TINJAUAN PUSTAKA
Selulosa
Selulosa merupakan polimer karbohidrat terbanyak yang terdapat di alam (Han & Chen 2007). Selulosa merupakan komponen utama penyusun dinding sel tumbuhan bersama-sama dengan hemiselulosa dan pektin. Komposisi selulosa dalam tumbuhan dapat mencapai 40-50% dari massa tumbuhan sehingga selulosa merupakan biopolimer terbarukan yang paling berlimpah di alam (Milala et al. 2005). Classen (1999) menambahkan bahwa diperkirakan 50% dari biomassa tumbuhan berupa selulosa dan jumlahnya sekitar 50 milyar ton. Selulosa merupakan polimer glukosa yang dihubungkan dengan ikatan β-1,4-D-glukosidik (Gambar 1).
Gambar 1 Struktur serat selulosa (Beguin & Aubert 1994).
Polimer glukosa tersusun secara paralel dan berikatan silang membentuk struktur kristalin yang disebut mikrofibril. Panjang mikrofibril ini bervariasi dari 2.000-15.000 unit glukosa, tergantung organismenya. Bentuk mikrofibril selulosa ditentukan oleh kompleks geometri sintase dan lingkungan lokal. Pada tumbuhan, unit mikrofibril mempunyai jumlah sekitar 3-4 unit dan terdiri atas sekitar 36 rantai selulosa dan seringkali dikemas dalam bentuk lebih besar (Doblin et al. 2002).
Mikrofibril pada selulosa memiliki orientasi beragam, tersusun secara pararel, dan setiap molekul glukosa dapat berotasi hingga 1800 (Beguin & Aubert 1994; Brown 1996). Mikrofibril ini pada tempat-tempat tertentu memiliki struktur yang teratur (crystalin) dan pada tempat-tempat tertentu memiliki struktur yang
kurang teratur (amorphous). Struktur amorphous terjadi karena proses kristalisasi yang berlangsung secara tidak sempurna pada mikrofibril yang terbentuk (Gambar 2). Dimensi serat selulosa dan proporsi dari bagian kristalin dan amorf sangat tergantung pada keadaan alaminya (Linder & Teeri 1997). Setiap serat selulosa tersusun oleh kira-kira 3.000 molekul glukosa dan berat molekulnya diperkirakan mencapai 500.000 (Hardjo et al. 1984).
Gambar 2 Struktur selulosa teratur (kristalin) dan kurang teratur (amorphous) (Beguin & Aubert 1994).
Secara alamiah molekul selulosa tersusun dalam fibril yang terdiri atas beberapa molekul glukosa yang dihubungkan dengan ikatan hidrogen yang kuat mengakibatkan dapat tahan terhadap tarikan tinggi. Fibril-fibril ini membentuk struktur kristal yang dibungkus oleh lignin, oleh karena itu sumber selulosa dari tumbuh-tumbuhan sulit sekali dihidrolisis secara langsung oleh katalis asam. Molekul selulosa berbentuk lurus dan tidak pernah bercabang, serta gugus hidroksilnya bebas membentuk ikatan hidrogen dengan gugus hidroksil molekul selulosa lainnya yang terletak sejajar (paralel) dengannya (Beguin & Aubert 1994).
Rumput Laut
Selulosa juga diproduksi oleh tanaman laut yaitu rumput laut (Linder & Teeri 1997). Rumput laut merupakan makroalga laut yang dapat digolongkan ke dalam alga merah, alga hijau, dan alga coklat. Rumput laut tidak memiliki daun, batang, dan akar sejati. Akan tetapi, bagian tubuhnya disebut dengan talus, dapat berupa filamen, lembaran tipis berdaun banyak, persegi dengan kulit keras, dan lumut raksasa. Uji proksimat yang dilakukan pada ampas rumput laut kering
didapatkan presentase masing-masing komponen kadar air sebesar 11.28%, kadar abu 36,05%, kadar lemak 0,42%, kadar protein 1,86%, kadar serat kasar 8,96% dan karbohidrat 41,43% (Harvey 2009).
Jenis rumput laut yang telah banyak dimanfaatkan berasal dari marga Euchema, Gelidium, Gracilaria, Hypnea, dan Sargassum. Selain itu, terdapat jenis lainnya seperti Caulerpa dan Dictosphaeria masih dimanfaatkan dalam skala kecil untuk konsumsi lokal (Atmadja et al. 1996). Beberapa jenis rumput laut memiliki komposisi kandungan selulosa maupun kandungan senyawa kimia lainnya yang berbeda. Berikut ini komposisi kimia dari beberapa jenis rumput laut (Tabel 1).
Tabel 1 Komposisi kimia rumput laut (Kim et al. 2008) Jenis alga Selulosa
(%) Galaktan (%) Karbo- hidrat (%) Protein (%) Lipid (%) Alga merah Gelidium amansii, marocco
Gelidium amansii, joju Glacilaria E. cottonii 16,8 23 19,7 7,1 55,2 56,4 54,4 43,4 72,0 79,4 74,1 50,5 21,1 11,8 11 4,9 6,9 8,8 14,9 44,6 Alga hijau Codium fragile 10,9 47,8 58,7 34,7 6,6 Alga coklat Undaria pinattinda Laminaria japonica 2,4 6,7 38,7 40,0 41,1 46,7 24,2 12,2 34,7 38,1
Rumput laut Glacilaria sp. banyak dimanfaatkan dalam industri pengolahan agar-agar. Limbah industri agar-agar yang dihasilkan mengandung selulosa sebesar 15-25% (Kim et al. 2008). Selain itu, limbah agar-agar Glacilaria sp. merupakan salah satu sumber bakteri yang berpotensi menghasilkan enzim selulase. Pemanfaatan limbah agar-agar dan enzim selulase dari bakteri tersebut memegang peranaan yang sangat penting dalam pengembangan bioenergi.
Enzim Selulase
Enzim selulase atau enzim yang dikenal dengan nama sistematik β-1,4 glukan-4-glukano hidrolase adalah enzim yang dapat menghidrolisis selulosa dengan memutus ikatan glikosidik β-1,4 dalam selulosa, selodektrin, selobiosa, dan turunan selulosa lainnya menjadi gula sederhana atau glukosa. Sistem
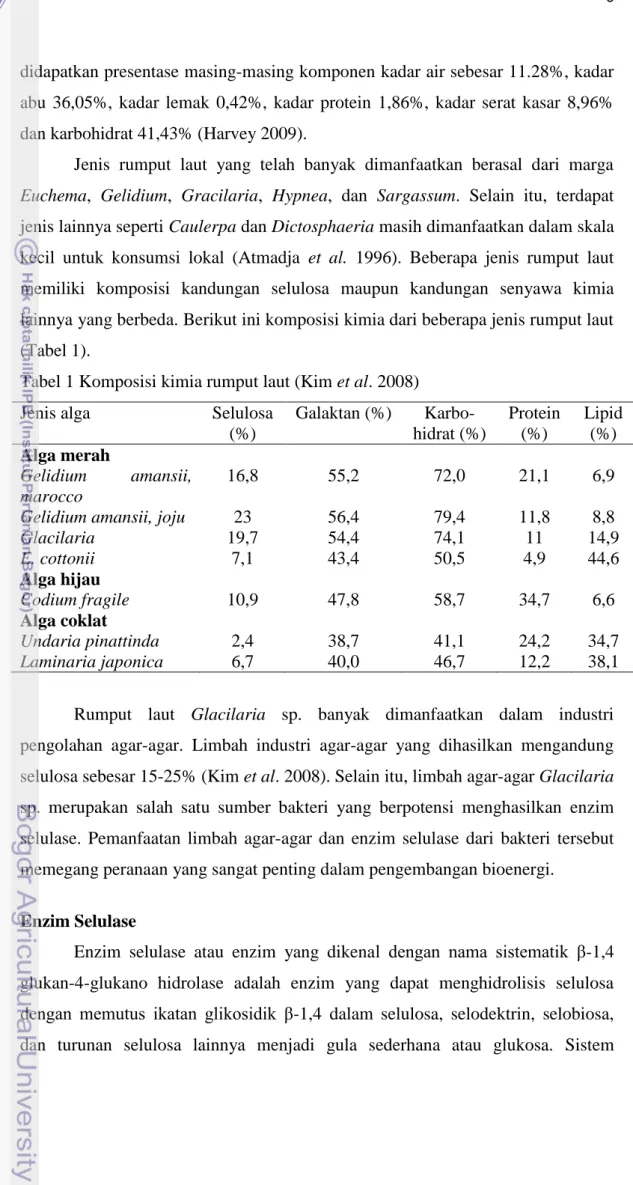
pemecahan selulosa menjadi glukosa terdiri atas tiga jenis enzim selulase yaitu endo-β-1,4-glukanase, ekso-β-1,4-glukanase, dan β-glukosidase. Endo-β-1,4-glukanase menyerang bagian tengah rantai secara random, ekso-β-1,4-Endo-β-1,4-glukanase (selobiohidrolase) memecah unit-unit disakarida (selobiosa) dari ujung rantai, dan β-glukosidase memecah selobiosa menjadi glukosa (Da silva et al. 2005) (Gambar 3).
Gambar 3 Pemecahan selulosa menjadi glukosa oleh enzim selulase.
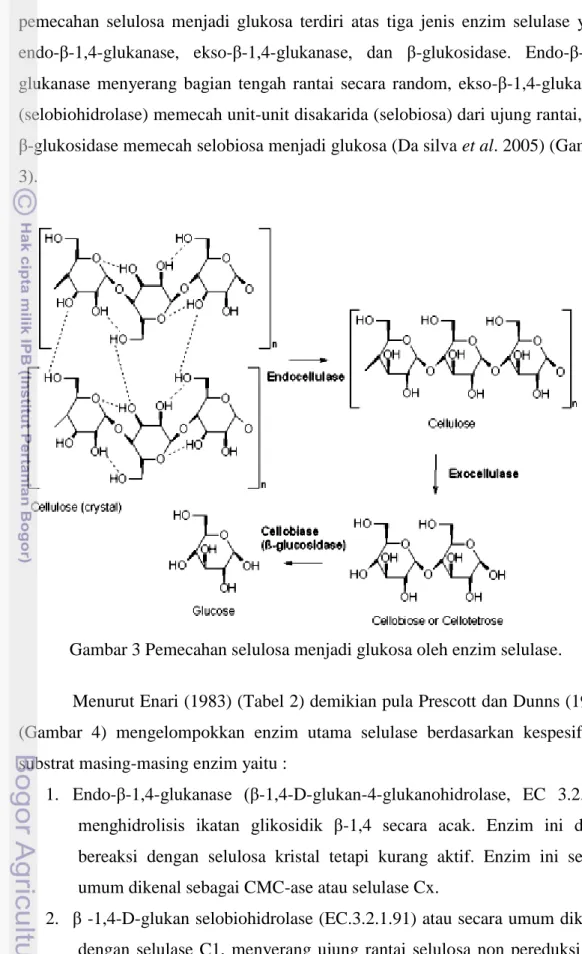
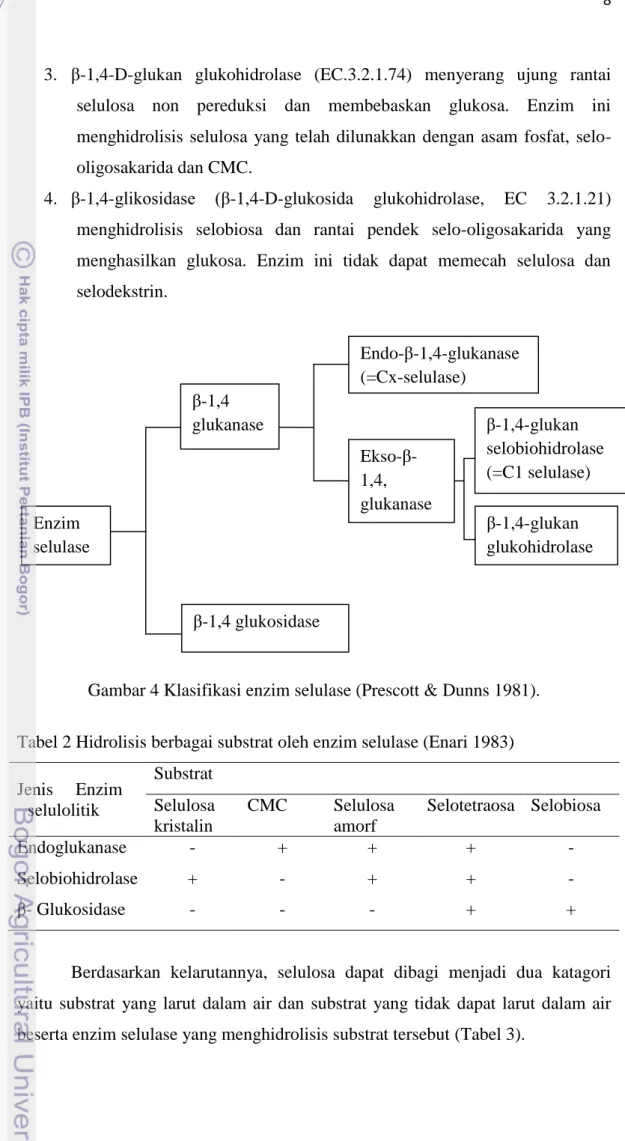
Menurut Enari (1983) (Tabel 2) demikian pula Prescott dan Dunns (1981) (Gambar 4) mengelompokkan enzim utama selulase berdasarkan kespesifikan substrat masing-masing enzim yaitu :
1. Endo-β-1,4-glukanase (β-1,4-D-glukan-4-glukanohidrolase, EC 3.2.1.4) menghidrolisis ikatan glikosidik β-1,4 secara acak. Enzim ini dapat bereaksi dengan selulosa kristal tetapi kurang aktif. Enzim ini secara umum dikenal sebagai CMC-ase atau selulase Cx.
2. β -1,4-D-glukan selobiohidrolase (EC.3.2.1.91) atau secara umum dikenal dengan selulase C1, menyerang ujung rantai selulosa non pereduksi dan membebaskan selobiosa.
3. β-1,4-D-glukan glukohidrolase (EC.3.2.1.74) menyerang ujung rantai selulosa non pereduksi dan membebaskan glukosa. Enzim ini menghidrolisis selulosa yang telah dilunakkan dengan asam fosfat, selo-oligosakarida dan CMC.
4. β-1,4-glikosidase (β-1,4-D-glukosida glukohidrolase, EC 3.2.1.21) menghidrolisis selobiosa dan rantai pendek selo-oligosakarida yang menghasilkan glukosa. Enzim ini tidak dapat memecah selulosa dan selodekstrin.
Gambar 4 Klasifikasi enzim selulase (Prescott & Dunns 1981).
Tabel 2 Hidrolisis berbagai substrat oleh enzim selulase (Enari 1983)
Jenis Enzim selulolitik Substrat Selulosa kristalin CMC Selulosa amorf Selotetraosa Selobiosa Endoglukanase - + + + - Selobiohidrolase + - + + - β- Glukosidase - - - + +
Berdasarkan kelarutannya, selulosa dapat dibagi menjadi dua katagori yaitu substrat yang larut dalam air dan substrat yang tidak dapat larut dalam air beserta enzim selulase yang menghidrolisis substrat tersebut (Tabel 3).
Enzim selulase β-1,4 glukanase β-1,4-glukan glukohidrolase β-1,4-glukan selobiohidrolase (=C1 selulase) Endo-β-1,4-glukanase (=Cx-selulase) Ekso-β-1,4, glukanase β-1,4 glukosidase
Tabel 3 Substrat selulosa berdasarkan kelarutan air dan jenis enzim selulase (Zhang et al. 2006)
Substrat Selulosa Enzim Selulase
Larut dalam air
- Rantai pendek (derajat polimerisasi rendah) Silodekstrin
Radio-labeled selodekstrin - Turunan silodekstrin
β-methyllumberlliferil oligosakarida p-nitrofenol oligosakarida
- Turunan selulosa dengan rantai panjang Carboxymethylecellulose (CMC) Dye CMC
Tidak larut dalam air - Selulosa kristalin
Katun, selulosa mikrokristalin (Avisel), selulosa bakteri
- Selulosa Amorf – PASC - Dyed Selulosa
- Kromogenik dan turunan fluoreforik Trinitrofenil-karboksimetilselulase (TNP-CMC) - Flurant Selulosa - α-selulosa Endo, ekso, BG Endo, ekso, BG Endo, ekso, BG Endo, ekso, BG Endo Endo Total,endo, ekso Total, endo.ekso Total, endo Endo Endo, total Total
Endo ; endoglukanase, Ekso ; eksoglukanase, BG ; glukosidase, Total ; ketiga tipe enzim selulase.
Perbedaan antara masing-masing enzim selulase terletak pada kespesifikan struktur di sekeliling substrat. Perbedaan kespesifikan dari enzim endoglukanase dan selobiohidrolase bersifat tidak mutlak karena kedua enzim tersebut dapat menghidrolisis ikatan β-1,4 glukosida dari selulosa amorf. Penentuan aktivitas enzim selulase akan sulit apabila filtrat yang akan diukur aktivitas enzimnya merupakan campuran dari berbagai enzim selulase. Enzim-enzim ini tidak hanya dapat menghidrolisis substrat yang sama tetapi juga dapat bekerja secara sinergis memecah substrat yang sama, sehingga menyebabkan aktivitas yang diukur dipengaruhi oleh proporsi dari masing-masing enzim yang ada (Enari 1983).
Aktivitas enzim endoglukanase pada umumnya dapat diuji dengan substrat CMC (Carboxymethyl cellulose) sehingga enzim endoglukanase juga disebut dengan istilah CMCase, sedangkan aktivitas enzim selobiohidrolase atau
eksoglukanase seringkali diuji dengan substrat avisel sehingga enzim eksoglukanase disebut dengan aviselase (Zhang et al. 2006).
Tahapan hidrolisis selulosa tergantung kepada struktur selulosa, interaksi antara enzim selulase dengan serat selulosa, mekanisme hidrolisis enzim tersebut di alam dan inhibitor yang terbentuk. Fase adsorbsi dan pembentukan kompleks enzim substrat adalah fase kritis di dalam hidrolisis selulosa. Glukosa dan selobiosa adalah inhibitor enzim dalam menghidrolisis selulosa. Selobiosa menghambat enzim selobiohidrolase dan glukosa menghambat enzim penghidrolisis selobiosa yaitu β-glukosidase pada kompleks enzim selulase. Selobiosa mempunyai potensi lebih kuat menjadi inhibitor dibandingkan dengan glukosa (Coughlan 1985). Laju hidrolisis enzim selulase ditentukan oleh struktur substrat (Mandels 1985). Struktur kristal lebih sulit dihidrolisis dibandingkan dengan struktur amorf maka hidrolisis dilakukan oleh enzim endoselulase atau endoglukanase (Coughlan 1985) (Gambar 5).
Gambar 5 Mekanisme degradasi selulosa (Beguin & Aubert 1994).
Aktivitas enzim selulase dipengaruhi oleh beberapa faktor antara lain derajat keasaman (pH), suhu, dan senyawa penghambat. Aktivitas enzim dipengaruhi oleh pH karena sifat ionik gugus karboksil dan gugus amino mudah dipengaruhi oleh pH sehingga apabila terjadi perubahan pH maka akan menyebabkan denaturasi enzim dan menghilangkan aktivitas enzim. Suhu memiliki peranan yang sangat penting dalam reaksi enzimatik. Ketika suhu
bertambah sampai suhu optimum, kecepatan reaksi enzim naik karena energi kinetik bertambah. Bertambahnya energi kinetik enzim akan mempercepat gerak vibrasi, translasi, dan rotasi baik enzim maupun substrat. Hal ini akan memperbesar peluang enzim dan substrat bereaksi. Ketika suhu lebih tinggi dari suhu optimum, protein enzim berubah konformasi sehingga gugus reaktif terhambat. Perubahan konformasi ini dapat menyebabkan enzim terdenaturasi. Substrat juga dapat berubah konformasinya pada suhu yang tidak sesuai, sehingga substrat tidak dapat masuk ke dalam sisi aktif enzim (Ottaway 1984).
Selain pH dan suhu, faktor lain yang mempengaruhi aktivitas selulase yaitu adanya senyawa penghambat berupa ion logam. Penghambatan tersebut dapat dinetralkan dengan menambahkan sistein sehingga aktivitas enzim dapat berlangsung kembali (Kulp 1975). Beberapa senyawa logam dan senyawa lainnya yang dapat menghambat aktivitas selulase ialah Hg2+, Ag2+, dan Cu2+ (Deng & Tabatai 1994; Oikawa et al. 1994), glukanolakton (Kulp 1975), surfaktan, senyawa pengkelat khususnya Sodium Dodecyl Sulphate (SDS), Ethylene Diamine Tetraacetyc Acid (EDTA) (Oikawa et al. 1994), laktat dalam konsentrasi agak rendah (Chesson 1987), dan etanol serta alkohol lainnya (Ooshima et al. 1985). Senyawa penghambat tersebut dapat menekan seluruh kecepatan hidrolisis dengan menghambat adsorbsi eksoglukanase dan endoglukanase pada selulosa, dan menghambat aksi sinergis eksoglukanase dan endoglukanase yang bekerja pada permukaan selulosa.
Mikroorganisme Penghasil Enzim Selulase
Mikroorganisme didefinisikan sebagai organisme yang berukuran sangat kecil (biasanya kurang dari 1 milimeter) sehingga untuk mengamatinya diperlukan bantuan mikroskop atau alat pembesar. Mikroorganisme dapat berupa sel tunggal atau kelompok sel yang mempunyai kemampuan untuk mengatur proses hidupnya tanpa bergantung sel lainnya. Mikroorganisme terdiri atas bakteri, virus, dan cendawan (fungi) yang masing-masing memiliki perbedaan karakteristik secara morfologi, ekologi, dan fisiologi. Bakteri merupakan sel prokariot dengan rRNA bakteri yang dihubungkan oleh ikatan ester dan membran lipid yang merupakan diasil gliserol dieter (Madigan et al. 2000).
Beberapa contoh genus bakteri yang diketahui mempunyai aktivitas selulolitik ialah Acetobacter, Bacillus, Clostridium, Cellulomonas, Pseudomonas, Cytophaga, Sarcina, dan Vibrio, sedangkan contoh genus cendawan yang mempunyai aktivitas selulolitik ialah Bulgaria, Chaetomium, Helotium, Coriolus, Phanerochaete, Poria, Schizophyllum, Serpula, Aspergillus, Cladosporium,
Fusarium, Geotrichum, Myrothecium, Paecilomyces, Penicillium, dan
Trichoderma (Rao 1994). Beberapa jenis organisme juga dapat menghasilkan enzim selulase seperti rayap (Watanabe & Tokuda 2001), remis (Xu et al. 2000), dan arabidopsis.
Di alam, degradasi selulosa kebanyakan dilakukan oleh mikroorganisme aerobik. Mikroorganisme aerobik menghasilkan enzim selulase nonkompleks yang terdiri atas endoglukanase, eksoglukanase, dan glukosidase yang bekerja secara sinergis untuk menghidrolisis selulosa. Mikroorganisme anaerobik menghasilkan enzim selulase kompleks yang disebut selulosom (Doi et al. 2003; Bayer et al. 2004). Meskipun mikroorganisme anaerobik hanya menyumbang sekitar 5-10% dari biodegradasi total selulosa di alam, namun peranannya sangat penting karena bertanggung jawab terhadap degradasi daerah anoksik pada danau, laut, dan saluran pencernaan hewan pemamah biak maupun rayap, yang tidak dapat dilakukan oleh mikroorganisme aerobik (Zhang et al. 2006).
Pemekatan Enzim
Pada tahap awal pemurnian enzim biasanya dilakukan klarifikasi dan pengendapan protein enzim. Klarifikasi berfungsi memisahkan larutan enzim dari partikel-partikel yang tidak larut, misalnya debris sel dan partikel substrat. Klarifikasi dapat dilakukan dengan penyaringan atau sentrifugasi. Pemekatan protein enzim merupakan tahap awal dari prosedur pemurnian enzim sebelum tahap pemurnian berikutnya atau dapat pula digunakan untuk keperluan analisis enzim. Pemekatan protein enzim berfungsi untuk meningkatkan konsentrasi protein enzim, mereduksi volume larutan enzim, dan memisahkan protein enzim dengan protein pengotor yang lain (Harris 1989).
Pemekatan protein dapat dilakukan dengan dua metode, yaitu analitik dan preparatif (penyiapan). Metode analitik menggunakan pengendapan asam (misalnya asam trikloroasetat), pengendapan organik (misalnya aseton atau
etanol), dan imunopresipitasi yang dapat menyebabkan denaturasi protein. Pemekatan protein dengan metode preparatif tetap mempertahankan aktivitas protein misalnya dengan menggunakan pengendapan garam, pengendapan dengan pelarut organik, pengendapan dengan polimer organik, ultrafiltrasi, liofilisasi, dan dialisis (Harris 1989).
Metode pengendapan protein yang biasa dilakukan dalam pengendapan selulase ialah dengan menggunakan amonium sulfat (Jung et al. 2008) dan ultrafiltrasi (Arifin 2006). Amonium sulfat merupakan garam yang paling sering digunakan untuk mengendapkan protein karena memiliki daya larut tinggi di dalam air, relatif tidak mahal, dan kestabilan protein di dalam larutan amonium sulfat (2M- 3M) tahan bertahun-tahun (Scopes 1987).
Prinsip pengendapan dengan garam berdasarkan pada kelarutan protein yang berinteraksi polar dengan molekul air, interaksi ionik protein dengan garam, dan daya tolak menolak protein yang bermuatan sama. Kelarutan protein (pada pH dan suhu tertentu) meningkat pada kenaikan konsentrasi garam (salting in). Kenaikan kelarutan protein akan meningkatkan kekuatan ion larutan. Pada penambahan garam dengan konsentrasi tertentu menyebabkan kelarutan protein menurun (salting out). Molekul air yang berikatan dengan ion-ion garam semakin banyak yang menyebabkan penarikan selubung air yang mengelilingi permukaan protein. Peristiwa ini mengakibatkan protein saling berinteraksi, beragregasi, dan kemudian mengendap (Harris 1989; Scopes 1987).
Garam berlebih yang terdapat di dalam larutan enzim setelah tahap fraksinasi dapat dihilangkan dengan cara dialisis. Pada tahap dialisis, protein ditempatkan di dalam kantung (membran) semipermeabel yang direndam di dalam larutan bufer tertentu. Molekul yang berukuran kecil akan ke luar melalui membran, dan molekul yang berukuran besar akan tertahan di dalam membran dialisis. Ukuran pori kantung dialisis yang terbuat dari bahan selulosa asetat berdiameter 1-20 nm. Ukuran ini menunjukkan berat molekul minimum yang dapat tertahan di dalam membran. Selain dengan dialisis, penghilangan garam dapat dilakukan dengan filtrasi gel. Metode ini biasanya diterapkan untuk sampel yang sedikit, yaitu tidak melampaui 25-30% volume kolom untuk mendapatkan resolusi yang memadai antara protein dan garam. Matriks filtrasi gel memiliki
pori yang berukuran kecil, misalnya Sephadex G-25 buatan Phamacia. Kekurangan metode ini adalah terjadi pengenceran sampel protein (Harris 1989). Ultrafiltrasi merupakan suatu metode untuk mengkonsentrasikan protein dengan menekan cairan larutan protein enzim supaya tertahan di dalam membran. Ukuran cairan yang akan ditahan (retentat) dan yang dikeluarkan (permeat) sesuai dengan ukuran membran yang digunakan. Prinsip pemisahan dengan ultrafiltrasi adalah pemisahan komponen berdasarkan berat molekul (Bollag & Edelstein 1991). Pemisahan komponen ini terjadi karena adanya membran ultrafiltrasi. Membran ultrafiltrasi berfungsi sebagai penghalang (barrier) tipis yang sangat selektif di antara dua fasa, hanya dapat melewatkan komponen tertentu dan menahan komponen lain dari suatu aliran fluida yang dilewatkan melalui membran (Mulder 1996). Proses membran ultrafiltrasi merupakan upaya pemisahan dengan membran yang menggunakan gaya dorong beda tekanan yang dipengaruhi oleh ukuran dan distribusi pori membran (Malleviale 1996).
Kromatografi Kolom
Kromatografi kolom pada prinsipnya yaitu pengaliran suatu cairan melalui kolom yang mengandung bahan pengisi dan substanta yang ingin dipisahkan menjadi beberapa komponen dengan adanya perbedaan terhadap daya ikat bahan pengisi (Tabel 4).
Tabel 4 Metode kromatografi untuk fraksinasi protein (Ersson et al. 1998) Sifat Protein Jenis Kromatografi Ukuran dan bentuk Filtrasi gel
Muatan neto dan distribusi grup bermuatan
Penukar ion Titik isoelektris Kromatofokusing
Hidrofobisitas Interaksi hidrofobik dan fase balik Pengikatan logam Afinitas ion logam terimobilisasi Kandungan tiol yang terbuka Kovalen
Afinitas biospesifik terhadap ligan, inhibitor, reseptor, antibodi, dsb
Afinitas
Teknik kromatografi kolom banyak digunakan dalam bioteknologi untuk mengamati tingkat kemurnian dan stabilitas protein (Neville 1998). Beberapa peneliti melakukan pemurnian enzim selulase yang dihasilkan oleh bakteri dengan berbagai teknik kromatografi kolom (Tabel 5).
Tabel 5 Teknik kromatografi yang digunakan pada pemurnian selulase Selulase Metode Kromatografi Sumber Endoglukanase dari
Sinorhizobium fredii
Penukar ion, interaksi hidrofobisitas
Po et al. (2004) Endoglukanase dari
Mucor circinelloides
Gel filtrasi Saha (2003) Endoglukanase dari
Bacillus sp
Penukar ion, gel filtrasi Mawadza et al. (2000) Endoglukanase dari
Bacillus sp
Penukar ion Singh et al. (2004) Endoglukanase dari
Pseudomonas fluorescens
Penukar ion, gel filtrasi Bakare et al. (2005) Endoglukanase dari
Bacillus sp
Penukar ion Ji et al. (2005) Endoglukanase dari
Bacillus pumilus
Gel filtrasi, penukar ion Christakopoulus et al. (1999)
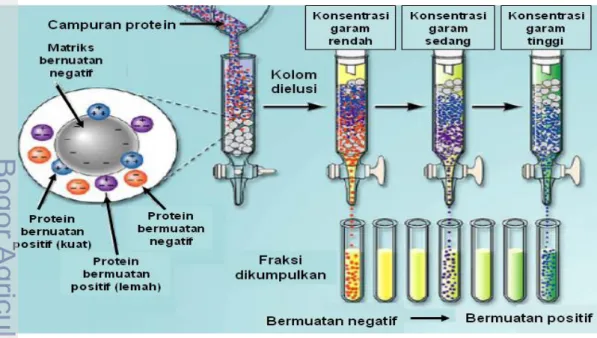
Kromatografi penukar ion memanfaatkan perbedaan afinitas antara molekul bermuatan di dalam larutan dengan senyawa yang tidak reaktif yang bermuatan berlawanan sebagai pengisi kolom (Scopes 1987). Kromatografi penukar ion memisahkan protein berdasarkan muatan bersih protein dan kekuatan relatif dari muatan bersih protein tersebut. Kromatografi penukar ion memerlukan fase diam yang biasanya merupakan polimer terhidratasi yang bersifat tidak larut seperti selulosa, dekstran dan agarosa. Gugus penukar ion diimobilisasikan pada matriks. Beberapa gugus penukar anion yaitu aminoetil (AE-), kuaternari aminoetil (QAE-), dan dietilaminoetil (DEAE-). Gugus penukar kation yaitu sulfopropil (SP-), metil sulfonat dan karboksimetil (CM-). Penukar ion lemah seperti DEAE- (penukar anion lemah) dan CM- (penukar kation lemah) hanya dapat mempertahankan kondisi terionisasi pada rentang pH sempit dan kehilangan muatannya pada pH tertentu. Gugus penukar anion lemah DEAE- terionisasi sempurna di bawah pH 6,0 dan akan kehilangan muatannya pada pH 9,0, sedangkan gugus penukar kation lemah CM- akan kehilangan muatannya di bawah pH 4,5. Penukar ion kuat dapat mempertahankan kondisi terionisasi pada rentang pH yang luas. Gugus penukar ion QAE- (penukar anion kuat) dan SP- (penukar kation kuat) dapat mempertahankan kondisi terionisasi pada rentang pH 1-10 (Coligan et al. 2003).
Kolom untuk kromatografi penukar ion biasanya tidak panjang dan memiliki diameter lebih besar dari pada kolom untuk filtrasi gel. Banyaknya sampel yang dimasukkan umumnya sekitar 10-20% dari kapasitas kolom. Pembilasan dengan gradien konsentrasi NaCl yang linier baik digunakan untuk memisahkan molekul-molekul yang memiliki perbedaan muatan bersih yang tidak terlalu besar sedangkan gradien NaCl bertahap baik digunakan untuk memisahkan molekul-molekul yang memiliki perbedaan muatan bersih yang besar.
Pada dasarnya prinsip kromatografi penukar ion adalah ion bermuatan bebas dipertukarkan dengan ion yang memiliki tipe muatan yang sama. Protein yang bermuatan negatif dapat ditukar dengan ion klorida. Awalnya gugus fungsional matriks yang bermuatan negatif mengikat ion dari bufer (misalnya Na+). Pada saat sampel dimasukkan ke dalam kolom, maka protein yang bermuatan positif akan menggantikan ion Na+ sedangkan protein yang bermuatan negatif atau netral tidak akan terikat. Protein yang tidak terikat dibilas dengan menggunakan bufer (biasanya dengan konsentrasi 10-50 mM). Selanjutnya ikatan protein yang terikat gugus fungsional matriks akan terlepas setelah dibilas dengan bufer yang mengandung NaCl atau KCl secara linier atau bertahap sehingga protein yang memiliki ikatan lemah dengan matriks akan lepas terlebih dahulu dan diikuti oleh protein yang memiliki ikatan lebih kuat (Gambar 6).
Gambar 6 Pemurnian enzim dengan kromatografi pertukar ion
Pemilihan penukar ion tergantung pada muatan protein target. Muatan bersih protein tergantung pada pH yaitu protein akan bermuatan positif dengan menurunkan pH dan bermuatan negatif dengan menaikkan pH. Pada saat menentukan pH untuk kromatografi, kestabilan protein target pada pH yang dipilih perlu dijaga. Apabila protein stabil pada pH di atas titik isoelektriknya (pI) maka digunakan penukar anion (positif), tetapi bila protein stabil pada pH di bawah pI nya maka digunakan penukar kation (negatif). Jika protein stabil pada rentang 1 unit di atas dan di bawah pI maka kedua penukar ion dapat digunakan. Matriks yang mengikat gugus fungsional menentukan sifat aliran, ion yang dapat diikat, kestabilan mekanik dan kimia. Ada 3 kelompok matriks yang biasanya digunakan, yaitu: 1) polistiren, poliakrilik atau polifenol; 2) selulosa; dan 3) dekstran (Sephadex) atau agarosa (Sepharose). Matriks polistiren dan polifenolik lebih sering digunakan untuk memisahkan molekul-molekul kecil seperti asam-asam amino, peptida kecil, nukleotida, nukleotida siklik, asam-asam-asam-asam organik. Matriks selulosa biasanya digunakan untuk memisahkan protein (termasuk enzim), polisakarida dan asam nukleat. Matriks DEAE-selulosa, CM-selulosa dan fosfoselulosa paling sering digunakan. Matriks polidekstran dan agarosa (misalnya DEAE-Sephadex, CM-Sephadex) digunakan untuk memisahkan protein, hormon, tRNA dan polisakarida (Scopes 1987).
Pemilihan penukar ion kuat atau lemah tergantung pada pH molekul target. Molekul yang memerlukan pH sangat rendah atau sangat tinggi untuk dapat berionisasi atau apabila molekul stabil pada pH ekstrem maka penukar ion kuat harus digunakan. Penukar ion lemah akan memberikan hasil pemisahan yang lebih baik untuk protein-protein yang memiliki muatan bersih yang berdekatan. Keuntungan kromatografi penukar ion diantaranya adalah tidak merusak protein yang dimurnikan dan pada umumnya memiliki kapasitas pengikatan yang tinggi. Kelemahannya adalah protein-protein yang memiliki distribusi gugus bermuatan pada permukaannya atau memiliki pI yang sama atau mirip akan sulit dipisahkan dengan cara kromatografi penukar ion. Selain itu larutan enzim hasil kromatografi penukar ion mengandung kadar garam cukup tinggi yang harus dihilangkan untuk proses pemurnian selanjutnya (Scopes 1987).
Elektroforesis
Elektroforesis adalah suatu proses perpindahan partikel-partikel bermuatan atau suatu cara untuk memisahkan fraksi-fraksi suatu campuran berdasarkan atas pergerakan partikel koloid yang bermuatan di bawah pengaruh medan listrik (Suhartono 1989). Elektoforesis dengan menggunakan gel polakrilamida sodium dodesil sulfat (SDS-PAGE) merupakan teknik elektroforesis gel yang menggunakan poliakrilamida untuk memisahkan protein yang bermuatan berdasarkan berat molekulnya. Penentuan berat molekul yang menyusun enzim selulase dianalisis dengan menggunakan metode SDS-PAGE (Sodium dodecyl sulphate-polyacrylamide gel electrophoresis). Pada metode ini digunakan 2 gel yaitu gel penahan (stacking gel) dan gel pemisah (separating gel). Gel akrilamida diperoleh dengan cara polimerisasi akrilamida dengan sejumlah crosslinking agent metilen bis akrilamida dan amonium persulfat (APS) sebagai katalisator. Radikal bebas yang terbentuk dari pelarutan amonium persulfat dalam air akan bereaksi dengan akrilamida membentuk akrilamida aktif yang dapat bereaksi satu dengan yang lain membentuk polimer (Janson & Ryden 1998).
Ada beberapa jenis elektroforesis, yaitu elektroforesis kertas, elektroforesis selulosa asetat/nitrat dan elektroforesis gel. Elektroforesis gel berguna untuk pemisahan protein, sedangkan dua jenis lainnya berguna untuk memisahkan molekul yang lebih kecil. Matriks gel dapat berupa pati, agarosa atau poliakrilamida. Saat ini gel poliakrilamida lebih sering digunakan. Matriks ini disusun oleh akrilamida dan N,N’-metilen-bis-akrilamida yang berpolimerisasi dengan bantuan katalisator amonium persulfat dan N,N,N’,N’tetrametilen diamin (TEMED). Elektroforesis gel dengan SDS digunakan untuk meneliti jumlah dan ukuran rantai protein atau rantai subunit protein. SDS merupakan detergen lemah anionik yang akan memutuskan ikatan di antara subunit penyusun dan membentuk kompleks yang bermuatan negatif sehingga pergerakan protein dalam medan listrik hanya berdasarkan pada ukuran molekul sedangkan β-merkaptoetanol digunakan untuk mereduksi ikatan disulfida pada protein. Protein yang berukuran kecil akan bergerak lebih cepat dibandingkan yang berukuran besar (Copeland 1994).
Elektroforesis protein dapat dilakukan dengan proses denaturasi (SDS-PAGE) dan nondenaturasi (Native-(SDS-PAGE). Mekanisme pada SDS-PAGE dijelaskan bahwa protein akan bereaksi dengan SDS yang merupakan detergen anionik membentuk kompleks yang bermuatan negatif. Protein akan terdenaturasi dan terlarut membentuk kompleks berikatan dengan SDS yang berbentuk elips atau batang yang ukurannya sebanding dengan berat molekul protein. Protein dalam bentuk kompleks yang bermuatan negatif ini akan dapat terpisahkan berdasarkan muatan dan ukurannya secara elektroforesis di dalam matriks gel poliakrilamida (Smith 1984).
Berbeda dengan SDS-PAGE, pada gel pemisah disisipi substrat yang akan dihidrolisis oleh enzim selama masa inkubasi yang disebut sebagai zimogram. Elektroforesis zimogram memisahkan protein terlarut yang tidak mengendap atau beragregasi selama elektroforesis. Pada elektroforesis gel yang terdenaturasi, seperti pada SDS-PAGE, molekul-molekul protein yang telah terpisah dengan elektroforesis dapat kehilangan aktivitas biologi dan biokimianya, tetapi pada elektroforesis zimogram aktivitas tersebut masih bertahan (Dunn 1989). Enzim dipisahkan dalam gel denaturasi (SDS), namun dalam kondisi tidak tereduksi. SDS dilepaskan dengan penambahan larutan renaturasi (misalnya detergen Triton X-100) dan kembali terjadi pelipatan protein. Kemudian gel diwarnai dengan pewarna yang sesuai dengan enzim yang diujikan. Metode zimogram bersifat mudah, sensitif, dan kuantitatif dalam menganalisis aktivitas enzim (Kleiner & Stetler-Stevenson 1994; Leber & Balkwil 1997).
Berat molekul protein dapat ditetapkan dengan menggunakan protein standar yang telah diketahui berat molekulnya dan memperbandingkan nilai Rf (mobilitas relatif) yang diperoleh. Pita pada gel dapat divisualisasi dengan pewarnaan, misalnya menggunakan pewarna coomasie blue atau pewarna perak nitrat (Suhartono 1989).
Identifikasi Mikroorganisme dengan 16S-rRNA
Madigan et al. (2000) menyatakan bahwa pada bakteri atau prokariot memiliki tiga macam ribosom RNA (rRNA) yaitu 23S-rRNA (2900 unit nukleotida), 16S-rRNA (1500 nukleotida) dan 5S-rRNA (sekitar 120 nukleotida). Gen penyandi 16S-rRNA mempunyai daerah sekuen yang konservatif yang dapat
digunakan untuk menduga hubungan kekerabatan secara alami antara spesies yang mempunyai kekerabatan dekat sehingga sangat menguntungkan untuk analisis filogenetik bakteri di tingkat famili, genus, spesies, maupun subspesies. (Chen et al. 2000). Woese (1987) menambahkan bahwa molekul 16S-rRNA paling banyak digunakan sebagai target asam nukleat untuk mendeteksi dan mengidentifikasi bakteri yang belum pernah terdeteksi sebelumnya. Sekuen variabel berevolusi pada laju yang berbeda sehingga memberikan cukup informasi untuk menentukan kedekatan atau jauhnya hubungan filogenetik suatu organisme (Woese 1987).
Madigan et al. (2000) menyatakan sekuen gen penyandi 16S-rRNA digunakan untuk menentukan pohon filogenetik dari keragaman makhluk hidup di bumi. Kekerabatan evolusi antar spesies dalam keseluruhan sistem biologi diperlukan parameter yang memenuhi persyaratan sebagai berikut : 1) terdapat pada semua makhluk hidup, 2) fungsinya identik, 3) dapat dibandingkan secara obyektif, dan 4) parameter tersebut berubah sesuai dengan jarak evolusinya sehingga dapat dijadikan sebagai kronometer evolusi yang handal.
Analisis molekuler dengan sekuen gen penyandi 16S-rRNA pada prinsipnya meliputi ekstraksi DNA total, amplifikasi gen penyandi 16S-rRNA, penentuan sekuen klon yang mengandung gen 16S-rRNA dan analisis perbandingan sekuen yang telah diketahui dalam database (Madigan et al. 2000).