinao Bay, Philippines Hundreed Is., Philippines Calatagan, Philippines Papua New Guinea Pulau Pari 1.57 1.55 1.41 1.08 0.68 0.44 0.37 0.25 1.19 – 1.70 MORO (1988) FORTES (1986) AZKAB (1988) ESTACION (1985) FORTES (1984) FORTES (1986) FORTES (1982) JOHSTONE (1982) Penelitian ini Thalassia hemprichii Laccadivas Calatagan, Philippines Seribu Is., Indonesia Banten Bay, Indonesia Balinao Bay, Philippines Pulau Pari 5.81 5.72 0.75 0.50 0.13 0.75 – 1.83
QASIM & BHATTATHIRI (1971) FORTES (1982) AZKAB (1988) MORO (1988) FORTES (1984) Penelitian ini Cymodocea rotundata
Balinao Bay, Philippines Pulau Pari 0.15 0.20 – 0.43 FORTES (1984) Penelitian ini Cymodocea serrulata
Banten Bay, Indonesia 0.25 MORO (1988)
Thalassodendron ciliatum
Taka Bone Rate, Indonesia 1.93 BROUNS (1985) Keterangan : * in dry weight
Tabel 9: Persentase kandungan karbon (% C) dalam serasah dan sedimen dasar. Table 9: Percentage of carbon content in the litter fall and bottom sediment (% C).
Station Remarks (% C)
West Pari Litter fall
Floating
Lying on substrate
39.68 – 45.15 27.81 – 36.56
North Pari Litter fall
Floating
Lying on substrate
32.10 30.27
Tabel 10 : Biomas serasah daun lamun yang melayang dan di dasar (g.C.m-2). n = 5; ± (Sd)
Table 10 : Biomass of litter fall leaves of seagrasses floating and on the substrate (g.C.m-2). n = 5; ± (Sd)
Month Station Remarks Biomass
(g.C.m-2.day-1) April West Pari-1 Litter floating
Litter lying on substrate
0.10 ± 0.02 0.63 ± 0.17 West Pari-2 Litter floating
Litter lying on substrate
0.30 ± 0.01 1.13 ± 0.19 North Pari Litter floating
Litter lying on substrate
0.11 ± 0.03 0.51 ± 0.09 July West Pari-1 Litter floating
Litter lying on substrate
0.25 ± 0.36 0.42 ± 0.36 West Pari-2 Litter floating
Litter lying on substrate
0.41 ± 0.06 0.50 ± 0.35 North Pari Litter floating
Litter lying on substrate
0.46 ± 0.18 0.21 ± 0.09
KESIMPULAN
Kerapatan rata-rata lamun berkisar antara 58,80 tunas.m-2 (E. acoroides) sampai 650,00 tunas.m-2 (T. hemprichii). Luas tutupan rata-rata lamun di Barat Pulau Pari berkisar antara 70 – 83,33 % dan di Utara Pulau Pari berkisar antara 21,5 – 30 %. Persentase karbon pada bagian tanaman lamun tertinggi didapat pada rimpang yang berkisar antara 41,53 – 50,63 %. Biomas total rata-rata lamun berkisar antara 30,62 g.C.m-2 (C. rotundata) sampai 545,33 g.C.m-2 (E. acoroides). Kecepatan pertumbuhan daun rata-rata berkisar antara 0,01 cm/hari (daun ke-5 T. hemprichii) sampai 1,79 cm/hari (daun ke-1 E. acoroides). Produksi karbon oleh daun rata-rata berkisar antara 0,20 g.C.m-2.h-1 (C. rotundata) sampai 1,83 g.C.m -2
.h-1 (E. acoroides). Biomass serasah bagian atas (melayang) berkisar antara 0,10 – 0,46 g.C.m-2 dan biomas serasah di dasar berkisar antara 0,21 – 1,13 g.C.m-2. Karbon rosot di padang lamun Pulau Pari berkisar antara 30,63 – 545,33 g.C.m-2 dan penyerapan karbon berkisar antara 0,20 – 1,83 g.C.m-2.h-1.
Penelitian ini baru menghitung produksi harian dari 3 jenis lamun yang ada di Pulau Pari. Untuk mendapatkan informasi yang lengkap tentang karbon rosot dan penyerapannya di Pulau Pari, masih diperlukan penelitian lebih lanjut untuk mengukur produksi harian jenis-jenis lamun lainnya a.l. C. serrulata, H. ovalis, H. uninervis dan S. isoetifolium serta pemetaan sebarannya untuk mengukur luasnya. Data tersebut digabung dengan hasil pemetaan dapat dipakai untuk menghitung karbon rosot dan penyerapannya oleh padang lamun di Pulau Pari. Hasil penelitian ini dapat dipakai sebagai dasar penghitungan karbon rosot dan penyerapannya di padang lamun berbagai lokasi di perairan Indonesia.
PERSANTUNAN
Dengan selesainya penelitian penyimpanan dan penyerapan karbon oleh beberapa jenis
lamun di Pulau Pari, diucapkan terima kasih kepada Sdr. Achmad Arifin, Muhayar dan Imam yang telah membantu pengamatan lapangan dan pengolahan sampel sehingga datanya siap untuk dianalisa.DAFTAR PUSTAKA
ALONGI, D.M. 2002. Present state and future of the world’s mangrove forest. Environ. Conser. 29: 331 -349.
AZKAB, M.H. 1988. Study on seagrass community structure and biomass in the southern part of Seribu Islands. Paper presented at the First Regional Symposium of the ASEAN-Australia Coastal Living Resources Project. Manila, Philippines, 30 Jan. – 1 Feb. 1988. 353-362.
BARRÓN, C., N. MARBÁ, J. TERRADOS, H. KENNEDY and C.M. DUARTE 2004. Community metabolism and carbon budget along a gradient of seagrass (Cymodoea
nodosa) colonizaation. Limnol. Oceanogr. 49(5): 1642-1651.
BROUNS, J.J.W.M. 1985. A comparison of the annual production and biomass in three monospecific stands of the seagrass Thalassia hemprichii (Ehrenb.) Aschers. Aquat. Bot. 23: 149-175.
DE BOER, W.F. 2000. Biomass dynamics of seagrasses and the role of mangrove and seagrass vegetation as different nutrient sources for an intertidal ecosystems. Aquat. Bot. 66: 225-239.
DE IONGH, H.H. 1996. Plant-herbivore interactions between seagrasses and dugongs in a tropical small ecosystem. Katholieke Universiteit Nijmegen, Thesis PhD, 205 pp. DUARTE, C. M. and J. CEBRIAN 1996. The fate of marine autotroph production. Limnol.
Oceanogr. 41: 1758 – 1766.
DUARTE, C.M. and M.H. HEMMINGA 2000. Seagrass ecology. Cambridge Univ. Press: 251 pp.
DUARTE, C.M.; J.J. MIDDELBURG and C. CARACO 2005. Major role of marine vegetation on the oceanic carbon cycle. Biogeoscience 2: 1-8.
ERFTEMEIJER, P.L.A. 1993. Factors limiting growth and reproduction of tropical seagrasses: nutrients dynamics in Indonesian seagrass beds. PhD Thesis. Catholic Nijmegen University. 173 pp.
ESTACION, J.S. 1985. Growth rates and organic matter production of the seagrass, Enhalus acoroides (L. f.) Royle in Lag-it, North Bais Bay. Master Thesis, University of the Philippines, Diliman, Quezon City, 78 pp.
ESTACION, J.E. and M.D. FORTES 1988. Growth rates and primary production of Enhalus acoroides (L.f.) Royle from Lag-it, North Bais Bay, The Phillippines. Aquat. Bot, 29: 347-356.
FORTES, M.D. 1982. Mangrove, seagrass, and algal productivity at Calatagan, Batangas, Philippines. Biotrop Spec. Publ. 17: 17-24.
FORTES, M.D. 1984. Ecological assessment and cultivation of seagrasses at Balinao Bay for biomass production. NCRP Res. Publ. 39(1): 60-112.
FORTES, M.D. 1986. Taxonomy and ecology of Philippines seagrasses. PhD Dissertation, University of the Philippines, Diliman, Quezon City, 254 pp.
FORTES, M.D. 1992. Comparative study of structure and productivity of seagrass communities in the ASEAN Region. In: CHOU, L.M. and C.R. WILKINSON (eds.) Third ASEAN Science and Technology Week Conference Proceeding, Vol. 6 Marine Science: Living Coastal Resources, 21-23 Sept. 1992, Singapore Dept. of Zoology, National University of Singapore and National Science and Technology Board, Singapore: 223-228.
JOHNSTONE, I.M. 1982. Ecology and distribution of seagrasses. In: J.L. GRESSIT (ed.) Biogeography and Ecology of New Guinea. Monographiae Biologicae 42.
KALDY, J.E., C.P. ONUF, P.M. ELDRIDGE and L.A. CIFUANTES 2002. Carbon budget for a substropical seagrass dominated coastal lagoon: How importance are seagrasses to total ecosystem net primary production. Estuaries 25(4A): 528-539.
MIDDELBURG, J.J., C.M. DUARTE and R.K. GATTUSO 2004. Respiration in coastal benthic communities. In: DEL GIORGIO, P.A. and P.J. LE B. WILLIAMS (eds.) Respiration in aquatic ecosystems. Oxford Univ. Press.
MORO, D.S. 1988. Pertumbuhan dan produktivitas beberapa jenis lamun di Pulau Panjang, Teluk Banten. Skripsi, Fakultas Biologi, Universitas Nasional Jakarta. 75 pp.
NIENHUIS, P.H., J. COOSEN and W. KISWARA 1989. Community stucture and biomass distribution of seagrasses and macrofauna in the Flores Sea, Indonesia. Neth. J. of Sea Res. 23(2): 197-214.
NOJIMA, S. and H. MUKAI 1996. The rate and fate of production of seagarass debris in cages over Syringodium isoetofolium (Aschers) Dandy meadows. In: KUO, J.; R.C. Phillips; D.I. WALKER and H. KIRKMAN (eds.) Proc. Intern. Seagrass Biol. Workshop. Rottnest Island, Western Australia 25-29 January 1996: 149-154.
PERGENT, G., J. ROMERO, C. PERGENT-MARTINI, M.A. MATEO and C.F. BOUDOURESQUE 1994. Primary production, stocks and fluxes in the Mediteranian seagrass Posidonia oceanica. Mar. Ecol. Prog. Ser. 106: 139-146.
QASIM, S.Z. and P.M.A. BHATTATHIRI 1971. Primary production of a seagrass bed on Kararatti Atoll (Laccadives). Hydrobiologia 38(1): 24-38.
SCHUMACHER, B.A. 2002. Methods for the determination of total organic carbon (TOC) in soils and sediments. United States Environment Protection Agency. Environmental Science Division National. 25 pp.
TOMLINSON, P.B. 1994. The botany of mangroves. Cambridge Univ. Press. 413 pp. WESTLAKE, D.F. 1963. Comparisons of plant productivity. Biol. Rev. 38: 385-425.
ZIEMAN, J.C. 1974. Methods for the study of the growth of turtle grass Thalassia testudinum König. Aquaculture 4: 139-143.