Trialeurodes vaporariorum WESTWOOD (HEMIPTERA:
ALEYRODIDAE) PADA TANAMAN TOMAT (Lycopersicum
esculentum MILL.): PANJANG ROSTRUM DAN SAYAP DI
BEBERAPA KETINGGIAN TEMPAT SERTA PERIODE
RETENSI Tomato infectious chlorosis virus (TICV)
RR. LARAS ANJARSARI
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
2011
ABSTRAK
RR. LARAS ANJARSARI. Trialeurodes vaporariorum Westwood (Hemiptera: Aleyrodidae) pada Tanaman Tomat (Lycopersicum esculentum Mill.): Panjang
Rostrum dan Sayap di Beberapa Ketinggian Tempat serta Periode Retensi Tomato
infectious chlorosis virus (TICV). Dibimbing oleh DEWI SARTIAMI dan
GEDE SUASTIKA.
Penelitian ini bertujuan untuk mengetahui variasi panjang rostrum dan sayap kutukebul T. vaporariorum pada beberapa ketinggian tempat serta lama waktu retensi TICV dalam tubuh serangga vektor T. vaporariorum. Sampel kutukebul diambil dari sentra produksi tomat di Kecamatan Cikajang, Kabupaten Garut (1287 mdpl, dengan suhu rata-rata 19,8 ˚C), Kecamatan Pacet, Kabupaten Cianjur (1125 mdpl, 20,93 ˚C), Kecamatan Cikole, Kabupaten Sukabumi (1022 mdpl, 22,22 ˚C), dan Kecamatan Batu, Kotamadya Batu, Malang (675 mdpl, 22,43 ˚C). Pengamatan variasi kutukebul T. vaporariorum dilakukan dengan mengukur panjang rostrum dan sayap 30 ekor imago betina kutukebul dari masing-masing lokasi. Data yang diperoleh kemudian diuji menggunakan uji t sampel bebas dengan program Microsoft Excel. Penelitian periode retensi TICV
dilakukan dengan melakukan inokulasi berseri menggunakan imago
T. vaporariorum yang viruliferus (mengandung virus) ke bibit tanaman tomat
sehat (sebagai tanaman uji) sampai pemindahan pada tanaman ke-7. Lama periode akuisisi dan inokulasi masing-masing 24 jam, dengan 10 ulangan.
Panjang rostrum rata-rata imago betina T. vaporariorum yang berasal dari Cikajang adalah 293,18 µm, dari Pacet 275,76 µm, dari Cikole 256,36 µm, dan dari Batu adalah 251,81 µm. Panjang sayap rata-rata imago yang berasal dari Cikajang, Pacet, Cikole, dan Batu berturut-turut adalah 1286,33 µm, 1212 µm, 1162,67 µm, dan 1119,33 µm. Panjang rostrum dan sayap imago betina
T. vaporariorum berbeda nyata antar masing-masing lokasi, kecuali panjang
rostrum imago betina dari Cikole dan Batu yang tidak berbeda nyata. Ukuran panjang rostrum dan sayap kutukebul mengalami pertambahan seiring dengan pertambahan ketinggian tempat dan penurunan suhu habitat kutukebul. Rata-rata periode retensi TICV dalam tubuh serangga vektor T. vaporariorum adalah selama tiga hari. Hal tersebut menunjukkan bahwa periode retensinya relatif lama, karena satu ekor imago mampu menularkan virus TICV selama tiga hari.
Kata kunci: Trialeurodes vaporariorum, rostrum, sayap, periode retensi,
Trialeurodes vaporariorum WESTWOOD (HEMIPTERA:
ALEYRODIDAE) PADA TANAMAN TOMAT (Lycopersicum
esculentum MILL.): PANJANG ROSTRUM DAN SAYAP DI
BEBERAPA KETINGGIAN TEMPAT SERTA PERIODE
RETENSI Tomato infectious chlorosis virus (TICV)
RR. LARAS ANJARSARI
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian di Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
2011
Judul Penelitian : Trialeurodes vaporariorum Westwood (Hemiptera: Aleyrodidae) pada Tanaman Tomat (Lycopersicum
esculentum Mill.): Panjang Rostrum dan Sayap di Beberapa
Ketinggian Tempat serta Periode Retensi Tomato infectious
chlorosis virus (TICV)
Nama Mahasiswa : Rr. Laras Anjarsari
NRP : A34062876
Program Studi : Proteksi Tanaman
Disetujui,
Pembimbing I Pembimbing II
Dra. Dewi Sartiami, M.Si Dr.Ir. Gede Suastika, M.Sc NIP. 19641204 199103 2 001 NIP. 19620607 198703 1 003 Mengetahui, Ketua Departemen Dr.Ir. Dadang, M.Sc NIP. 19640204 199002 1 002 Tanggal Lulus :
RIWAYAT HIDUP
Penulis dilahirkan dari pasangan Bapak R. Dwiono Rahardjo dan Ibu Effy Kurniati di Jombang, Jawa Timur pada tanggal 29 September 1988 sebagai anak kedua dari tiga bersaudara.
Penulis menempuh pendidikan dasar di SDN Panggang I Jepara, SDN Kraton I Pekalongan, dan terakhir di SDN Sale I Rembang pada tahun 2000. Pada tahun 2000-2001 penulis menempuh pendidikan di SLTPN Jatirogo I Tuban dan menyelesaikan sisanya di SMPN 2 Jombang pada tahun 2003. Pada tahun tersebut penulis melanjutkan pendidikan di SMAN 2 Jombang sampai lulus pada tahun 2006. Selama SMP sampai SMA penulis aktif dalam kegiatan ekstrakurikuler Pramuka, tari, renang, dan PMR. Pada tahun yang sama penulis melanjutkan pendidikan di Institut Pertanian Bogor (IPB) melalui Undangan Seleksi Masuk IPB (USMI) dan diterima di Departemen Proteksi Tanaman pada tahun selanjutnya.
Selama menjadi mahasiswa penulis aktif di kegiatan kemahasiswaan, di antaranya sebagai pengurus Himpunan Mahasiswa Proteksi Tanaman (HIMASITA) Biro Keprofesian periode 2007/2008 serta Biro Ekonomi dan Bisnis periode 2008/2009. Penulis juga aktif di organisasi mahasiswa daerah (OMDA) JAC (Jombang Agrostudent Community) sebagai bendahara. Penulis pernah mengikuti kepanitiaan dalam Seminar Nasional Pertanian dan Peternakan Jombang tahun 2007, Musyawarah Nasional Himpunan Mahasiswa Proteksi Tanaman Indonesia (HMPTI) pada tahun 2008, Seminar Aku dan Serangga, dan Lokakarya Nasional Wereng Cokelat pada tahun 2010. Penulis juga aktif dalam bidang olahraga dan klub kemahasiswaan seperti Organic Farming (OF), Entomologi Club, dan klub fotografi Capung. Penulis tergabung menjadi anggota PKKM IPB pada 2010.
Penulis menjadi asisten mata kuliah Entomologi Umum pada tahun 2009 dan asisten mata kuliah Dasar-dasar Proteksi Tanaman tahun 2010.
PRAKATA
Puji syukur kepada Allah Swt., atas rahmat dan karunia-Nya penulis dapat menyelesaikan penelitian dan penulisan skripsi yang berjudul “Trialeurodes
vaporariorum Westwood (Hemiptera: Aleyrodidae) pada Tanaman Tomat
(Lycopersicum esculentum Mill.): Panjang Rostrum dan Sayap di Beberapa Ketinggian Tempat Berbeda serta Periode Retensi Tomato infectious chlorosis
virus (TICV)”. Sebagian dari penelitian ini dibiayai dari kerjasama dengan
Utsunomiya University, Japan melalui Dr. Ir. Gede Suastika, M.Sc.
Penulis mengucapkan rasa terima kasih setulusnya kepada ibu Dra. Dewi Sartiami, M.Si dan bapak Dr. Ir. Gede Suastika, M.Sc selaku pembimbing yang telah memberi perhatian, arahan, bimbingan, masukan, saran, serta koreksi dalam penulisan skripsi ini. Terima kasih kepada ibu Ir. Titiek Siti Yuliani, SU. selaku penguji sekaligus pembimbing akademik yang telah bersedia menguji, memberi masukan untuk perbaikan skripsi ini, serta nasihat sehari-harinya. Penulis merasa sangat beruntung bisa memperoleh tambahan ilmu pengetahuan dari ibu dan bapak dosen. Bak pelita yang menjadi penerang dalam gulita, jasamu tiada tara.
Penulis juga mengucapkan terima kasih kepada teman-teman yang telah membantu terlaksananya penelitian ini: Satrio Harjono, Sari Nurulita, Amelia Andriani, Herlie, Rifqi, Septa, Mba Tuti Legiastuti, Ibu Rika, Ibu Aisyah, mbak Elsa, mbak Lia, mbak Rika, mbak Atik, Fitrah M, dan Gilang. Terima kasih atas dukungan dan jalinan persaudaraan teman-teman angkatan 43, kakak kelas angkatan 42, adik kelas angkatan 44 dan 45, serta keluarga besar Departemen Proteksi Tanaman yang hangat dan bersahabat. Terima kasih pula kepada petani tomat di Garut (Pak H. Oman), Cianjur (Pak Ateng), Sukabumi (Pak Ajum), dan Batu, yang telah membantu proses pengerjaan penelitian di lapangan. Kepada warga Desa Cibodas, Kecamatan Cikajang, Garut, penulis berterimakasih atas keramahan dan bantuan selama penelitian di sana.
Tak lupa penulis mengucapkan terima kasih yang sebesar-besarnya kepada yang tersayang, orangtua: Ir. R. H. Dwiono Rahardjo dan Drg. H. Effy Kurniati; kakak: dr. R. Bagas Widhiarso; serta adikku: Rr. Niken Ambarsari, Yang Ti, Yang Santo, tante Liliek, keluarga besar di Jombang, Jakarta, Bogor, Jogja, Gresik, dan Kolaka yang selalu memberi nasihat, motivasi, dukungan, serta doanya kepada penulis. Keluarga adalah mutiara tiada tara, untuk merekalah penelitian ini dipersembahkan.
Akhir kata, penulis berharap semoga penelitian ini dapat bermanfaat bagi pembaca, petani, pemerhati bidang pertanian, dan menjadi masukan bagi perkembangan ilmu pengetahuan. Amin.
Bogor, 22 Januari 2011
DAFTAR ISI
Halaman
DAFTAR TABEL ... viii
DAFTAR GAMBAR ... ix
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan Penelitian ... 3
TINJAUAN PUSTAKA ... 4
Taksonomi dan Morfologi Trialeurodes vaporariorum (Westwood) 4 Pengaruh Ketinggian dan Suhu terhadap Serangga ... 7
Tomato infectious chlorosis virus (TICV) ... 9
Hubungan Virus dengan Serangga Vektor serta Periode Retensi ... 10
BAHAN DAN METODE ... 11
Tempat dan Waktu Penelitian ... 11
Metode Penelitian ... 11
Pengambilan Sampel Kutukebul dan Tanaman Tomat Sumber TICV ... 11
Pembuatan Preparat dan Identifikasi T. vaporariorum ... 11
Pengukuran Panjang Rostrum dan Sayap T. vaporariorum ... 12
Pengumpulan Data Cuaca ... 13
Pengukuran Periode Retensi TICV dalam T. vaporariorum ... 13
HASIL DAN PEMBAHASAN ... 16
Identitas T. vaporariorum ... 16
Panjang Rostrum dan Sayap T. vaporariorum ... 17
Periode Retensi TICV dalam Tubuh T. vaporariorum ... 19
KESIMPULAN DAN SARAN ... 23
Kesimpulan ... 23
Saran ... 23
DAFTAR PUSTAKA ... 24
DAFTAR TABEL
Halaman
1. Panjang rostrum dan sayap T. vaporariorum ... 18 2. Masa infektif T. vaporariorum ... 21DAFTAR GAMBAR
Halaman
1. Kutukebul T. vaporariorum ... 5
2. Pengukuran T. vaporariorum ... 13
3. Bibit tomat yang sudah diinokulasi TICV ... 14
4. Pupa T. vaporariorum ... 16
5. Morfologi imago T. vaporariorum ... 17
6. Daun tanaman tomat uji ... 20
7. Hasil amplifikasi DNA genom virus dengan metode RT-PCR menggunakan pasangan primer spesifik TICV-CF dan TICV-CR ... 22
DAFTAR LAMPIRAN
Halaman 1. Panjang rostrum dan panjang sayap T. vaporariorum dari Kecamatan
Cikajang, Kabupaten Garut ... 28 2. Panjang rostrum dan panjang sayap T. vaporariorum dari Kecamatan
Pacet, Kabupaten Cianjur ... 29 3. Panjang rostrum dan panjang sayap T. vaporariorum dari Kecamatan
Cikole, Kabupaten Sukabumi ... 30 4. Panjang rostrum dan panjang sayap T. vaporariorum dari Kecamatan
Batu, Kotamadya Batu ... 31 5. Data suhu bulanan tahun 2010 ... 32 6. Data kelembaban bulanan tahun 2010 ... 32
PENDAHULUAN
Latar Belakang
Kutukebul adalah kelompok serangga berukuran kecil yang tubuhnya lunak dan berwarna kekuningan. Kutukebul termasuk dalam ordo Hemiptera, subordo Sternorrhyncha, superfamili Aleyrodoidea, famili Aleyrodidae, dan subfamili Aleurodicinae (Martin 1987). Kutukebul menjadi masalah bagi petani di seluruh belahan dunia. Kutukebul Trialeurodes vaporariorum (Westwood) pertama kali dilaporkan di Inggris pada tahun 1856, kemudian dilaporkan di Amerika pada tahun 1870 (CABI 2005). Semua stadia kutukebul ini hidup dan makan di permukaan bawah daun, namun stadia yang merusak adalah nimfa dan imago (Morales 2001).
Semua stadia T. vaporariorum memiliki kemampuan beradaptasi secara fisiologi di suhu dingin dibandingkan kutukebul Bemisia tabaci (Xie et al. 2006). Spesies B. tabaci dan T. vaporariorum juga berbeda secara morfologi. Bentuk sayap imago T. vaporariorum menyerupai tenda segitiga dan menutupi hampir seluruh abdomennya (Smith 2009). Variasi fisiologi dan morfologi pada serangga selain dipengaruhi oleh faktor genetik, juga dipengaruhi oleh posisi dan ketinggian tempat (Oliveira et al. 2004), serta tingkat radiasi dan suhu (Digby 1954). Menurut Roermund & Lenteren (1992), suhu lingkungan dapat mempengaruhi parameter kehidupan T. vaporariorum, seperti tingkat perkembangan nimfa, kematian nimfa, seks rasio, lama hidup, periode preoviposisi, fekunditas, frekuensi oviposisi, serta peningkatan periode oviposisi harian. Murai & Toda (2002) menyatakan bahwa individu Thrips tabaci yang hidup pada suhu yang lebih rendah mempunyai warna tubuh yang lebih gelap dan ukuran tubuh yang lebih besar daripada suhu yang lebih tinggi. Suhu di negara tropis seperti Indonesia sangat dipengaruhi oleh ketinggian tempat dan kelembaban udara (Wisnubroto et al. 1986). Namun sampai saat ini belum ada laporan mengenai pengaruh ketinggian terhadap morfologi atau variasi ukuran kutukebul.
Kutukebul T. vaporariorum bersifat polifag, baik pada tanaman pangan, sayuran maupun buah-buahan (CABI 2005). Kutukebul ini dilaporkan
mempunyai distribusi yang luas dan dapat ditemukan hampir di seluruh belahan dunia beriklim tropis dan subtropis. Di Indonesia T. vaporariorum dapat ditemukan di sentra pertanian tomat (Lycopersicum esculentum Mill.) provinsi Jawa Barat dan Jawa Tengah (Hartono & Wijonarko 2007). Fitriasari (2010) mengemukakan bahwa pada umumnya distribusi kutukebul T. vaporariorum berada pada lahan dataran tinggi. Distribusi kutukebul dipengaruhi oleh beberapa faktor, salah satunya adalah kemampuan serangga tersebut untuk terbang dengan sayapnya.
Sayap dan rostrum merupakan anggota tubuh yang dimiliki
T. vaporariorum dan anggota Aleyrodidae lainnya. Rostrum merupakan bentuk
alat mulut menusuk-menghisap yang terdiri dari empat struktur tubular yang dinamakan stilet. Hama ini dapat menimbulkan kerusakan baik secara langsung maupun tidak langsung pada tanaman tomat dengan rostrumnya. Kerusakan langsung yang ditimbulkan yaitu dengan menghisap cairan tanaman, akibatnya menimbulkan gangguan secara fisiologi. Kerusakan tidak langsung yaitu berupa akibat yang ditimbulkannya sebagai vektor virus penyebab penyakit tanaman. Kutukebul T. vaporariorum mampu menularkan TICV (Tomato infectious
chlorosis virus) secara semipersisten (Duffus et al. 1994; Wisler et al. 1996).
TICV adalah virus tanaman yang berasal dari famili Closteroviridae dan genus
Crinivirus (Martelli et al. 2002). Closterovirus ini menginduksi penyakit klorosis
pada bagian jaringan floem (Duffus et al. 1996).
TICV merupakan virus dengan partikel berbentuk batang lentur dengan ukuran sekitar 650 nm (Jones 2003). Gejala TICV pada daun tomat umumnya tampak jelas pada daun yang lebih tua di bagian bawah tanaman (Wisler et al. 1998). Gejala awal muncul berupa jaringan di antara tulang daun mengalami klorosis dan berwarna kuning (Duffus et al. 1996; Hirota et al. 2010), adanya bintik-bintik nekrotik kecil (Wintermantel & Wisler 2006), namun pertumbuhan baru terus berlangsung. Gejala kuning terlihat pada seluruh daun-daun di bagian bawah, tengah, maupun atas (Anfoka & Abhary 2007), namun mulai muncul dari daun bagian bawah tanaman dan berlanjut ke bagian pucuk. Gejala lanjutan berupa warna merah-keunguan pada daun tua (Wisler et al. 1998), daun menggulung ke bawah, mengering dan rapuh yang diikuti dengan kehilangan hasil
yang banyak. Kehilangan hasil ini disebabkan karena area fotosintesis pada daun yang berkurang. Selain itu kualitas buah tomat yang dihasilkan rendah mutunya, karena pada umumnya buah masak tidak merata sebelum waktunya dan berukuran lebih kecil.
TICV dilaporkan ditularkan hanya oleh T. vaporariorum dan tidak dapat ditularkan oleh spesies kutukebul lainnya (Duffus et al. 1996). Dengan demikian,
T. vaporariorum saja yang berperan sangat penting dalam penyebaran TICV. Ada
kemungkinan satu ekor imago T. vaporariorum mampu menularkan virus lebih dari sekali. Meski demikian, penelitian mengenai berapa lama virus tersebut dapat bertahan dalam tubuh vektornya (periode retensi) belum diketahui, oleh karena itu penelitian ini juga dilakukan untuk mengetahui periode retensi TICV dalam tubuh vektor T. vaporariorum. Indikasi tanaman terinfeksi atau tidak oleh suatu patogen adalah dengan melihat gejala yang muncul. Konfirmasi gejala yang diakibatkan oleh virus TICV yang dimaksud atau tidak, dapat dilakukan dengan uji molekuler menggunakan Polymerase Chain Reaction (PCR) pada daun yang terinfeksi.
Tujuan
Penelitian ini bertujuan mengukur panjang rostrum dan sayap kutukebul
T. vaporariorum pada beberapa ketinggian tempat berbeda serta periode retensi
TICV yang ditularkannya pada tanaman tomat.
TINJAUAN PUSTAKA
Taksonomi dan Morfologi Trialeurodes vaporariorum (Westwood) Kutukebul T. vaporariorum merupakan hama utama pada berbagai tanaman hortikultura dan tanaman hias. Kutukebul ini bersifat polifag. Tanaman inang T. vaporariorum meliputi beberapa tanaman seperti tomat, cabai, terung, buncis, timunsuri, paria, kacang hijau, ubi jalar, stroberi, tembakau, dan bahkan mawar. Secara taksonomi, T. vaporariorum termasuk dalam ordo Hemiptera, subordo Sternorrhyncha, superfamili Aleyrodoidea, famili Aleyrodidae, dan sub famili Alerodicinae (Martin 1987).
Kutukebul T. vaporariorum ini secara umum dikenal sebagai greenhouse
whitefly atau kutukebul rumah kaca yang habitatnya di daerah beriklim sedang di
dunia (Kessing & Mau 2009). Siklus hidup kutukebul terdiri dari empat fase perkembangan, yaitu telur, nimfa, pupa, dan imago. Kutukebul dapat bereproduksi secara seksual maupun partenogenesis. Imago betina yang sudah dibuahi imago jantan biasanya akan memilih salah satu tempat di permukaan bawah daun dan diam di tempat tersebut sampai ia meletakkan telurnya.
Menurut Roermund & Lenteren (1992), ciri morfologi T. vaporariorum adalah sebagai berikut: telur berbentuk bulat panjang (± 0,25 mm), permukaannya licin dengan tangkai yang pendek pada salah satu ujungnya. Posisi telur pada umumnya tegak lurus atau vertikal, diletakkan dalam kumpulan lilin putih berpola melingkar dengan tangkai telur menempel pada daun inangnya. Telur yang baru dikeluarkan imago betina berwarna putih pucat agak krem, kemudian setelah 24 jam berubah warnanya menjadi kekuningan. Warna telur yang berumur lebih tua akan semakin gelap dan akhirnya kehitaman menjelang keluarnya nimfa instar satu sekitar hari ke-8 (Nielsen 2000).
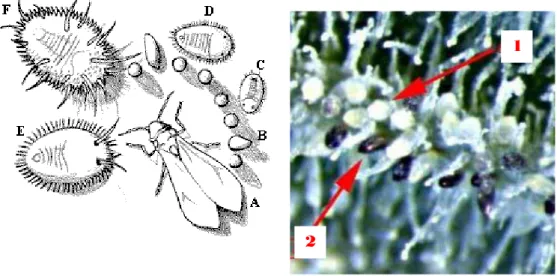
(a) (b)
Gambar 1 Kutukebul T. vaporariorum. (a) Tahapan perkembangan
T. vaporariorum: A= imago, B= telur, C= nimfa i.1, D= nimfa i.2,
E= nimfa i.3, F= pupa, dan (b) telur T. vaporariorum: 1= telur berumur 1-2 hari yang berwarna kuning pucat, 2= Telur yang berumur lebih tua, berwarna lebih hitam (Nielsen 2000).
Nimfa instar 1 berbentuk bulat panjang, berwarna krem cerah dengan panjang tubuh 0,30 mm. Nimfa ini bergerak secara aktif dan disebut “crawler”, biasanya selama 1-2 hari. Nimfa menghisap sap atau cairan daun tanaman yang mengandung protein dan ekstrak nutrisi lainnya.
Nimfa instar 2 berwarna krem lebih gelap daripada instar satu dengan antena sangat pendek dan tungkai yang tereduksi. Nimfa instar 3 berwarna krem dengan ukuran yang lebih besar daripada instar 2, dengan segera tinggal dan masuk fase istirahat (immobile). Nimfa instar 2 dan 3 tidak bergerak dan menetap di permukaan bawah daun. Tahap perkembangan nimfa akhir terdiri dari nimfa instar 4, prapupa, dan pupa, namun biasanya ketiga tahap ini disebut dengan fase “pupa” saja. Pupa berbentuk bulat panjang, dibagian toraks agak melebar dan cembung dengan abdomen yang tampak jelas. Pupa berukuran 0,73-0,75 mm, lebih tebal daripada nimfa instar sebelumnya dan berwarna keputihan.
Pupa memiliki semacam rambut panjang yang keluar pada bagian dorsal dan margin tubuhnya. Susunan mata dan jaringan tubuh di dalam pupa menjadi jelas terlihat selama fase ini (Roermund & Lenteren 1992). Lama stadium pupa berkisar antara 3-7 hari. Ciri khas pupa T. vaporariorum ditandai pada bagian-bagian seperti lingual, vasiform orifice, dan papila submargin.
1 T . v a p o r a r i o r u m 2 T . v a p o r a r i o r u m
T. vaporariorum memiliki lingual yang membulat, barisan papila pada
submarginnya, serta basal tungkai tengah dan belakang mempunyai seta yang halus dan kecil (Martin 1987). Setelah imago keluar, maka kulit/ kantung pupa yang berwarna transparan akan ditinggalkan. Antena imago biasanya panjang dan berbentuk moniliform, dengan 3-7 ruas. Mata majemuk berkembang baik. Imago kutukebul ini panjangnya 1-2 mm dengan warna tubuh kekuningan dan memiliki empat sayap berlapis lilin yang hampir sejajar dengan permukaan daun. Sayap imago baik jantan maupun betina bentuknya menyerupai tenda segitiga dan menutupi hampir seluruh tubuhnya (Smith 2009).
Kutukebul T. vaporariorum yang memasuki stadium imago pada umumnya menetap pada daun-daun muda dekat titik tumbuh tanaman dan bertelur di tempat tersebut. Kutukebul rumah kaca ini reproduksinya relatif lambat, yakni satu generasi tiap 30 sampai 45 hari. Akan tetapi seekor imago betina mampu bertelur sampai 250 butir. Lama perkembangan serangga tergantung temperatur dan jenis tanaman inang. Temperatur perkembangan optimum berkisar antara 21-24 ºC. Pada tanaman tomat, durasi perkembangan T. vaporariorum pada stadium telur adalah sekitar 8 hari, nimfa instar 1 sekitar 6 hari, nimfa instar 2 selama 2 hari, nimfa instar 3 sekitar 3 hari, stadium pupa sekitar 9 hari, dan imago rata-rata sekitar 7-8 hari (Roermund & Lenteren 1992).
Tahapan perkembangan T. vaporariorum merupakan peralihan antara paurometabola dan holometabola, karena nimfa instar akhir hanya diam dan seperti pupa. Imago T. vaporariorum baik jantan maupun betina dapat terbang dengan baik karena keduanya mempunyai dua pasang sayap yang berselaput tipis. Sepasang sayap depan mempunyai sifat yang seragam seluruhnya, demikian pula halnya dengan sepasang sayap belakang. Warna sayapnya keruh, keputihan, dan tertutup dengan serbuk yang putih, sayap-sayap belakang hampir sama besarnya dengan sayap depan, dan tidak ada kornikelnya. Pada waktu serangga sedang istirahat sayap-sayapnya diletakkan seperti atap di atas tubuh.
Kutukebul T. vaporariorum dan anggota Aleyrodidae lainnya memiliki bentuk alat mulut menusuk-menghisap yang terdiri dari empat struktur tubular yang disebut stilet. Dua stilet mandibulata melakukan aktivitas mekanik dan perpindahan secara bebas pada tiap-tiap penetrasi melalui ruang interseluler. Dua
stilet maksila menghubungkan dua bentuk pembuluh utama, saluran makanan, dan kelenjar ludah (Forbes 1969). Kutukebul dapat mengeluarkan lapisan lilin berwarna putih dari kelenjar khusus yang ada pada bagian abdomen. Lapisan lilin ini bervariasi bentuknya pada masing-masing spesies, baik pada stadium nimfa maupun imago, sehingga dapat dijadikan dasar identifikasi (Botha et al. 2000).
Stadium nimfa instar 1, instar 2, instar 3, dan imago menyebabkan kerusakan tanaman dengan cara memasukkan stiletnya ke dalam tulang daun dan mengekstrak sap floem untuk memperoleh makanannya (Wintermantel 2004). Cara makan nimfa yaitu dengan menghisap ekstrak protein dan nutrisi dari sap tanaman serta mengekskresikan kelebihan gula. Cairan gula ini merupakan limbah pencernaan yang dikenal dengan istilah embun madu. Pada populasi yang tinggi, jumlah embun madu pun melimpah dan menyebabkan tanaman atau buah-buahan menjadi lengket. Bila embun madu ini ditumbuhi cendawan embun jelaga
Capnodium sp., maka akan menyebabkan terbatasnya sistem fotosintesis. Selain
nimfa, stadium yang potensinya lebih berbahaya yaitu imago, karena imago dapat menularkan virus tanaman.
Musuh alami kutukebul T. vaporariorum di alam adalah parasitoid
Encarsia formosa (Hymenoptera: Aphelinidae). Imago betina parasitoid ini
meletakkan telurnya pada larva T. vaporariorum, sehingga pada saatnya telur parasitoid akan menetas di dalam tubuh inang dan mengambil nutrisi dari larva
T. vaporariorum (parasit larva). Akibatnya larva T. vaporariorum lama-kelamaan
akan mati karena kekurangan nutrisi dan infeksi (Roermund & Lenteren 1992). Musuh alami lainnya yaitu Eretmocerus eremicus (Hymenoptera: Aphelinidae),
Macrolophus caliginosus (Hemiptera: Miridae), dan Amblyseius swirskii
(Mesostigmata: Phytoseiidae).
Pengaruh Ketinggian dan Suhu terhadap Serangga
Variasi ukuran pada serangga dapat dipengaruhi oleh faktor genetik, lingkungan, ataupun interaksi antara keduanya (Zera 2004). Variasi morfologi serangga juga dipengaruhi oleh posisi dan ketinggian tempat. Serangga yang berada pada posisi yang lebih tinggi berukuran lebih besar, lebih berat, dan berwarna lebih gelap daripada posisi yang lebih rendah. Kutukebul T. vaporariorum dapat ditemukan pada ketinggian 950 – 1500 mdpl (Nurrohman 2003). Kecenderungan
peningkatan bobot tubuh, lebar kapsul kepala, dan panjang sayap yang berada pada ketinggian tempat yang lebih tinggi terjadi pada serangga Dalbulus maydis (Hemiptera: Cicadellidae) (Oliveira et al. 2004). Populasi serangga Lycaena sp. (Lepidoptera: Lycaenidae) yang berada pada tempat yang lebih tinggi menunjukkan peningkatan ukuran telur dan panjang sayap (Fischer & Karl 2010). Tinggi atau rendahnya suatu tempat di permukaan bumi berpengaruh terhadap suhu udaranya. Suhu di negara tropis seperti Indonesia menunjukkan adanya penurunan seiring dengan makin tingginya tempat. Namun kenyataannya, terdapat faktor lainnya yang juga mempengaruhi fluktuasi suhu harian, misalnya kelembapan udara. Kelembapan udara menyatakan banyaknya uap air dalam udara. Uap air ini mempunyai sifat menyerap radiasi bumi sehingga menentukan cepatnya kehilangan panas dari bumi dan dengan sendirinya juga ikut mengatur suhu (Wisnubroto et al. 1986).
Suhu mempengaruhi ukuran tubuh kutu daun dan ukuran tubuh beberapa serangga lainnya. Serangga-serangga tersebut akan berukuran lebih kecil ketika berkembang pada suhu yang lebih tinggi (Dixon 1985). Hal tersebut dibuktikan oleh Blackman (1994) pada spesies Myzus persicae (Hemiptera: Aphididae) yang mengalami penurunan ukuran tubuh pada pertumbuhan di suhu yang lebih tinggi. Menurut Digby (1954), suhu dan tingkat radiasi mempengaruhi aktivitas dan ukuran panjang tubuh beberapa spesies lalat. Murai & Toda (2001) juga menyatakan bahwa individu Thrips tabaci (Thysanoptera: Thripidae) yang hidup pada suhu yang lebih rendah mempunyai warna tubuh yang lebih gelap dan ukuran tubuh yang lebih besar daripada suhu yang lebih tinggi.
Semua stadium T. vaporariorum memiliki kemampuan beradaptasi di suhu dingin dibandingkan B. tabaci (Xie et al. 2006). Menurut Roermund & Lenteren (1992), suhu mempengaruhi parameter kehidupan T. vaporariorum, seperti tingkat perkembangan nimfa, kematian nimfa, seks rasio, lama hidup, periode preoviposisi, fekunditas, frekuensi oviposisi, serta peningkatan periode oviposisi harian. Smith (2009) menyatakan bahwa imago T. vaporariorum mampu hidup normal pada suhu antara 22 ˚C sampai 25 ˚C, sedangkan pada suhu di atas 30 ˚C imago tidak mampu berkembang dan pada suhu 35 ˚C imago akan mati.
Tomato infectious chlorosis virus (TICV)
TICV merupakan virus dengan partikel berukuran sekitar 650 nm, berbentuk batang lentur. Virus ini berkembang hanya pada bagian jaringan floem saja (Duffus et al. 1996). TICV termasuk kelompok genom bipartite RNA untai tunggal (ssRNA), dengan panjang genom RNA1 7,8 kb dan RNA2 7,4 kb (Liu et al. 2000). TICV merupakan anggota famili Closteroviridae dan genus
Crinivirus (Hull 2002, Martelli et al. 1999 di dalam Liu et al. 2000). Keberadaan
TICV telah dilaporkan di beberapa wilayah penghasil tomat dunia seperti di California (Duffus et al. 1994), North Carolina (Vaira et al. 2002), Spanyol (Font et al. 2002), Yunani (Dovas et al. 2002), dan Perancis (Dalmon et al. 2005). TICV adalah masalah tomat yang penting dalam pertanian dunia. Bahkan, TICV telah menyebabkan kerugian sekitar $ 2 juta di Orange County pada tahun 1993 (Wisler et al. 1997). Di belahan Asia, TICV terdeteksi pertama kalinya pada
tanaman tomat di Indonesia dan Jepang (Verhoeven et al. 2003; Hartono et al. 2003), serta Yordania (Anfoka & Abhary 2007).
Gejala TICV pada daun tomat umumnya tampak jelas pada daun yang lebih tua di bagian bawah tanaman (Wisler et al. 1998). Gejala awal muncul berupa jaringan di antara tulang daun menguning (Duffus et al. 1996; Hirota et al. 2010), adanya bintik nekrotik kecil (Wintermantel & Wisler 2006), namun pertumbuhan baru terus berlangsung. Gejala kuning terlihat jelas pada seluruh daun (Anfoka & Abhary 2007), namun mulai muncul dari daun bagian bawah tanaman dan berlanjut ke bagian atas tanaman tomat. Gejala lanjutan berupa warna merah-keunguan pada daun tua (Wisler et al. 1998), daun menggulung ke bawah, daun mengering dan rapuh yang diikuti dengan kehilangan hasil yang banyak. Kehilangan hasil ini disebabkan karena area fotosintesis pada daun yang berkurang. Selain itu kualitas buah tomat yang dihasilkan rendah mutunya, karena pada umumnya buah masak sebelum waktunya dan berukuran lebih kecil. Kesenjangan kualitas inilah yang menjadi faktor pembatas bagi petani tomat. Sayangnya terkadang gejala disalahartikan sebagai penuaan alami, kekurangan nutrisi, gangguan fisiologis, atau bahkan fitotoksisitas dari pestisida.
TICV dilaporkan hanya dapat ditularkan oleh T. vaporariorum dan tidak oleh spesies kutukebul lainnya (Duffus et al 1996). Virus ini juga tidak dapat
ditularkan melalui benih maupun perasan tanaman sakit, sehingga
T. vaporariorum berperan sangat penting dalam penyebaran patogen TICV.
TICV merupakan patogen penyebab penyakit klorosis pada tanaman tomat. Kehadiran patogen ini tentunya mengancam produksi tomat di Indonesia maupun di seluruh dunia.
Hubungan Virus dengan Serangga Vektor serta Periode Retensi
Penggolongan virus yang ditularkan oleh kutukebul berdasarkan lamanya vektor mempertahankan virus dalam tubuhnya. Penggolongan ini berupa nonpersisten, semipersisten, dan persisten. Virus dianggap golongan non-persisten bila kemampuan vektor menularkan virus hilang dalam beberapa menit atau beberapa jam saja. Bila kemampuan vektor menularkan virus hilang setelah beberapa jam, maka digolongkan ke dalam semipersisten. Bila kemampuan vektor untuk menularkan virus tersimpan dalam kurun waktu beberapa hari atau selama hidupnya, maka digolongkan ke dalam persisten (Watson & Robers 1939; Sylvester 1956). TICV ditularkan oleh serangga vektor T. vaporariorum secara semipersisten.
Periode retensi adalah selang waktu vektor masih dapat menularkan patogen sampai serangga tersebut tidak dapat menimbulkan gejala atau menularkan virus lagi. Lama periode retensi ini tergantung spesies kutukebul serta jenis virus yang ditularkannya.
BAHAN DAN METODE
Tempat dan Waktu
Kegiatan survei dan pengambilan sampel kutukebul dilakukan di sentra produksi tomat di Kecamatan Cikajang (kabupaten Garut), Kecamatan Pacet (Kabupaten Cianjur), Kecamatan Cikole (kabupaten Sukabumi), dan Kecamatan Batu (kotamadya Batu, Malang). Penelitian mengenai lama waktu retensi virus dilakukan di rumah kaca Cikajang, Garut. Identifikasi kutukebul dilaksanakan di Laboratorium Biosistematika Serangga, sedangkan identifikasi virus dilakukan di Laboratorium Virologi Tumbuhan, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor. Data mengenai unsur cuaca diperoleh dari Badan Meteorologi dan Geofisika (BMKG) Darmaga, Bogor dan BMKG Karangploso, Malang. Penelitian dilaksanakan dari bulan Maret sampai November 2010.
Metode Penelitian
Pengambilan Sampel Kutukebul dan Tanaman Tomat Sumber TICV
Sampel kutukebul diambil dari sentra produksi tomat di Kecamatan Cikajang di Kabupaten Garut (1287 m dpl) pada lintang 7˚21’35,05” S dan bujur 107˚48’50,82” T, Kecamatan Pacet di Kabupaten Cianjur (1205 m dpl) yang terletak di 6˚73’59.51” S & 107˚41’77.12” T, Kecamatan Cikole di Kabupaten Sukabumi (1022 m dpl) yang terletak di 6˚51’48.57”S dan 106˚56’56.37” T, dan Kecamatan Batu di Kotamadya Batu, Malang (675 m dpl) yang berada di 7˚53’19.32” S dan 112˚35’29.14” T. Sampel imago kutukebul dengan ciri-ciri
T. vaporariorum diambil dari daun tomat dengan menggunakan aspirator,
sedangkan pupa dan kantung pupa diambil dari bagian daun tempat melekatnya serangga ini. Tanaman tomat sumber inokulum TICV untuk penelitian periode retensi diambil dari daerah Cikajang, Garut.
Pembuatan Preparat dan Identifikasi T. vaporariorum
Pupa dan kantung pupa kutukebul dari lapangan dilepaskan dari daun tomat dengan menggunakan jarum dan dimasukkan ke dalam tabung reaksi yang berisi larutan alkohol 80%. Tabung reaksi tersebut dimasukkan ke dalam gelas piala berisi kapas dan air, kemudian dipanaskan pada suhu 100 ºC selama 10 menit. Hasil rebusan tersebut dituang ke dalam cawan syracuse dengan memakai penjepit. Tahap selanjutnya pupa dimasukkan ke dalam tabung reaksi berisi larutan KOH 10% dan direbus seperti sebelumnya. Setelah pupa lunak dan berwarna transparan, tabung reaksi diangkat dan dituang ke dalam cawan syracuse. Supaya cairan isi pupa keluar dan pupa tidak sobek, maka secara perlahan pupa ditekan dengan jarum halus. Pupa yang telah bersih isinya kemudian dipindahkan ke cawan yang baru dan dibilas dengan aquades.
Proses selanjutnya adalah pewarnaan. Pupa yang sudah dicuci bersih dipindahkan ke dalam cawan syracuse baru yang berisi campuran 1 ml asam asetik glasial serta 1 ml asam fuchsin dan direndam selama 20 menit. Pupa yang telah berwarna merah kemudian direndam ke dalam larutan alkohol 80% selama 5 menit agar mendapatkan warna merah yang optimum. Pupa selanjutnya direndam selama satu menit dalam cawan baru berisi carbol xylene. Tahapan berikutnya, pupa direndam selama 5 menit dalam alkohol absolut. Lemak pada pupa akan hancur, kemudian direndam selama sepuluh menit ke dalam minyak cengkeh. Pupa siap untuk dibuat preparat.
Preparat kutukebul dibuat dengan meletakkan pupa di bagian tengah kaca objek. Kemudian canada balsam diteteskan di atas pupa dan diratakan. Setelah kaca penutup dipasang pada kaca objek, preparat selanjutnya diletakkan ke dalam elemen pengering selama tujuh hari. Preparat awetan puparium diidentifikasi menggunakan kunci identifikasi dari Martin (1987). Serangga yang telah diidentifikasi sebagai T. vaporariorum kemudian diperbanyak pada tanaman tomat, Selanjutnya imago yang muncul digunakan untuk percobaan periode retensi TICV.
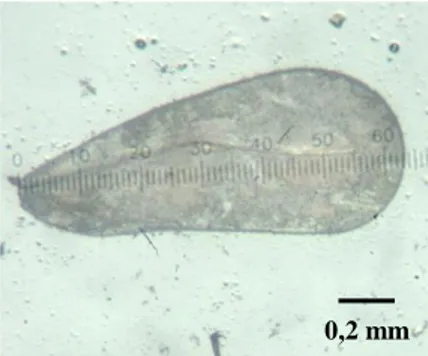
Sebanyak 30 ekor imago betina T. vaporariorum dari masing-masing lokasi yang telah dipilih diukur panjang rostrum (1a) dan panjang sayap depannya (1b) menggunakan mikroskop stereo bermikrometer.
Perbedaan panjang rostrum dan panjang sayap kutukebul dari empat ketinggian tempat berbeda diuji menggunakan uji t sampel bebas dengan program Microsoft Excel.
(a) (b)
Gambar 2 Pengukuran T. vaporariorum: (a) rostrum dengan perbesaran 11x10 dan (b) sayap depan dengan perbesaran 5x10
Pengumpulan Data Cuaca
Data cuaca seperti suhu (T) dan kelembaban (RH) diperoleh dari Badan Meteorologi dan Geofisika (BMKG) Darmaga, Bogor dan BMKG Karangploso, Batu. Namun data suhu di Kecamatan Cikole, Sukabumi tidak akurat dikarenakan kesalahan pengamatan (pengamat belum terampil), sehingga untuk memperoleh data suhu di tempat tersebut dihitung dengan menggunakan rumus Mock (1969) sebagai berikut:
T = 0,006 (x1 – x2 ) . 1° C Keterangan :
T = Selisih suhu udara antara lokasi 1 dengan lokasi 2 (°C). x1= Tinggi tempat yang diketahui suhu udaranya (m). x2= Tinggi tempat yang dicari suhu udaranya (m).
Pengukuran Periode Retensi TICV dalam Kutukebul T. vaporariorum
Sumber inokulum berasal dari tanaman tomat kultivar Marta yang positif terinfeksi TICV dengan gejala khas Crinivirus (seperti warna kuning-keunguan dan munculnya dimulai dari bagian bawah ke bagian atas tanaman). Tanaman uji yang digunakan dalam percobaan ini adalah tomat kultivar Marta. Benih tomat terlebih dahulu disemai dalam tray yang telah diisi tanah dan humus. Kemudian setelah berkecambah dan keluar dua daun pertama (berumur kurang lebih 2 MST), bibit tomat dipindahtanamkan ke dalam polybag yang telah diisi dengan tanah dan pupuk kandang dengan perbandingan 2:1. Tiap polybag hanya diisi satu bibit tomat saja. Bibit tomat siap digunakan untuk percobaan.
Penelitian periode retensi dilaksanakan dengan melakukan inokulasi berseri menggunakan imago kutukebul hasil pemeliharaan yang viruliferus (mengandung virus) ke bibit tomat (sebagai tanaman uji). Sejumlah imago
T. vaporariorum dipindahkan ke dalam kurungan kasa yang berisi tanaman sakit
(sumber inokulum) dan dibiarkan makan selama 24 jam (periode akuisisi). Kutukebul yang viruliferus kemudian dipindahkan ke bibit tanaman tomat sehat sejumlah satu ekor tiap tanaman uji, kemudian disungkup menggunakan plastik mika yang sudah dimodifikasi dasarnya. Kutukebul dibiarkan selama 24 jam pada tanaman tersebut (periode inokulasi). Kemudian masing-masing kutukebul tersebut dipindahkan lagi ke bibit tanaman tomat sehat lainnya dan dibiarkan selama 24 jam, begitu seterusnya sampai kutukebul tersebut mati. Tanaman hasil perlakuan selanjutnya diinkubasi di rumah kaca selama tiga minggu. Perawatan tanaman dilakukan setiap hari. Parameter yang diamati meliputi ada atau tidaknya gejala klorosis pada daun. Pengamatan dilakukan setiap hari dengan mencatat hari kemunculan gejala mulai inokulasi kutukebul viruliferus tersebut hingga tanaman uji tidak bergejala lagi. Masing-masing seri inokulasi menggunakan sepuluh ulangan. Sebagai kontrol adalah tanaman uji yang ditulari dengan kutukebul yang tidak viruliferus dengan cara diakuisisikan pada tanaman tomat sehat.
Gambar 3 Bibit tomat yang sudah diinokulasi TICV
Gejala klorosis yang muncul pada tanaman uji diuji secara molekuler dengan menggunakan Polymerase Chain Reaction (PCR) untuk meyakinkan bahwa gejala tersebut benar-benar disebabkan oleh infeksi TICV. Ekstraksi RNA daun tanaman uji dilakukan menggunakan Rneasy Plant Mini Kits (Qiagen Inc., Chatsworth, CA, USA) sesuai dengan prosedur yang ditetapkan oleh Qiagen. RNA hasil ekstraksi selanjutnya dipakai sebagai template dalam Reverse
Transcriptase (RT) atau transkripsi balik. Larutan untuk reaksi RT-PCR
mengandung 2 µl RNA total, 1 µl buffer RT 10X, 0,35 µl 50 mM DTT (dithiothreitol), 2 µl 10 mM dNTP (deoksiribonukleotida triphosphat), 0,35 µl M-MuLV Rev, 0,35 µl RNase inhibitor, 0,75 µl oligo (dT), dan 3,2 µl H2O.
Amplifikasi dilakukan dalam sebuah Automated Thermal cycler (Gene Amp PCR
System 9700; PE Applied Biosystem, USA) yang diprogram untuk satu siklus pada
suhu 25 ºC selama 5 menit, 42 ºC selama 60 menit, dan 70 ºC selama 15 menit.
Complementary DNA (cDNA) yang didapat dipakai sebagai template dalam
proses PCR dengan campuran 1 µl primer spesifik untuk mendeteksi virus TICV, yaitu TICV-CF (5’-AATCGGTAGTGACACGAGTAGCATC-3’) dan TICV-CR (5’-CTTCAAACATCCTCCATCTGCC-3’) dengan ukuran produk PCR 417 bp, 2,5 µl buffer PCR 10X + Mg2+, 0,5 µl 10 mM dNTP, 2,5 µl sucrose cresol 10X, 0,3 µl Taq DNA polymerase, 15,2 µl H2O, dan 1 µl cDNA. Amplifikasi DNA
diawali dengan pradenaturasi pada suhu 94 ºC selama 4 menit. Selanjutnya secara berturut-turut berlangsung fase denaturasi pada 94 ºC selama 1 menit, annealing pada suhu 62 ºC selama 1 menit, dan elongasi pada suhu 72 ºC selama 2 menit yang diteruskan tahap pascaextention selama 10 menit serta penyimpanan pada suhu 4 ºC. Produk PCR selanjutnya dielektroforesis pada gel agarose 1%, dengan tegangan 50 Volt selama 45 menit. Visualisasi pita DNA dengan transluminator
UV. Hasil daun tomat yang positif terinduksi TICV akan terlihat bila pita DNA
berada pada 417 bp. Hasil visualisasi kemudian difoto menggunakan kamera digital.
HASIL DAN PEMBAHASAN
Identitas T. vaporariorum
Berdasarkan pengamatan, imago kutukebul T. vaporariorum baik jantan maupun betina mempunyai dua pasang sayap yang berselaput tipis. Sepasang sayap depan mempunyai sifat yang seragam seluruhnya, demikian pula halnya dengan sepasang sayap belakang. Warna sayapnya keruh, keputihan, dan tertutup dengan serbuk yang putih. Pada waktu serangga sedang istirahat sayap-sayapnya diletakkan seperti atap di atas tubuh, dengan tepi bagian dalam yang agak tumpang tindih di bagian ujungnya.
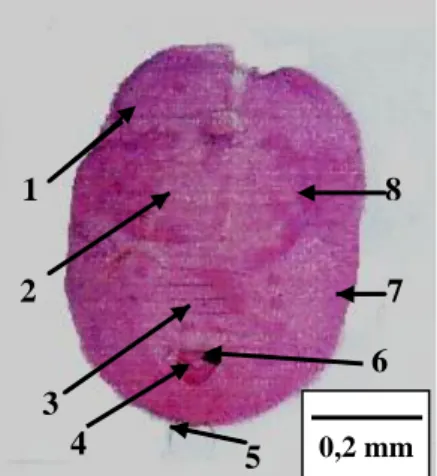
(a) (b)
Gambar 4 Pupa T. vaporariorum (a) pada daun tomat dan (b) preparat pupa: (1) pinggiran trakea, (2) rambut dorsal, (3) ruas abdomen VII, (4)
vasiform orifice, (5) seta kaudal, (6) lingula, (7) submarginal, (8) basal
tungkai tengah dan belakang
Tahapan perkembangan T. vaporariorum merupakan peralihan antara paurometabola dan holometabola, karena nimfa instar akhir hanya diam dan seperti pupa. Pupa T. vaporariorum berwarna kuning pucat, agak tebal, dan panjangnya sekitar 0,73 mm (gambar 4a). Setelah dibuat preparat, pupa kutukebul
yang diidentifikasi untuk penelitian mempunyai ciri khas pupa spesies 1 2 3 4 5 6 7 8 0,15 mm 0,2 mm
T. vaporariorum, yakni berbentuk ovoid, pinggiran trakeanya tidak seperti sisir,
tidak mempunyai rambut dorsal, lingula membulat, terdapat barisan papila pada submarginal, serta basal tungkai tengah dan belakang mempunyai seta yang kecil dan halus (gambar 4b). Hal tersebut berdasarkan deskripsi pada Martin (1987).
Panjang Rostrum dan Sayap T. vaporariorum
Imago betina T. vaporariorum yang diambil dari empat lokasi berbeda, yakni Cikajang, Pacet, Cikole, dan Batu, memiliki ciri-ciri yang sama. Pada waktu makan, rostrum ditusukkan secara tegak lurus pada permukaan daun (biasanya diikuti gerakan sayap, abdomen, dan tungkainya). Rostrum T. vaporariorum bentuknya tidak benar-benar lurus, namun agak membengkok pada bagian tengah. Ujung rostrum berwarna coklat tua pada bagian ujung.
(a) (b)
Gambar 5 Morfologi imago T. vaporariorum, (a) saat sedang istirahat dan (b) saat sedang makan
Panjang rata-rata rostrum imago betina T. vaporariorum yang berasal dari Cikajang adalah 293,18 µm, dari Pacet sebesar 275,76 µm, dari Cikole sebesar 256,36 µm, dan dari Batu adalah 251,81 µm. Berdasarkan data yang didapat dari pengukuran panjang rostrum dan sayap imago betina T. vaporariorum, terlihat
hasil yang berbeda nyata (Tabel 1). Panjang rostrum imago betina
T. vaporariorum yang berasal dari Cikajang berbeda nyata dengan panjang
rostrum imago betina dari Pacet, Cikole, dan Batu. Panjang rostrum sampel yang berasal dari Pacet berbeda nyata dengan panjang rostrum dari Cikajang, Cikole, dan Batu. Panjang rostrum sampel imago betina kutukebul T. vaporariorum yang berasal dari Cikole berbeda nyata dengan panjang rostrum imago betina dari
Cikajang dan Pacet, namun tidak berbeda nyata dengan sampel dari Batu. Panjang rostrum imago betina yang berasal dari Batu berbeda nyata dengan panjang rostrum imago betina dari Cikajang dan Pacet, namun tidak berbeda nyata dengan sampel dari Cikole. Hal ini kemungkinan disebabkan oleh suhu di kedua tempat tersebut yang tidak jauh berbeda, sehingga memberikan pengaruh yang sama terhadap perkembangan kutukebul. Menurut Digby (1954) semakin tinggi suhu lingkungan yang diterima, maka ukuran serangga akan semakin kecil.
Tabel 1 Panjang rostrum dan sayap T. vaporariorum
Ketinggian (m dpl) Suhu (˚C) Kelembaban (%) Panjang ± SBa (µm) Rostrum Sayapb
Cikajang 1287 19,8 87,3 293,18 ± 25,62a 1286,33 ± 41,89a Pacet 1125 20,9 83 275,76 ± 13,15b 1212,00 ± 21,40b Cikole 1022 22,2 80 256,36 ± 15,55c 1162,67 ± 19,46c Batu 675 22,4 78,2 251,81 ± 13,54c 1119,33 ± 27,03d
a
SB = simpangan baku
bangka yang diikuti huruf yang berbeda pada kolom yang sama menunjukkan hasil yang berbeda nyata (uji t pada α = 0,01)
Panjang rata-rata sayap imago yang berasal dari Cikajang, Pacet, Cikole, dan Batu berturut-turut adalah 1286,33 µm, 1212 µm, 1162,67 µm, dan 1119,33 µm. Panjang sayap imago betina T. vaporariorum yang berasal dari Cikajang berbeda nyata dengan panjang sayap imago betina dari Pacet, Cikole, dan Batu. Panjang sayap imago yang berasal dari Pacet berbeda nyata dengan panjang sayap dari Cikajang, Cikole, dan Batu. Panjang sayap imago betina T. vaporariorum yang berasal dari Cikole berbeda nyata dengan panjang sayap imago betina dari Cikajang, Pacet, dan Batu. Panjang sayap imago betina T. vaporariorum yang berasal dari Batu berbeda nyata dengan panjang sayap imago betina dari Cikajang, Pacet, dan Cikole.
Rostrum dan sayap imago betina yang berasal dari Cikajang memiliki ukuran paling panjang dibandingkan lokasi lainnya. Sebaliknya, panjang rostrum dan sayap imago betina yang berasal dari Batu memiliki ukuran paling pendek dibandingkan lokasi lainnya. Hal ini kemungkinan disebabkan karena ketinggian tempat di Cikajang paling tinggi (1287 m dpl) daripada lokasi lainnya, sedangkan Batu mempunyai ketinggian tempat yang paling rendah (675 m dpl) dibandingkan
lokasi pengamatan lainnya. Tinggi rendahnya suatu tempat di permukaan bumi berpengaruh terhadap suhu udaranya. Semakin tinggi letak suatu tempat, maka suhunya pun akan semakin rendah. Ukuran rostrum dan sayap kutukebul mengalami pertambahan seiring dengan pertambahan ketinggian tempat dan penurunan suhu habitat kutukebul. Atau dengan kata lain, semakin tinggi suhu habitat perkembangan T. vaporariorum maka ukuran rostrum dan sayapnya akan semakin pendek. Hal tersebut dibuktikan pula oleh Blackman (1994) pada spesies
Myzus persicae (Hemiptera: Aphididae) serta Murai & Toda (2002) pada spesies Thrips tabaci (Thysanoptera: Thripidae) yang mengalami penurunan ukuran tubuh
imago bila perkembangan pradewasanya berada pada suhu yang lebih tinggi. Suhu mempengaruhi ukuran kutu daun dan beberapa serangga lainnya, yaitu serangga-serangga yang berukuran kecil ketika berkembang pada suhu yang lebih tinggi (Dixon 1985). Hal tersebut dikarenakan fenomena adaptasi dan interaksi suhu dengan proses metabolisme serangga (Blackman 1994).
Suhu di negara tropis seperti Indonesia sangat dipengaruhi oleh kelembaban udara (Wisnubroto et al. 1986). Kelembaban di Cikajang paling tinggi (87,33%) dibandingkan lokasi lainnya. Kelembaban yang tinggi disebabkan letak daerah Cikajang yang berada di lereng gunung dan sebagian besar kawasannya merupakan tadah hujan (rain fed area), sehingga banyak mendapat curah hujan. Letak daerah dan kelembaban yang tinggi tersebut mempengaruhi suhu lingkungan, sehingga suhu di Cikajang juga paling rendah dibandingkan lokasi lainnya, yaitu sebesar 19,8 ˚C.
Periode Retensi TICV dalam Tubuh T. vaporariorum
Tanaman tomat kultivar Marta mengekspresikan gejala infeksi oleh virus TICV lebih jelas dibandingkan tomat kultivar lain (Fitriasari 2010). Tomat kultivar ini banyak ditanam oleh petani tomat baik di wilayah Bogor, Cianjur, Sukabumi, maupun Garut.
Setelah pengamatan selama kurang lebih tiga minggu, terdapat variasi gejala antara tanaman tomat yang diinokulasi imago T. vaporariorum tidak viruliferus dengan yang viruliferus. Daun tanaman tomat yang diinokulasi dengan imago tidak viruliferus terlihat tetap berwarna hijau daun dan segar (gambar 6a). Ukuran dan perkembangan tanaman tomat tampak normal seperti pada umumnya.
Sebaliknya, tanaman tomat yang diberi perlakuan inokulasi kutukebul yang viruliferus menunjukkan ekspresi khas TICV (gambar 6b). Ekspresi atau gejala tersebut berkembang dari bagian bawah ke atas tanaman berupa daun yang klorosis, kemudian berwarna agak coklat keunguan, lebih tebal, lebih kaku, agak menggulung, dan nekrosis. Gejala pada tanaman uji mirip dengan gejala TICV yang ditemukan di lapangan, hanya saja tidak separah gejala di lapangan. Hal ini dikarenakan umur tanaman di lapangan lebih tua daripada tanaman uji, sehingga perkembangan gejalanya pun lebih lama dan gejala yang terlihat lebih parah.
(a) (b)
Gambar 6 Daun tanaman tomat uji yang tidak memperlihatkan gejala klorosis setelah diinokulasi melalui T. vaporariorum yang tidak viruliferus (a) dan yang memperlihatkan gejala klorosis setelah diinokulasi TICV melalui T. vaporariorum yang viruliferus (b).
Data pada Tabel 2 memperlihatkan bahwa periode retensi TICV pada
T. vaporariorum adalah 3 hari. Hal tersebut terjadi karena hubungan antara virus
dan serangga vektornya adalah semipersisten. Pada hubungan semacam ini, virus tidak memperbanyak diri dalam tubuh vektornya, sehingga konsentrasi virus akan terus menurun sampai akhirnya tidak cukup untuk menimbulkan penyakit pada tanaman yang diinokulasinya (misalnya pada hari ke-4).
Tabel 2 Masa infektif T. vaporariorum dalam penularan berseri TICV pada bibit tomat setelah 24 jam periode akuisisia
Pemindahan vektor pada tanaman ke-
Imago T. vaporariorum yang ke-
1 2 3 4 5 6 7 8 9 10 1 + + + + + + + + + + 2 + + + + + + + + + + 3 + + + + + + + + + - 4 - - - - 5 - - - - 6 - - m - m - - - - - 7 m m m m m m m m m m
a Keterangan: + = tanaman terinfeksi, - = tanaman tidak terinfeksi, m = serangga mati
Amplifikasi DNA dilakukan untuk memastikan bahwa gejala klorosis yang muncul pada tanaman uji disebabkan oleh TICV. Amplifikasi DNA dengan metode PCR menggunakan pasangan primer spesifik TICV-CF dan TICV-CR sebagai kontrol positif dengan ukuran 417 bp (lajur K+). Pasangan primer TICV hanya mengamplifikasi virus spesifik TICV saja, sehingga bila tanaman yang diuji tidak mengandung partikel TICV maka hasilnya akan negatif. Dari hasil visualisasi, terbukti bahwa daun tanaman tomat yang diambil dari pemindahan vektor pada tanaman uji ke-3 (memperlihatkan gejala klorosis) positif terinfeksi oleh TICV, hal ini tampak pada lajur 1 pada Gambar 7. Daun tanaman tomat yang diambil dari pemindahan vektor pada tanaman uji ke-4 (tidak memperlihatkan gejala klorosis) ternyata hasilnya negatif atau dengan kata lain tidak mengandung partikel TICV (lajur 2). Penelitian periode retensi ini menunjukkan bahwa semua imago T. vaporariorum yang tidak viruliferus tidak mampu menginfeksi tanaman uji. Seekor imago T. vaporariorum yang viruliferus dapat menularkan TICV sampai pemindahan pada tanaman ke-3 atau sebelum serangga vektor tersebut mati pada pemindahan ke-7 (Tabel 2).
M K+ 1 2 K-
Gambar 7 Hasil amplifikasi DNA genom virus dengan metode RT-PCR menggunakan pasangan primer spesifik TICV-CF dan TICV-CR. Lajur M: Marker 100 bp DNA ladder, lajur K+ : kontrol positif TICV, lajur 1: daun tanaman uji yang terinfeksi atau bergejala klorosis pada pemindahan vektor ke tanaman uji ke-3, lajur 2: daun tanaman uji yang tidak terinfeksi atau tidak bergejala klorosis pada pemindahan vektor ke tanaman uji ke-4, dan lajur K- : kontrol negatif (tanaman tomat sehat).
Menurut Fitriasari (2010), seekor T. vaporariorum sudah mampu menyebabkan infeksi per tanaman sebesar 13,33% dalam masa inkubasi 11 hari, sedangkan sepuluh ekor T. vaporariorum dapat menyebabkan infeksi per tanaman
sebesar 100% dalam masa inkubasi 9 hari. Seekor imago kutukebul
T. vaporariorum yang viruliferus dalam seri inokulasi ke bibit tanaman tomat
sehat mampu menularkan virus TICV sampai inokulasi hari ke-3. Meski ada tanaman uji yang tidak terinfeksi, namun sebagian besar tanaman uji positif terinfeksi TICV. Jadi, rata-rata periode retensi TICV dalam serangga vektor
T. vaporariorum adalah selama tiga hari. Hal tersebut menunjukkan bahwa
periode retensinya relatif lama, karena satu ekor imago mampu menularkan virus TICV lebih dari satu kali inokulasi.
KESIMPULAN DAN SARAN
Kesimpulan
Panjang rostrum dan sayap imago betina T. vaporariorum berbeda nyata antar masing-masing lokasi, kecuali panjang rostrum imago betina dari Cikole dan Batu yang tidak berbeda nyata. Ukuran panjang rostrum dan sayap kutukebul mengalami pertambahan seiring dengan pertambahan ketinggian tempat dan penurunan suhu habitat kutukebul. Periode retensi TICV dalam serangga vektor
T. vaporariorum adalah selama tiga hari.
Saran
Perlu dilakukan penelitian lebih lanjut mengenai pengukuran
T. vaporariorum dari daerah lain dengan ketinggian tempat yang berbeda pula.
Selain itu juga penelitian mengenai periode retensi virus lain di dalam tubuh
T. vaporariorum, karena kutukebul tersebut juga dapat menularkan virus lainnya,
DAFTAR PUSTAKA
Anfoka GH, Abhary MK. 2007. Occurrence of Tomato infectious chlorosis
virus (TICV) in Jordan. Bulletin OEPP/EPPO 37: 186-190.
Blackman RL, Spence JM. 1994. The effects of temperature on aphid morphology, using a multivariate approach. European Journal of Plant
Pathology 91: 7-22.
Botha J, Hardie D, Power G. 2000. Spiraling whitefly Aleurodicus dispersus.
Exotic Threat to western Australia. Fact sheet no. 18/2000.
[CABI] Centre of Agriculture and Biological Institute. 2005. Crop Protection Compendium. London: CABI.
Dalmon A, Bouyer S, Cailly M et al. 2005. First report of Tomato clorosis virus and Tomato infectious chlorosis virus in France. Plant Disease 89, 1243. Digby PSB. 1954. Factors Affecting The Temperature Excess of Insects in
Sunshine. London: University of Oxford.
Dixon AFG. 1985. Aphid Ecology. London: Blackie and Son.
Dovas CI, Katis NI, and Avgelis AD. 2002. Multiplex detection of criniviruses associated with epidemics of a yellowing disease of tomato in Greece.
Plant Disease. 86: 1345-1349.
Duffus JE, Liu HY, Cobn S. 1994. Partial characterization of a new closterovirus, the causal agent of cucurbit yellow stunting disorder. Page 49 in Sweetpotato Whitefy: 1994 Suppleent to the Five-Year Plan US Dep.
Agric. Res. Serv. Publ.112.
Duffus JE, Liu HY, Wisler GC. 1996. Tomato infectious chlorosis virus-a new clostero-like virus transmitted by Trialeurodes vaporariorum. European
Journal of Plant Pathology 102: 219-226.
Fischer K, Karl I. 2010. Exploring plastic and genetic responses to temperature variation using copper butterflies. Climate Research 43: 17-30.
Fitriasari ED. 2010. Keefektifan kutukebul dalam menularkan virus penyebab penyakit kuning pada tanaman tomat [tesis]. Bogor: Sekolah Pascasarjana, Institut Pertanian Bogor.
Font I, Martinez-Culebras P, Jorda MC, Louro D, Vaira AM, et al. 2002. First report of Tomato infectious chlorosis virus in Spain. Plant Disease 86: 696.
Forbes AR. 1969. The stylets of the green peach aphid, Myzus persicae (Homoptera: Aphididae). Can. Entomol. 101, 31-41.
Hartono S, Natsuaki T, Sayama H, Atarashi H, Okuda S. 2003. Yellowing disease of tomatoes caused by Tomato infectious chlorosis virus newly recognized in Japan. J Gen Plant Pathol 69: 61-64.
Hartono S, Wijonarko A. 2007. Karakterisasi biologi molekuler Tomato
infectious chlorosis virus penyebab penyakit kuning pada tanaman tomat
di Indonesia. Jurnal Akta Agrosia Edisi Khusus 2: 139-146.
Hirota T, Natsuaki T, Murai T, Nishigawa H, Niibori K, et al. 2010. Yellowing disease of tomato caused by Tomato chlorosis virus newly recognized in Japan. J Gen Plant Pathol 76: 168-171.
Jones DR. 2003. Plant viruses transmitted by whitelies. European Journal of
Plant Pathology 109: 195-219.
Kessing JLM, Mau RL. 2009. Trialeurodes vaporariorum. http// www.Extento.hawaii.edu/dbase/crop/type/t.vap.html. [Februari 2010]. Liu HY, Wisler GC, Duffus JE. 2000. Particle lengths of whitefly-transmitted
criniviruses. Plant Disease 84: 803-805.
Martelli GP, Agranovsky AA, Bar-Joseph M. 2002. The family Closteroviridae revised. Archives of Virology 147: 2039-2044.
Martin JH. 1987. An Identification guide to common whitefly pest spesies of the world (Homoptera: Aleyrodidae). Top Pest Manag 33(4): 298 – 322. Miller A, Thompson JC, Peterson RE, Haragan DR. 1970. Elements of
Meteorology. Columbus: Charles E. Merrill Publishing Company.
Mock. 1969. Land Capability Appraisal in Indonesia. Soil Research Institute, Bogor.
Morales FJ. 2001. Conventional breeding for resistance Bemisia tabaci-transmitted geminiviruses. Crop Prot 20: 825-843.
Murai T, Toda S. 2002. Variation of Thrips tabaci in colour and size. Proc.
Thrips, Plants, Tospoviruses: the Millennial Review. CSIRO Australia 7,
377-378.
Nielsen MG. 2000. Plantedokteren. Institut for Plantebeskyttelse og Skadedyr
Forskningscenter Flakkebjerg Forsøgsvej 14200 Slagelse.
Nurrohman I. 2003. Inventarisasi dan perkembangan populasi kutukebul (Hemiptera: Aleyrodidae) pada pertanaman tomat di Bogor dan sekitarnya [skripsi]. Bogor: Institut Pertanian Bogor.
Oliveira CM, Lopes JRS, Dias CTDS, Nault LR. 2004. Influence of latitude and elevation on polymorphism among populations of the corn leafhopper,
Dalbulus maydis (DeLong & Wolcott) (Hemiptera: Cicadellidae), in
Brazil. Environmental Entomology 33: 1192-1199.
Roermund HJW van, Lenteren JC van. 1992. Life-history parameters of the greenhouse whitely, Trialeurodes vaporariorum and the parasitoid
Smith, PE. 2009. Whitefly: identification and biology in New Zealand
greenhouse tomato crops. Factsheet 1. Horticulture New Zealand. Fresh
tomato product group.
Sylvester ES. 1956. Beet yellows virus transmission by the Green peach aphid.
J. Economic Entomology. 49,789-800.
Vaira AM, Accotto GP, Vecchiati M, Bracaloni M. 2002. tomato infectious
chlorosis virus causes leaf yellowing and reddening of tomato in Italy. Phytoparasitica 30: 290-294.
Verhoeven JTJ, Willemen TM, Roenhorst JW, van der Vlugt RAA. 2003. First report of tomato infectious chlorosis virus in Indonesia. Plant Disease 87: 872.
Watson MA, Robers FM. 1939. A comparative study of the transmission of Hyoscyamus virus 3, Potato virus Y and Cucumber virus 1 by the vectors
Myzus persicae (Sulz), M. circumflexus (Buckton), and Macrosiphum gei
(K). Proc. R. Soc. Lond. B 127, 543-577.
Wintermantel WM. 2004. Emergence of greenhouse whitefly (Trialeurodes
vaporariorum) transmitted criniviruses as threats to vegetables and fruit
production in north America. APSnet feature. http// www.apsnet.org/online/eature/whitefly/html. [19 Maret 2010].
Wisler GC, Liu HY, Klaasen VA, Duffus JE, Falk BW. 1996. Tomato infectious
chlorosis virus has a bipartite genome and induces phloem limited
inclusions characteristic of the closteroviruses. Phytopathology 86: 622-626.
Wisler GC, Duffus JE, Liu HY, Li RH. 1998. Ecology and epidemiology of whitefly-transmitted closteroviruses. Plant Disease 82(3): 270-280.
Wisnubroto S, Aminah SL, Nitisapto M. 1986. Asas-Asas Meteorologi
Pertanian. Yogyakarta: Ghalia Indonesia.
Xie M, Chen YH, Wan H. 2006. Responses of two whitefly spesies:
Trialeurodes vaporariorum (Westwood) and Bemisia tabaci (Gennadius)
B-biotype, to low temperature. J of Insect Sci. 8: 1-53.
Zera AJ. 2004. The endocrine regulation of wing polymorphism in insects: state of the art, recent surprises, and future directions. Integr. Comp. Biol. 43: 607-616.
Tabel Lampiran 1 Panjang rostrum dan panjang sayap T. vaporariorum dari Kecamatan Cikajang, Kabupaten Garut
Individu Panjang sayap (µm) Panjang rostrum (µm)
1 1280 272,73 2 1310 318,18 3 1300 290,91 4 1280 327,27 5 1300 309,09 6 1260 290,91 7 1340 281,82 8 1340 309,09 9 1280 268,18 10 1280 218,18 11 1300 336,36 12 1260 281,82 13 1280 309,09 14 1420 354,55 15 1280 290,91 16 1260 281,82 17 1260 290,91 18 1320 309,09 19 1240 318,18 20 1260 309,09 21 1240 272,72 22 1240 272,72 23 1240 272,72 24 1260 281,82 25 1260 290,91 26 1320 309,09 27 1280 272,72 28 1260 272,72 29 1380 300 30 1260 281,82 Rata-rata 1286,33 293,18
Tabel Lampiran 2 Panjang rostrum dan panjang sayap T. vaporariorum dari Kecamatan Pacet, Kabupaten Cianjur
Individu Panjang sayap (µm) Panjang rostrum (µm)
1 1200 290,91 2 1260 272,73 3 1200 281,82 4 1220 272,73 5 1240 263,63 6 1240 281,82 7 1180 254,54 8 1220 254,54 9 1220 263,63 10 1200 254,54 11 1180 254,54 12 1180 263,63 13 1220 290,91 14 1220 290,91 15 1200 281,82 16 1220 281,82 17 1220 281,82 18 1220 281,82 19 1200 281,82 20 1200 281,82 21 1200 300 22 1220 281,82 23 1200 272,73 24 1240 281,82 25 1240 254,54 26 1220 290,91 27 1180 263,63 28 1180 272,73 29 1200 281,82 30 1240 290,91 Rata-rata 1212 275,76
Tabel Lampiran 3 Panjang rostrum dan panjang sayap T. vaporariorum dari Kecamatan Cikole, Kabupaten Sukabumi
Individu Panjang sayap (µm) Panjang rostrum (µm)
1 1180 300 2 1160 254,54 3 1160 245,45 4 1140 254,54 5 1180 272,73 6 1160 272,73 7 1160 263,63 8 1100 227,27 9 1180 272,73 10 1160 227,27 11 1180 254,54 12 1160 236,36 13 1160 245,45 14 1160 254,54 15 1160 263,63 16 1160 254,54 17 1180 245,45 18 1200 245,45 19 1140 254,54 20 1140 245,45 21 1160 245,45 22 1140 254,54 23 1180 254,54 24 1160 254,54 25 1140 245,45 26 1180 281,82 27 1180 254,54 28 1180 272,73 29 1160 272,73 30 1180 263,63 Rata-rata 1162,67 256,36
Tabel Lampiran 4 Panjang rostrum dan panjang sayap T. vaporariorum dari Kecamatan Batu, Kotamadya Batu
Individu Panjang sayap (µm) Panjang rostrum (µm)
1 1120 236,36 2 1100 254,54 3 1100 254,54 4 1080 254,54 5 1140 236,36 6 1120 245,45 7 1100 245,45 8 1140 245,45 9 1100 254,54 10 1140 263,63 11 1100 263,63 12 1160 236,36 13 1160 254,54 14 1160 281,82 15 1160 263,63 16 1140 245,45 17 1100 236,36 18 1120 245,45 19 1100 227,27 20 1080 227,27 21 1080 263,63 22 1140 254,54 23 1100 254,54 24 1080 245,45 25 1140 245,45 26 1100 281,82 27 1140 263,63 28 1120 263,63 29 1160 245,45 30 1100 263,63 Rata-rata 1119,33 251,81
Tabel Lampiran 5 Data suhu bulanan tahun 2010
Bulan Suhu (°C)
Cikajanga Paceta Cikoleb Batua
Januari 19,9 20,2 22,2 22,2 Februari 19,8 20,8 22,1 22,4 Maret 20,4 21,1 22,4 22,6 April 21,0 21,7 23,2 23,5 Mei 21,1 21,9 23,3 23,8 Juni 20,2 21,3 22,4 21,8 Juli 19,6 21,1 21,9 21,7 Agustus 19,9 21,1 22,0 21,7 September 19,9 20,8 22,0 22,1 Oktober 20,2 20,8 22,1 22,3 November 20,2 20,6 22,1 22,8 Desember 19,6 20,8 21,9 22,0 Rata-rata 20,1 21,0 22,3 22,4 a
Sumber: Badan Meteorologi Klimatologi dan Geofisika b Hasil konversi menggunakan rumus Mock (1969)
Tabel Lampiran 6 Data kelembaban bulanan tahun 2010
Bulan Kelembaban (%)
Cikajanga Paceta Cikoleb Batua
Januari 88 87 83 80 Februari 89 86 82 82 Maret 88 87 83 81 April 83 83 79 82 Mei 86 84 80 80 Juni 86 81 80 79 Juli 87 82 79 79 Agustus 87 84 79 78 September 88 85 78 77 Oktober 86 83 79 78 November 89 81 83 80 Desember 87 84 81 77 Rata-rata 87 84 80,5 79,4
a Sumber: Badan Meteorologi Klimatologi dan Geofisika b